
Identification of GOLPH3 Partners in Drosophila Unveils Potential Novel Roles in Tumorigenesis and Neural Disorders


 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Stocks and Transgenes
2.2. Molecular Cloning
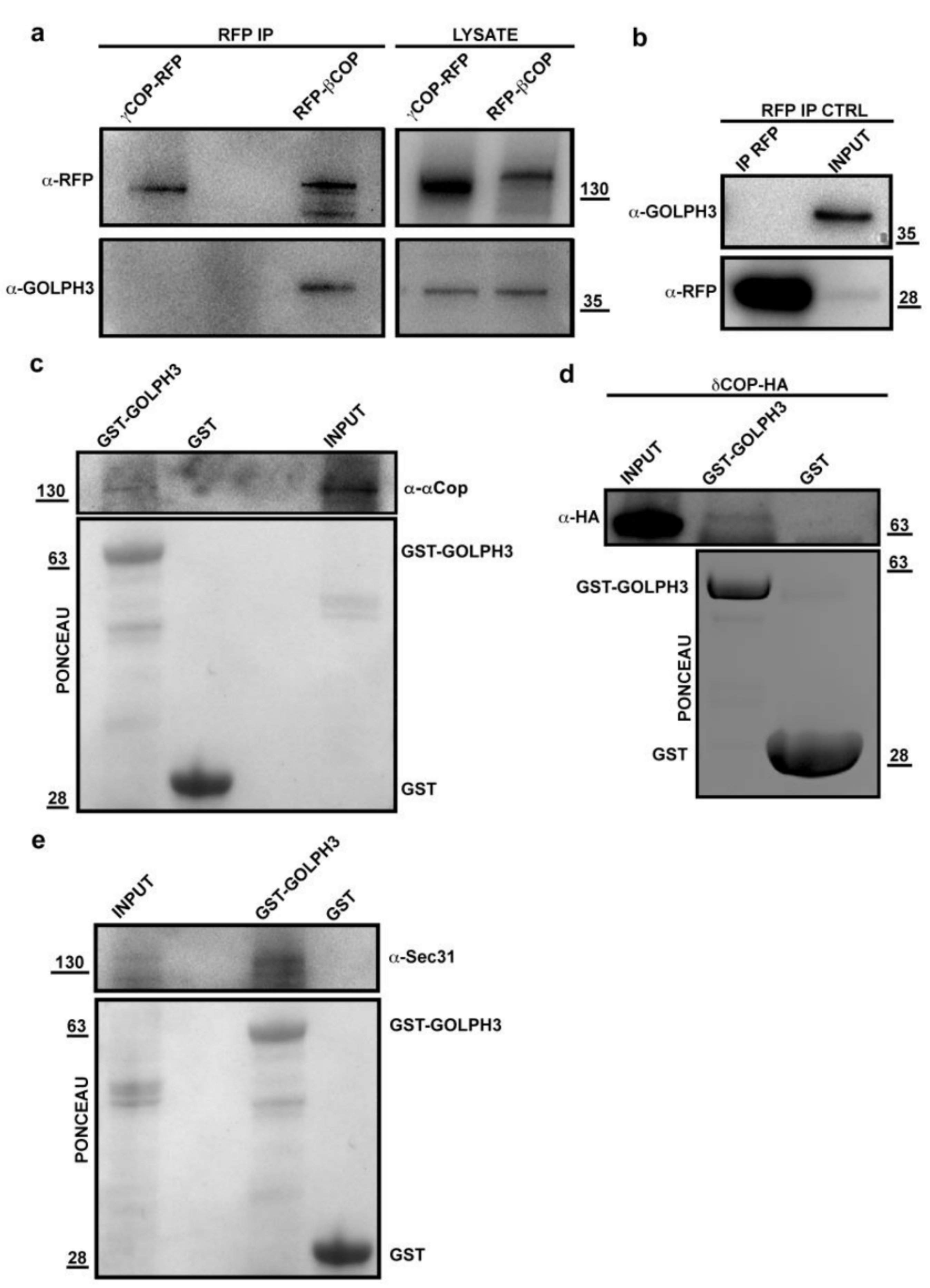
2.3. Co-Immunoprecipitation Experiments
2.4. Western Blotting
2.5. Glutathione S-Transferase (GST) Pull-Down Assays
2.6. Proteomics and Data Analysis
2.7. Computational Analysis of the dGOLPH3 Interactome
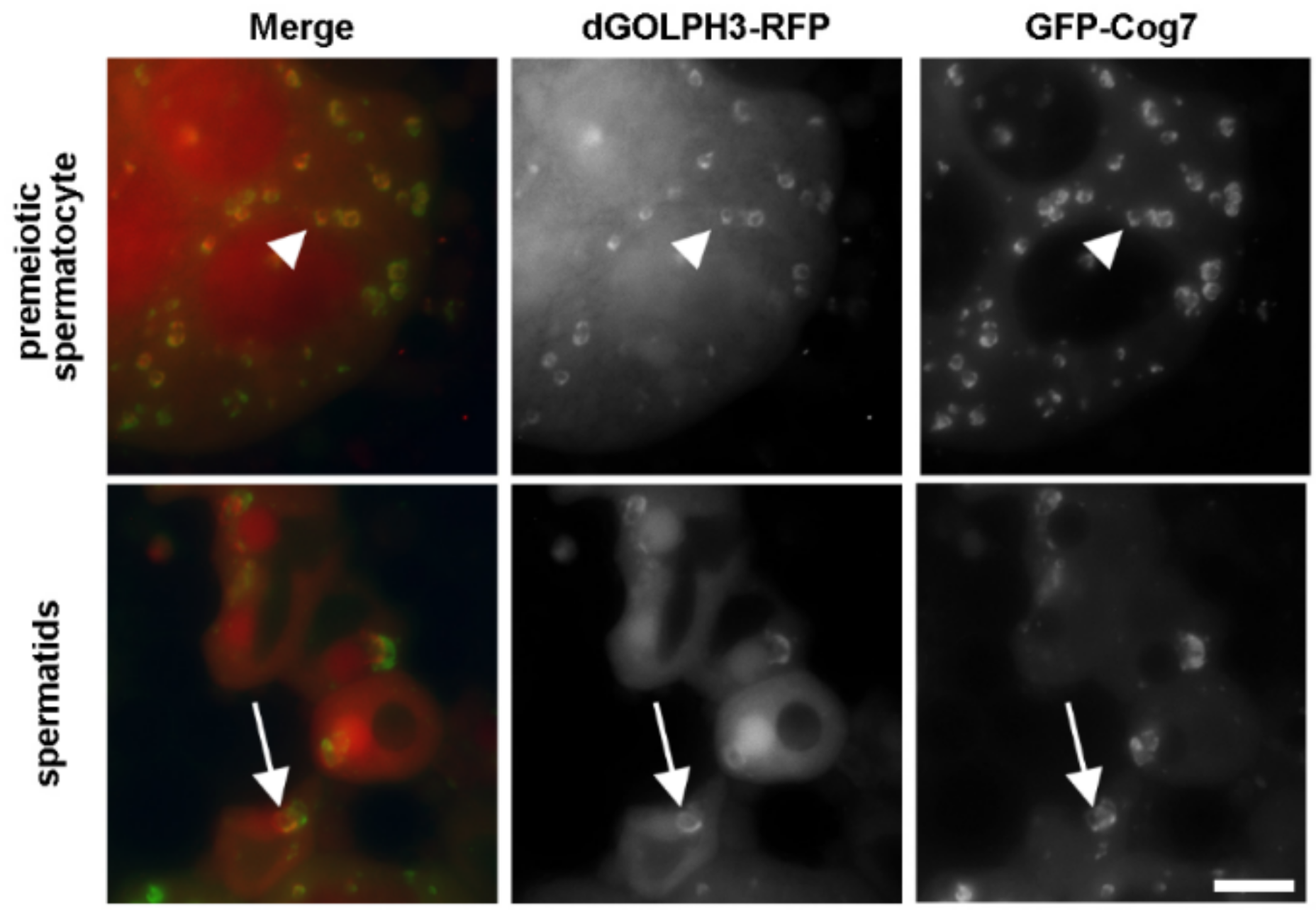
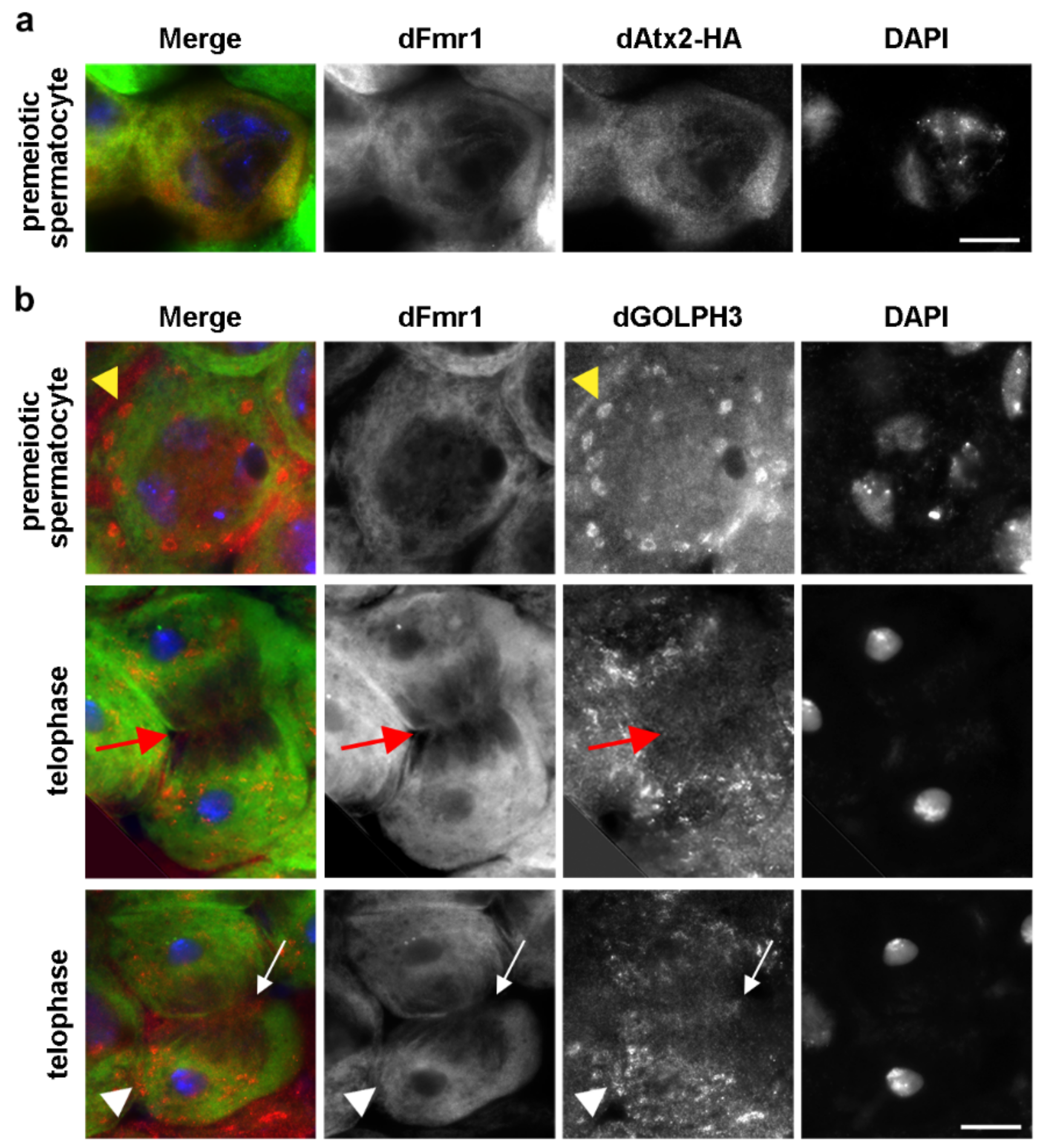
2.8. Immunofluorescence Analysis and Live Imaging of Testes
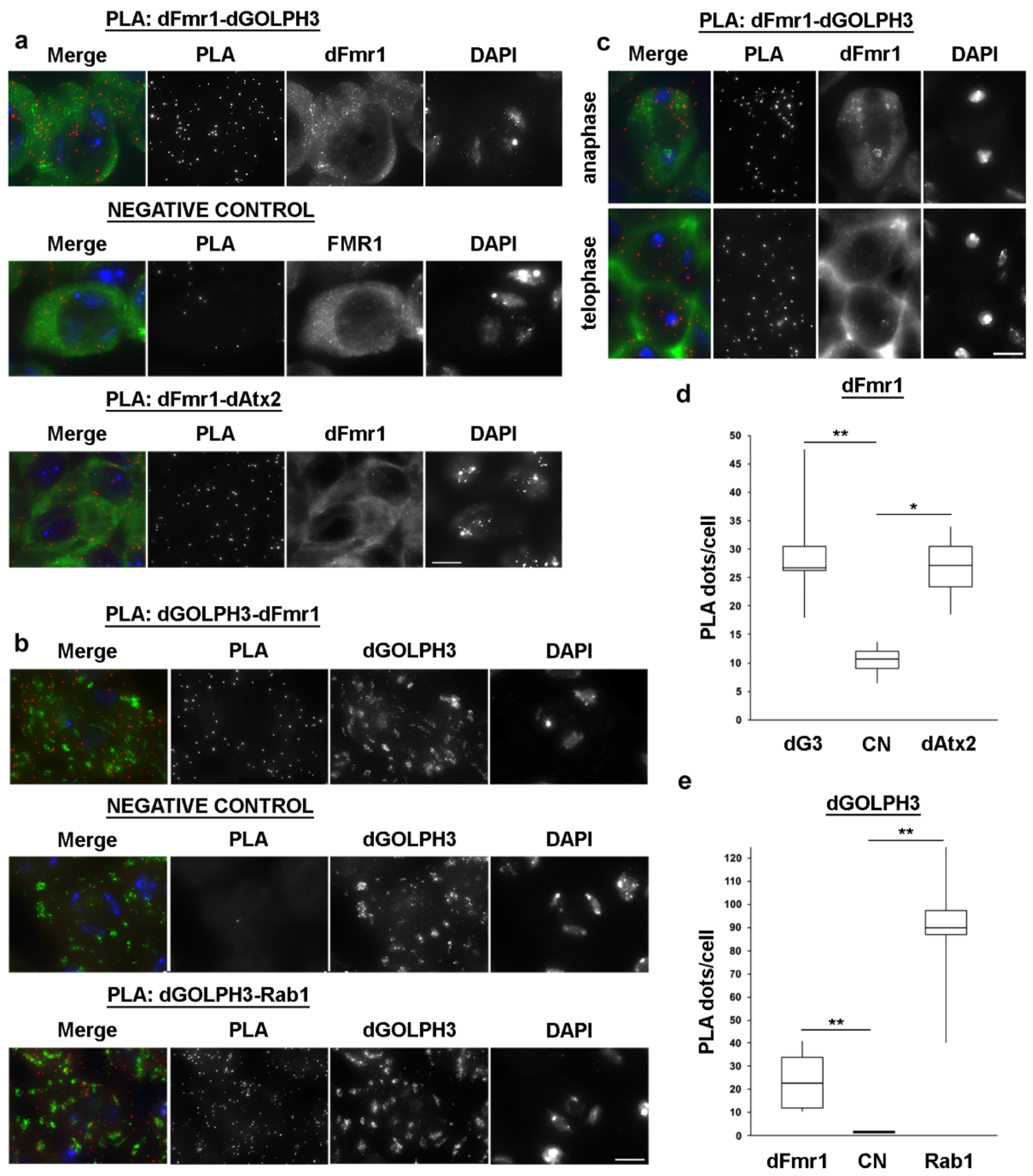
2.9. Proximity Ligation Assay
3. Results
3.1. Identification of the dGOLPH3 Interactome in Drosophila melanogaster
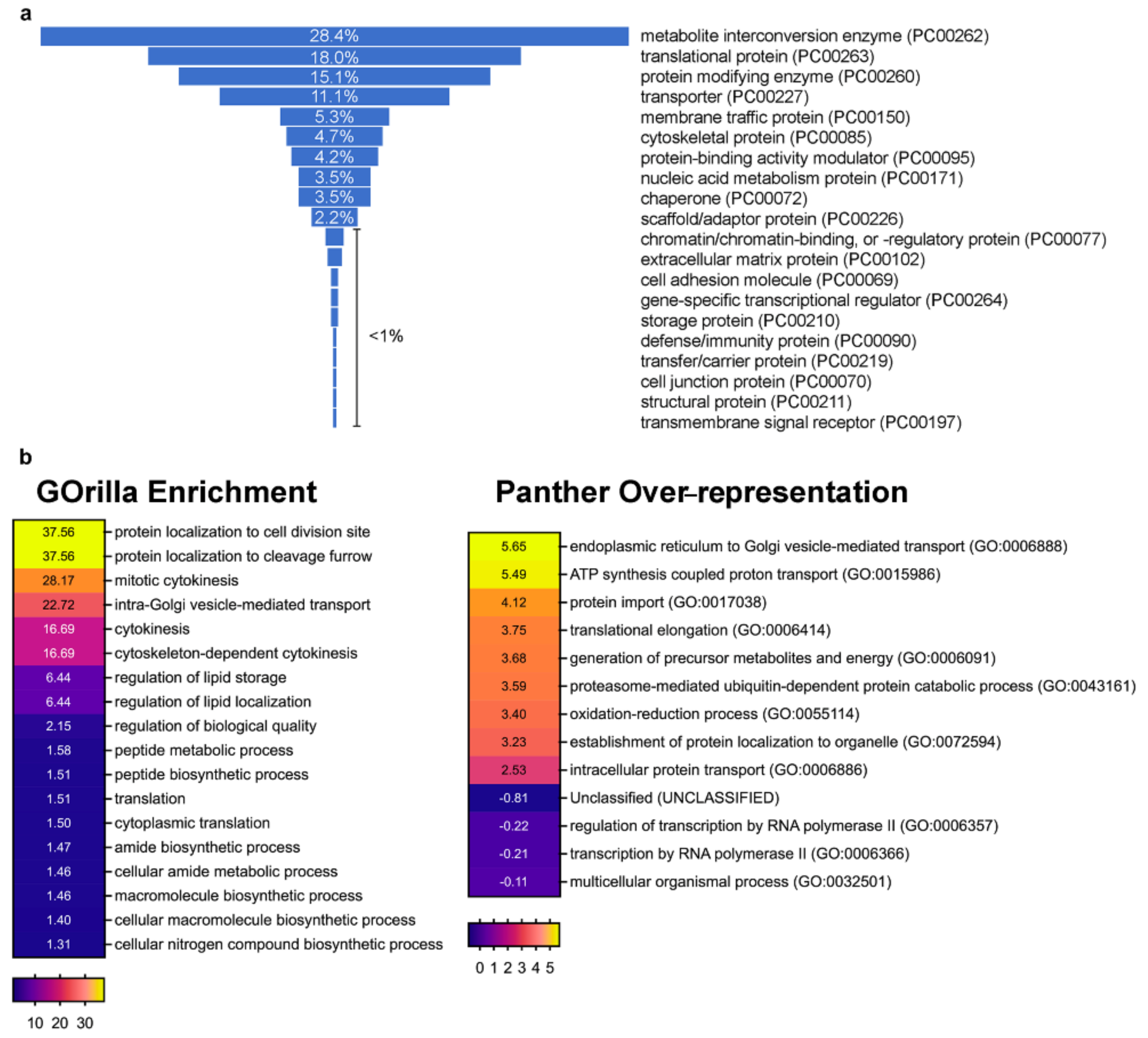
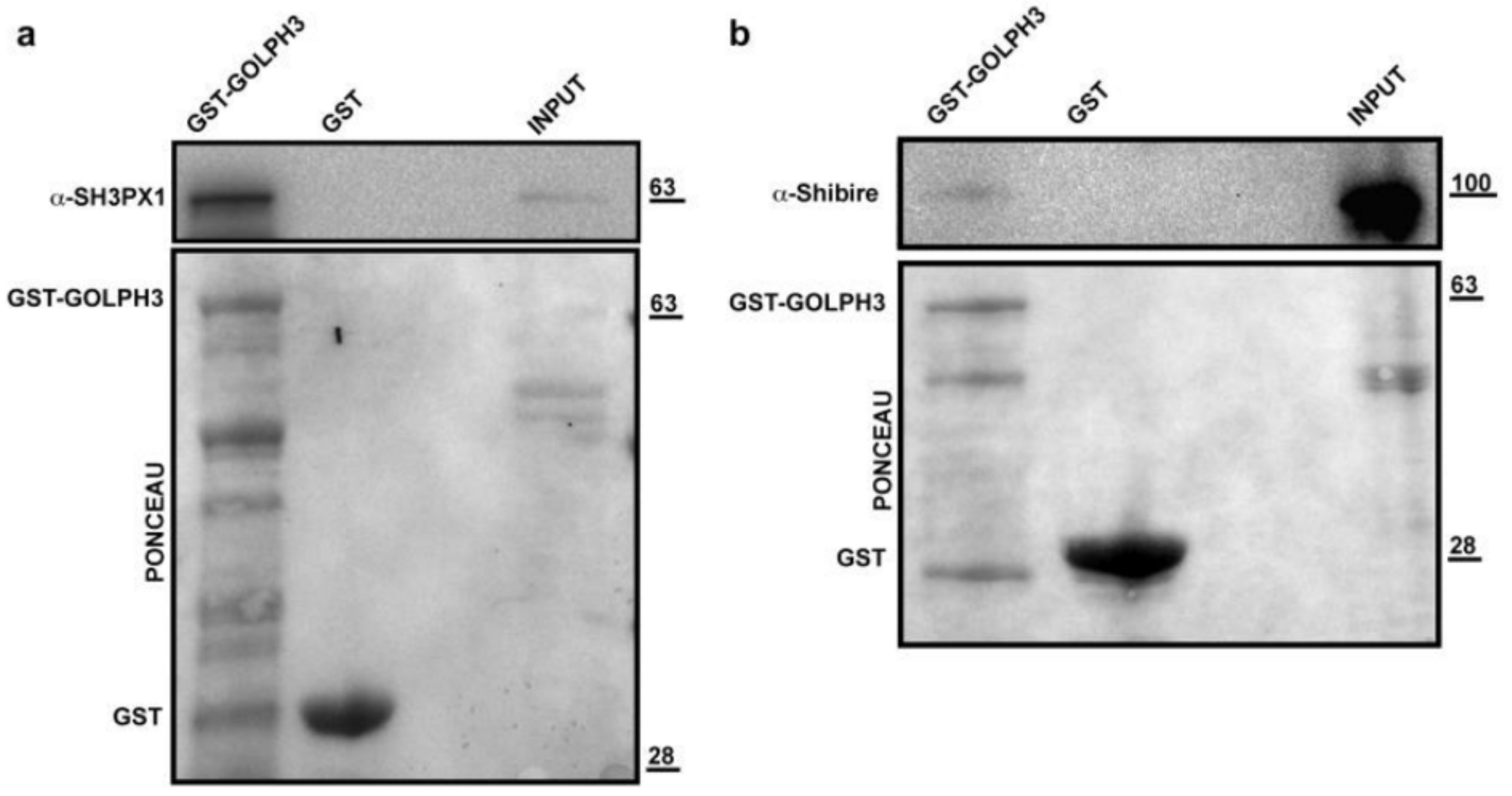
3.2. Profiling dGOLPH3 Interactors Reveals an Enrichment of Membrane Trafficking Proteins
3.3. The dGOLPH3 Interactome Reveals Functions in Several Glycosylation Pathways
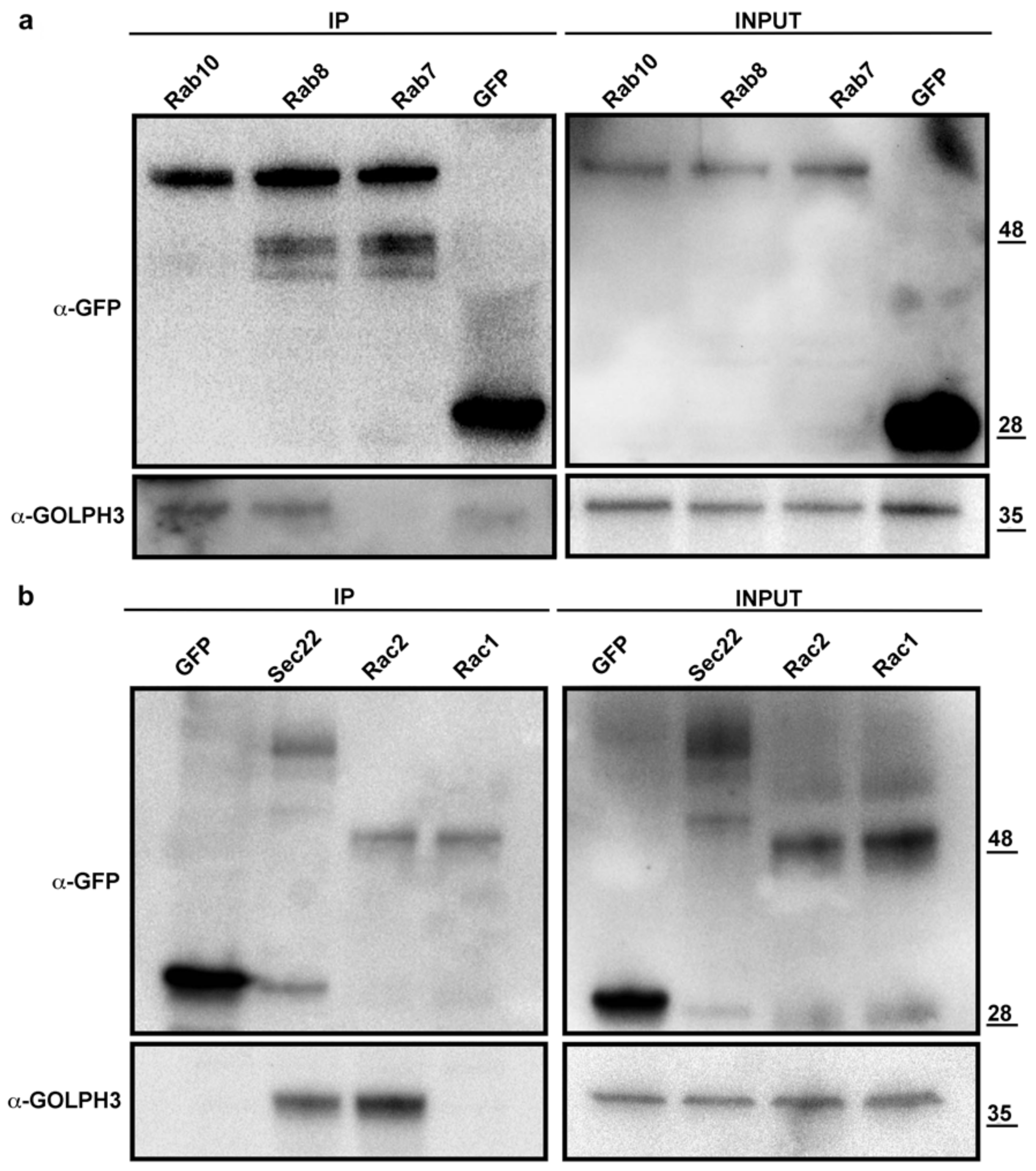
3.4. dGOLPH3 Partners Control Lipid Homeostasis and Golgi Architecture
3.5. dGOLPH3 Partners Regulate Cell Cycle Progression and Cell Signaling
3.6. dGOLPH3 Interactors Suggest an Involvement in the Assembly or Organization of Ciliary and Flagellar Axonemes
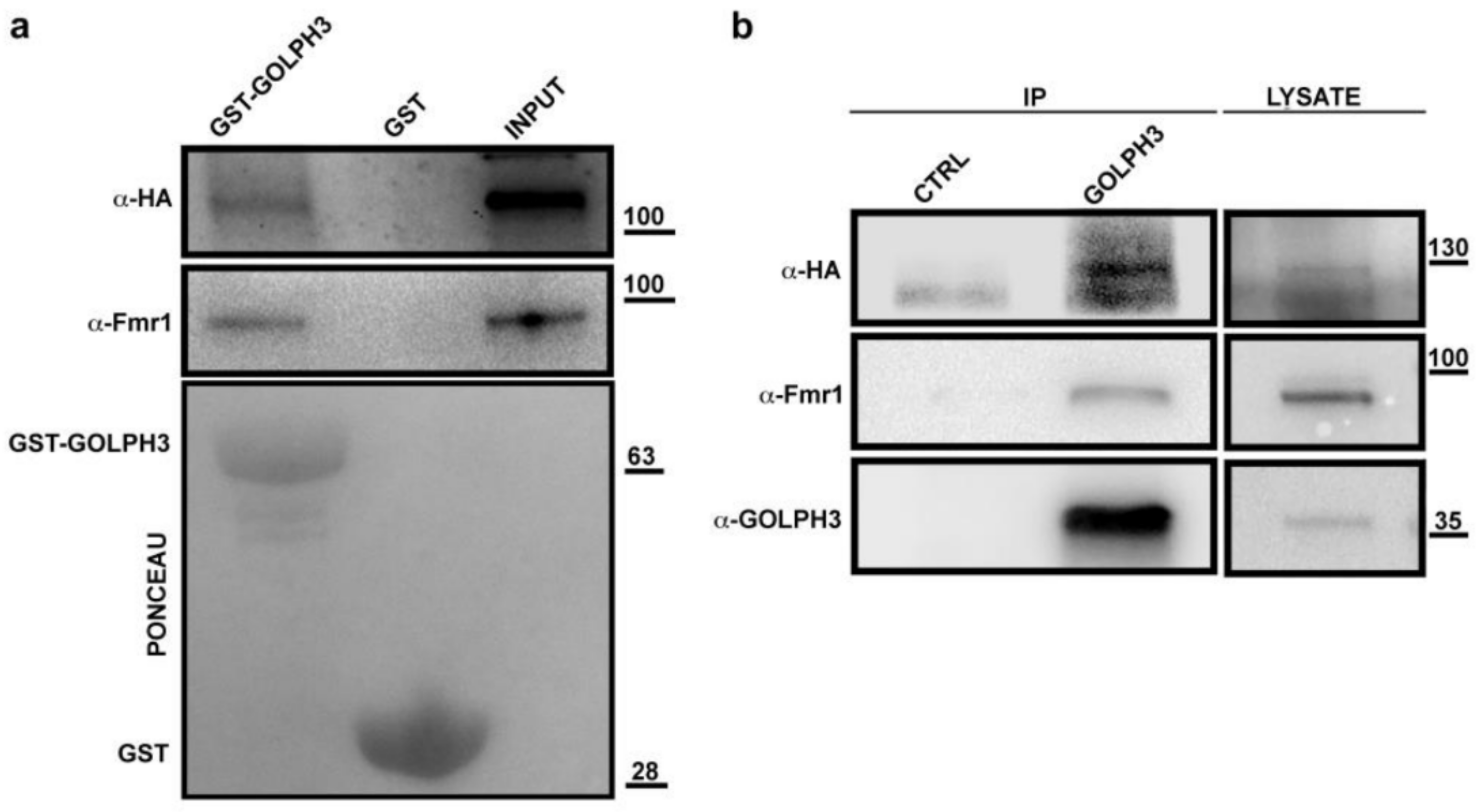
3.7. dGOLPH3 Interacts with Drosophila Ataxin-2 and Fragile Mental Retardation Protein
4. Discussion
4.1. The dGOLPH3 Interactome Reveals an Enrichment in Vesicle-Mediated Trafficking and Cytokinesis Proteins
4.2. dGOLPH3 Interacts with Proteins Required for Protein Glycosylation and Lipid Homeostasis
4.3. The dGOLPH3 Interactome Indicates Molecular Targets that Might Be Relevant for Therapeutic Intervention in Cancer and Other Neurological Diseases
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dippold, H.C.; Ng, M.M.; Farber-Katz, S.E.; Lee, S.K.; Kerr, M.L.; Peterman, M.C.; Sim, R.; Wiharto, P.A.; Galbraith, K.A.; Madhavarapu, S.; et al. GOLPH3 bridges phosphatidylinositol-4- phosphate and actomyosin to stretch and shape the Golgi to promote budding. Cell 2009, 139, 337–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, P.; Bhave, M.; Jain, B.K.; RoyChowdhury, S.; Bhattacharyya, D. Vps74p controls Golgi size in an Arf1-dependent manner. FEBS Lett. 2018, 592, 3720–3735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahajeng, J.; Kuna, R.S.; Makowski, S.L.; Tran, T.T.T.; Buschman, M.D.; Li, S.; Cheng, N.; Ng, M.M.; Field, S.J. Efficient Golgi Forward Trafficking Requires GOLPH3-Driven, PI4P-Dependent Membrane Curvature. Dev. Cell 2019, 50, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Sechi, S.; Colotti, G.; Belloni, G.; Mattei, V.; Frappaolo, A.; Giansanti, M.G. GOLPH3 Is Essential for Contractile Ring Formation and Rab11 Localization to the Cleavage Site during Cytokinesis in Drosophila melanogaster. PLoS Genet. 2014, 10, e1004305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sechi, S.; Frappaolo, A.; Karimpour-Ghahnavieh, A.; Piergentili, R.; Giansanti, M.G. Oncogenic Roles of GOLPH3 in the Physiopathology of Cancer. Int. J. Mol. Sci. 2020, 21, 933. [Google Scholar] [CrossRef] [Green Version]
- Snyder, C.M.; Mardones, G.A.; Ladinsky, M.S.; Howell, K.E. GMx33 associates with the trans-Golgi matrix in a dynamic manner and sorts within tubules exiting the Golgi. Mol. Biol. Cell. 2006, 17, 511–524. [Google Scholar] [CrossRef]
- Eckert, E.S.P.; Reckmann, I.; Hellwig, A.; Röhling, S.; El-Battari, A.; Wieland, F.T.; Popoff, V. Golgi phosphoprotein 3 triggers signal-mediated incorporation of glycosyltransferases into coatomer-coated (COPI) vesicles. J. Biol. Chem. 2014, 289, 31319–31329. [Google Scholar] [CrossRef] [Green Version]
- Isaji, T.; Im, S.; Gu, W.; Wang, Y.; Hang, Q.; Lu, J.; Fukuda, T.; Hashii, N.; Takakura, D.; Kawasaki, N.; et al. An oncogenic protein Golgi phosphoprotein 3 up-regulates cell migration via sialylation. J. Biol. Chem. 2014, 289, 20694–20705. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Doray, B.; Kornfeld, S. Recycling of Golgi glycosyltransferases requires direct binding to coatomer. Proc. Natl. Acad. Sci. USA 2018, 115, 8984–8989. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, R.; Russo, D.; Kurokawa, K.; Sahu, P.; Lombardi, B.; Supino, D.; Zhukovsky, M.A.; Vocat, A.; Pothukuchi, P.; Kunnathully, V.; et al. Golgi maturation-dependent glycoenzyme recycling controls glycosphingolipid biosynthesis and cell growth via GOLPH3. EMBO J. 2021, 40, e107238. [Google Scholar] [CrossRef]
- Schmitz, K.R.; Liu, J.; Li, S.; Setty, T.G.; Wood, C.S.; Burd, C.G.; Ferguson, K.M. Golgi localization of glycosyltransferases requires a Vps74p oligomer. Dev. Cell 2008, 14, 523–534. [Google Scholar] [CrossRef] [Green Version]
- Tu, L.; Tai, W.C.; Chen, L.; Banfield, D.K. Signal-mediated dynamic retention of glycosyltransferases in the Golgi. Science 2008, 321, 404–407. [Google Scholar] [CrossRef]
- Wood, C.S.; Schmitz, K.R.; Bessman, N.J.; Setty, T.G.; Ferguson, K.M.; Burd, C.G. PtdIns4P recognition by Vps74/GOLPH3 links PtdIns 4-kinase signaling to retrograde Golgi trafficking. J. Cell Biol. 2009, 187, 967–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, L.; Banfield, D.K. Localization of Golgi-resident glycosyltransferases. Cell Mol. Life Sci. 2010, 67, 29–41. [Google Scholar] [CrossRef]
- Ali, M.F.; Chachadi, V.B.; Petrosyan, A.; Cheng, P.W. Golgi phosphoprotein 3 determines cell binding properties under dynamic flow by controlling Golgi localization of core 2 N-acetylglucosaminyltransferase 1. J. Biol. Chem. 2012, 287, 39564–39577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.L.; Chang, C.W.; Chang, Y.Y.; Sung, H.H.; Lin, M.D.; Chang, S.C.; Chen, C.H.; Huang, C.W.; Tung, K.S.; Chou, T.B. The Drosophila GOLPH3 homolog regulates the biosynthesis of heparan sulfate proteoglycans by modulating the retrograde trafficking of exostosins. Development 2013, 140, 2798–2807. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.L.; Kabbarah, O.; Liang, M.-C.; Ivanova, E.; Anagnostou, V.; Wu, J.; Dhakal, S.; Wu, M.; Chen, S.; Feinberg, T.; et al. GOLPH3 modulates mTOR signalling and rapamycin sensitivity in cancer. Nature 2009, 459, 1085–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokuda, E.; Itoh, T.; Hasegawa, J.; Ijuin, T.; Takeuchi, Y.; Irino, Y.; Fukumoto, M.; Takenawa, T. Phosphatidylinositol 4-phosphate in the Golgi apparatus regulates cell-cell adhesion and invasive cell migration in human breast cancer. Cancer Res. 2014, 74, 3054–3066. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Ding, Z.; Mo, J.; Sang, B.; Shi, Q.; Hu, J.; Xie, S.; Zhan, W.; Lu, D.; Yang, M.; et al. GOLPH3 promotes glioblastoma cell migration and invasion via the mTOR-YB1 pathway in vitro. Mol. Carcinog. 2015, 54, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xu, T.; Qin, R.; Yan, Y.; Chen, C.; Chen, Y.; Yu, H.; Xia, C.; Lu, Y.; Ding, X.; et al. Overexpression of Golgi phosphoprotein-3 (GOLPH3) in glioblastoma multiforme is associated with worse prognosis. J. Neurooncol. 2012, 110, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Sechi, S.; Frappaolo, A.; Belloni, G.; Giansanti, M.G. The roles of the oncoprotein GOLPH3 in contractile ring assembly and membrane trafficking during cytokinesis. Biochem. Soc. Trans. 2015, 43, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Sechi, S.; Frappaolo, A.; Karimpour-Ghahnavieh, A.; Fraschini, R.; Giansanti, M.G. A novel coordinated function of Myosin II with GOLPH3 controls centralspindlin localization during cytokinesis in Drosophila. J. Cell Sci. 2020, 133, jcs252965. [Google Scholar] [CrossRef]
- Demarco, R.S.; Eikenes, Å.H.; Haglund, K.; Jones, D.L. Investigating spermatogenesis in Drosophila melanogaster. Methods 2014, 68, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Fabian, L.; Brill, J.A. Drosophila spermiogenesis: Big things come from little packages. Spermatogenesis 2012, 2, 197–212. [Google Scholar] [CrossRef] [Green Version]
- Frappaolo, A.; Sechi, S.; Belloni, G.; Piergentili, R.; Giansanti, M.G. Visualization of cleavage furrow proteins in fixed dividing spermatocytes. Methods Cell Biol. 2017, 137, 85–103. [Google Scholar]
- Giansanti, M.G.; Fuller, M.T. What Drosophila spermatocytes tell us about the mechanisms underlying cytokinesis. Cytoskeleton 2012, 69, 869–881. [Google Scholar] [CrossRef] [Green Version]
- Sechi, S.; Frappaolo, A.; Fraschini, R.; Capalbo, L.; Gottardo, M.; Belloni, G.; Glover, D.M.; Wainman, A.; Giansanti, M.G. Rab1 interacts with GOLPH3 and controls Golgi structure and contractile ring constriction during cytokinesis in Drosophila melanogaster. Open Biol. 2017, 7, 160257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Schulze, K.L.; Hiesinger, P.R.; Suyama, K.; Wang, S.; Fish, M.; Acar, M.; Hoskins, R.A.; Bellen, H.J.; Scott, M.P. Thirty-one flavors of Drosophila Rab Proteins. Genetics 2007, 176, 1307–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarov, M.; Barz, C.; Jambor, H.; Hein, M.Y.; Schmied, C.; Suchold, D.; Stender, B.; Janosch, S.; Kj, V.V.; Krishnan, R.T.; et al. A genome-wide resource for the analysis of protein localisation in Drosophila. eLife 2016, 5, e12068. [Google Scholar] [CrossRef]
- Bischof, J.; Björklund, M.; Furger, E.; Schertel, C.; Taipale, J.; Basler, K. A versatile platform for creating a comprehensive UAS-ORFeome library in Drosophila. Development 2013, 140, 2434–2442. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; McKearin, D. Dpp signaling silences bam transcription directly to establish asymmetric divisions of germline stem cells. Curr. Biol. 2003, 13, 1786–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sechi, S.; Frappaolo, A.; Karimpour-Ghahnavieh, A.; Gottardo, M.; Burla, R.; Di Francesco, L.; Szafer-Glusman, E.; Schininà, E.; Fuller, M.T.; Saggio, I.; et al. Drosophila Doublefault protein coordinates multiple events during male meiosis by controlling mRNA translation. Development. 2019, 146, dev183053. [Google Scholar] [CrossRef]
- Belloni, G.; Sechi, S.; Riparbelli, M.G.; Fuller, M.T.; Callaini, G.; Giansanti, M.G. Mutations in Cog7 affect Golgi structure, meiotic cytokinesis and sperm development during Drosophila spermatogenesis. J. Cell Sci. 2012, 125, 5441–5542. [Google Scholar] [PubMed] [Green Version]
- Kitazawa, D.; Yamaguchi, M.; Mori, H.; Inoue, Y.H. COPI-mediated membrane trafficking is required for cytokinesis in Drosophila male meiotic divisions. J. Cell Sci. 2012, 125, 3649–3660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shugrue, C.A.; Kolen, E.R.; Peters, H.; Czernik, A.; Kaiser, C.; Matovcik, L.; Hubbard, A.L.; Gorelick, F. Identification of the putative mammalian orthologue of Sec31P, a component of the COPII coat. J. Cell Sci. 1999, 112, 4547–4556. [Google Scholar] [CrossRef]
- Hicks, L.; Liu, G.; Ukken, F.P.; Lu, S.; Bollinger, K.E.; O’Connor-Giles, K.; Gonsalvez, G.B. Depletion or over-expression of Sh3px1 results in dramatic changes in cell morphology. Biol. Open 2015, 4, 1448–1461. [Google Scholar] [CrossRef] [Green Version]
- Giansanti, M.G.; Vanderleest, T.E.; Jewett, C.E.; Sechi, S.; Frappaolo, A.; Fabian, L.; Robinett, C.C.; Brill, J.A.; Loerke, D.; Fuller, M.T.; et al. Exocyst-Dependent Membrane Addition Is Required for Anaphase Cell Elongation and Cytokinesis in Drosophila. PLoS Genet. 2015, 11, e1005632. [Google Scholar] [CrossRef] [Green Version]
- Frappaolo, A.; Sechi, S.; Kumagai, T.; Robinson, S.; Fraschini, R.; Karimpour-Ghahnavieh, A.; Belloni, G.; Piergentili, R.; Tiemeyer, K.H.; Tiemeyer, M.; et al. COG7 deficiency in Drosophila generates multifaceted developmental, behavioral and protein glycosylation phenotypes. J. Cell Sci. 2017, 130, 3637–3649. [Google Scholar]
- Di Francesco, L.; Correani, V.; Fabrizi, C.; Fumagalli, L.; Mazzanti, M.; Maras, B.; Schininà, M.E. 14-3-3ε marks the amyloid-stimulated microglia long-term activation. Proteomics 2012, 12, 124–134. [Google Scholar] [CrossRef]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, D183–D189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eden, E.; Navon, R.; Steinfeld, I.; Lipson, D.; Yakhini, Z. GOrilla: A tool for discovery and visualization of enriched GO terms in ranked gene lists. Bioinformatics 2009, 10, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, L.; Chen, L.; Banfield, D.K. A conserved N-terminal arginine-motif in GOLPH3-family proteins mediates binding to coatomer. Traffic 2012, 13, 1496–1507. [Google Scholar] [CrossRef]
- Wood, C.S.; Hung, C.S.; Huoh, Y.S.; Mousley, C.J.; Stefan, C.J.; Bankaitis, V.; Ferguson, K.M.; Burd, C.G. Local control of phosphatidylinositol 4-phosphate signaling in the Golgi apparatus by Vps74 and Sac1 phosphoinositide phosphatase. Mol. Biol. Cell 2012, 23, 2527–2536. [Google Scholar] [CrossRef] [PubMed]
- Mima, J. Reconstitution of membrane tethering mediated by Rab-family small GTPases. Biophys Rev. 2018, 10, 543–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavieres, V.A.; Cerda-Troncoso, C.; Rivera-Dictter, A.; Castro, R.I.; Luchsinger, C.; Santibañez, N.; Burgos, P.V.; Mardones, G.A. Human Golgi phosphoprotein 3 is an effector of RAB1A and RAB1B. PLoS ONE 2020, 15, e0237514. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Zou, S.; Chen, Y.; Lipatova, Z.; Sun, D.; Zhu, X.; Li, R.; Wu, Z.; You, W.; Cong, X.; et al. A Rab5 GTPase module is important for autophagosome closure. PLoS Genet. 2017, 13, e1007020. [Google Scholar] [CrossRef] [Green Version]
- Hutagalung, A.H.; Novick, P.J. Role of Rab GTPases in membrane traffic and cell physiology. Physiol Rev. 2011, 91, 119–149. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Plesken, H.; Treisman, J.E.; Edelman-Novemsky, I.; Ren, M. Lightoid and Claret: A rab GTPase and its putative guanine nucleotide exchange factor in biogenesis of Drosophila eye pigment granules. Proc. Natl. Acad. Sci. USA 2004, 101, 11652–11657. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, Z.; Huang, X. Rab32 is important for autophagy and lipid storage in Drosophila. PLoS ONE 2012, 7, e32086. [Google Scholar] [CrossRef]
- Adnan, M.; Islam, W.; Zhang, J.; Zheng, W.; Lu, G.D. Diverse Role of SNARE Protein Sec22 in Vesicle Trafficking, Membrane Fusion, and Autophagy. Cells 2019, 8, 337. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Holthuis, J.C.M. Membrane contact sites, ancient and central hubs of cellular lipid logistics. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1450–1458. [Google Scholar] [CrossRef] [PubMed]
- Halachmi, N.; Lev, Z. The Sec1 family: A novel family of proteins involved in synaptic transmission and general secretion. J. Neurochem. 1996, 66, 889–897. [Google Scholar] [CrossRef]
- Shen, J.; Tareste, D.C.; Paumet, F.; Rothman, J.E.; Melia, T.J. Selective activation of cognate SNAREpins by Sec1/Munc18 proteins. Cell 2007, 128, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Südhof, T.C.; Rothman, J.E. Membrane fusion: Grappling with SNARE and SM proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Dulubova, I.; Min, S.W.; Chen, X.; Rizo, J.; Südhof, T.C. Sly1 binds to Golgi and ER syntaxins via a conserved N-terminal peptide motif. Dev. Cell 2002, 2, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Akopian, D.; Shen, K.; Zhang, X.; Shan, S.O. Signal recognition particle: An essential protein-targeting machine. Annu. Rev. Biochem. 2013, 82, 693–721. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zhu, X.; Zhou, L.; Wang, J.; Tao, X.; Wang, S.; Sun, F.; Kan, X.; Han, Z.; Gu, Y. Gilgamesh is required for the maintenance of germline stem cells in Drosophila testis. Sci. Rep. 2017, 7, 5737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nerusheva, O.O.; Dorogova, N.V.; Gubanova, N.V.; Yudina, O.S.; Omelyanchuk, L.V. A GFP trap study uncovers the functions of Gilgamesh protein kinase in Drosophila melanogaster spermatogenesis. Cell Biol. Int. 2009, 33, 586–593. [Google Scholar] [CrossRef]
- Gault, W.J.; Olguin, P.; Weber, U.; Mlodzik, M. Drosophila CK1-γ, gilgamesh, controls PCP-mediated morphogenesis through regulation of vesicle trafficking. J. Cell Biol. 2012, 196, 605–621. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, K.; Yamamoto-Hino, M.; Matsuyama, N.; Suzuki, E.; Goto, S. Subunits of the GPI transamidase complex localize to the endoplasmic reticulum and nuclear envelope in Drosophila. FEBS Lett. 2021, 595, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.; Dudek, J.; Gogala, M.; Schorr, S.; Linxweiler, J.; Lang, S.; Becker, T.; Beckmann, R.; Zimmermann, R.; Förster, F. Structure of the mammalian oligosaccharyl-transferase complex in the native ER protein translocon. Nat. Commun. 2014, 5, 3072. [Google Scholar] [CrossRef] [Green Version]
- Hebert, D.N.; Molinari, M. Flagging and docking: Dual roles for N-glycans in protein quality control and cellular proteostasis. Trends Biochem Sci. 2012, 37, 404–410. [Google Scholar] [CrossRef]
- Määttänen, P.; Gehring, K.; Bergeron, J.J.; Thomas, D.Y. Protein quality control in the ER: The recognition of misfolded proteins. Semin. Cell Dev. Biol. 2010, 21, 500–511. [Google Scholar] [CrossRef]
- Parkinson, W.M.; Dookwah, M.; Dear, M.L.; Gatto, C.L.; Aoki, K.; Tiemeyer, M.; Broadie, K. Synaptic roles for phosphomannomutase type 2 in a new Drosophila congenital disorder of glycosylation disease model. Dis. Model Mech. 2016, 9, 513–527. [Google Scholar] [PubMed] [Green Version]
- Rind, N.; Schmeiser, V.; Thiel, C.; Absmanner, B.; Lübbehusen, J.; Hocks, J.; Apeshiotis, N.; Wilichowski, E.; Lehle, L.; Körner, C. A severe human metabolic disease caused by deficiency of the endoplasmatic mannosyltransferase hALG11 leads to congenital disorder of glycosylation-Ip. Hum. Mol. Genet. 2010, 19, 1413–1424. [Google Scholar] [CrossRef]
- Andreotti, G.; Cabeza de Vaca, I.; Poziello, A.; Monti, M.C.; Guallar, V.; Cubellis, M.V. Conformational response to ligand binding in phosphomannomutase2: Insights into inborn glycosylation disorder. J. Biol. Chem. 2014, 289, 34900–34910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeze, H.H.; Chong, J.X.; Bamshad, M.J.; Ng, B.G. Solving glycosylation disorders: Fundamental approaches reveal complicated pathways. Am. J. Hum. Genet. 2014, 94, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Freeze, H.H.; Eklund, E.A.; Ng, B.G.; Patterson, M.C. Neurological aspects of human glycosylation disorders. Annu. Rev. Neurosci. 2015, 38, 105–125. [Google Scholar] [CrossRef] [Green Version]
- Schollen, E.; Pardon, E.; Heykants, L.; Renard, J.; Doggett, N.A.; Callen, D.F.; Cassiman, J.J.; Matthijs, G. Comparative analysis of the phosphomannomutase genes PMM1, PMM2 and PMM2psi: The sequence variation in the processed pseudogene is a reflection of the mutations found in the functional gene. Hum. Mol. Genet. 1998, 7, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Tran, D.T.; Zhang, L.; Zhang, Y.; Tian, E.; Earl, L.A.; Ten Hagen, K.G. Multiple members of the UDP-GalNAc: Polypeptide N-acetylgalactosaminyltransferase family are essential for viability in Drosophila. J. Biol. Chem. 2012, 287, 5243–5252. [Google Scholar] [CrossRef] [Green Version]
- Thackeray, J.R.; Gaines, P.C.; Ebert, P.; Carlson, J.R. Small wing encodes a phospholipase C-(gamma) that acts as a negative regulator of R7 development in Drosophila. Development 1998, 125, 5033–5042. [Google Scholar] [CrossRef] [PubMed]
- McElwain, M.A.; Ko, D.C.; Gordon, M.D.; Fyrst, H.; Saba, J.D.; Nusse, R. A suppressor/enhancer screen in Drosophila reveals a role for wnt-mediated lipid metabolism in primordial germ cell migration. PLoS ONE 2011, 6, e26993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, A.; Aoki, J. Phospholipase A1: Structure, distribution and function. Future Lipidol. 2006, 1, 687–700. [Google Scholar] [CrossRef]
- Sheng, X.; Yung, Y.C.; Chen, A.; Chun, J. Lysophosphatidic acid signalling in development. Development 2015, 142, 1390–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gáliková, M.; Klepsatel, P.; Münch, J.; Kühnlein, R.P. Spastic paraplegia-linked phospholipase PAPLA1 is necessary for development, reproduction, and energy metabolism in Drosophila. Sci. Rep. 2017, 7, 46516. [Google Scholar] [CrossRef]
- Kunduri, G.; Yuan, C.; Parthibane, V.; Nyswaner, K.M.; Kanwar, R.; Nagashima, K.; Britt, S.G.; Mehta, N.; Kotu, V.; Porterfield, M.; et al. Phosphatidic acid phospholipase A1 mediates ER-Golgi transit of a family of G protein-coupled receptors. J. Cell Biol. 2014, 206, 79–95. [Google Scholar] [CrossRef] [Green Version]
- Richmond, G.S.; Smith, T.K. Phospholipases A1. Int. J. Mol. Sci. 2011, 12, 588–612. [Google Scholar] [CrossRef] [Green Version]
- Oh, C.S.; Toke, D.A.; Mandala, S.; Martin, C.E. ELO2 and ELO3, homologues of the Saccharomyces cerevisiae ELO1 gene, function in fatty acid elongation and are required for sphingolipid formation. J. Biol. Chem. 1997, 272, 17376–17384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szafer-Glusman, E.; Giansanti, M.G.; Nishihama, R.; Bolival, B.; Pringle, J.; Gatti, M.; Fuller, M.T. A role for very-long-chain fatty acids in furrow ingression during cytokinesis in Drosophila spermatocytes. Curr. Biol. 2008, 18, 1426–1431. [Google Scholar] [CrossRef] [Green Version]
- Ben-David, G.; Miller, E.; Steinhauer, J. Drosophila spermatid individualization is sensitive to temperature and fatty acid metabolism. Spermatogenesis 2015, 5, e1006089. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Liu, Z.; Huang, X. Drosophila models of peroxisomal biogenesis disorder: Peroxins are required for spermatogenesis and very-long-chain fatty acid metabolism. Hum. Mol. Genet. 2010, 19, 494–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herr, D.R.; Fyrst, H.; Phan, V.; Heinecke, K.; Georges, R.; Harris, G.L.; Saba, J.D. Sply regulation of sphingolipid signaling molecules is essential for Drosophila development. Development 2003, 130, 2443–2453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolz, A.; Hilt, W.; Buchberger, A.; Wolf, D.H. Cdc48: A power machine in protein degradation. Trends Biochem. Sci. 2011, 36, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Appenzeller-Herzog, C.; Nyfeler, B.; Burkhard, P.; Santamaria, I.; Lopez-Otin, C.; Hauri, H.P. Carbohydrate- and conformation-dependent cargo capture for ER-exit. Mol. Biol. Cell. 2005, 16, 1258–1267. [Google Scholar] [CrossRef] [Green Version]
- Mitrovic, S.; Ben-Tekaya, H.; Koegler, E.; Gruenberg, J.; Hauri, H.P. The cargo receptors Surf4, endoplasmic reticulum-Golgi intermediate compartment (ERGIC)-53, and p25 are required to maintain the architecture of ERGIC and Golgi. Mol. Biol. Cell. 2008, 19, 1976–1990. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.C.; Zhou, Y.; Yang, C.Z.; Xiong, D.S. A review of ERGIC-53: Its structure, functions, regulation and relations with diseases. Histol. Histopathol. 2009, 24, 1193–1204. [Google Scholar]
- Hauri, H.P.; Kappeler, F.; Andersson, H.; Appenzeller, C. ERGIC-53 and traffic in the secretory pathway. J. Cell Sci. 2000, 113, 587–596. [Google Scholar] [CrossRef]
- Mayer-Jaekel, R.E.; Baumgartner, S.; Bilbe, G.; Ohkura, H.; Glover, D.M.; Hemmings, B.A. Molecular cloning and developmental expression of the catalytic and 65-kDa regulatory subunits of protein phosphatase 2A in Drosophila. Mol. Biol. Cell 1992, 3, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Archambault, V.; Kar, A.; Lio’, P.; D’Avino, P.P.; Sinka, R.; Lilley, K.; Laue, E.D.; Deak, P.; Capalbo, L.; et al. Multiple protein phosphatases are required for mitosis in Drosophila. Curr. Biol. 2007, 17, 293–303. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Pinson, X.; Archambault, V. PP2A-twins is antagonized by greatwall and collaborates with polo for cell cycle progression and centrosome attachment to nuclei in drosophila embryos. PLoS Genet. 2011, 7, e1002227. [Google Scholar] [CrossRef] [Green Version]
- Bánréti, Á.; Lukácsovich, T.; Csikós, G.; Erdélyi, M.; Sass, M. PP2A regulates autophagy in two alternative ways in Drosophila. Autophagy 2012, 8, 623–636. [Google Scholar] [CrossRef] [Green Version]
- Krahn, M.P.; Egger-Adam, D.; Wodarz, A. PP2A antagonizes phosphorylation of Bazooka by PAR-1 to control apical-basal polarity in dividing embryonic neuroblasts. Dev. Cell 2009, 16, 901–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brownlee, C.W.; Klebba, J.E.; Buster, D.W.; Rogers, G.C. The Protein Phosphatase 2A regulatory subunit Twins stabilizes Plk4 to induce centriole amplification. J. Cell Biol. 2011, 195, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobbelaere, J.; Josué, F.; Suijkerbuijk, S.; Baum, B.; Tapon, N.; Raff, J. A genome-wide RNAi screen to dissect centriole duplication and centrosome maturation in Drosophila. PLoS Biol. 2008, 6, e224. [Google Scholar] [CrossRef] [PubMed]
- Kotadia, S.; Kao, L.R.; Comerford, S.A.; Jones, R.T.; Hammer, R.E.; Megraw, T.L. PP2A-dependent disruption of centrosome replication and cytoskeleton organization in Drosophila by SV40 small tumor antigen. Oncogene 2008, 27, 6334–6346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, B.S.; Orr-Weaver, T.L. Drosophila protein phosphatases 2A B′ Wdb and Wrd regulate meiotic centromere localization and function of the MEI-S332 Shugoshin. Proc. Natl. Acad. Sci. USA 2017, 114, 12988–12993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolterhoff, N.; Gigengack, U.; Rumpf, S. PP2A phosphatase is required for dendrite pruning via actin regulation in Drosophila. EMBO Rep. 2020, 21, e48870. [Google Scholar] [CrossRef]
- Bastos, R.N.; Cundell, M.J.; Barr, F.A. KIF4A and PP2A-B56 form a spatially restricted feedback loop opposing Aurora B at the anaphase central spindle. J. Cell Biol. 2014, 207, 683–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cundell, M.J.; Bastos, R.N.; Zhang, T.; Holder, J.; Gruneberg, U.; Novak, B.; Barr, F.A. The BEG (PP2A-B55/ENSA/Greatwall) pathway ensures cytokinesis follows chromosome separation. Mol. Cell. 2013, 52, 393–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baril, C.; Therrien, M. Alphabet, a Ser/Thr phosphatase of the protein phosphatase 2C family, negatively regulates RAS/MAPK signaling in Drosophila. Dev. Biol. 2006, 294, 232–245. [Google Scholar] [CrossRef] [Green Version]
- Baril, C.; Sahmi, M.; Ashton-Beaucage, D.; Stronach, B.; Therrien, M. The PP2C Alphabet is a negative regulator of stress-activated protein kinase signaling in Drosophila. Genetics 2009, 181, 567–579. [Google Scholar] [CrossRef] [Green Version]
- Geuking, P.; Narasimamurthy, R.; Lemaitre, B.; Basler, K.; Leulier, F. A non-redundant role for Drosophila Mkk4 and hemipterous/Mkk7 in TAK1-mediated activation of JNK. PLoS ONE 2009, 4, e7709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leibfried, A.; Müller, S.; Ephrussi, A. A Cdc42-regulated actin cytoskeleton mediates Drosophila oocyte polarization. Development 2013, 140, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Riesgo-Escovar, J.R.; Jenni, M.; Fritz, A.; Hafen, E. The Drosophila Jun-N-terminal kinase is required for cell morphogenesis but not for DJun-dependent cell fate specification in the eye. Genes Dev. 1996, 10, 2759–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolner, S.; Jacinto, A.; Martin, P. The small GTPase Rac plays multiple roles in epithelial sheet fusion--dynamic studies of Drosophila dorsal closure. Dev. Biol. 2005, 282, 163–173. [Google Scholar] [CrossRef]
- Georgiou, M.; Marinari, E.; Burden, J.; Baum, B. Cdc42, Par6, and aPKC regulate Arp2/3-mediated endocytosis to control local adherens junction stability. Curr. Biol. 2008, 18, 1631–1638. [Google Scholar] [CrossRef] [Green Version]
- Harris, K.P.; Tepass, U. Cdc42 and Par proteins stabilize dynamic adherens junctions in the Drosophila neuroectoderm through regulation of apical endocytosis. J. Cell Biol. 2008, 183, 1129–1143. [Google Scholar] [CrossRef] [Green Version]
- Fucini, R.V.; Navarrete, A.; Vadakkan, C.; Lacomis, L.; Erdjument-Bromage, H.; Tempst, P.; Stamnes, M. Activated ADP-ribosylation factor assembles distinct pools of actin on golgi membranes. J. Biol. Chem. 2000, 275, 18824–18829. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.L.; Fucini, R.V.; Lacomis, L.; Erdjument-Bromage, H.; Tempst, P.; Stamnes, M. Coatomer-bound Cdc42 regulates dynein recruitment to COPI vesicles. J. Cell Biol. 2005, 169, 383–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Yang, J.S.; Schmider, A.B.; Soberman, R.J.; Hsu, V.W. Coordinated regulation of bidirectional COPI transport at the Golgi by CDC42. Nature 2015, 521, 529–532. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.C.; Chern, J.J.; Cai, Y.; Liu, M.; Choi, K.W. Drosophila TCTP is essential for growth and proliferation through regulation of dRheb GTPase. Nature 2007, 445, 785–788. [Google Scholar] [CrossRef]
- Le, T.P.; Vuong, L.T.; Kim, A.R.; Hsu, Y.C.; Choi, K.W. 14-3-3 proteins regulate Tctp-Rheb interaction for organ growth in Drosophila. Nat. Commun. 2016, 7, 11501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, D.J.; Grewal, S.S.; De la Cruz, A.F.; Edgar, B.A. Rheb-TOR signaling promotes protein synthesis, but not glucose or amino acid import, in Drosophila. BMC Biol. 2007, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, P.H.; Thapar, N.; Guo, L.; Martinez, M.; Maris, J.; Gau, C.L.; Lengyel, J.A.; Tamanoi, F. Drosophila Rheb GTPase is required for cell cycle progression and cell growth. J. Cell Sci. 2003, 116, 3601–3610. [Google Scholar] [CrossRef] [Green Version]
- Saucedo, L.J.; Gao, X.; Chiarelli, D.A.; Li, L.; Pan, D.; Edgar, B.A. Rheb promotes cell growth as a component of the insulin/TOR signalling network. Nat. Cell Biol. 2003, 5, 566–571. [Google Scholar] [CrossRef]
- Stocker, H.; Radimerski, T.; Schindelholz, B.; Wittwer, F.; Belawat, P.; Daram, P.; Breuer, S.; Thomas, G.; Hafen, E. Rheb is an essential regulator of S6K in controlling cell growth in Drosophila. Nat. Cell Biol. 2003, 5, 559–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adami, A.; García-Alvarez, B.; Arias-Palomo, E.; Barford, D.; Llorca, O. Structure of TOR and its complex with KOG1. Mol. Cell. 2007, 27, 509–516. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Blumhagen, R.; Lao, U.; Kuo, Y.; Edgar, B.A. LST8 regulates cell growth via target-of-rapamycin complex 2 (TORC2). Mol. Cell. Biol. 2012, 32, 2203–2213. [Google Scholar] [CrossRef] [Green Version]
- Yip, C.K.; Murata, K.; Walz, T.; Sabatini, D.M.; Kang, S.A. Structure of the human mTOR complex I and its implications for rapamycin inhibition. Mol. Cell. 2010, 38, 768–774. [Google Scholar] [CrossRef] [Green Version]
- Zur Lage, P.; Newton, F.G.; Jarman, A.P. Survey of the Ciliary Motility Machinery of Drosophila Sperm and Ciliated Mechanosensory Neurons Reveals Unexpected Cell-Type Specific Variations: A Model for Motile Ciliopathies. Front. Genet. 2019, 10, 24. [Google Scholar] [CrossRef]
- Legendre, M.; Zaragosi, L.-E.; Mitchison, H.M. Motile cilia and airway disease. Semin. Cell Dev. Biol. 2021, 110, 19–33. [Google Scholar] [CrossRef]
- Dick, T.; Ray, K.; Salz, H.K.; Chia, W. Cytoplasmic dynein (ddlc1) mutations cause morphogenetic defects and apoptotic cell death in Drosophila melanogaster. Mol. Cell Biol. 1996, 16, 1966–1977. [Google Scholar] [CrossRef] [Green Version]
- Dorus, S.; Freeman, Z.N.; Parker, E.R.; Heath, B.D.; Karr, T.L. Recent origins of sperm genes in Drosophila. Mol. Biol. Evol. 2008, 25, 2157–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, S.; Januschke, J.; Rossi, F.; Izumi, Y.; Garcia-Alvarez, G.; Gwee, S.S.; Soon, S.B.; Sidhu, H.K.; Yu, F.; et al. An ana2/ctp/mud complex regulates spindle orientation in Drosophila neuroblasts. Dev. Cell 2011, 21, 520–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duquesnoy, P.; Escudier, E.; Vincensini, L.; Freshour, J.; Bridoux, A.M.; Coste, A.; Deschildre, A.; De Blic, J.; Legendre, M.; Montantin, G.; et al. Loss-of-function mutations in the human ortholog of Chlamydomonas reinhardtii ODA7 disrupt dynein arm assembly and cause primary ciliary dyskinesia. Am. J. Hum. Genet. 2009, 85, 890–896. [Google Scholar] [CrossRef] [Green Version]
- Dobbelaere, J.; Schmidt Cernohorska, M.; Huranova, M.; Slade, D.; Dammermann, A. Cep97 Is Required for Centriole Structural Integrity and Cilia Formation in Drosophila. Curr. Biol. 2020, 30, 3045–3056.e7. [Google Scholar] [CrossRef]
- Persico, V.; Callaini, G.; Riparbelli, M.G. The Microtubule-Depolymerizing Kinesin-13 Klp10A Is Enriched in the Transition Zone of the Ciliary Structures of Drosophila melanogaster. Front. Cell Dev. Biol. 2019, 7, 173. [Google Scholar] [CrossRef]
- Spektor, A.; Tsang, W.Y.; Khoo, D.; Dynlacht, B.D. Cep97 and CP110 suppress a cilia assembly program. Cell 2007, 130, 678–690. [Google Scholar] [CrossRef] [Green Version]
- Vieillard, J.; Paschaki, M.; Duteyrat, J.L.; Augière, C.; Cortier, E.; Lapart, J.A.; Thomas, J.; Durand, B. Transition zone assembly and its contribution to axoneme formation in Drosophila male germ cells. J. Cell Biol. 2016, 214, 875–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elden, A.C.; Kim, H.J.; Hart, M.P.; Chen-Plotkin, A.S.; Johnson, B.S.; Fang, X.; Armakola, M.; Geser, F.; Greene, R.; Lu, M.M.; et al. Ataxin-2 intermediate-length polyglutamine expansions are associated with increased risk for ALS. Nature 2010, 466, 1069–1075. [Google Scholar] [CrossRef] [PubMed]
- Pyronneau, A.; He, Q.; Hwang, J.-Y.; Porch, M.; Contractor, A.; Zukin, R.S. Aberrant Rac1-cofilin signaling mediates defects in dendritic spines, synaptic function, and sensory perception in fragile X syndrome. Sci. Signal. 2017, 10, eaan0852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudhakaran, I.P.; Hillebrand, J.; Dervan, A.; Das, S.; Holohan, E.E.; Hülsmeier, J.; Sarov, M.; Parker, R.; VijayRaghavan, K.; Ramaswami, M. FMRP and Ataxin-2 function together in long-term olfactory habituation and neuronal translational control. Proc. Natl. Acad. Sci. USA 2014, 111, E99–E108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satterfield, T.F.; Jackson, S.M.; Pallanck, L.J. A Drosophila homolog of the polyglutamine disease gene SCA2 is a dosage-sensitive regulator of actin filament formation. Genetics 2002, 162, 1687–1702. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Dockendorff, T.C.; Jongens, T.A.; Dreyfuss, G. Characterization of dFMR1, a Drosophila melanogaster homolog of the fragile X mental retardation protein. Mol. Cell Biol. 2000, 20, 8536–8547. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Q.; Bailey, A.M.; Matthies, H.J.; Renden, R.B.; Smith, M.A.; Speese, S.D.; Rubin, G.M.; Broadie, K. Drosophila fragile X-related gene regulates the MAP1B homolog Futsch to control synaptic structure and function. Cell 2001, 107, 591–603. [Google Scholar] [CrossRef] [Green Version]
- Imbert, G.; Saudou, F.; Yvert, G.; Devys, D.; Trottier, Y.; Garnier, J.M.; Weber, C.; Mandel, J.L.; Cancel, G.; Abbas, N.; et al. Cloning of the gene for spinocerebellar ataxia 2 reveals a locus with high sensitivity to expanded CAG/glutamine repeats. Nat. Genet. 1996, 14, 285–291. [Google Scholar] [CrossRef]
- Pulst, S.M.; Nechiporuk, A.; Nechiporuk, T.; Gispert, S.; Chen, X.N.; Lopes-Cendes, I.; Pearlman, S.; Starkman, S.; Orozco-Diaz, G.; Lunkes, A.; et al. Moderate expansion of a normally biallelic trinucleotide repeat in spinocerebellar ataxia type 2. Nat. Genet. 1996, 14, 269–276. [Google Scholar] [CrossRef]
- Sanpei, K.; Takano, H.; Igarashi, S.; Sato, T.; Oyake, M.; Sasaki, H.; Wakisaka, A.; Tashiro, K.; Ishida, Y.; Ikeuchi, T.; et al. Identification of the spinocerebellar ataxia type 2 gene using a direct identification of repeat expansion and cloning technique, DIRECT. Nat. Genet. 1996, 14, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, R.N.; Feinberg, R.L.; Vaurio, R.; Passanante, N.M.; Thompson, R.E.; Kaufmann, W.E. Autism spectrum disorder in fragile X syndrome: A longitudinal evaluation. Am. J. Med. Genet. Part A 2009, 149A, 1125–1137. [Google Scholar] [CrossRef] [Green Version]
- Jin, P.; Warren, S.T. Understanding the molecular basis of fragile X syndrome. Hum. Mol. Genet. 2000, 9, 901–908. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, W.E.; Cortell, R.; Kau, A.S.M.; Bukelis, I.; Tierney, E.; Gray, R.M.; Cox, C.; Capone, G.T.; Stanard, P. Autism spectrum disorder in fragile X syndrome: Communication, social interaction, and specific behaviors. Am. J. Med. Genet. A 2004, 129A, 225–234. [Google Scholar] [CrossRef] [PubMed]
- O′Donnell, W.T.; Warren, S.T. A decade of molecular studies of fragile X syndrome. Annu. Rev. Neurosci. 2002, 25, 315–338. [Google Scholar] [CrossRef] [Green Version]
- Swanson, M.S.; Orr, H.T. Fragile X tremor/ataxia syndrome: Blame the messenger! Comment Neuron. 2007, 55, 535–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamanini, F.; Willemsen, R.; van Unen, L.; Bontekoe, C.; Galjaard, H.; Oostra, B.A.; Hoogeveen, A.T. Differential expression of FMR1, FXR1 and FXR2 proteins in human brain and testis. Hum. Mol. Genet. 1997, 6, 1315–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verkerk, A.J.; Pieretti, M.; Sutcliffe, J.S.; Fu, Y.H.; Kuhl, D.P.; Pizzuti, A.; Reiner, O.; Richards, S.; Victoria, M.F.; Zhang, F.P.; et al. Identification of a gene (FMR-1) containing a CGG repeat coincident with a breakpoint cluster region exhibiting length variation in fragile X syndrome. Cell 1991, 65, 905–914. [Google Scholar] [CrossRef]
- Cai, Y.; Deng, Y.; Horenkamp, F.; Reinisch, K.M.; Burd, C.G. Sac1-Vps74 structure reveals a mechanism to terminate phosphoinositide signaling in the Golgi apparatus. J. Cell Biol. 2014, 206, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Iwano, T.; Kunii, M.; Matsuda, S.; Mizuguchi, R.; Jung, Y.; Hagiwara, H.; Yoshihara, Y.; Yuzaki, M.; Harada, R.; et al. Rab8a and Rab8b are essential for several apical transport pathways but insufficient for ciliogenesis. J. Cell Sci. 2014, 127, 422–431. [Google Scholar]
- Ordónez, A.J.L.; Fernández, B.; Fdez, E.; Romo-Lozano, M.; Madero-Pérez, J.; Lobbestael, E.; Baekelandt, V.; Aiastui, A.; López de Munaín, A.; Melrose, H.L.; et al. RAB8, RAB10 and RILPL1 contribute to both LRRK2 kinase-mediated centrosomal cohesion and ciliogenesis deficits. Hum. Mol. Genet. 2019, 28, 3552–3568. [Google Scholar] [CrossRef] [Green Version]
- Mayinger, P. Phosphoinositides and vesicular membrane traffic. Biochim. Biophys. Acta 2012, 1821, 1104–1113. [Google Scholar] [CrossRef] [Green Version]
- Skop, A.R.; Bergmann, D.; Mohler, W.A.; White, J.G. Completion of cytokinesis in C. elegans requires a brefeldin A-sensitive membrane accumulation at the cleavage furrow apex. Curr. Biol. 2001, 11, 735–746. [Google Scholar] [CrossRef] [Green Version]
- D′Avino, P.P.; Giansanti, M.G.; Petronczki, M. Cytokinesis in animal cells. Cold Spring Harb. Perspect. Biol. 2015, 7, a015834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frémont, S.; Echard, A. Membrane Traffic in the Late Steps of Cytokinesis. Curr. Biol. 2018, 28, R458–R470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horgan, C.P.; Walsh, M.; Zurawski, T.H.; McCaffrey, M.W. Rab11-FIP3 localises to a Rab11-positive pericentrosomal compartment during interphase and to the cleavage furrow during cytokinesis. Biochem. Biophys. Res. Commun. 2004, 319, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Fielding, A.B.; Schonteich, E.; Matheson, J.; Wilson, G.; Yu, X.; Hickson, G.R.; Srivastava, S.; Baldwin, S.A.; Prekeris, R.; Gould, G.W. Rab11-FIP3 and FIP4 interact with Arf6 and the exocyst to control membrane traffic in cytokinesis. EMBO J. 2005, 24, 3389–3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, G.M.; Fielding, A.B.; Simon, G.C.; Yu, X.; Andrews, P.D.; Hames, R.S.; Frey, A.M.; Peden, A.A.; Gould, G.W.; Prekeris, R. Molecular characterization of Rab11 interactions with members of the family of Rab11-interacting proteins. Mol. Biol. Cell. 2005, 16, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Dambournet, D.; Machicoane, M.; Chesneau, L.; Sachse, M.; Rocancourt, M.; El Marjou, A.; Formstecher, E.; Salomon, R.; Goud, B.; Echard, A. Rab35 GTPase and OCRL phosphatase remodel lipids and F-actin for successful cytokinesis. Nat. Cell Biol. 2011, 13, 981–988. [Google Scholar] [CrossRef]
- Schiel, J.A.; Simon, G.C.; Zaharris, C.; Weisz, J.; Castle, D.; Wu, C.C.; Prekeris, R. FIP3-endosome-dependent formation of the secondary ingression mediates ESCRT-III recruitment during cytokinesis. Nat. Cell Biol. 2012, 14, 1068–1078. [Google Scholar] [CrossRef] [Green Version]
- Kouranti, I.; Sachse, M.; Arouche, N.; Goud, B.; Echard, A. Rab35 regulates an endocytic recycling pathway essential for the terminal steps of cytokinesis. Curr. Biol. 2006, 16, 1719–1725. [Google Scholar] [CrossRef] [Green Version]
- Giansanti, M.G.; Belloni, G.; Gatti, M. Rab11 is required for membrane trafficking and actomyosin ring constriction in meiotic cytokinesis of Drosophila males. Mol. Biol. Cell. 2007, 18, 5034–5047. [Google Scholar] [CrossRef] [Green Version]
- Polevoy, G.; Wei, H.C.; Wong, R.; Szentpetery, Z.; Kim, Y.J.; Goldbach, P.; Steinbach, S.K.; Balla, T.; Brill, J.A. Dual roles for the Drosophila PI 4-kinase four wheel drive in localizing Rab11 during cytokinesis. J. Cell Biol. 2009, 187, 847–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelissier, A.; Chauvin, J.P.; Lecuit, T. Trafficking through Rab11 endosomes is required for cellularization during Drosophila embryogenesis. Curr. Biol. 2003, 13, 1848–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riggs, B.; Rothwell, W.; Mische, S.; Hickson, G.R.; Matheson, J.; Hays, T.S.; Gould, G.W.; Sullivan, W. Actin cytoskeleton remodeling during early Drosophila furrow formation requires recycling endosomal components Nuclear-fallout and Rab11. J. Cell Biol. 2003, 163, 143–154. [Google Scholar] [CrossRef]
- Wienke, D.C.; Knetsch, M.L.; Neuhaus, E.M.; Reedy, M.C.; Manstein, D.J. Disruption of a dynamin homologue affects endocytosis, organelle morphology, and cytokinesis in Dictyostelium discoideum. Mol. Biol. Cell. 1999, 10, 225–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerald, N.J.; Damer, C.K.; O′Halloran, T.J.; De Lozanne, A. Cytokinesis failure in clathrin-minus cells is caused by cleavage furrow instability. Cell Motil. Cytoskeleton. 2001, 48, 213–223. [Google Scholar] [CrossRef]
- Thompson, H.M.; Skop, A.R.; Euteneuer, U.; Meyer, B.J.; McNiven, M.A. The large GTPase dynamin associates with the spindle midzone and is required for cytokinesis. Curr. Biol. 2002, 12, 2111–2117. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, S.M.; Raimondi, A.; Paradise, S.; Shen, H.; Mesaki, K.; Ferguson, A.; Destaing, O.; Ko, G.; Takasaki, J.; Cremona, O.; et al. Coordinated actions of actin and BAR proteins upstream of dynamin at endocytic clathrin-coated pits. Dev. Cell 2009, 17, 811–822. [Google Scholar] [CrossRef] [Green Version]
- Hinshaw, J.E. Dynamin and its role in membrane fission. Annu. Rev. Cell Dev. Biol. 2000, 16, 483–519. [Google Scholar] [CrossRef]
- Taylor, M.J.; Perrais, D.; Merrifield, C.J. A high precision survey of the molecular dynamics of mammalian clathrin-mediated endocytosis. PLoS Biol. 2011, 9, e1000604. [Google Scholar] [CrossRef] [Green Version]
- Lundmark, R.; Carlsson, S.R. Sorting nexin 9 participates in clathrin-mediated endocytosis through interactions with the core components. J. Biol. Chem. 2003, 278, 46772–46781. [Google Scholar] [CrossRef] [Green Version]
- Soulet, F.; Yarar, D.; Leonard, M.; Schmid, S.L. SNX9 regulates dynamin assembly and is required for efficient clathrin-mediated endocytosis. Mol. Biol. Cell. 2005, 16, 2058–2067. [Google Scholar] [CrossRef] [Green Version]
- Poodry, C.A. Shibire, a neurogenic mutant of Drosophila. Dev. Biol. 1990, 138, 464–472. [Google Scholar] [CrossRef]
- Van der Bliek, A.M.; Meyerowrtz, E.M. Dynamin-like protein encoded by the Drosophila shibire gene associated with vesicular traffic. Nature 1991, 351, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Worby, C.A.; Simonson-Leff, N.; Clemens, J.C.; Kruger, R.P.; Muda, M.; Dixon, J.E. The sorting nexin, DSH3PX1, connects the axonal guidance receptor, Dscam, to the actin cytoskeleton. J. Biol. Chem. 2001, 276, 41782–41789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundmark, R.; Carlsson, S.R. SNX9—a prelude to vesicle release. J. Cell Sci. 2009, 122, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pylypenko, O.; Lundmark, R.; Rasmuson, E.; Carlsson, S.R.; Rak, A. The PX-BAR membrane-remodeling unit of sorting nexin 9. EMBO J. 2007, 26, 4788–4800. [Google Scholar] [CrossRef] [Green Version]
- Teasdale, R.D.; Collins, B.M. Insights into the PX (phox-homology) domain and SNX (sorting nexin) protein families: Structures, functions and roles in disease. Biochem. J. 2012, 441, 39–59. [Google Scholar] [CrossRef]
- Håberg, K.; Lundmark, R.; Carlsson, S.R. SNX18 is an SNX9 paralog that acts as a membrane tubulator in AP-1-positive endosomal trafficking. J. Cell Sci. 2008, 121, 1495–1505. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.P.C.; Chircop, M. SNX9, SNX18 and SNX33 are required for progression through and completion of mitosis. J. Cell Sci. 2012, 125, 4372–4382. [Google Scholar] [CrossRef] [Green Version]
- Yarar, D.; Surka, M.C.; Leonard, M.C.; Schmid, S.L. SNX9 activities are regulated by multiple phosphoinositides through both PX and BAR domains. Traffic 2008, 9, 133–146. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Guo, Y.; Xu, L.; Pei, D. Sorting nexin 33 induces mammalian cell micronucleated phenotype and actin polymerization by interacting with Wiskott-Aldrich syndrome protein. J. Biol. Chem. 2009, 284, 21659–21669. [Google Scholar] [CrossRef] [Green Version]
- Habermann, B. The BAR-domain family of proteins: A case of bending and binding? EMBO Rep. 2004, 5, 250–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, B.J.; Kent, H.M.; Mills, I.G.; Vallis, Y.; Butler, P.J.; Evans, P.R.; McMahon, H.T. BAR domains as sensors of membrane curvature: The amphiphysin BAR structure. Science 2004, 303, 495–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badour, K.; McGavin, M.K.; Zhang, J.; Freeman, S.; Vieira, C.; Filipp, D.; Julius, M.; Mills, G.B.; Siminovitch, K.A. Interaction of the Wiskott-Aldrich syndrome protein with sorting nexin 9 is required for CD28 endocytosis and cosignaling in T cells. Proc. Natl. Acad. Sci. USA 2007, 104, 1593–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, N.; Lee, S.; Ahn, N.; Kim, S.A.; Ahn, S.G.; YongPark, Z.; Chang, S. Sorting nexin 9 interacts with dynamin 1 and N-WASP and coordinates synaptic vesicle endocytosis. J. Biol. Chem. 2007, 282, 28939–28950. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.; Ahn, N.; Chang-Ileto, B.; Park, J.; Takei, K.; Ahn, S.G.; Kim, S.A.; Di Paolo, G.; Chang, S. SNX9 regulates tubular invagination of the plasma membrane through interaction with actin cytoskeleton and dynamin 2. J. Cell Sci. 2008, 121, 1252–1263. [Google Scholar] [CrossRef] [Green Version]
- Gallop, J.L.; Walrant, A.; Cantley, L.C.; Kirschner, M.W. Phosphoinositides and membrane curvature switch the mode of actin polymerization via selective recruitment of toca-1 and Snx9. Proc. Natl. Acad. Sci. USA 2013, 110, 7193–7198. [Google Scholar] [CrossRef] [Green Version]
- Posor, Y.; Eichhorn-Gruenig, M.; Puchkov, D.; Schöneberg, J.; Ullrich, A.; Lampe, A.; Müller, R.; Zarbakhshisa, S.; Gulluni, F.; Hirsch, E.; et al. Spatiotemporal control of endocytosis by phosphatidylinositol-3,4-bisphosphate. Nature 2013, 499, 233–237. [Google Scholar] [CrossRef]
- Yarar, D.; Waterman-Storer, C.M.; Schmid, S.L. SNX9 couples actin assembly to phosphoinositide signals and is required for membrane remodeling during endocytosis. Dev. Cell 2007, 13, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Prekeris, R.; Gould, G.W. Breaking up is hard to do—membrane traffic in cytokinesis. J. Cell Sci. 2008, 121, 1569–1576. [Google Scholar] [CrossRef] [Green Version]
- Skop, A.R.; Liu, H.; Yates, J., 3rd; Meyer, B.J.; Heald, R. Dissection of the mammalian midbody proteome reveals conserved cytokinesis mechanisms. Science 2004, 305, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Bobinnec, Y.; Marcaillou, C.; Morin, X.; Debec, A. Dynamics of the endoplasmic reticulum during early development of Drosophila melanogaster. Cell Motil. Cytoskeleton. 2003, 54, 217–225. [Google Scholar] [CrossRef]
- Giansanti, M.G.; Bonaccorsi, S.; Kurek, R.; Farkas, R.M.; Dimitri, P.; Fuller, M.T.; Gatti, M. The class I PITP giotto is required for Drosophila cytokinesis. Curr. Biol. 2006, 16, 195–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudelka, M.R.; Ju, T.; Heimburg-Molinaro, J.; Cummings, R.D. Simple sugars to complex disease--mucin-type O-glycans in cancer. Adv. Cancer Res. 2015, 126, 53–135. [Google Scholar] [PubMed] [Green Version]
- Wymann, M.P.; Schneiter, R. Lipid signalling in disease. Nat. Rev. Mol. Cell Biol. 2008, 9, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Buschman, M.D.; Xing, M.; Field, S.J. The GOLPH3 pathway regulates Golgi shape and function and is activated by DNA damage. Front. Neurosci. 2015, 9, 362. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Menárguez, J.Á.; Tomás, M.; Martínez-Martínez, N.; Martínez-Alonso, E. Golgi Fragmentation in Neurodegenerative Diseases: Is There a Common Cause? Cells 2019, 8, 748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciosk, R.; DePalma, M.; Priess, J.R. ATX-2, the C. elegans ortholog of ataxin 2, functions in translational regulation in the germline. Development 2004, 131, 4831–4841. [Google Scholar] [CrossRef] [Green Version]
- Fittschen, M.; Lastres-Becker, I.; Halbach, M.V.; Damrath, E.; Gispert, S.; Azizov, M.; Walter, M.; Müller, S.; Auburger, G. Genetic ablation of ataxin-2 increases several global translation factors in their transcript abundance but decreases translation rate. Neurogenetics 2015, 16, 181–192. [Google Scholar] [CrossRef] [Green Version]
- McCann, C.; Holohan, E.E.; Das, S.; Dervan, A.; Larkin, A.; Lee, J.A.; Rodrigues, V.; Parker, R.; Ramaswami, M. The Ataxin-2 protein is required for microRNA function and synapse-specific long-term olfactory habituation. Proc. Natl. Acad. Sci. USA 2011, 108, E655–E662. [Google Scholar] [CrossRef] [Green Version]
- Satterfield, T.F.; Pallanck, L.J. Ataxin-2 and its Drosophila homolog, ATX2, physically assemble with polyribosomes. Hum. Mol. Genet. 2006, 15, 2523–2532. [Google Scholar] [CrossRef] [Green Version]
- Yokoshi, M.; Li, Q.; Yamamoto, M.; Okada, H.; Suzuki, Y.; Kawahara, Y. Direct binding of Ataxin-2 to distinct elements in 3′ UTRs promotes mRNA stability and protein expression. Mol. Cell. 2014, 55, 186–198. [Google Scholar] [CrossRef] [Green Version]
- Nonis, D.; Schmidt, M.H.H.; Van de Loo, S.; Eich, F.; Dikic, I.; Nowock, J.; Auburger, G. Ataxin-2 associates with the endocytosis complex and affects EGF receptor trafficking. Cell. Signal. 2008, 20, 1725–1739. [Google Scholar] [CrossRef]
- Ralser, M.; Nonhoff, U.; Albrecht, M.; Lengauer, T.; Wanker, E.E.; Lehrach, H.; Krobitsch, S. Ataxin-2 and huntingtin interact with endophilin-A complexes to function in plastin-associated pathways. Hum. Mol. Genet. 2005, 14, 2893–2909. [Google Scholar] [CrossRef]
- Huynh, D.P.; Yang, H.T.; Vakharia, H.; Nguyen, D.; Pulst, S.M. Expansion of the polyQ repeat in ataxin-2 alters its Golgi localization, disrupts the Golgi complex and causes cell death. Hum. Mol. Genet. 2003, 12, 1485–1496. [Google Scholar] [CrossRef]
- Van de Loo, S.; Eich, F.; Nonis, D.; Auburger, G.; Nowock, J. Ataxin-2 associates with rough endoplasmic reticulum. Exp. Neurol. 2009, 215, 110–118. [Google Scholar] [CrossRef]
- Del Castillo, U.; Gnazzo, M.M.; Sorensen Turpin, C.G.; Nguyen, K.C.Q.; Semaya, E.; Lam, Y.; De Cruz, M.A.; Bembenek, J.N.; Hall, D.H.; Riggs, B.; et al. Conserved role for Ataxin-2 in mediating endoplasmic reticulum dynamics. Traffic 2019, 20, 436–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devys, D.; Lutz, Y.; Rouyer, N.; Bellocq, J.P.; Mandel, J.L. The FMR-1 protein is cytoplasmic, most abundant in neurons and appears normal in carriers of a fragile X premutation. Nat. Genet. 1993, 4, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Li, W.; Xu, K.; Bogert, B.A.; Su, K.; Gao, F.B. Control of dendritic development by the Drosophila fragile X-related gene involves the small GTPase Rac1. Development. 2003, 130, 5543–5552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeve, S.P.; Bassetto, L.; Genova, G.K.; Kleyner, Y.; Leyssen, M.; Jackson, F.R.; Hassan, B.A. The Drosophila fragile X mental retardation protein controls actin dynamics by directly regulating profilin in the brain. Curr. Biol. 2005, 15, 1156–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caudy, A.A.; Myers, M.; Hannon, G.J.; Hammond, S.M. Fragile X-related protein and VIG associate with the RNA interference machinery. Genes Dev. 2002, 16, 2491–2496. [Google Scholar] [CrossRef] [Green Version]
- Ishizuka, A.; Siomi, M.C.; Siomi, H. A Drosophila fragile X protein interacts with components of RNAi and ribosomal proteins. Genes Dev. 2002, 16, 2497–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plante, I.; Davidovic, L.; Ouellet, D.L.; Gobeil, L.-A.; Tremblay, S.; Khandjian, E.W.; Provost, P. Dicer-Derived MicroRNAs Are Utilized by the Fragile X Mental Retardation Protein for Assembly on Target RNAs. J. Biomed. Biotechnol. 2006, 2006, 64347. [Google Scholar] [CrossRef] [PubMed]
- Bozzetti, M.P.; Specchia, V.; Cattenoz, P.B.; Laneve, P.; Geusa, A.; Sahin, H.B.; Di Tommaso, S.; Friscini, A.; Massari, S.; Diebold, C.; et al. The Drosophila fragile X mental retardation protein participates in the piRNA pathway. J. Cell Sci. 2015, 128, 2070–2084. [Google Scholar] [CrossRef] [Green Version]
- Specchia, V.; D′Attis, S.; Puricella, A.; Bozzetti, M.P. dFmr1 Plays Roles in Small RNA Pathways of Drosophila melanogaster. Int. J. Mol. Sci. 2017, 18, 1066. [Google Scholar] [CrossRef] [Green Version]
- Puthanveettil, S.V. RNA transport and long-term memory storage. RNA Biol. 2013, 10, 1765–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnazzo, M.M.; Uhlemann, E.E.; Villarreal, A.R.; Shirayama, M.; Dominguez, E.G.; Skop, A.R. The RNA-binding protein ATX-2 regulates cytokinesis through PAR-5 and ZEN-4. Mol. Biol. Cell. 2016, 27, 3052–3064. [Google Scholar] [CrossRef]
- Abekhoukh, S.; Sahin, H.B.; Grossi, M.; Zongaro, S.; Maurin, T.; Madrigal, I.; Kazue-Sugioka, D.; Raas-Rothschild, A.; Doulazmi, M.; Carrera, P.; et al. New insights into the regulatory function of CYFIP1 in the context of WAVE- and FMRP-containing complexes. Dis. Model Mech. 2017, 10, 463–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenck, A.; Bardoni, B.; Moro, A.; Bagni, C.; Mandel, J.-L. A highly conserved protein family interacting with the fragile X mental retardation protein (FMRP) and displaying selective interactions with FMRP-related proteins FXR1P and FXR2P. Proc. Nat. Acad. Sci. USA 2001, 98, 8844–8849. [Google Scholar] [CrossRef] [Green Version]
- Burman, J.L.; Bourbonniere, L.; Philie, J.; Stroh, T.; Dejgaard, S.Y.; Presley, J.F.; McPherson, P.S. Scyl1, mutated in a recessive form of spinocerebellar neurodegeneration, regulates COPI-mediated retrograde traffic. J. Biol. Chem. 2008, 283, 22774–22786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, C.J.; Evans, M.; Thayanithy, V.; Taniguchi-Ishigaki, N.; Bach, I.; Kolpak, A.; Bassell, G.J.; Rossoll, W.; Lorson, C.L.; Bao, Z.Z.; et al. The COPI vesicle complex binds and moves with survival motor neuron within axons. Hum. Mol. Genet. 2011, 20, 1701–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, A.G.; Lin, H.; Ebert, A.D.; Liu, Y.; Androphy, E.J. COPI transport complexes bind to specific RNAs in neuronal cells. Hum. Mol. Genet. 2013, 22, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Lucá, R.; Averna, M.; Zalfa, F.; Vecchi, M.; Bianchi, F.; La Fata, G.; Del Nonno, F.; Nardacci, R.; Bianchi, M.; Nuciforo, P.; et al. The fragile X protein binds mRNAs involved in cancer progression and modulates metastasis formation. EMBO Mol. Med. 2013, 5, 1523–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalfa, F.; Panasiti, V.; Carotti, S.; Zingariello, M.; Perrone, G.; Sancillo, L.; Pacini, L.; Luciani, F.; Roberti, V.; D’Amico, S.; et al. The fragile X mental retardation protein regulates tumor invasiveness-related pathways in melanoma cells. Cell Death Dis. 2017, 8, e3169. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProtKD Entry 1 | Drosophila Annotation Symbol 2 | Drosophila Symbol 3 | PEP 4 |
|---|---|---|---|
| Golgi vesicle transport—GO: 0048193 | |||
| Q9W0B8 | CG7961 | αCOP | 3.71 × 10−115 |
| P45437 | CG6223 | βCOP | 2.64 × 10−71 |
| O62621 | CG6699 | β’COP | 6.21 × 10−70 |
| Q8I0G5 | CG1528 | γCOP | 6.38 × 10−40 |
| Q9W555 | CG14813 | δCOP | 1.35 × 10−28 |
| A1Z7J7 | CG8266 | Sec31 | 2.17× 10−15 |
| M9PGI6 | CG7359 | Sec22 | 1.47 × 10−12 |
| Q9VLS7 | CG8552 | PAPLA1 | 1.59 × 10−10 |
| Q9VQ94 | CG10882 | Sec24CD | 3.34 × 10−10 |
| Q9VNF8 | CG1250 | Sec23 | 1.29 × 10−09 |
| Q24179 | CG3539 | Slh | 1.88 × 10−05 |
| Q9Y0Y5 | CG9543 | εCOP | 1.83 × 10−04 |
| Q9VAD6 | CG31040 | Cog7 | 4.91 × 10−03 |
| Rab protein signal transduction—GO: 0032482 | |||
| O18332 | CG3320 | Rab1 | 3.74 × 10−69 |
| A1Z7S3 | CG8024 | Rab32 | 5.99 × 10−23 |
| Q9V3I2 | CG3664 | Rab5 | 2.60 × 10−22 |
| O18338 | CG8287 | Rab8 | 5.90 × 10−06 |
| O15971 | CG17060 | Rab10 | 5.90 × 10−06 |
| Q86BK8 | CG4212 | Rab14 | 1.63 × 10−05 |
| O18335 | CG5771 | Rab11 | 8.83 × 10−05 |
| Transport—GO: 006810 | |||
| P29742 | CG9012 | Chc | 1.36 × 10−43 |
| Q9VS57 | CG8583 | Sec63 | 1.58 × 10−43 |
| Q9V3D9 | CG4659 | Srp54k | 1.90 × 10−22 |
| P27619 | CG18102 | Shi | 1.23 × 10−10 |
| Q9VEX2 | CG6963 | Gish | 1.21 × 10−05 |
| Q9NCC3 | CG6757 | SH3PX1 | 3.91 × 10−05 |
| Protein glycosylation—GO: 0006486 | |||
| Q7K110 | CG6370 | OstΔ | 5.06 × 10−11 |
| Q09332 | CG6850 | Uggt | 9.90 × 10−10 |
| Q9VP06 | CG11306 | Alg11 | 6.42 × 10−08 |
| Q9XZ08 | CG15110 | Botv | 9.17 × 10−06 |
| Q6WV17 | CG31651 | Pgant5 | 1.87 × 10−05 |
| Q9VTZ6 | CG10688 | Pmm2 | 3.61 × 10−04 |
| Q8MV48 | CG6394 | Pgant7 | 1.17 × 10−02 |
| Lipid metabolic process—GO: 0006629 | |||
| Q9W0I6 | CG9128 | Sac1 | 1.43 × 10−49 |
| Q9VXH3 | CG4200 | Sl | 1.04 × 10−10 |
| Q9VL10 | CG31873 | Mulk | 4.69 × 10−10 |
| Q9VGM0 | CG6790 | CG6790 | 4.19 × 10−09 |
| Q9VZL3 | CG10849 | Sc2 | 1.13 × 10−08 |
| Q9VGL9 | CG5342 | CG5342 | 2.21 × 10−08 |
| Q9VCV7 | CG4907 | CG4907 | 3.48 × 10−07 |
| Q9V7Y2 | CG8946 | Sply | 2.20 × 10−06 |
| Q9VCY7 | CG6921 | Bond | 5.56 × 10−04 |
| Q9W3G0 | CG11190 | PIG-T | 3.44 × 10−03 |
| Golgi organization—GO: 0007030 | |||
| Q7KN62 | CG2331 | TER94 | 5.55 × 10−45 |
| Q7KNA0 | CG8230 | CG8230 | 4.24 × 10−16 |
| Q9V3A8 | CG6822 | ergic53 | 1.23 × 10−05 |
| Cell cycle—GO: 0007049 | |||
| P36179 | CG17291 | Pp2A-29B | 2.55 × 10−11 |
| Q9VAK1 | CG1906 | Alph | 2.56 × 10−05 |
| P36872 | CG6235 | Tws | 2.18 × 10−03 |
| P40423 | CG3595 | Sqh | 4.19 × 10−03 |
| Signaling—GO: 0023052 | |||
| P29310 | CG17870 | 14-3-3ζ | 1.36 × 10−57 |
| P40792 | CG2248 | Rac1 | 1.01 × 10−41 |
| P48554 | CG8556 | Rac2 | 1.01 × 10−41 |
| Q9VGS2 | CG4800 | Tctp | 1.71 × 10−06 |
| O61444 | CG9738 | Mkk4 | 1.43 × 10-04 |
| Q9W328 | CG3004 | Lst8 | 8.71 × 10−04 |
| P40793 | CG12530 | Cdc42 | 1.72 × 10−03 |
| Cilium assembly—GO: 0060271 | |||
| Q8INT5 | CG31623 | Dtr | 2.96 × 10−15 |
| Q9V3M9 | CG4767 | Tektin-A | 4.85 × 10−13 |
| Q9W1D3 | CG3121 | Rsph4a | 1.12 × 10−08 |
| O96860 | CG5450 | Cdlc2 | 1.58 × 10−05 |
| Q24117 | CG6998 | Ctp | 1.58 × 10−05 |
| Q960Z0 | CG1453 | Klp10A | 1.61 × 10-05 |
| Q8T3V7 | CG31803 | Rsph9 | 1.72 × 10−05 |
| Q9W1U1 | CG12192 | Klp59D | 1.92 × 10−05 |
| Q9VQV7 | CG3980 | Cep97 | 3.60 × 10−05 |
| Q9VJY4 | CG10859 | CG10859 | 2.45 × 10−04 |
| Q8MSJ9 | CG9313 | CG9313 | 1.12 × 10−03 |
| Nervous system process—GO: 0050877 | |||
| Q9NFU0 | CG6203 | Fmr1 | 9.29 × 10−41 |
| Q8SWR8 | CG5166 | Atx2 | 8.52 × 10−11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sechi, S.; Karimpour-Ghahnavieh, A.; Frappaolo, A.; Di Francesco, L.; Piergentili, R.; Schininà, E.; D’Avino, P.P.; Giansanti, M.G. Identification of GOLPH3 Partners in Drosophila Unveils Potential Novel Roles in Tumorigenesis and Neural Disorders. Cells 2021, 10, 2336. https://doi.org/10.3390/cells10092336
Sechi S, Karimpour-Ghahnavieh A, Frappaolo A, Di Francesco L, Piergentili R, Schininà E, D’Avino PP, Giansanti MG. Identification of GOLPH3 Partners in Drosophila Unveils Potential Novel Roles in Tumorigenesis and Neural Disorders. Cells. 2021; 10(9):2336. https://doi.org/10.3390/cells10092336
Chicago/Turabian StyleSechi, Stefano, Angela Karimpour-Ghahnavieh, Anna Frappaolo, Laura Di Francesco, Roberto Piergentili, Eugenia Schininà, Pier Paolo D’Avino, and Maria Grazia Giansanti. 2021. "Identification of GOLPH3 Partners in Drosophila Unveils Potential Novel Roles in Tumorigenesis and Neural Disorders" Cells 10, no. 9: 2336. https://doi.org/10.3390/cells10092336
APA StyleSechi, S., Karimpour-Ghahnavieh, A., Frappaolo, A., Di Francesco, L., Piergentili, R., Schininà, E., D’Avino, P. P., & Giansanti, M. G. (2021). Identification of GOLPH3 Partners in Drosophila Unveils Potential Novel Roles in Tumorigenesis and Neural Disorders. Cells, 10(9), 2336. https://doi.org/10.3390/cells10092336