
Impact of Oxysterols on Cell Death, Proliferation, and Differentiation Induction: Current Status

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
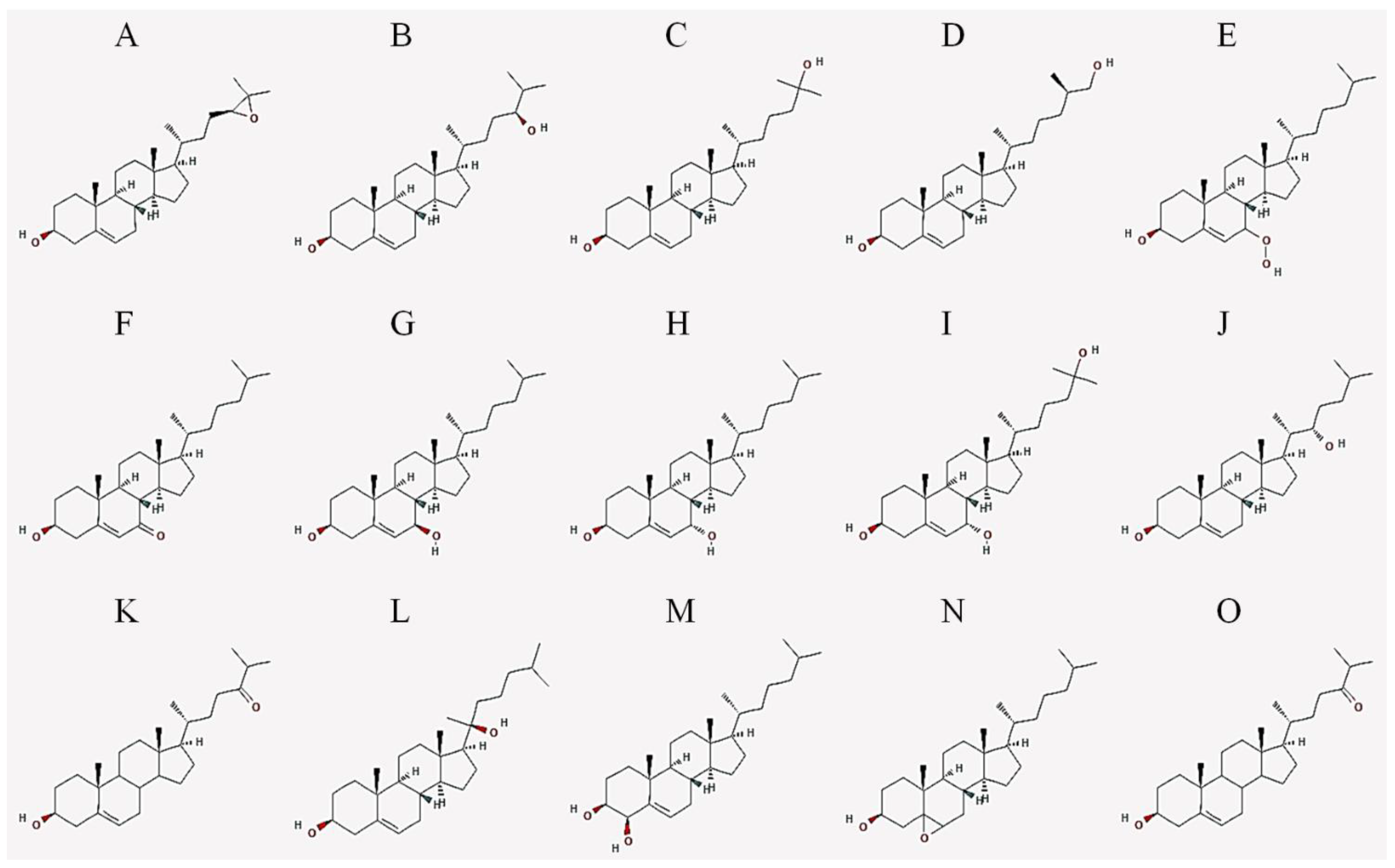
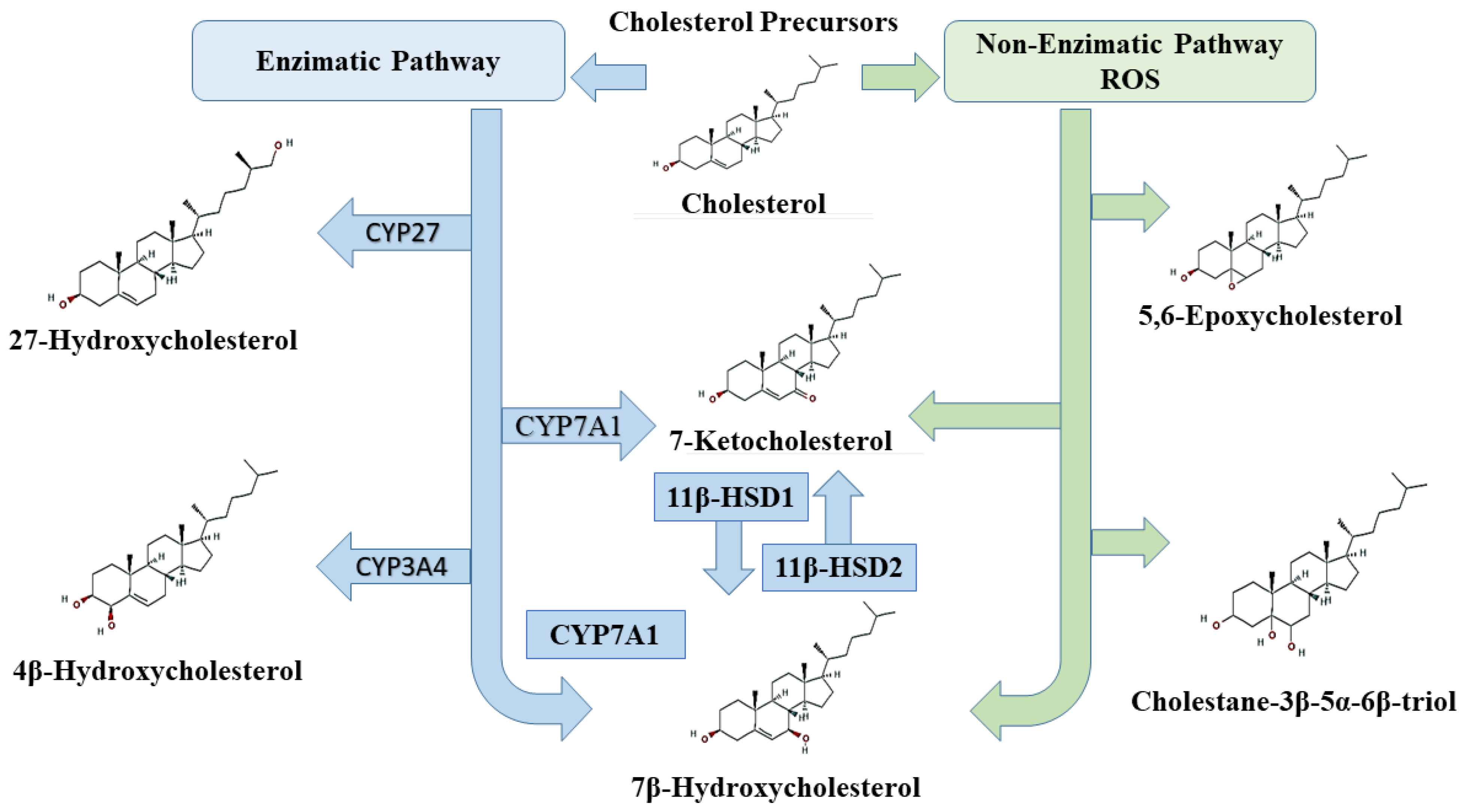
2. Oxysterols
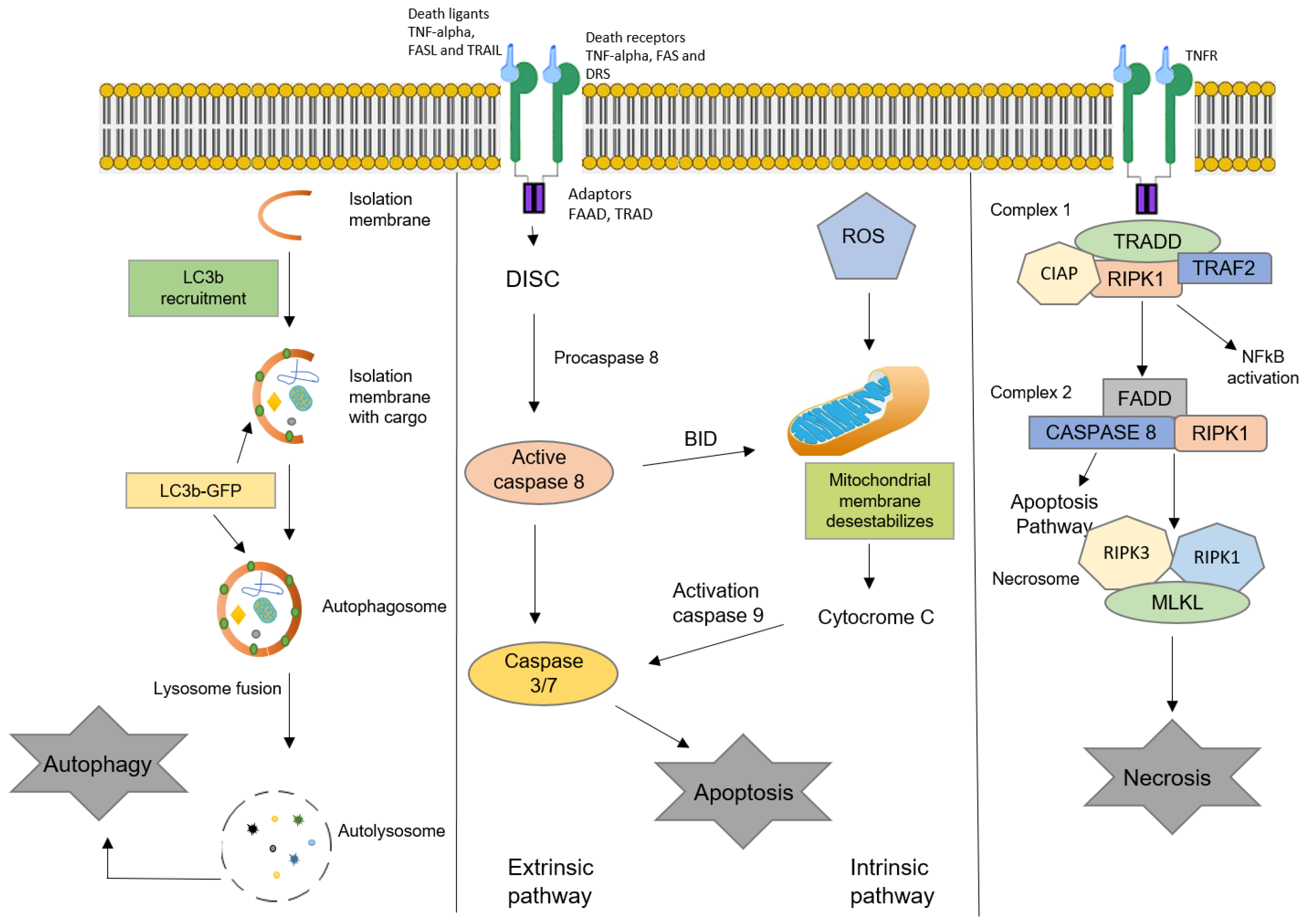
3. Oxysterols and Cell Death
3.1. Apoptosis
3.2. Autophagy
3.3. Necrosis
3.4. Main Oxysterols Involved in Cell Death
3.4.1. 7-Ketocholesterol (7-KC)
3.4.2. 7β-Hydroxycholesterol (7β-HC)
3.4.3. Cholestane-3β-5α-6β-triol
3.4.4. Effects of Other Oxysterols
4. Oxysterols and Cell Proliferation
5. Oxysterols and Cell Differentiation
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Faletrov, Y.V.; Efimova, V.S.; Horetski, M.; Tugaeva, K.V.; Frolova, N.S.; Lin, Q.; Isaeva, L.V.; Rubtsov, M.A.; Sluchanko, N.N.; Novikova, L.A.; et al. New 20-hydroxycholesterol-like compounds with fluorescent NBD or alkyne labels: Synthesis, in silico interactions with proteins and uptake by yeast cells. Chem. Phys. Lipids 2019, 227, 104850. [Google Scholar] [CrossRef]
- Levy, D.; Melo, T.; Ohira, B.Y.; Fidelis, M.L.; Ruiz, J.; Rodrigues, A.; Bydlowski, S.P. Oxysterols selectively promote short-term apoptosis in tumor cell lines. Biochem. Biophys. Res. Commun. 2018, 505, 1043–1049. [Google Scholar] [CrossRef]
- Perego, C.; Da Dalt, L.; Pirillo, A.; Galli, A.; Catapano, A.L.; Norata, G.D. Cholesterol metabolism, pancreatic β-cell function and diabetes. Biochimica Biophysica Acta (BBA)-Mol. Basis Dis. 2019, 1865, 2149–2156. [Google Scholar] [CrossRef] [PubMed]
- Ikonen, E. Cellular cholesterol trafficking and compartmentalization. Nat. Rev. Mol. Cell Biol. 2008, 9, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Yang, E.J.; Shim, J.S. Cholesterol Trafficking: An Emerging Therapeutic Target for Angiogenesis and Cancer. Cells 2019, 8, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulig, W.; Cwiklik, L.; Jurkiewicz, P.; Rog, T.; Vattulainen, I. Cholesterol oxidation products and their biological importance. Chem. Phys. Lipids 2016, 199, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Duc, D.; Vigne, S.; Pot, C. Oxysterols in autoimmunity. Int. J. Mol. Sci. 2019, 20, 4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, W.J.; Wang, Y. Oxysterols as lipid mediators: Their biosynthetic genes, enzymes and metabolites. Prostaglandins Other Lipid Mediat. 2019, 147, 106381. [Google Scholar] [CrossRef]
- Crick, P.J.; Yutuc, E.; Abdel-Khalik, J.; Saeed, A.; Betsholtz, C.; Genove, G.; Björkhem, I.; Wang, Y.; Griffiths, W.J. Formation and metabolism of oxysterols and cholestenoic acids found in the mouse circulation: Lessons learnt from deuterium-enrichment experiments and the CYP46A1 transgenic mouse. J. Steroid Biochem. Mol. Biol. 2019, 195, 105475. [Google Scholar] [CrossRef]
- Levy, D.; Melo, T.; Ruiz, J.; Bydlowski, S.P. Oxysterols and mesenchymal stem cell biology. Chem. Phys. Lipids 2017, 207, 223–230. [Google Scholar] [CrossRef]
- Zmysłowski, A.; Szterk, A. Oxysterols as a biomarker in diseases. Clin. Chim. Acta 2019, 491, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Ursan, R.; Odnoshivkina, U.G.; Petrov, A.M. Membrane cholesterol oxidation downregulates atrial β-adrenergic responses in ROS-dependent manner. Cell. Signal. 2020, 67, 109503. [Google Scholar] [CrossRef]
- Mutemberezi, V.; Guillemot-Legris, O.; Muccioli, G.G. Oxysterols: From cholesterol metabolites to key mediators. Prog. Lipid Res. 2016, 64, 152–169. [Google Scholar] [CrossRef]
- Riscal, R.; Skuli, N.; Simon, M.C. Even cancer cells watch their cholesterol! Mol. Cell 2019, 76, 220–231. [Google Scholar] [CrossRef]
- Nury, T.; Zarrouk, A.; Ragot, K.; Debbabi, M.; Riedinger, J.-M.; Vejux, A.; Aubourg, P.; Lizard, G. 7-Ketocholesterol is increased in the plasma of X-ALD patients and induces peroxisomal modifications in microglial cells: Potential roles of 7-ketocholesterol in the pathophysiology of X-ALD. J. Steroid Biochem. Mol. Biol. 2017, 169, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, P.; Holl, V.; Coelho, D.; Dufour, P.; Luu, B.; Weltin, D. Apoptosis at the Interface of Immunosuppressive and Anticancer Activities: The Examples of Two Classes of Chemical Inducers, Oxysterols and Alkylating Agents. Curr. Med. Chem. 2000, 7, 693–713. [Google Scholar] [CrossRef] [PubMed]
- Chimento, A.; Casaburi, I.; Avena, P.; Trotta, F.; De Luca, A.; Rago, V.; Pezzi, V.; Sirianni, R. Cholesterol and Its Metabolites in Tumor Growth: Therapeutic Potential of Statins in Cancer Treatment. Front. Endocrinol. 2019, 9, 807. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Estrada, M.; Cardenia, V.; Poirot, M.; Iuliano, L.; Lizard, G. Oxysterols and sterols: From lipidomics to food sciences. Elsevier 2020, 196, 105515. [Google Scholar] [CrossRef]
- Olkkonen, V.M.; Béaslas, O.; Nissilä, E. Oxysterols and Their Cellular Effectors. Biomolecules 2012, 2, 76–103. [Google Scholar] [CrossRef] [Green Version]
- Oguro, H. The Roles of Cholesterol and Its Metabolites in Normal and Malignant Hematopoiesis. Front. Endocrinol. 2019, 10, 204. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, W.J.; Wang, Y. An update on oxysterol biochemistry: New discoveries in lipidomics. Biochem. Biophys. Res. Commun. 2018, 504, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Kloudova, A.; Guengerich, F.P.; Soucek, P. The role of oxysterols in human cancer. Trends Endocrinol. Metab. 2017, 28, 485–496. [Google Scholar] [CrossRef]
- Hutchinson, S.A.; Lianto, P.; Moore, J.B.; Hughes, T.A.; Thorne, J.L. Phytosterols Inhibit Side-Chain Oxysterol Mediated Activation of LXR in Breast Cancer Cells. Int. J. Mol. Sci. 2019, 20, 3241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vejux, A.; Namsi, A.; Nury, T.; Moreau, T.; Lizard, G. Biomarkers of Amyotrophic Lateral Sclerosis: Current Status and Interest of Oxysterols and Phytosterols. Front. Mol. Neurosci. 2018, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Koppenol, W.P.; Knol, D.; Vermeer, M.A.; Hiemstra, H.; Friedrichs, S.; Lütjohann, D.; Trautwein, E.A.; Lin, K. Serum Concentration of Plant Sterol Oxidation Products (POP) Compared to Cholesterol Oxidation Products (COP) after Intake of Oxidized Plant Sterols: A Randomised, Placebo-Controlled, Double-Blind Dose‒Response Pilot Study. Nutrients 2019, 11, 2319. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.J.; Jessup, W. Oxysterols: Sources, cellular storage and metabolism, and new insights into their roles in cholesterol homeostasis. Mol. Asp. Med. 2009, 30, 111–122. [Google Scholar] [CrossRef]
- Ito, H.; Matsuo, K.; Hosono, S.; Watanabe, M.; Kawase, T.; Suzuki, T.; Hirai, T.; Yatabe, Y.; Tanaka, H.; Tajima, K. Association between CYP7A1 and the risk of proximal colon cancer in Japanese. Int. J. Mol. Epidemiol. Genet. 2009, 1, 35–46. [Google Scholar]
- Srivastava, A.; Choudhuri, G.; Mittal, B. CYP7A1 (−204 A>C; rs3808607 and −469 T>C; rs3824260) promoter polymorphisms and risk of gallbladder cancer in North Indian population. Metabolism 2010, 59, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-P.; Yao, F.; Luan, H.; Wang, Y.-L.; Dong, X.-H.; Zhou, W.-W.; Wang, Q.-H. CYP3A4* 1B polymorphism and cancer risk: A HuGE review and meta-analysis. Tumor Biol. 2013, 34, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhao, G.; Ouellet, J.; Fan, Z.; Labrie, F.; Pelletier, G. Expression of 11β-hydroxysteroid dehydrogenase type 1 in breast cancer and adjacent non-malignant tissue. An immunocytochemical study. Pathol. Oncol. Res. 2011, 17, 627–632. [Google Scholar] [CrossRef] [PubMed]
- Lipka, C.; Mankertz, J.; Fromm, M.; Lübbert, H.; Bühler, H.; Kühn, W.; Ragosch, V.; Hundertmark, S. Impairment of the antiproliferative effect of glucocorticosteroids by 11β-hydroxysteroid dehydrogenase type 2 overexpression in MCF-7 breast-cancer cells. Horm. Metab. Res. 2004, 36, 437–444. [Google Scholar] [CrossRef]
- Luu, W.; Sharpe, L.J.; Capell-Hattam, I.; Gelissen, I.C.; Brown, A.J. Oxysterols: Old tale, new twists. Annu. Rev. Pharmacol. Toxicol. 2016, 56, 447–467. [Google Scholar] [CrossRef] [PubMed]
- Kanner, J.; Lapidot, T. The stomach as a bioreactor: Dietary lipid peroxidation in the gastric fluid and the effects of plant-derived antioxidants. Free Radic. Biol. Med. 2001, 31, 1388–1395. [Google Scholar] [CrossRef]
- Dwyer, J.; Sever, N.; Carlson, M.; Nelson, S.F.; Beachy, P.A.; Parhami, F. Oxysterols Are Novel Activators of the Hedgehog Signaling Pathway in Pluripotent Mesenchymal Cells. J. Biol. Chem. 2007, 282, 8959–8968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Kang, J.; Shih, Y.; Lo, Y.; Wang, C. Cholesterol-3-beta, 5-alpha, 6-beta-triol induced genotoxicity through reactive oxygen species formation. Food Chem. Toxicol. 2005, 43, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Aghaloo, T.L.; Amantea, C.M.; Cowan, C.M.; Richardson, J.A.; Wu, B.M.; Parhami, F.; Tetradis, S. Oxysterols enhance osteoblast differentiation in vitro and bone healing in vivo. J. Orthop. Res. 2007, 25, 1488–1497. [Google Scholar] [CrossRef]
- Liu, X.; Tajima, N.; Taniguchi, M.; Kato, N. The enantiomer pair of 24S-and 24R-hydroxycholesterol differentially alter activity of large-conductance Ca2+-dependent K+ (slo1 BK) channel. Chirality 2020, 32, 223–230. [Google Scholar] [CrossRef]
- Amantea, C.M.; Kim, W.-K.; Meliton, V.; Tetradis, S.; Parhami, F. Oxysterol-induced osteogenic differentiation of marrow stromal cells is regulated by Dkk-1 inhibitable and PI3-kinase mediated signaling. J. Cell. Biochem. 2008, 105, 424–436. [Google Scholar] [CrossRef] [Green Version]
- Hong, W.; Guo, F.; Yang, M.; Xu, D.; Zhuang, Z.; Niu, B.; Bai, Q.; Li, X. Hydroxysteroid sulfotransferase 2B1 affects gastric epithelial function and carcinogenesis induced by a carcinogenic agent. Lipids Health Dis. 2019, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kloudova-Spalenkova, A.; Ueng, Y.-F.; Wei, S.; Kopeckova, K.; Guengerich, F.P.; Soucek, P. Plasma oxysterol levels in luminal subtype breast cancer patients are associated with clinical data. J. Steroid Biochem. Mol. Biol. 2019, 197, 105566. [Google Scholar] [CrossRef]
- Vejux, A.; Abed-Vieillard, D.; Hajji, K.; Zarrouk, A.; Mackrill, J.J.; Ghosh, S.; Nury, T.; Yammine, A.; Zaibi, M.; Mihoubi, W. 7-Ketocholesterol and 7β-hydroxycholesterol: In vitro and animal models used to characterize their activities and to identify molecules preventing their toxicity. Biochem. Pharmacol. 2020, 173, 113648. [Google Scholar] [CrossRef] [Green Version]
- Doria, M.; Maugest, L.; Moreau, T.; Lizard, G.; Vejux, A. Contribution of cholesterol and oxysterols to the pathophysiology of Parkinson’s disease. Free Radic. Biol. Med. 2016, 101, 393–400. [Google Scholar] [CrossRef]
- Jarvis, S.; Williamson, C.; Bevan, C.L. Liver X receptors and male (In) fertility. Int. J. Mol. Sci. 2019, 20, 5379. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Nelson, E.R. Oxysterols and nuclear receptors. Mol. Cell. Endocrinol. 2019, 484, 42–51. [Google Scholar] [CrossRef]
- Bensinger, S.J.; Bradley, M.N.; Joseph, S.; Zelcer, N.; Janssen, E.; Hausner, M.A.; Shih, R.; Parks, J.S.; Edwards, P.A.; Jamieson, B.D.; et al. LXR Signaling Couples Sterol Metabolism to Proliferation in the Acquired Immune Response. Cell 2008, 134, 97–111. [Google Scholar] [CrossRef] [Green Version]
- Sacchetti, P.; Sousa, K.M.; Hall, A.C.; Liste, I.; Steffensen, K.; Theofilopoulos, S.; Parish, C.; Hazenberg, C.; Richter, L.; Hovatta, O.; et al. Liver X Receptors and Oxysterols Promote Ventral Midbrain Neurogenesis In Vivo and in Human Embryonic Stem Cells. Cell Stem Cell 2009, 5, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, D.; Ruiz, J.; Celestino, A.T.; Silva, S.F.; Ferreira, A.K.; Isaac, C.; Bydlowski, S.P. Short-term effects of 7-ketocholesterol on human adipose tissue mesenchymal stem cells in vitro. Biochem. Biophys. Res. Commun. 2014, 446, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Brahmi, F.; Vejux, A.; Sghaier, R.; Zarrouk, A.; Nury, T.; Meddeb, W.; Rezig, L.; Namsi, A.; Sassi, K.; Yammine, A.; et al. Prevention of 7-ketocholesterol-induced side effects by natural compounds. Crit. Rev. Food Sci. Nutr. 2018, 59, 3179–3198. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Danial, N.N.; Hockenbery, D.M. Chapter 18—Cell death. In Hematology, 7th ed.; Hoffman, R., Benz, E.J., Silberstein, L.E., Heslop, H.E., Weitz, J.I., Anastasi, J., Salama, M.E., Abutalib, S.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 186–196. [Google Scholar]
- Zarrouk, A.; Ben Salem, Y.; Hafsa, J.; Sghaier, R.; Charfeddine, B.; Limem, K.; Hammami, M.; Majdoub, H. 7β-hydroxycholesterol-induced cell death, oxidative stress, and fatty acid metabolism dysfunctions attenuated with sea urchin egg oil. Biochimie 2018, 153, 210–219. [Google Scholar] [CrossRef]
- Paz, J.L.; Levy, D.; Oliveira, B.A.; Melo, T.; De Freitas, F.A.; Reichert, C.O.; Rodrigues, A.; Pereira, J.; Bydlowski, S.P. 7-Ketocholesterol Promotes Oxiapoptophagy in Bone Marrow Mesenchymal Stem Cell from Patients with Acute Myeloid Leukemia. Cells 2019, 8, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, B.; Agnihotri, N. Role of cholesterol homeostasis and its efflux pathways in cancer progression. J. Steroid Biochem. Mol. Biol. 2019, 191, 105377. [Google Scholar] [CrossRef] [PubMed]
- Vejux, A.; Malvitte, L.; Lizard, G. Side effects of oxysterols: Cytotoxicity, oxidation, inflammation, and phospholipidosis. Braz. J. Med. Biol. Res. 2008, 41, 545–556. [Google Scholar] [CrossRef] [Green Version]
- Green, D.R.; Llambi, F. Cell death signaling. Cold Spring Harbor Perspect. Biol. 2015, 7, a006080. [Google Scholar] [CrossRef]
- Castelli, G.; Pelosi, E.; Testa, U. Emerging Therapies for Acute Myelogenus Leukemia Patients Targeting Apoptosis and Mitochondrial Metabolism. Cancers 2019, 11, 260. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, L.M.; Smith, S.W.; Jones, M.; Osborne, B.A. Do all programmed cell deaths occur via apoptosis? Proc. Natl. Acad. Sci. USA 1993, 90, 980–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposti, M.D. Apoptosis: Who was first? Cell Death Differ. 1998, 5, 719. [Google Scholar] [CrossRef]
- Wlodkowic, D.; Skommer, J.; Darzynkiewicz, Z. Cytometry of apoptosis. Historical perspective and new advances. Exp. Oncol. 2012, 34, 255–262. [Google Scholar]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wideranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, A.; Nomura, M.; Shuck, R.; Yustein, J. Cancer’s Achilles’ heel: Apoptosis and necroptosis to the rescue. Int. J. Mol. Sci. 2017, 18, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Ashrafi, G.H.; Schwarz, T.L. The pathways of mitophagy for quality control and clearance of mitochondria. Cell Death Differ. 2012, 20, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsetti, G.; Chen-Scarabelli, C.; Romano, C.; Pasini, E.; Dioguardi, F.S.; Onorati, F.; Knight, R.; Patel, H.; Saravolatz, L.; Faggian, G.; et al. Autophagy and Oncosis/Necroptosis Are Enhanced in Cardiomyocytes from Heart Failure Patients. Med. Sci. Monit. Basic Res. 2019, 25, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Kirkin, V. History of the selective autophagy research: How did it begin and where does it stand today? J. Mol. Biol. 2020, 432, 3–27. [Google Scholar] [CrossRef]
- Jha, S.; Kitsis, R.N. Myocardial Basis for Heart Failure. Role of Cell Death. In Heart Failure; Elsevier Inc.: Philadelphia, PA, USA, 2011; pp. 85–102. [Google Scholar]
- Klionsky, D.J. Autophagy revisited: A conversation with Christian de Duve. Autophagy 2008, 4, 740–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.; Cai, N.; Li, M.; Liu, G.H.; Izpisua Belmonte, J.C. Autophagic control of cell ‘stemness’. EMBO Mol. Med. 2013, 5, 327–331. [Google Scholar] [CrossRef]
- Gump, J.M.; Thorburn, A. Autophagy and apoptosis: What is the connection? Trends Cell Biol. 2011, 21, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Kuma, A.; Hatano, M.; Matsui, M.; Yamamoto, A.; Nakaya, H.; Yoshimori, T.; Ohsumi, Y.; Tokuhisa, T.; Mizushima, N. The role of autophagy during the early neonatal starvation period. Nature 2004, 432, 1032–1036. [Google Scholar] [CrossRef]
- Yun, C.W.; Lee, S.H. The roles of autophagy in cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef] [Green Version]
- Stenson, W.F.; Ciorba, M.A. Chapter 9—Cell death. In Physiology of the Gastrointestinal Tract, 6th ed.; Said, H.M., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 221–234. [Google Scholar]
- Rock, K.L.; Kono, H. The inflammatory response to cell death. Annu. Rev. Pathol. Mech. Dis. 2008, 3, 99–126. [Google Scholar] [CrossRef]
- Proskuryakov, S.Y.; Konoplyannikov, A.G.; Gabai, V.L. Necrosis: A specific form of programmed cell death? Exp. Cell. Res. 2003, 283, 1–16. [Google Scholar] [CrossRef]
- Mbaveng, A.; Bitchagno, G.T.; Kuete, V.; Tane, P.; Efferth, T. Cytotoxicity of ungeremine towards multi-factorial drug resistant cancer cells and induction of apoptosis, ferroptosis, necroptosis and autophagy. Phytomedicine 2019, 60, 152832. [Google Scholar] [CrossRef]
- Monier, S.; Samadi, M.; Prunet, C.; Denance, M.; Laubriet, A.; Athias, A.; Berthier, A.; Steinmetz, E.; Jürgens, G.; Nègre-Salvayre, A. Impairment of the cytotoxic and oxidative activities of 7β-hydroxycholesterol and 7-ketocholesterol by esterification with oleate. Biochem. Biophys. Res. Commun. 2003, 303, 814–824. [Google Scholar] [CrossRef]
- Testa, G.; Rossin, D.; Poli, G.; Biasi, F.; Leonarduzzi, G.M. Implication of oxysterols in chronic inflammatory human diseases. Biochimie 2018, 153, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Nury, T.; Zarrouk, A.; Vejux, A.; Doria, M.; Riedinger, J.M.; Delage-Mourroux, R.; Lizard, G. Induction of oxiapoptophagy, a mixed mode of cell death associated with oxidative stress, apoptosis and autophagy, on 7-ketocholesterol-treated 158N murine oligodendrocytes: Impairment by α-tocopherol. Biochem. Biophys. Res. Commun. 2014, 446, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Nury, T.; Zarrouk, A.; Mackrill, J.J.; Samadi, M.; Durand, P.; Riedinger, J.-M.; Doria, M.; Vejux, A.; Limagne, E.; Delmas, D. Induction of oxiapoptophagy on 158N murine oligodendrocytes treated by 7-ketocholesterol-, 7β-hydroxycholesterol-, or 24 (S)-hydroxycholesterol: Protective effects of α-tocopherol and docosahexaenoic acid (DHA.; C22: 6 n-3). Steroids 2015, 99, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Favero, G.M.; Paz, J.L.; Otake, A.H.; Maria, D.A.; Caldini, E.; De Medeiros, R.S.; Deus, D.F.; Chammas, R.; Maranhão, R.C.; Bydlowski, S.P. Cell internalization of 7-ketocholesterol-containing nanoemulsion through LDL receptor reduces melanoma growth in vitro and in vivo: A preliminary report. Oncotarget 2018, 9, 14160–14174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, L.R.; Stern, A.C.B.; Cavaglieri, R.D.C.; Nogueira, F.C.S.; Domont, G.; Palmisano, G.; Bydlowski, S.P. 7-Ketocholesterol overcomes drug resistance in chronic myeloid leukemia cell lines beyond MDR1 mechanism. J. Proteom. 2017, 151, 12–23. [Google Scholar] [CrossRef]
- He, C.; Zhu, H.; Zhang, W.; Okon, I.; Wang, Q.; Li, H.; Le, Y.-Z.; Xie, Z. 7-Ketocholesterol Induces Autophagy in Vascular Smooth Muscle Cells through Nox4 and Atg4B. Am. J. Pathol. 2013, 183, 626–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiduschka, G.; Erovic, B.M.; Vormittag, L.; Skoda, C.; Martinek, H.; Brunner, M.; Ehrenberger, K.; Thurnher, D. 7β-hydroxycholesterol induces apoptosis and regulates cyclooxygenase 2 in head and neck squamous cell carcinoma. Arch. Otolaryngol.–Head Neck Surg. 2009, 135, 261–267. [Google Scholar] [CrossRef]
- Lordan, S.; Mackrill, J.J.; O’Brien, N.M. Involvement of Fas Signalling in 7 β-Hydroxycholesterol-and Cholesterol-5 β, 6 β-Epoxide-Induced Apoptosis. Int. J. Toxicol. 2008, 27, 279–285. [Google Scholar] [CrossRef]
- Sassi, K.; Nury, T.; Zarrouk, A.; Sghaier, R.; Khalafi-Nezhad, A.; Vejux, A.; Samadi, M.; Ben Aissa-Fennira, F.; Lizard, G. Induction of a non-apoptotic mode of cell death associated with autophagic characteristics with steroidal maleic anhydrides and 7β-hydroxycholesterol on glioma cells. J. Steroid Biochem. Mol. Biol. 2019, 191, 105371. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-Y.; Huo, C.; Kuo, L.-K.; Hiipakka, R.A.; Jones, R.B.; Lin, H.-P.; Hung, Y.-W.; Su, L.-C.; Tseng, J.-C.; Kuo, Y.-Y.; et al. Cholestane-3β, 5α, 6β-triol Suppresses Proliferation, Migration, and Invasion of Human Prostate Cancer Cells. PLoS ONE 2013, 8, e65734. [Google Scholar] [CrossRef] [Green Version]
- Vejux, A.; Lizard, G. Cytotoxic effects of oxysterols associated with human diseases: Induction of cell death (apoptosis and/or oncosis), oxidative and inflammatory activities, and phospholipidosis. Mol. Asp. Med. 2009, 30, 153–170. [Google Scholar] [CrossRef]
- Nakazawa, T.; Miyanoki, Y.; Urano, Y.; Uehara, M.; Saito, Y.; Noguchi, N. Effect of vitamin E on 24(S)-hydroxycholesterol-induced necroptosis-like cell death and apoptosis. J. Steroid Biochem. Mol. Biol. 2016, 169, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.F.; Levy, D.; Ruiz, J.; Melo, T.; Isaac, C.; Fidelis, M.L.; Rodrigues, A.; Bydlowski, S.P. Oxysterols in adipose tissue-derived mesenchymal stem cell proliferation and death. J. Steroid Biochem. Mol. Biol. 2017, 169, 164–175. [Google Scholar] [CrossRef]
- Fernandes, L.R.; Maselli, L.M.; Maeda, N.Y.; Palmisano, G.; Bydlowski, S.P. Outside-in, inside-out: Proteomic analysis of endothelial stress mediated by 7-ketocholesterol. Chem. Phys. Lipids 2017, 207, 231–238. [Google Scholar] [CrossRef]
- Brahmi, F.; Nury, T.; Debbabi, M.; Hadj-Ahmed, S.; Zarrouk, A.; Prost, M.; Madani, K.; Boulekbache-Makhlouf, L.; Lizard, G. Evaluation of Antioxidant, Anti-Inflammatory and Cytoprotective Properties of Ethanolic Mint Extracts from Algeria on 7-Ketocholesterol-Treated Murine RAW 264.7 Macrophages. Antioxidants 2018, 7, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarrouk, A.; Vejux, A.; Mackrill, J.; O’Callaghan, Y.; Hammami, M.; O’Brien, N.; Lizard, G. Involvement of oxysterols in age-related diseases and ageing processes. Ageing Res. Rev. 2014, 18, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.; de Melo, T.C.; Oliveira, B.A.; Paz, J.L.; de Freitas, F.A.; Reichert, C.O.; Rodrigues, A.; Bydlowski, S.P. 7-Ketocholesterol and cholestane-triol increase expression of SMO and LXRα signaling pathways in a human breast cancer cell line. Biochem. Biophys. Rep. 2019, 19, 100604. [Google Scholar] [CrossRef]
- Nury, T.; Zarrouk, A.; Yammine, A.; Mackrill, J.J.; Vejux, A.; Lizard, G. Oxiapoptophagy: A type of cell death induced by some oxysterols. Br. J. Pharmacol. 2021, 178, 3115–3123. [Google Scholar] [CrossRef]
- Nury, T.; Yammine, A.; Ghzaiel, I.; Sassi, K.; Zarrouk, A.; Brahmi, F.; Samadi, M.; Rup-Jacques, S.; Vervandier-Fasseur, D.; de Barros, J.P. Attenuation of 7-ketocholesterol-and 7β-hydroxycholesterol-induced oxiapoptophagy by nutrients, synthetic molecules and oils: Potential for the prevention of age-related diseases. Ageing Res. Rev. 2021, 68, 101324. [Google Scholar] [CrossRef]
- Hu, H.; Zhou, Y.; Leng, T.; Liu, A.; Wang, Y.; You, X.; Chen, J.; Tang, L.; Chen, W.; Qiu, P. The major cholesterol metabolite cholestane-3β, 5α, 6β-triol functions as an endogenous neuroprotectant. J. Neurosci. 2014, 34, 11426–11438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urano, Y.; Vo, D.-K.H.; Hirofumi, A.; Noguchi, N. 24(S)-Hydroxycholesterol induces ER dysfunction-mediated unconventional cell death. Cell Death Discov. 2019, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- De Weille, J.; Fabre, C.; Bakalara, N. Oxysterols in cancer cell proliferation and death. Biochem. Pharmacol. 2013, 86, 154–160. [Google Scholar] [CrossRef]
- El Roz, A.; Bard, J.-M.; Huvelin, J.-M.; Nazih, H. LXR agonists and ABCG1-dependent cholesterol efflux in MCF-7 breast cancer cells: Relation to proliferation and apoptosis. Anticancer. Res. 2012, 32, 3007–3013. [Google Scholar] [PubMed]
- Ruiz, J.L.; Fernandes, L.R.; Levy, D.; Bydlowski, S.P. Interrelationship between ATP-binding cassette transporters and oxysterols. Biochem. Pharmacol. 2013, 86, 80–88. [Google Scholar] [CrossRef]
- Shahoei, S.H.; Nelson, E.R. Nuclear receptors, cholesterol homeostasis and the immune system. J. Steroid Biochem. Mol. Biol. 2019, 191, 105364. [Google Scholar] [CrossRef]
- Javitt, N.B. 26-Hydroxycholesterol: Synthesis, metabolism, and biologic activities. J. Lipid Res. 1990, 31, 1527–1533. [Google Scholar] [CrossRef]
- Griffiths, W.J.; Wang, Y. Oxysterol research: A brief review. Biochem. Soc. Trans. 2019, 47, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Baek, A.E.; Yu, Y.-R.A.; He, S.; Wardell, S.E.; Chang, C.-Y.; Kwon, S.; Pillai, R.V.; McDowell, H.B.; Thompson, J.W.; Dubois, L.G.; et al. The cholesterol metabolite 27 hydroxycholesterol facilitates breast cancer metastasis through its actions on immune cells. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Sirianni, R.; Umetani, M.; Pezzi, V. Editorial: Cholesterol and Oxysterols as Signal Molecules in Human Pathophysiology and Cancer: Implications for New Therapeutic Strategies. Front. Endocrinol. 2019, 10, 732. [Google Scholar] [CrossRef]
- Soules, R.; Noguer, E.; Iuliano, L.; Zerbinati, C.; Leignadier, J.; Rives, A.; de Medina, P.; Silvente-Poirot, S.; Poirot, M. Improvement of 5, 6α-epoxycholesterol, 5, 6β-epoxycholesterol, cholestane-3β, 5α, 6β-triol and 6-oxo-cholestan-3β, 5α-diol recovery for quantification by GC/MS. Chem. Phys. Lipids 2017, 207, 92–98. [Google Scholar] [CrossRef]
- Kim, M.-W.; An, B.M.; Wang, L.; Lee, K.Y.; Bang, J.; Joo, B.S. Effects of Oxysterols on Chondrogenesis of Human Adipose-derived Stem Cells. Ann. of Clin. Lab. Sci. 2020, 50, 190–198. [Google Scholar]
- Moseti, D.; Regassa, A.; Chen, C.; Kim, W.K. 25-Hydroxycholesterol Inhibits Adipogenic Differentiation of C3H10T1/2 Pluripotent Stromal Cells. Int. J. Mol. Sci. 2020, 21, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevi, L.; Costantini, D.; Safarikia, S.; DI Matteo, S.; Melandro, F.; Berloco, P.B.; Cardinale, V. Cholest-4,6-Dien-3-One Promote Epithelial-To-Mesenchymal Transition (EMT) in Biliary Tree Stem/Progenitor Cell Cultures In Vitro. Cells 2019, 8, 1443. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Lin, Y.; Rong, M.; Liu, W.; He, J.; Zhou, L. 20(S)-hydroxycholesterol and simvastatin synergistically enhance osteogenic differentiation of marrow stromal cells and bone regeneration by initiation of Raf/MEK/ERK signaling. J. Mater. Sci. Mater. Med. 2019, 30, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Kim, B.-Y.; Eo, S.-K.; Park, Y.C.; Kim, K. Dexamethasone Suppresses Oxysterol-Induced Differentiation of Monocytic Cells. Oxidative Med. Cell. Longev. 2016, 2016, 2915382. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, M.; Liu, J.; Li, X.; Yang, M.; Su, B.; Lin, Y. 27-Hydroxycholesterol enhanced osteoclastogenesis in lung adenocarcinoma microenvironment. J. Cell. Physiol. 2018, 234, 12692–12700. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.; DuSell, C.D.; Wang, X.; Howe, M.K.; Evans, G.; Michalek, R.D.; Umetani, M.; Rathmell, J.C.; Khosla, S.; Gesty-Palmer, D.; et al. The Oxysterol, 27-Hydroxycholesterol, Links Cholesterol Metabolism to Bone Homeostasis through Its Actions on the Estrogen and Liver X Receptors. Endocrinology 2011, 152, 4691–4705. [Google Scholar] [CrossRef] [PubMed]
- Theofilopoulos, S.; de Oliveira, W.A.A.; Yang, S.; Yutuc, E.; Saeed, A.; Abdel-Khalik, J.; Ullgren, A.; Cedazo-Minguez, A.; Björkhem, I.; Wang, Y.; et al. 24(S),25-Epoxycholesterol and cholesterol 24S-hydroxylase (CYP46A1) overexpression promote midbrain dopaminergic neurogenesis in vivo. J. Biol. Chem. 2019, 294, 4169–4176. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-S.; Kim, E.; Han, S.; Kang, K.L.; Heo, J.S. Evaluating the oxysterol combination of 22(S)-hydroxycholesterol and 20(S)-hydroxycholesterol in periodontal regeneration using periodontal ligament stem cells and alveolar bone healing models. Stem Cell Res. Ther. 2017, 8, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murdolo, G.; Piroddi, M.; Tortoioli, C.; Bartolini, D.; Schmelz, M.; Luchetti, F.; Canonico, B.; Papa, S.; Zerbinati, C.; Iuliano, L.; et al. Free Radical-derived Oxysterols: Novel Adipokines Modulating Adipogenic Differentiation of Adipose Precursor Cells. J. Clin. Endocrinol. Metab. 2016, 101, 4974–4983. [Google Scholar] [CrossRef] [PubMed]
- Marengo, B.; Bellora, F.; Ricciarelli, R.; De Ciucis, C.; Furfaro, A.; Leardi, R.; Colla, R.; Pacini, D.; Traverso, N.; Moretta, A.; et al. Oxysterol mixture and, in particular, 27-hydroxycholesterol drive M2 polarization of human macrophages. BioFactors 2015, 42, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Wöltje, M.; Böbel, M.; Heiland, M.; Beck-Broichsitter, B.; Al-Dam, A.; Gröbe, A.; Friedrich, R.E.; Hanken, H.; Smeets, R. Purmorphamine and oxysterols accelerate and promote osteogenic differentiation of mesenchymal stem cells in vitro. In Vivo 2015, 29, 247–254. [Google Scholar]
- Kwon, I.K.; Lee, S.C.; Hwang, Y.-S.; Heo, J.S. Mitochondrial function contributes to oxysterol-induced osteogenic differentiation in mouse embryonic stem cells. Biochim. Biophys. Acta (BBA) Bioenerg. 2015, 1853, 561–572. [Google Scholar] [CrossRef] [Green Version]
- Soroosh, P.; Wu, J.; Xue, X.; Song, J.; Sutton, S.W.; Sablad, M.; Yu, J.; Nelen, M.I.; Liu, X.; Castro, G. Oxysterols are agonist ligands of RORγt and drive Th17 cell differentiation. Proc. Natl. Acad. Sci. USA 2014, 111, 12163–12168. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, S.R.; Nargizyan, T.; Meliton, V.; Nachtergaele, S.; Rohatgi, R.; Stappenbeck, F.; Jung, M.E.; Johnson, J.S.; Aghdasi, B.; Tian, H.; et al. A Novel Osteogenic Oxysterol Compound for Therapeutic Development to Promote Bone Growth: Activation of Hedgehog Signaling and Osteogenesis through Smoothened Binding. J. Bone Miner. Res. 2014, 29, 1872–1885. [Google Scholar] [CrossRef] [Green Version]
- Hokugo, A.; Sorice, S.; Yalom, A.; Lee, J.C.; Li, A.; Zuk, P.; Jarrahy, R. In Vitro Study of a Novel Oxysterol for Osteogenic Differentiation on Rabbit Bone Marrow Stromal Cells. Plast. Reconstr. Surg. 2013, 132, 70e–80e. [Google Scholar] [CrossRef]
- Asghari, A.; Ishikawa, T.; Hiramitsu, S.; Lee, W.-R.; Umetani, J.; Bui, L.; Korach, K.S.; Umetani, M. 27-Hydroxycholesterol Promotes Adiposity and Mimics Adipogenic Diet-Induced Inflammatory Signaling. Endocrinology 2019, 160, 2485–2494. [Google Scholar] [CrossRef] [Green Version]
- Son, Y.; Choi, J.; Kim, B.; Park, Y.C.; Eo, S.-K.; Cho, H.-R.; Bae, S.S.; Kim, C.D.; Kim, K. Cyclosporin A inhibits differentiation and activation of monocytic cells induced by 27-hydroxycholesterol. Int. Immunopharmacol. 2019, 69, 358–367. [Google Scholar] [CrossRef]
- Aye, I.; Waddell, B.; Mark, P.; Keelan, J. Oxysterols inhibit differentiation and fusion of term primary trophoblasts by activating liver X receptors. Placenta 2011, 32, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Jamadagni, P.; Patten, S.A. 25-hydroxycholesterol impairs neuronal and muscular development in zebrafish. NeuroToxicology 2019, 75, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Rossin, D.; Calfapietra, S.; Sottero, B.; Poli, G.; Biasi, F. HNE and cholesterol oxidation products in colorectal inflammation and carcinogenesis. Free Radic. Biol. Med. 2017, 111, 186–195. [Google Scholar] [CrossRef]
- Sunjic, S.; Gasparovic, A.; Jaganjac, M.; Rechberger, G.; Meinitzer, A.; Grune, T.; Kohlwein, S.; Mihaljevic, B.; Zarkovic, N. Sensitivity of Osteosarcoma Cells to Concentration-Dependent Bioactivities of Lipid Peroxidation Product 4-Hydroxynonenal Depend on their Level of Differentiation. Cells 2021, 10, 269. [Google Scholar] [CrossRef]
- Grimsrud, P.A.; Xie, H.; Griffin, T.; Bernlohr, D.A. Oxidative Stress and Covalent Modification of Protein with Bioactive Aldehydes. J. Biol. Chem. 2008, 283, 21837–21841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poli, G.; Schaur, R.J.; Siems, W.G.; Leonarduzzi, G.M. 4-Hydroxynonenal: A membrane lipid oxidation product of medicinal interest. Med. Res. Rev. 2008, 28, 569–631. [Google Scholar] [CrossRef]
- Dianzani, M.U.; Barrera, G.; Parola, M. 4-Hydroxy-2,3-nonenal as a signal for cell function and differentiation. Acta Biochim. Pol. 1999, 46, 61–75. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Common Name | IUPAC Name |
|---|---|---|
| 24,25-EC | 24(S),25-epoxycholesterol | (3S,8S,9S,10R,13R,14S,17R)-17-[(2R)-4-[(2S)-3,3-dimethyloxiran-2-yl]butan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol |
| 24-HC | 24(S)-hydroxycholesterol | (3S,8S,9S,10R,13R,14S,17R)-17-[(2R,5S)-5-hydroxy-6-methylheptan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol |
| 25-HC | 25-hydroxycholesterol | (3S,8S,9S,10R,13R,14S,17R)-17-[(2R)-6-hydroxy-6-methylheptan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol |
| 27-HC | 27-hydroxycholesterol | (3S,8S,9S,10R,13R,14S,17R)-17-[(2R,6R)-7-hydroxy-6-methylheptan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol |
| 7-OOHC | 7-hydroperoxycholesterol | (3S,8S,9S,10R,13R,14S,17R)-7-hydroperoxy-10,13-dimethyl-17-[(2R)-6-methylheptan-2-yl]-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol |
| 7α-HC | 7α-hydroxycholesterol | (3S,7S,8S,9S,10R,13R,14S,17R)-10,13-dimethyl-17-[(2R)-6-methylheptan-2-yl]-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthrene-3,7-diol |
| 7β-HC | 7β-hydroxycholesterol | (3S,4R,8S,9S,10R,13R,14S,17R)-10,13-dimethyl-17-[(2R)-6-methylheptan-2-yl]-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthrene-3,4-diol |
| 7-KC | 7-ketocholesterol | (3S,8S,9S,10R,13R,14S,17R)-3-hydroxy-10,13-dimethyl-17-[(2R)-6-methylheptan-2-yl]-1,2,3,4,8,9,11,12,14,15,16,17-dodecahydrocyclopenta[a]phenanthren-7-one |
| 7α,25-DHC | 7α,25-dihydroxycholesterol | (3S,7S,8S,9S,10R,13R,14S,17R)-17-[(2R)-6-hydroxy-6-methylheptan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthrene-3,7-diol |
| 7β,27-DHC | 7β,27-dihydroxycholesterol | (3S,7R,8S,9S,10R,13R,14S,17R)-17-[(2R)-7-hydroxy-6-methylheptan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthrene-3,7-diol |
| 22-HC | 22(S)-hydroxycholesterol | (3S,8S,9S,10R,13S,14S,17R)-17-[(2S,3S)-3-hydroxy-6-methylheptan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol |
| 20-HC | 20(S)-hydroxycholesterol | (3S,8S,9S,10R,13S,14S,17S)-17-[(2S)-2-hydroxy-6-methylheptan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol |
| 4β-HC | 4β-hydroxycholesterol | (3S,4R,8S,9S,10R,13R,14S,17R)-10,13-dimethyl-17-[(2R)-6-methylheptan-2-yl]-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthrene-3,4-diol |
| 7,25-DHC | 7,25-dihydroxycholesterol | (3S,8S,9S,10R,13R,14S,17R)-17-[(2R)-6-hydroxy-6-methylheptan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthrene-3,7-diol |
| 5,6-EC | 5,6-epoxycholesterol | (1S,2R,5S,11S,12S,15R,16R)-2,16-dimethyl-15-[(2R)-6-methylheptan-2-yl]-8-oxapentacyclo[9.7.0.02,7.07,9.012,16]octadecan-5-ol |
| 24-OXO | 24-oxocholesterol | (6R)-6-[(3S,10R,13R,17R)-3-hydroxy-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-17-yl]-2-methylheptan-3-one |
| Oxysterol | Cell Death | Cell Lineage or Animal Model | References |
|---|---|---|---|
| 7-ketocholesterol | Oxiapoptophagy | MSC * | [52] |
| Human monocytic U937 cells | [77,78] | ||
| Murine oligodendrocytes 158N cells | [79,80] | ||
| Short-term apoptosis | MSC ** | [47] | |
| Mus musculus skin melanoma cells (B16-F10) | [2,81] | ||
| Human mammary gland/breast cells (MDA-MB-231) | [2] | ||
| Human endothelial umbilical vein/vascular endothelium cells (HUV-EC-C) | [2] | ||
| Apoptosis | Lucena | [82] | |
| Phospholipidosis | Human monocytic U937 cells | [54] | |
| Autophagy | Human aortic smooth muscle cells (HASMCs) | [83] | |
| 7β-hydroxycholesterol | Oxiapoptophagy | Human monocytic U937 cells | [77] |
| Murine oligodendrocytes 158N cells | [80] | ||
| Cell death associated with oxidative stress, and metabolic dysfunctions | Murine oligodendrocytes 158N cells | [51] | |
| Apoptosis | Human tongue squamous cell carcinoma (SCC9) | [84] | |
| Human tongue squamous cell carcinoma (SCC25) | [84] | ||
| Human tongue squamous cell carcinoma (CAL27) | [84] | ||
| Human pharynx squamous cell carcinoma FaDu | [84] | ||
| Human monocytic U937 cells | [85] | ||
| Cell death process associated with several characteristics of survival autophagy | C6 rat glioma cells | [86] | |
| 24(S)-hydroxycholesterol | Oxiapoptophagy | Murine oligodendrocytes 158N cells | [80] |
| Cholestane-3β-5α-6β-triol | Short-term apoptosis | Mus musculus skin melanoma cells (B16-F10) | [2] |
| Human mammary gland/breast cells (MDA-MB-231) | [2] | ||
| Human endothelial umbilical vein/vascular endothelium cells (HUV-EC-C) | [2] | ||
| Human lung fibroblasts (LL 24) | [2] | ||
| Apoptosis | Human prostate cancer cell line (PC-3) | [87] | |
| Human prostate cancer cell line (DU-145 | [87] | ||
| Human LNCaP CDXR-3 | [87] | ||
| Rat bone marrow cells | [88] | ||
| 5α-cholestane-3β,6β-diol | Short-term apoptosis | Mus musculus skin melanoma cells (B16-F10) | [2] |
| Human mammary gland/breast cells (MDA-MB-231) | [2] | ||
| Human endothelial umbilical vein/vascular endothelium cells (HUV-EC-C) | [2] | ||
| Human lung fibroblasts (LL 24) | [2] | ||
| 25-hydroxycholesterol | Apoptosis | Human acute lymphoblastic leukemia T lymphoblast (CEM-C1) | [16] |
| Human acute lymphoblastic leukemia T lymphoblast (CEM-C7) | [16] | ||
| Murine thymocytes | [16] | ||
| 27-hydroxycholesterol | Apoptosis | C6 glioma cells | [53] |
| Oxisterol mixture *** | Apoptosis | Human monocytic U937 cells | [78] |
| 24-hydroxycholesterol | necroptosis-like | human neuroblastoma cells (SH-SY5Y) | [89] |
| Oxysterol | Differentiation | Cell Lineage or Animal Model | References |
|---|---|---|---|
| Cholestan-3β,5α,6β-triol | ↓ Osteogenesis | Rat bone marrow cells | [88] |
| 20(S)-hydroxycholesterol | ↑ Osteogenesis | Bone marrow stromal cells | [10] |
| Embryonic stem cells | [10] | ||
| ↓ Adipogenesis | Bone marrow stromal cells | [10] | |
| 22(R)-hydroxycholesterol | ↑ Chondrogenesis | Human adipose-derived mesenchymal stem cells | [108] |
| 25-hydroxycholesterol | ↓ Adipogenesis | Mouse pluripotent cell line (C3H10T1/2) | [109] |
| Cholest-4,6-Dien-3-One | Epithelial-to-mesenchymal transition | Human biliary tree stem/progenitor cells | [110] |
| 20(S)-hydroxycholesterol associated with simvastatin | ↑ Osteogenesis | Rats bone marrow mesenchymal stem cells (BNCC100381) | [111] |
| 27-Hydroxycholesterol | Differentiation and activation of monocytes into mature dendritic cell | Human monocytes cells from acute monocytic leukemia (THP-1) | [112] |
| ↑ Osteoclastogenesis | Mice macrophage cells (RAW264.7) | [113] | |
| ↓ Osteoblastogenesis | Mice primary calvarial (pre)osteoblast | [114] | |
| 24(S),25-Epoxycholesterol | Midbrain dopaminergic neurogenesis | Mice model | [115] |
| 22(S)-hydroxycholesterol and 20(S)-hydroxycholesterol combination | ↑ in vitro osteogenesis | Human periodontal ligament stem cells | [116] |
| ↑ in vivo alveolar bone regeneration | Rat model | [116] | |
| 7-ketocholesterol | ↓ Adipogenesis | Human adipose-derived stem cells | [117] |
| Oxysterol mixture * | Macrophage (M0) polarization to M2 immunomodulatory functional phenotype | Human macrophage | [118] |
| 22(R)-hydroxycholesterol and 20(S) hydroxycholesterol combination | ↑ Osteogenesis | Rats bone marrow mesenchymal stem cell | [119] |
| Mouse embryonic stem cells line (D3) | [120] | ||
| 7β,27-dihydroxycholesterol and 7α,27-dihydroxycholesterol | IL-17–producing CD4+ Th17 T cells | Murine and human primary CD4+ T cells | [121] |
| Oxy133 | ↑ Osteogenesis | Mouse multipotent bone marrow stromal cell (M2-10B4) | [122] |
| Mouse embryonic fibroblast cell (C3H10T1/2) | [122] | ||
| ↓ Adipogenesis | Mouse embryonic fibroblast cell (C3H10T1/2) | [122] | |
| Stimulates bone formation and spinal fusion | In vivo rat model | [122] | |
| Oxysterol 49 | ↑ Osteogenesis | Primary rabbit mesenchymal stem cells | [123] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Freitas, F.A.; Levy, D.; Zarrouk, A.; Lizard, G.; Bydlowski, S.P. Impact of Oxysterols on Cell Death, Proliferation, and Differentiation Induction: Current Status. Cells 2021, 10, 2301. https://doi.org/10.3390/cells10092301
de Freitas FA, Levy D, Zarrouk A, Lizard G, Bydlowski SP. Impact of Oxysterols on Cell Death, Proliferation, and Differentiation Induction: Current Status. Cells. 2021; 10(9):2301. https://doi.org/10.3390/cells10092301
Chicago/Turabian Stylede Freitas, Fábio Alessandro, Débora Levy, Amira Zarrouk, Gérard Lizard, and Sérgio Paulo Bydlowski. 2021. "Impact of Oxysterols on Cell Death, Proliferation, and Differentiation Induction: Current Status" Cells 10, no. 9: 2301. https://doi.org/10.3390/cells10092301
APA Stylede Freitas, F. A., Levy, D., Zarrouk, A., Lizard, G., & Bydlowski, S. P. (2021). Impact of Oxysterols on Cell Death, Proliferation, and Differentiation Induction: Current Status. Cells, 10(9), 2301. https://doi.org/10.3390/cells10092301