
Cauliflower mosaic virus P6 Dysfunctions Histone Deacetylase HD2C to Promote Virus Infection


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Virus Inoculation
2.2. Yeast Two-Hybrid Assay
2.3. Bimolecular Fluorescence Complementation (BiFC) Assay
2.4. Pull-Down Assay
2.5. Co-Immunoprecipitation (Co-IP) Assay
2.6. Detection of Histone H3 Modifications
2.7. Treatments and Germination Rates
2.8. RNA Extraction and Quantitative Real-Time PCR
2.9. Chromatin Immunoprecipitation (ChIP)
3. Results
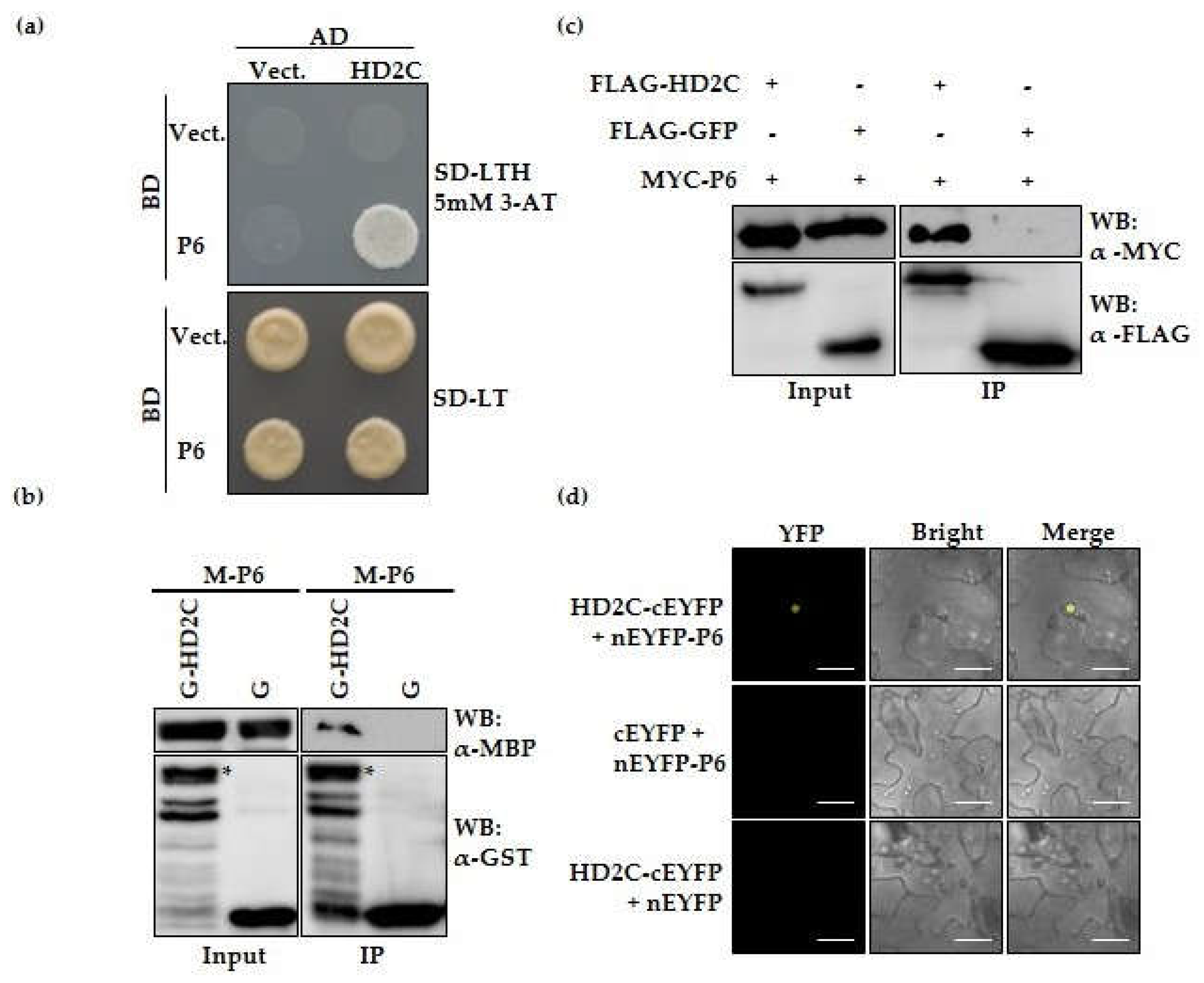
3.1. CaMV P6 Interacts with Histone Acetyltransferase AtHD2C in Nucleoli
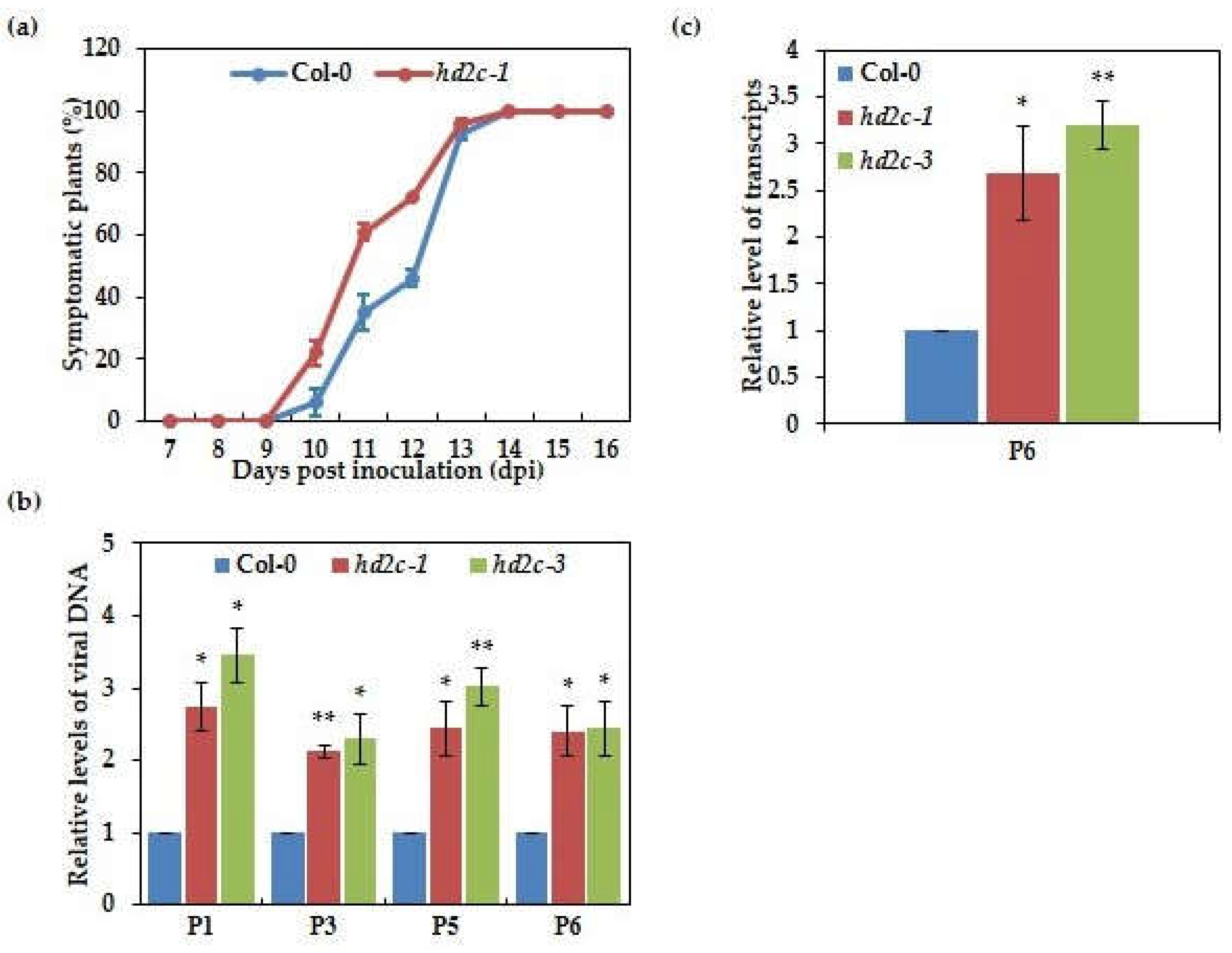
3.2. Mutants of AtHD2C Are More Susceptible to CaMV Infection
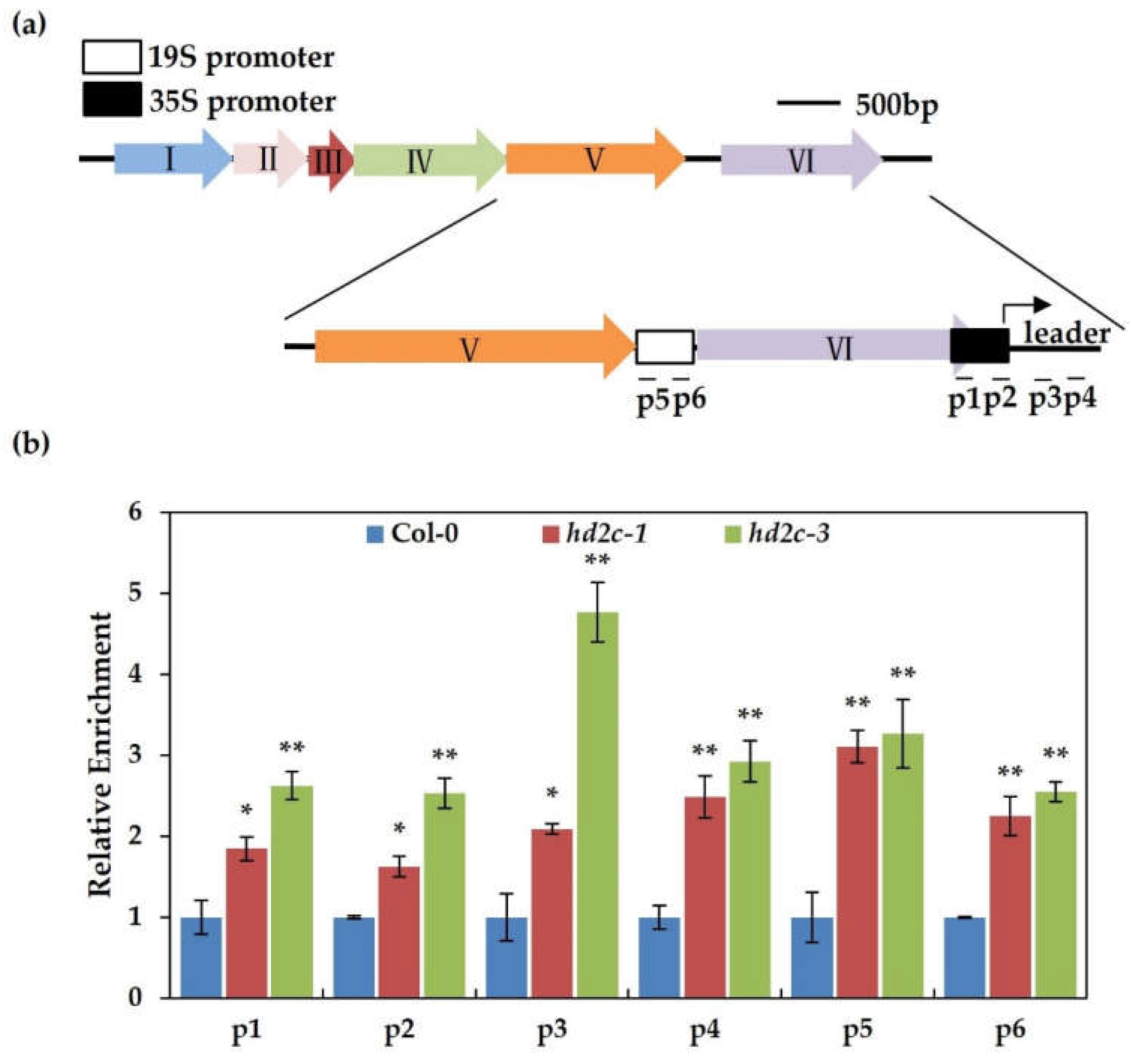
3.3. AtHD2C Functions to Inhibit the Viral Gene Expression on Minichromosome through Histone Deacetylation
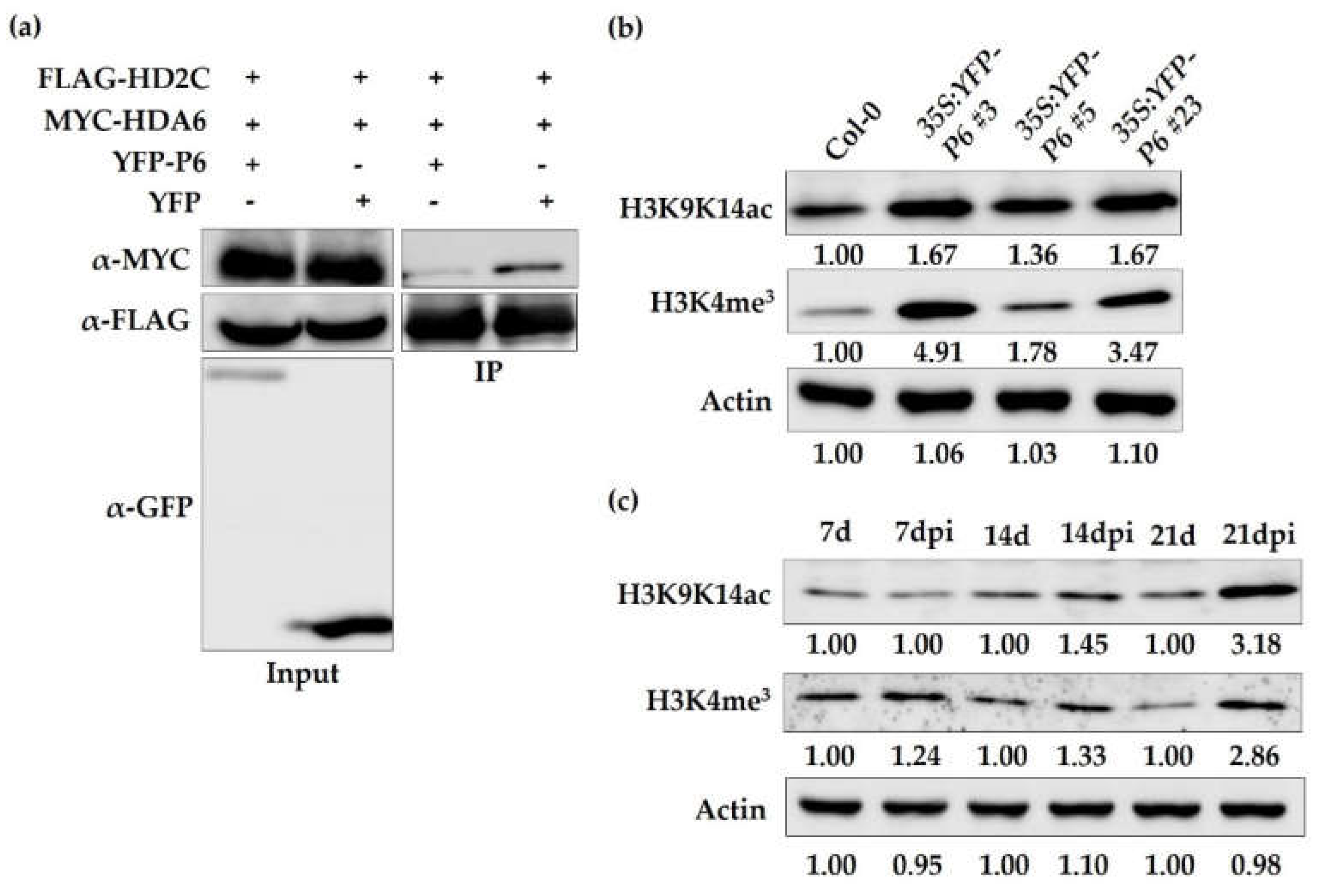
3.4. CaMV Infection and Overexpression of P6 Dysfunction AtHD2C
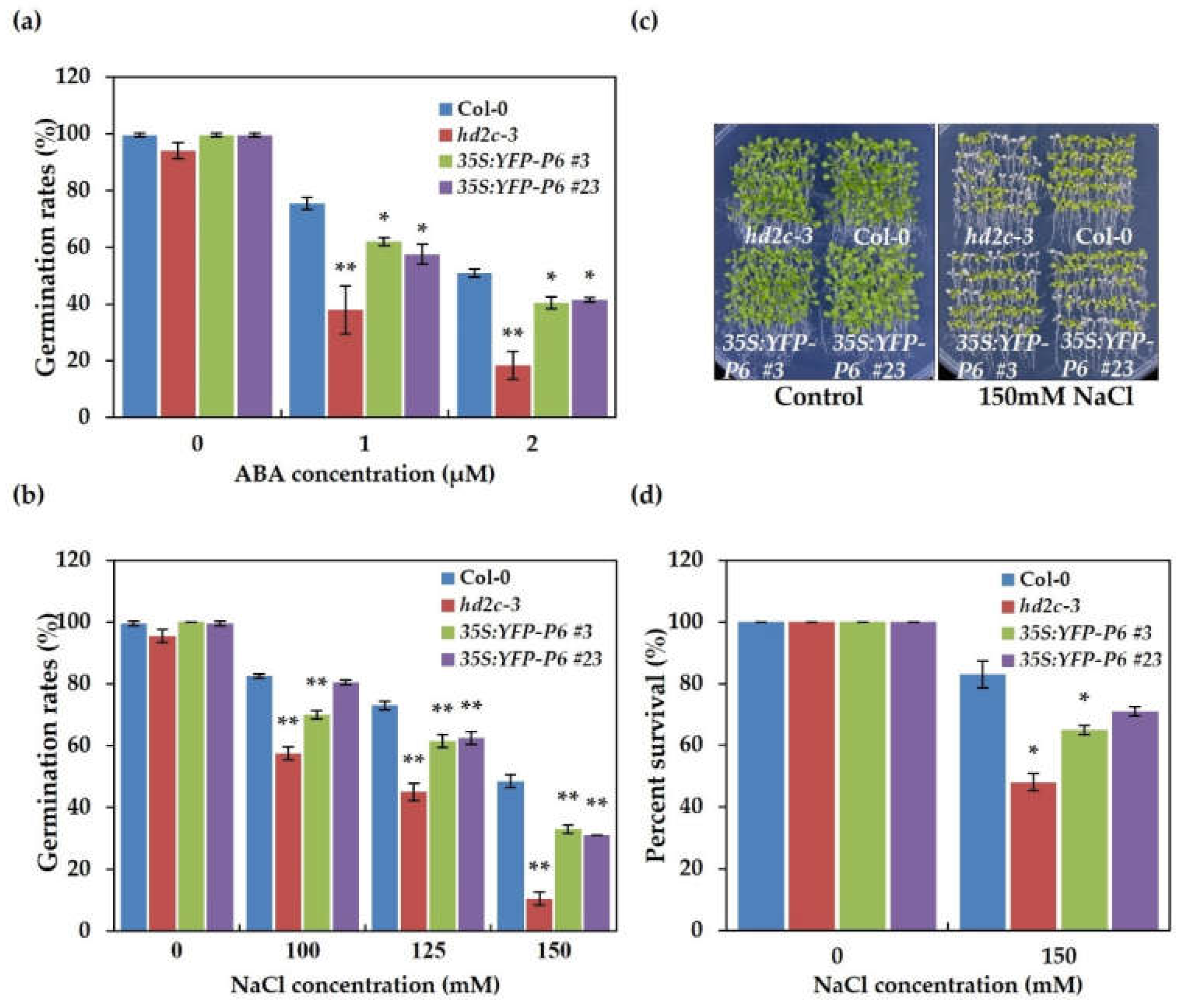
3.5. Overexpression of P6 Increases Sensitivity to ABA and NaCl
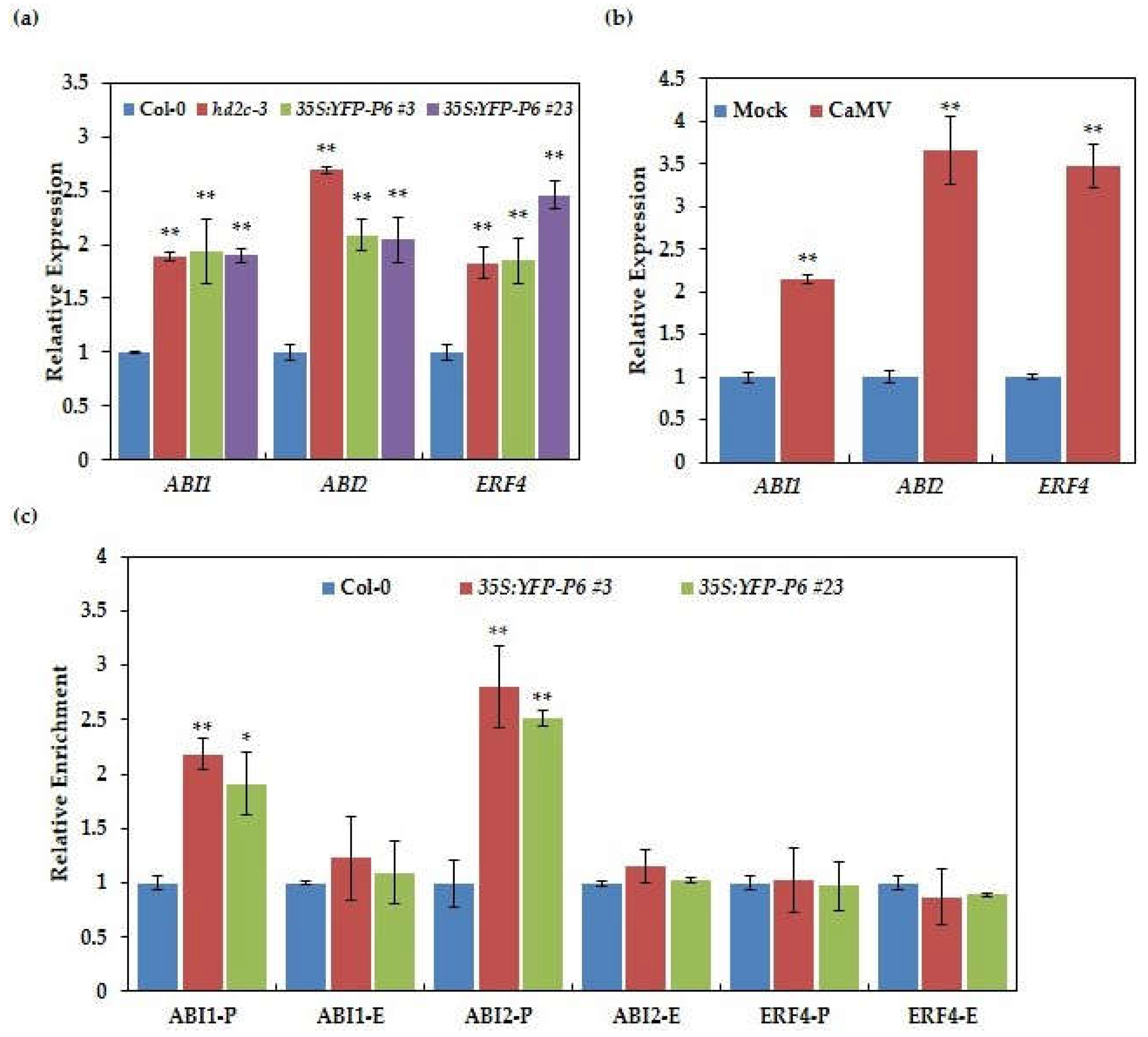
3.6. Histone H3K9K14ac Levels of ABI1 and ABI2 Are Changed in P6 Overexpression Lines
4. Discussion
4.1. Histone Acetylation Participates in Biotic Stress Responses against DNA Viral Infection
4.2. CaMV P6 Interacts with HD2C to Interfere with Histone Deacetylation
4.3. HD2C Functions as a Positive Regulator in Defense Response to DNA Virus Infection
4.4. CaMV Facilitates Host Adaptation under Abiotic Stress by Histone Deacetylation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, L.; Liu, X.; Luo, M.; Yang, S.; Wu, K. Involvement of Histone Modifications in Plant Abiotic Stress Responses. J. Integr. Plant Biol. 2013, 55, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.; MuÈller, A.; Napoli, C.A.; Selinger, D.A.; Pikaard, C.S.; Richards, E.J.; Bender, J.; Mount, D.W.; Jorgensen, R.A. Analysis of histone acetyltransferase and histone deacetylase families of Arabidopsis thaliana suggests functional diversification of chromatin modification among multicellular eukaryotes. Nucleic Acids Res. 2002, 30, 5036–5055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahbazian, M.D.; Grunstein, M. Functions of Site-Specific Histone Acetylation and Deacetylation. Annu. Rev. Biochem. 2007, 76, 75–100. [Google Scholar] [CrossRef] [PubMed]
- Lusser, A.; Brosch, G.; Loidl, A.; Haas, H.; Loidl, P. Identification of maize histone deacetylase HD2 as an acidic nucleolar phosphoprotein. Science 1997, 277, 88–91. [Google Scholar] [CrossRef]
- Wu, K.Q.; Tian, L.N.; Malik, K.; Brown, D.; Miki, B. Functional analysis of HD2 histone deacetylase homologues in Arabidopsis thaliana. Plant J. 2000, 22, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Aravind, L.; Koonin, E.V. Second Family of Histone Deacetylases. Science 1998, 280, 1167. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.M.; Yao, Y.L.; Seto, E. The FK506-binding protein 25 functionally associates with histone deacetylases and with transcription factor YY1. EMBO J. 2001, 20, 4814–4825. [Google Scholar] [CrossRef] [Green Version]
- Dangl, M.; Brosch, G.; Haas, H.; Loidl, P.; Lusser, A. Comparative analysis of HD2 type histone deacetylases in higher plants. Planta 2001, 213, 280–285. [Google Scholar] [CrossRef]
- Zhou, C.; Labbe, H.; Sridha, S.; Wang, L.; Tian, L.; Latoszek-Green, M.; Yang, Z.; Brown, D.; Miki, B.; Wu, K. Expression and function of HD2-type histone deacetylases in Arabidopsis development. Plant J. 2004, 38, 715–724. [Google Scholar] [CrossRef]
- Guo, Z.; Li, Z.; Liu, Y.; An, Z.; Peng, M.; Shen, W.; Dong, A.; Yu, Y. MRG1/2 histone methylation readers and HD2C histone deacetylase associate in repression of the florigen gene FT to set a proper flowering time in response to day-length changes. New Phytol. 2020, 227, 1453–1466. [Google Scholar] [CrossRef]
- Wu, K.Q.; Tian, L.N.; Zhou, C.H.; Brown, D.; Miki, B. Repression of gene expression by Arabidopsis HD2 histone deacetylases. Plant J. 2003, 34, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, L.; Qian, S.; Scalf, M.; Smith, L.M.; Zhong, X. Canonical and Noncanonical Actions of Arabidopsis Histone Deacetylases in Ribosomal RNA Processing. Plant Cell 2018, 30, 134–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colville, A.; Alhattab, R.; Hu, M.; Labbe, H.; Xing, T.; Miki, B. Role of HD2 genes in seed germination and early seedling growth in Arabidopsis. Plant Cell Rep. 2011, 30, 1969–1979. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.S.; Tian, L. HD2-type histone deacetylases: Unique regulators of plant development and stress responses. Plant Cell Rep. 2021, 40, 1603–1615. [Google Scholar] [CrossRef]
- Sridha, S.; Wu, K.Q. Identification of AtHD2C as a novel regulator of abscisic acid responses in Arabidopsis. Plant J. 2006, 46, 124–133. [Google Scholar] [CrossRef]
- Luo, M.; Wang, Y.-Y.; Liu, X.; Yang, S.; Lu, Q.; Cui, Y.; Wu, K. HD2C interacts with HDA6 and is involved in ABA and salt stress response in Arabidopsis. J. Exp. Bot. 2012, 63, 3297–3306. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Wang, Y.Y.; Liu, X.; Yang, S.; Wu, K. HD2 proteins interact with RPD3-type histone deacetylases. Plant Signal. Behav. 2012, 7, 608–610. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.T.; Wu, K. Role of histone deacetylases HDA6 and HDA19 in ABA and abiotic stress response. Plant Signal. Behav. 2010, 5, 1318–1320. [Google Scholar] [CrossRef] [Green Version]
- Buszewicz, D.; Archacki, R.; Palusiński, A.; Kotliński, M.; Fogtman, A.; Iwanicka-Nowicka, R.; Sosnowska, K.; Kuciński, J.; Pupel, P.; Oledzki, J.; et al. HD2C histone deacetylase and a SWI/SNF chromatin remodelling complex interact and both are involved in mediating the heat stress response in Arabidopsis. Plant Cell Environ. 2016, 39, 2108–2122. [Google Scholar] [CrossRef]
- Park, J.; Lim, C.J.; Shen, M.; Park, H.J.; Cha, J.-Y.; Iniesto, E.; Rubio, V.; Mengiste, T.; Zhu, J.-K.; Bressan, R.A.; et al. Epigenetic switch from repressive to permissive chromatin in response to cold stress. Proc. Natl. Acad. Sci. USA 2018, 115, E5400–E5409. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.J.; Park, J.; Shen, M.; Park, H.J.; Cheong, M.S.; Park, K.S.; Baek, D.; Bae, M.J.; Ali, A.; Jan, M.; et al. The Histone-Modifying Complex PWR/HOS15/HD2C Epigenetically Regulates Cold Tolerance. Plant Physiol. 2020, 184, 1097–1111. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.J.; Ali, A.; Park, J.; Shen, M.; Park, K.S.; Baek, D.; Yun, D.-J. HOS15-PWR chromatin remodeling complex positively regulates cold stress in Arabidopsis. Plant Signal. Behav. 2021, 16, 1893978. [Google Scholar] [CrossRef] [PubMed]
- Latrasse, D.; Jégu, T.; Li, H.; de Zelicourt, A.; Raynaud, C.; Legras, S.; Gust, A.; Samajova, O.; Veluchamy, A.; Rayapuram, N.; et al. MAPK-triggered chromatin reprogramming by histone deacetylase in plant innate immunity. Genome Biol. 2017, 18, 131–149. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Bellizzi, M.D.R.; Ning, Y.; Meyers, B.C.; Wang, G.L. HDT701, a Histone H4 Deacetylase, Negatively Regulates Plant Innate Immunity by Modulating Histone H4 Acetylation of Defense-Related Genes in Rice. Plant Cell 2012, 24, 3783–3794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.H.; Zhang, L.; Duan, J.; Miki, B.; Wu, K.Q. HISTONE DEACETYLASE19 is involved in jasmonic acid and ethylene signaling of pathogen response in Arabidopsis. Plant Cell 2005, 17, 1196–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.C.; Lai, Z.; Fan, B.; Chen, Z. Arabidopsis WRKY38 and WRKY62 Transcription Factors Interact with Histone Deacetylase 19 in Basal Defense. Plant Cell 2008, 20, 2357–2371. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Gao, F.; Wu, J.; Dai, J.; Wei, C.; Li, Y. Arabidopsis Putative Deacetylase AtSRT2 Regulates Basal Defense by Suppressing PAD4, EDS5 and SID2 Expression. Plant Cell Physiol. 2010, 51, 1291–1299. [Google Scholar] [CrossRef]
- Wang, B.; Yang, X.; Wang, Y.; Xie, Y.; Zhou, X. Tomato Yellow Leaf Curl Virus V2 Interacts with Host Histone Deacetylase 6 To Suppress Methylation-Mediated Transcriptional Gene Silencing in Plants. J. Virol. 2018, 92, e00036-18. [Google Scholar] [CrossRef] [Green Version]
- Haq, M.I.; Thakuri, B.K.C.; Hobbs, T.; Davenport, M.L.; Kumar, D. Tobacco SABP2-interacting protein SIP428 is a SIR2 type deacetylase. Plant Physiol. Biochem. 2020, 152, 72–80. [Google Scholar] [CrossRef]
- Jin, P.; Gao, S.; He, L.; Xu, M.; Zhang, T.; Zhang, F.; Jiang, Y.; Liu, T.; Yang, J.; Yang, J.; et al. Genome-Wide Identification and Expression Analysis of the Histone Deacetylase Gene Family in Wheat (Triticum aestivum L.). Plants 2021, 10, 19. [Google Scholar] [CrossRef]
- Haas, M.; Bureau, M.; Geldreich, A.; Yot, P.; Keller, M. Cauliflower mosaic virus: Still in the news. Mol. Plant Pathol. 2002, 3, 419–429. [Google Scholar] [CrossRef]
- Olszewski, N.; Hagen, G.; Guilfoyle, T.J. A transcriptionally active, covalently closed minichromosome of cauliflower mosaic virus DNA isolated from infected turnip leaves. Cell 1982, 29, 395–402. [Google Scholar] [CrossRef]
- Omae, N.; Suzuki, M.; Ugaki, M. The genome of theCauliflower mosaic virus, a plant pararetrovirus, is highly methylated in the nucleus. FEBS Lett. 2020, 594, 1974–1988. [Google Scholar] [CrossRef]
- Raja, P.; Sanville, B.C.; Buchmann, R.C.; Bisaro, D.M. Viral genome methylation as an epigenetic defense against geminiviruses. J. Virol. 2008, 82, 8997–9007. [Google Scholar] [CrossRef] [Green Version]
- Love, A.J.; Geri, C.; Laird, J.; Carr, C.; Yun, B.-W.; Loake, G.J.; Tada, Y.; Sadanandom, A.; Milner, J.J. Cauliflower mosaic virus protein P6 inhibits signaling responses to salicylic acid and regulates innate immunity. PLoS ONE 2012, 7, e47535. [Google Scholar] [CrossRef] [Green Version]
- Love, A.J.; Laval, V.; Geri, C.; Laird, J.; Tomos, D.; Hooks, M.A.; Milner, J.J. Components of Arabidopsis defense- and ethylene-signaling pathways regulate susceptibility to Cauliflower mosaic virus by restricting long-distance movement. Mol. Plant-Microbe Interact. 2007, 20, 659–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geri, C.; Love, A.J.; Cecchini, E.; Barrett, S.J.; Laird, J.; Covey, S.N.; Milner, J.J. Arabidopsis mutants that suppress the phenotype induced by transgene-mediated expression of cauliflower mosaic virus (CaMV) gene VI are less susceptible to CaMV-infection and show reduced ethylene sensitivity. Plant Mol. Biol. 2004, 56, 111–124. [Google Scholar] [CrossRef]
- Tsuge, S.; Kobayashi, K.; Nakayashiki, H.; Okuno, T.; Furusawa, I. Replication of Cauliflower Mosaic Virus ORF I Mutants in Turnip Protoplasts. Jpn. J. Phytopathol. 1994, 60, 27–35. [Google Scholar] [CrossRef]
- Adhab, M.; Finke, D.; Schoelz, J. Turnip aphids (Lipaphis erysimi) discriminate host plants based on the strain of Cauliflower mosaic virus infection. Emir. J. Food Agric. 2019, 31, 69–75. [Google Scholar] [CrossRef]
- Sparkes, I.A.; Runions, J.; Kearns, A.; Hawes, C. Rapid, transient expression of fluorescent fusion proteins in tobacco plants and generation of stably transformed plants. Nat. Protoc. 2006, 1, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Iwasaki, M.; Machida, C.; Machida, Y.; Zhou, X.; Chua, N.H. betaC1, the pathogenicity factor of TYLCCNV, interacts with AS1 to alter leaf development and suppress selective jasmonic acid responses. Genes Dev. 2008, 22, 2564–2577. [Google Scholar] [CrossRef] [Green Version]
- Gendrel, A.V.; Lippman, Z.; Martienssen, R.; Colot, V. Profiling histone modification patterns in plants using genomic tiling microarrays. Nat. Methods 2005, 2, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Abouzid, A.M.; Frischmuth, T.; Jeske, H. A putative replicative form of the abutilon mosaic virus (gemini group) in a chromatin-like structure. Mol. Gen. Genet. MGG 1988, 212, 252–258. [Google Scholar] [CrossRef]
- Ceniceros-Ojeda, E.A.; Rodríguez-Negrete, E.A.; Rivera-Bustamante, R.F. Two Populations of Viral Minichromosomes Are Present in a Geminivirus-Infected Plant Showing Symptom Remission (Recovery). J. Virol. 2016, 90, 3828–3838. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Li, L.; Han, X.; Liu, T.; Jin, P.; Cai, L.; Xu, M.; Zhang, T.; Zhang, F.; Chen, J.; et al. Genome-wide identification of the histone acetyltransferase gene family in Triticum aestivum. BMC Genom. 2021, 22. [Google Scholar] [CrossRef]
- Chen, L.T.; Luo, M.; Wang, Y.Y.; Wu, K. Involvement of Arabidopsis histone deacetylase HDA6 in ABA and salt stress response. J. Exp. Bot. 2010, 61, 3345–3353. [Google Scholar] [CrossRef] [Green Version]
- Zhi, P.; Kong, L.; Liu, J.; Zhang, X.; Wang, X.; Li, H.; Sun, M.; Li, Y.; Chang, C. Histone Deacetylase TaHDT701 Functions in TaHDA6-TaHOS15 Complex to Regulate Wheat Defense Responses to Blumeria graminis f.sp. tritici. Int. J. Mol. Sci. 2020, 21, 2640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; He, J.; Velanis, C.N.; Zhu, Y.; He, Y.; Tang, K.; Zhu, M.; Graser, L.; de Leau, E.; Wang, X.; et al. A domesticated Harbinger transposase forms a complex with HDA6 and promotes histone H3 deacetylation at genes but not TEs in Arabidopsis. J. Integr. Plant Biol. 2021. [Google Scholar] [CrossRef]
- Zijlstra, C.; Schärer-Hernández, N.; Gal, S.; Hohn, T. Arabidopsis thaliana expressing the cauliflower mosaic virus ORF VI transgene has a late flowering phenotype. Virus Genes 1996, 13, 5–17. [Google Scholar] [CrossRef]
- Zarreen, F.; Chakraborty, S. Epigenetic regulation of geminivirus pathogenesis: A case of relentless recalibration of defence responses in plants. J. Exp. Bot. 2020, 71, 6890–6906. [Google Scholar] [CrossRef]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. (vol 180, pg 911, 2008). New Phytol. 2009, 184, 275. [Google Scholar] [CrossRef]
- Westwood, J.H.; McCann, L.; Naish, M.; Dixon, H.; Murphy, A.; Stancombe, M.A.; Bennett, M.H.; Powell, G.; Webb, A.; Carr, J. A viral RNA silencing suppressor interferes with abscisic acid-mediated signalling and induces drought tolerance in Arabidopsis thaliana. Mol. Plant Pathol. 2013, 14, 158–170. [Google Scholar] [CrossRef]
- Aguilar, E.; Cutrona, C.; Del Toro, F.J.; Vallarino, J.G.; Osorio, S.; Bueno, M.L.P.; Barón, M.; Chung, B.-N.; Canto, T.; Tenllado, F. Virulence determines beneficial trade-offs in the response of virus-infected plants to drought via induction of salicylic acid. Plant Cell Environ. 2017, 40, 2909–2930. [Google Scholar] [CrossRef]
- Anfoka, G.; Moshe, A.; Fridman, L.; Amrani, L.; Rotem, O.; Kolot, M.; Zeidan, M.; Czosnek, H.; Gorovits, R. Tomato yellow leaf curl virus infection mitigates the heat stress response of plants grown at high temperatures. Sci. Rep. 2016, 6, 19715. [Google Scholar] [CrossRef] [Green Version]
- Gorovits, R.; Sobol, I.; Altaleb, M.; Czosnek, H.; Anfoka, G. Taking advantage of a pathogen: Understanding how a virus alleviates plant stress response. Phytopathol. Res. 2019, 1. [Google Scholar] [CrossRef]
- Yung, W.S.; Li, M.W.; Sze, C.C.; Wang, Q.; Lam, H.M. Histone modifications and chromatin remodelling in plants in response to salt stress. Physiol. Plant 2021. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Lyu, S.; Liu, Y.; Luo, M.; Shi, S.; Deng, S. Cauliflower mosaic virus P6 Dysfunctions Histone Deacetylase HD2C to Promote Virus Infection. Cells 2021, 10, 2278. https://doi.org/10.3390/cells10092278
Li S, Lyu S, Liu Y, Luo M, Shi S, Deng S. Cauliflower mosaic virus P6 Dysfunctions Histone Deacetylase HD2C to Promote Virus Infection. Cells. 2021; 10(9):2278. https://doi.org/10.3390/cells10092278
Chicago/Turabian StyleLi, Shun, Shanwu Lyu, Yujuan Liu, Ming Luo, Suhua Shi, and Shulin Deng. 2021. "Cauliflower mosaic virus P6 Dysfunctions Histone Deacetylase HD2C to Promote Virus Infection" Cells 10, no. 9: 2278. https://doi.org/10.3390/cells10092278
APA StyleLi, S., Lyu, S., Liu, Y., Luo, M., Shi, S., & Deng, S. (2021). Cauliflower mosaic virus P6 Dysfunctions Histone Deacetylase HD2C to Promote Virus Infection. Cells, 10(9), 2278. https://doi.org/10.3390/cells10092278