
Differentially Regulated miRNAs and Their Related Molecular Pathways in Lichen Sclerosus


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Samples
2.2. Cell Lines
2.3. RNA Extraction, RNA Library Preparation and RNASeq Assays
2.4. Data Preprocessing
2.5. Identification of Differentially Expressed miRNAs
2.6. Pathway Analysis
2.7. Quantitative Real-Time Reverse Transcription-PCR (qRT-PCR)
2.8. Target Gene Prediction
2.9. miRNA Precursors and Plasmid Transfection
2.10. Matrigel Invasion Assays
2.11. Statistical Analysis
3. Results
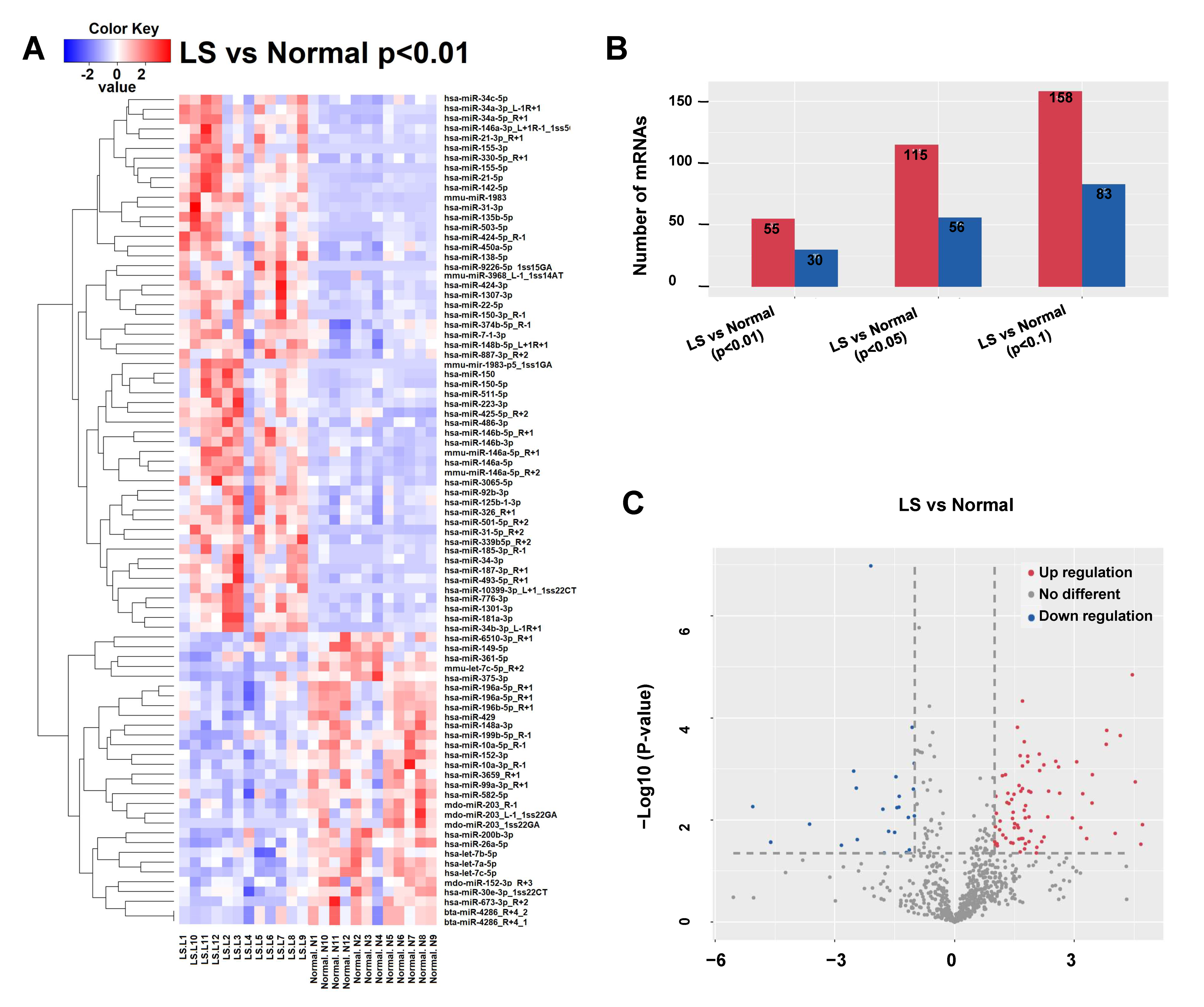
3.1. miRNA Profiles in LS and Matched Adjacent Tissue
3.2. Verifying the Dysregulation of miRNAs in Peripheral Blood
3.3. Gene Network and Pathway Analysis
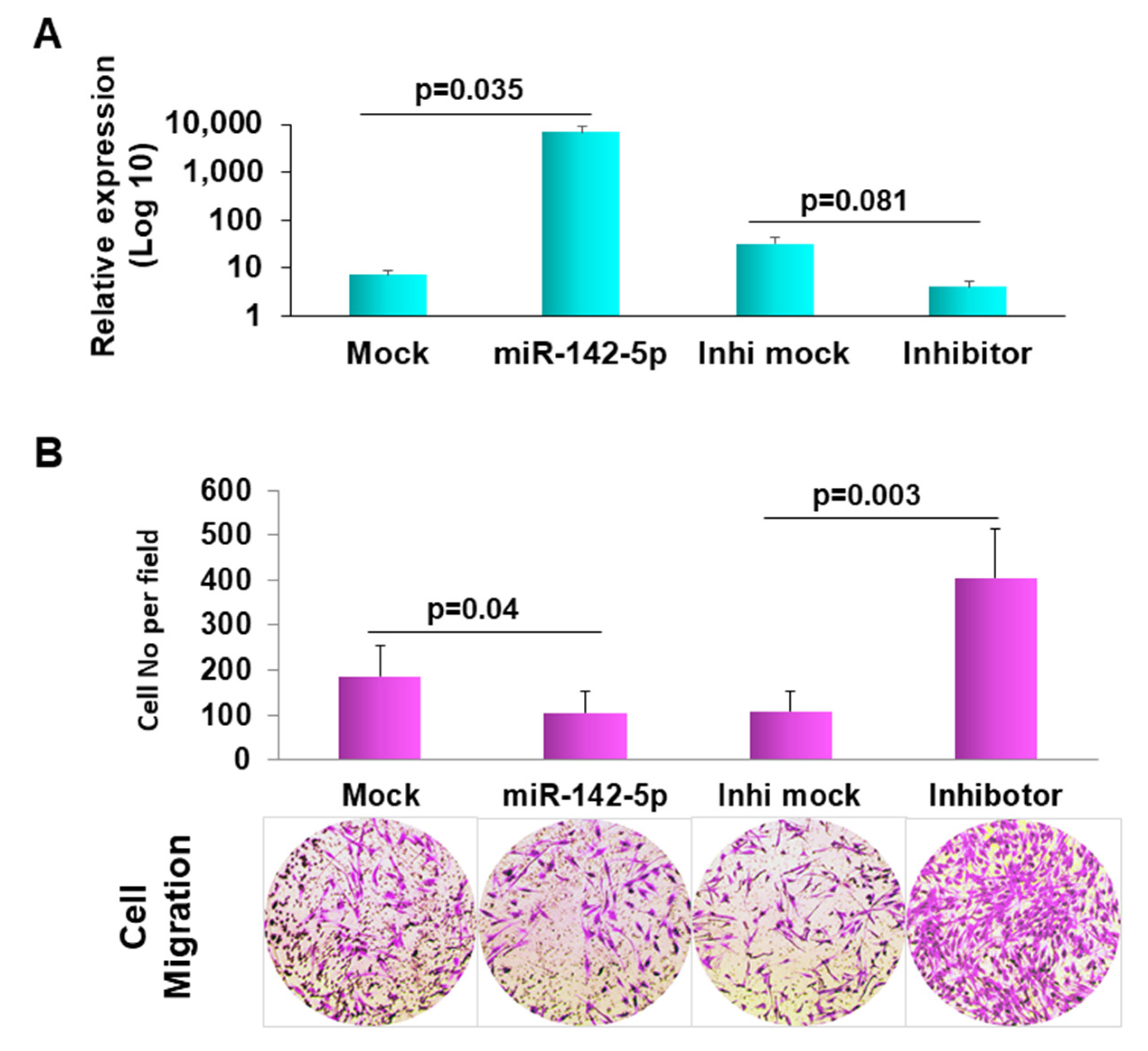
3.4. Forced Expression of miR-142-5p in Human Dermal Fibroblast PCS-201-010 Resulted in Decreased Cell Migration
3.5. Target Gene Prediction and Verification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dongre, H.; Rana, N.; Fromreide, S.; Rajthala, S.; Engelsen, I.B.; Paradis, J.; Gutkind, J.S.; Vintermyr, O.K.; Johannessen, A.C.; Bjørge, L.; et al. Establishment of a novel cancer cell line derived from vulvar carcinoma associated with lichen sclerosus exhibiting a fibroblast-dependent tumorigenic potential. Exp. Cell Res. 2019, 386, 111684. [Google Scholar] [CrossRef] [PubMed]
- Kirtschig, G.; Becker, K.L.; Gunthert, A.R.; Jasaitiene, D.; Cooper, S.; Chi, C.-C.; Kreuter, A.; Rall, K.; Aberer, W.; Riechardt, S.; et al. Evidence-based (S3) guideline on (anogenital) lichen sclerosus. J. Eur. Acad. Dermatol. Venereol. 2015, 29, e1–e43. [Google Scholar] [CrossRef]
- Kreuter, A.; Kryvosheyeva, Y.; Terras, S.; Moritz, R.; Möllenhoff, K.; Altmeyer, P.; Scola, N.; Gambichler, T. Association of autoimmune diseases with lichen sclerosus in 532 male and female patients. Acta Derm. Venereol. 2013, 93, 238–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheinis, M.; Selk, A. Development of the adult vulvar lichen sclerosus severity scale—A delphi consensus exercise for item generation. J. Low. Genit. Tract Dis. 2018, 22, 66–73. [Google Scholar] [CrossRef]
- Haefner, H.K.; Aldrich, N.Z.; Dalton, V.K.; Gagné, H.M.; Marcus, S.B.; Patel, D.A.; Berger, M. The impact of vulvar lichen sclerosus on sexual dysfunction. J. Women’s Health 2014, 23, 765–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, E.G.; MacLaren, N.K. Genetic aspects of vulvar lichen sclerosus. Am. J. Obstet. Gynecol. 1984, 150, 161–166. [Google Scholar] [CrossRef]
- Lambert, J. Pruritus in female patients. BioMed Res. Int. 2014, 2014, 1–6. [Google Scholar] [CrossRef]
- Niamh, L.; Naveen, S.; Hazel, B. Diagnosis of vulval inflammatory dermatoses: A pathological study with clinical correlation. Int. J. Gynecol. Pathol. 2009, 28, 554–558. [Google Scholar] [CrossRef] [PubMed]
- McPherson, T.; Cooper, S. Vulval lichen sclerosus and lichen planus. Dermatol. Ther. 2010, 23, 523–532. [Google Scholar] [CrossRef]
- Fistarol, S.K.; Itin, P.H. Diagnosis and treatment of lichen sclerosus. Am. J. Clin. Dermatol. 2013, 14, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Krapf, J.M.; Mitchell, L.; Holton, M.A.; Goldstein, A.T. Vulvar lichen sclerosus: Current perspectives. Int. J. Women’s Health 2020, 12, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, D.A.; Tan, X.; Macri, C.J.; Goldstein, A.T.; Fu, S.W. Lichen sclerosus: An autoimmunopathogenic and genomic enigma with emerging genetic and immune targets. Int. J. Biol. Sci. 2019, 15, 1429–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marren, P.; Yell, J.; Charnock, F.; Bunce, M.; Welsh, K.; Wojnarowska, F.; Jell, J. The association between lichen sclerosus and antigens of the HLA system. Br. J. Dermatol. 2006, 132, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Burrows, L.J.; Creasey, A.; Goldstein, A. The Treatment of vulvar lichen sclerosus and female sexual dysfunction. J. Sex. Med. 2011, 8, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.; Burrows, L.J. Surgical treatment of clitoral phimosis caused by lichen sclerosus. Am. J. Obstet. Gynecol. 2007, 196, 126.e1–126.e4. [Google Scholar] [CrossRef] [PubMed]
- Bracken, C.P.; Scott, H.S.; Goodall, G.J. A network-biology perspective of microRNA function and dysfunction in cancer. Nat. Rev. Genet. 2016, 17, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Rieger, J.K.; Klein, K.; Winter, S.; Zanger, U.M. Expression variability of absorption, distribution, metabolism, excretion–Related microRNAs in human liver: Influence of nongenetic factors and association with gene expression. Drug Metab. Dispos. 2013, 41, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef]
- Singh, R.P.; Massachi, I.; Manickavel, S.; Singh, S.; Rao, N.P.; Hasan, S.; Mc Curdy, D.K.; Sharma, S.; Wong, D.; Hahn, B.H.; et al. The role of miRNA in inflammation and autoimmunity. Autoimmun. Rev. 2013, 12, 1160–1165. [Google Scholar] [CrossRef] [PubMed]
- Talebi, F.; Ghorbani, S.; Chan, W.F.; Boghozian, R.; Masoumi, F.; Ghasemi, S.; Vojgani, M.; Power, C.; Noorbakhsh, F. MicroRNA-142 regulates inflammation and T cell differentiation in an animal model of multiple sclerosis. J. Neuroinflamm. 2017, 14, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Meng, Z.; Zhang, H.; Li, L.; Wang, K. Clinical significance of miR-142-3p in oral lichen planus and its regulatory role in keratinocyte proliferation. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2021. [Google Scholar] [CrossRef]
- Nymark, P.; Guled, M.; Borze, I.; Faisal, A.; Lahti, L.; Salmenkivi, K.; Kettunen, E.; Anttila, S.; Knuutila, S. Integrative analysis of microRNA, mRNA and aCGH data reveals asbestos- and histology-related changes in lung cancer. Genes Chromosom. Cancer 2011, 50, 585–597. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Fu, Y.; Chen, L.; Lee, W.; Lai, Y.; Rezaei, K.; Tabbara, S.; Latham, P.; Teal, C.B.; Man, Y.G.; et al. MiR-671-5p inhibits epithelial-to-mesenchymal transition by downregulating FOXM1 expression in breast cancer. Oncotarget 2016, 7, 293–307. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. MiRBase: MicroRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Saini, H.K.; van Dongen, S.; Enright, A.J. MiRBase: Tools for microRNA genomics. Nucleic Acids Res. 2008, 36, D154–D158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. MiRBase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. MiRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Peng, J.; Fu, Y.; An, S.; Rezaei, K.; Tabbara, S.; Teal, C.B.; Man, Y.G.; Brem, R.F.; Fu, S.W. MiR-638 mediated regulation of BRCA1 affects DNA repair and sensitivity to UV and cisplatin in triple-negative breast cancer. Breast Cancer Res. 2014, 16, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Ren, S.; Fu, M.Z.; Ren, S.; Yang, C.; Wu, X.; Chen, T.; Latham, P.S.; Meltzer, S.J.; Fu, S.W. microRNA-196b promotes esophageal squamous cell carcinogenesis and chemoradioresistance by inhibiting EPHA7, thereby restoring EPHA2 activity. Am. J. Cancer Res. 2021, 11, 3594–3610. [Google Scholar]
- Jadideslam, G.; Ansarin, K.; Sakhinia, E.; Babaloo, Z.; Abhari, A.; Alipour, S.; Farhadi, J.; Shirvani, S.S.; Ghojazadeh, M.; Khabbazi, A. Expression levels of miR-21, miR-146b and miR-326 as potential biomarkers in Behcet’s disease. Biomark. Med. 2019, 13, 1339–1348. [Google Scholar] [CrossRef]
- Rożalski, M.; Rudnicka, L.; Samochocki, Z. MiRNA in atopic dermatitis. Adv. Dermatol. Allergol. 2016, 33, 157–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Specjalski, K.; Jassem, E. MicroRNAs: Potential biomarkers and targets of therapy in allergic diseases? Arch. Immunol. Ther. Exp. 2019, 67, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Zou, L.; Lu, F.; Ye, L.; Su, B.; Yang, K.; Lin, M.; Fu, J.; Li, Y. MiR-142-5p promotes renal cell tumorigenesis by targeting TFAP2B. Oncol. Lett. 2020, 20, 324. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Z.; Fang, X.; Yang, H. MiR-142-5p Suppresses tumorigenesis by targeting PIK3CA in non-small cell lung cancer. Cell. Physiol. Biochem. 2017, 43, 2505–2515. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.; Zhang, Y.; Yan, R.; Huang, L.; Mellor, A.L.; Yang, Y.; Chen, X.; Wei, W.; Wu, X.; Yu, L.; et al. Exosome-derived miR-142-5p remodels lymphatic vessels and induces IDO to promote immune privilege in the tumour microenvironment. Cell Death Differ. 2021, 28, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Distler, J.H.W.; Akhmetshina, A.; Schett, G.; Distler, O. Monocyte chemoattractant proteins in the pathogenesis of systemic sclerosis. Rheumatology 2008, 48, 98–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X. MiRDB: An online database for prediction of functional microRNA targets. Nucleic Acids Res. 2020, 48, D127–D131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paraskevopoulou, M.D.; Georgakilas, G.; Kostoulas, N.; Vlachos, I.; Vergoulis, T.; Reczko, M.; Filippidis, C.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-microT web server v5.0: Service integration into miRNA functional analysis workflows. Nucleic Acids Res. 2013, 41, W169–W173. [Google Scholar] [CrossRef] [Green Version]
- Reczko, M.; Maragkakis, M.; Alexiou, P.; Grosse, I.; Hatzigeorgiou, A.G. Functional microRNA targets in protein coding sequences. Bioinformatics 2012, 28, 771–776. [Google Scholar] [CrossRef]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seddiki, N.; Brezar, V.; Ruffin, N.; Lévy, Y.; Swaminathan, S. Role of miR-155 in the regulation of lymphocyte immune function and disease. Immunology 2014, 142, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, G.; Biswas, R. MicroRNA-155: A master regulator of inflammation. J. Interf. Cytokine Res. 2019, 39, 321–330. [Google Scholar] [CrossRef]
- Testa, U.; Pelosi, E.; Castelli, G.; Labbaye, C. MiR-146 and miR-155: Two key modulators of immune response and tumor development. Noncoding RNA 2017, 3, 22. [Google Scholar] [CrossRef]
- Vigorito, E.; Kohlhaas, S.; Lu, D.; Leyland, R. MiR-155: An ancient regulator of the immune system. Immunol. Rev. 2013, 253, 146–157. [Google Scholar] [CrossRef]
- Wang, L.; Wu, W.; Chen, J.; Li, Y.; Xu, M.; Cai, Y. MicroRNA microarray-based identification of involvement of miR-155 and miR-19a in development of oral lichen planus (OLP) by modulating Th1/Th2 balance via targeting eNOS and toll-like receptor 2 (TLR2). Med. Sci. Monit. 2018, 24, 3591–3603. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Zhao, Y.; Huo, X.; Wu, X. MiR-155-5p promotes fibroblast cell proliferation and inhibits FOXO signaling pathway in vulvar lichen sclerosis by targeting FOXO3 and CDKN1B. Gene 2018, 653, 43–50. [Google Scholar] [CrossRef]
- Lind, E.F.; Millar, D.G.; Dissanayake, D.; Savage, J.C.; Grimshaw, N.K.; Kerr, W.G.; Ohashi, P.S. MiR-155 upregulation in dendritic cells is sufficient to break tolerance in vivo by negatively regulating SHIP1. J. Immunol. 2015, 195, 4632–4640. [Google Scholar] [CrossRef] [Green Version]
- Ji, W.G.; Zhang, X.D.; Sun, X.D.; Wang, X.Q.; Chang, B.P.; Zhang, M.Z. MiRNA-155 modulates the malignant biological characteristics of NK/T-cell lymphoma cells by targeting FOXO3a gene. J. Huazhong Univ. Sci. Technolog. Med. Sci. 2014, 34, 882–888. [Google Scholar] [CrossRef]
- Hu, J.Y.; Zhang, J.; Ma, J.Z.; Liang, X.Y.; Chen, G.Y.; Lu, R.; Du, G.F.; Zhou, G. MicroRNA-155-IFN-gamma feedback loop in CD4(+)t cells of erosive type oral lichen planus. Sci. Rep. 2015, 5, 16935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terlou, A.; Santegoets, L.A.; van der Meijden, W.I.; Heijmans-Antonissen, C.; Swagemakers, S.M.; van der Spek, P.J.; Ewing, P.C.; van Beurden, M.; Helmerhorst, T.J.; Blok, L.J. An autoimmune phenotype in vulvar lichen sclerosus and lichen planus: A Th1 response and high levels of microRNA. J. Investig. Dermatol. 2012, 132, 658–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrestha, A.; Carraro, G.; El Agha, E.; Mukhametshina, R.; Chao, C.-M.; Rizvanov, A.; Barreto, G.; Bellusci, S. Generation and validation of miR-142 knock out mice. PLoS ONE 2015, 10, e0136913. [Google Scholar] [CrossRef]
- Islam, F.; Gopalan, V.; Vider, J.; Lu, C.-T.; Lam, A.K.-Y. MiR-142-5p act as an oncogenic microRNA in colorectal cancer: Clinicopathological and functional insights. Exp. Mol. Pathol. 2018, 104, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Su, S.; Zhao, Q.; He, C.; Huang, D.; Liu, J.; Chen, F.; Chen, J.; Liao, J.Y.; Cui, X.; Zeng, Y.; et al. MiR-142-5p and miR-130a-3p are regulated by IL-4 and IL-13 and control profibrogenic macrophage program. Nat. Commun. 2015, 6, 8523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Du, G.; Wang, Y.; Shi, L.; Mi, J.; Tang, G. Integrative analysis of mRNA and miRNA expression profiles in oral lichen planus: Preliminary results. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2017, 124, 390–402.e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Day, D.H.; Taylor, R.J.; Myre, M.A. Calmodulin and calmodulin binding proteins in dictyostelium: A primer. Int. J. Mol. Sci. 2020, 21, 1210. [Google Scholar] [CrossRef] [Green Version]
- Lingle, W.L.; Lukasiewicz, K.; Salisbury, J. Deregulation of the centrosome cycle and the origin of chromosomal instability in cancer. Chem. Biol. Pteridines Folates 2006, 570, 393–421. [Google Scholar] [CrossRef]
- Paksa, A.; Bandemer, J.; Hoeckendorf, B.; Razin, N.; Tarbashevich, K.; Minina, S.; Meyen, D.; Biundo, A.; Leidel, S.; Peyrieras, N.; et al. Repulsive cues combined with physical barriers and cell–cell adhesion determine progenitor cell positioning during organogenesis. Nat. Commun. 2016, 7, 11288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrington, C.B.; Patel, A.; Bacino, C.A.; Bowles, N.E. Haploinsufficiency of the LIM domain containing preferred translocation partner in lipoma (LPP) gene in patients with tetralogy of Fallot and VACTERL association. Am. J. Med Genet. Part A 2010, 152A, 2919–2923. [Google Scholar] [CrossRef] [PubMed]
- Petit, M.M.R.; Meulemans, S.M.P.; Alen, P.; Ayoubi, T.A.Y.; Jansen, E.; Van De Ven, W.J.M. The tumor suppressor Scrib interacts with the zyxin-related protein LPP, which shuttles between cell adhesion sites and the nucleus. BMC Cell Biol. 2005, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Tissue | Blood |
|---|---|---|
| 1 * | Normal + LS | Yes |
| 2 * | Normal + LS | Yes |
| 3 | N/A | Yes |
| 4 * | Normal + LS | Yes |
| 5 | N/A | Yes |
| 6 | N/A | Yes |
| 7 * | Normal + LS | Yes |
| 8 | N/A | Yes |
| 9 | N/A | Yes |
| 10 | N/A | Yes |
| 11 | N/A | Yes |
| 12 * | Normal + LS | Yes |
| 13 | N/A | Yes |
| 14 | N/A | Yes |
| 15 * | Normal + LS | Yes |
| 16 * | Normal + LS | Yes |
| 17 * | Normal + LS | Yes |
| 18 | N/A | Yes |
| 19 | N/A | Yes |
| 20 * | Normal + LS | Yes |
| 21 * | Normal + LS | Yes |
| 22 | N/A | Yes |
| 23 * | Normal + LS | Yes |
| 24 * | Normal + LS | Yes |
| 25 | Normal + LS | Yes |
| 26 | N/A | Yes |
| 27 | Normal + LS | Yes |
| 28 | N/A | Yes |
| 29 | N/A | Yes |
| 30 | Normal + LS | Yes |
| 31 | Normal + LS | Yes |
| 32 | Normal + LS | Yes |
| 33 | Normal + LS | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, X.; Ren, S.; Yang, C.; Ren, S.; Fu, M.Z.; Goldstein, A.R.; Li, X.; Mitchell, L.; Krapf, J.M.; Macri, C.J.; et al. Differentially Regulated miRNAs and Their Related Molecular Pathways in Lichen Sclerosus. Cells 2021, 10, 2291. https://doi.org/10.3390/cells10092291
Tan X, Ren S, Yang C, Ren S, Fu MZ, Goldstein AR, Li X, Mitchell L, Krapf JM, Macri CJ, et al. Differentially Regulated miRNAs and Their Related Molecular Pathways in Lichen Sclerosus. Cells. 2021; 10(9):2291. https://doi.org/10.3390/cells10092291
Chicago/Turabian StyleTan, Xiaohui, Shuyang Ren, Canyuan Yang, Shuchang Ren, Melinda Z. Fu, Amelia R. Goldstein, Xuelan Li, Leia Mitchell, Jill M. Krapf, Charles J. Macri, and et al. 2021. "Differentially Regulated miRNAs and Their Related Molecular Pathways in Lichen Sclerosus" Cells 10, no. 9: 2291. https://doi.org/10.3390/cells10092291
APA StyleTan, X., Ren, S., Yang, C., Ren, S., Fu, M. Z., Goldstein, A. R., Li, X., Mitchell, L., Krapf, J. M., Macri, C. J., Goldstein, A. T., & Fu, S. W. (2021). Differentially Regulated miRNAs and Their Related Molecular Pathways in Lichen Sclerosus. Cells, 10(9), 2291. https://doi.org/10.3390/cells10092291