
Current Understanding of the Neutrophil Transcriptome in Health and Disease


Abstract
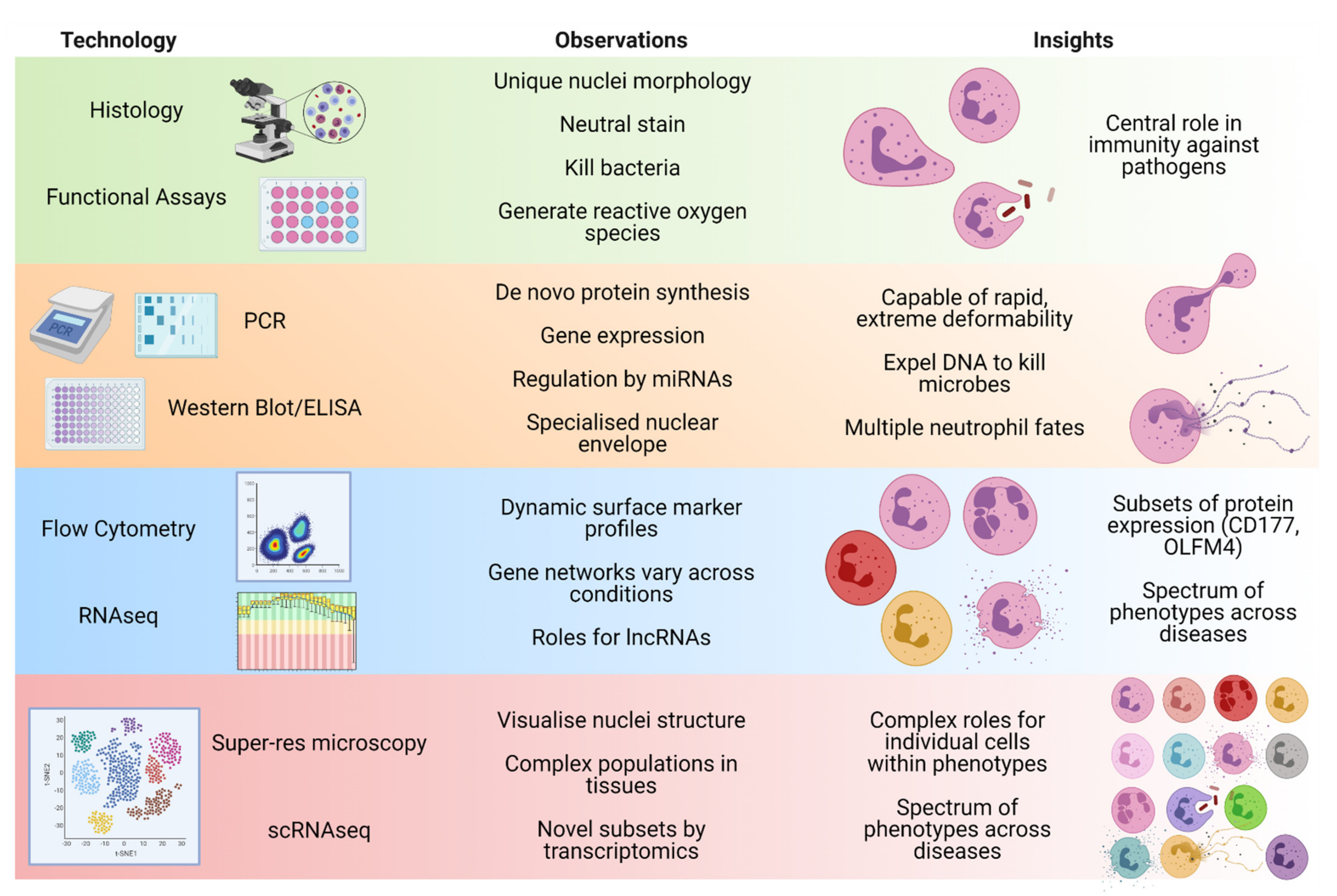
:1. Introduction
2. Neutrophil Development
3. Neutrophil Transcriptional Infrastructure
3.1. The Distinctive Neutrophil Nucleus
3.2. Transcription and Nuclei Structure Intertwine
3.3. Homeostatic Transcription
4. Transcriptional Responses during Disease
4.1. Infection Response
4.2. Chronic Inflammation
4.3. Cancer
5. Priority Research Areas
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liew, P.X.; Kubes, P. The Neutrophil’s role during health and disease. Physiol. Rev. 2019, 99, 1223–1248. [Google Scholar] [CrossRef] [PubMed]
- Granelli-Piperno, A.; Vassalli, J.D.; Reich, E. RNA and protein synthesis in human peripheral blood polymorphonuclear leukocytes. J. Exp. Med. 1979, 149, 284–289. [Google Scholar] [CrossRef]
- Itoh, K.; Okubo, K.; Utiyama, H.; Hirano, T.; Yoshii, J.; Matsubara, K. Expression Profile of Active Genes in Granulocytes. Blood 1998, 92, 1432–1441. [Google Scholar] [CrossRef] [PubMed]
- Newburger, P.E.; Subrahmanyam, Y.; Weissman, S.M. Global analysis of neutrophil gene expression. Curr. Opin. Hematol. 2000, 7, 16–20. [Google Scholar] [CrossRef]
- Xie, X.; Shi, Q.; Wu, P.; Zhang, X.; Kambara, H.; Su, J.; Yu, H.; Park, S.Y.; Guo, R.; Ren, Q.; et al. Single-cell transcriptome profiling reveals neutrophil heterogeneity in homeostasis and infection. Nat. Immunol. 2020, 21, 1119–1133. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.J.; Fan, J. Discovering myeloid cell heterogeneity in the lung by means of next generation sequencing. Mil. Med. Res. 2019, 6, 1–10. [Google Scholar] [CrossRef]
- Ai, Z.; Udalova, I.A. Transcriptional regulation of neutrophil differentiation and function during inflammation. J. Leukoc. Biol. 2020, 107, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Laslo, P.; Spooner, C.J.; Warmflash, A.; Lancki, D.W.; Lee, H.-J.; Sciammas, R.; Gantner, B.N.; Dinner, A.R.; Singh, H. Multilineage transcriptional priming and determination of alternate hematopoietic cell fates. Cell 2006, 126, 755–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahoz-Beneytez, J.; Elemans, M.; Zhang, Y.; Ahmed, R.; Salam, A.; Block, M.; Niederalt, C.; Asquith, B.; Macallan, D. Human neutrophil kinetics: Modeling of stable isotope labeling data supports short blood neutrophil half-lives. Blood 2016, 127, 3431–3438. [Google Scholar] [CrossRef] [Green Version]
- Hoogendijk, A.J.; Pourfarzad, F.; Aarts, C.E.M.; Tool, A.T.J.; Hiemstra, I.H.; Grassi, L.; Frontini, M.; Meijer, A.B.; van den Biggelaar, M.; Kuijpers, T.W. Dynamic Transcriptome-Proteome Correlation Networks Reveal Human Myeloid Differentiation and Neutrophil-Specific Programming. Cell Rep. 2019, 29, 2505–2519.e4. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, A.; Chilvers, E.R.; Summers, C.; Koenderman, L. The Neutrophil Life Cycle. Trends Immunol. 2019, 40, 584–597. [Google Scholar] [CrossRef] [PubMed]
- Silvestre-Roig, C.; Fridlender, Z.G.; Glogauer, M.; Scapini, P. Neutrophil Diversity in Health and Disease. Trends Immunol. 2019, 40, 565–583. [Google Scholar] [CrossRef]
- Goldschmeding, R.; van Dalen, C.M.; Faber, N.; Calafat, J.; Huizinga, T.W.; van der Schoot, C.E.; Clement, L.T.; von dem Borne, A.E. Further characterization of the NB 1 antigen as a variably expressed 56-62 kD GPI-linked glycoprotein of plasma membranes and specific granules of neutrophils. Br. J. Haematol. 1992, 81, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Welin, A.; Amirbeagi, F.; Christenson, K.; Björkman, L.; Björnsdottir, H.; Forsman, H.; Dahlgren, C.; Karlsson, A.; Bylund, J. The Human Neutrophil Subsets Defined by the Presence or Absence of OLFM4 Both Transmigrate into Tissue In Vivo and Give Rise to Distinct NETs In Vitro. PLoS ONE 2013, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Marini, O.; Costa, S.; Bevilacqua, D.; Calzetti, F.; Tamassia, N.; Spina, C.; De Sabata, D.; Tinazzi, E.; Lunardi, C.; Scupoli, M.T.; et al. Mature CD10+ and immature CD10− neutrophils present in G-CSF–treated donors display opposite effects on T cells. Blood 2017, 129, 1343–1356. [Google Scholar] [CrossRef] [Green Version]
- Giacalone, V.D.; Margaroli, C.; Mall, M.A.; Tirouvanziam, R. Neutrophil adaptations upon recruitment to the lung: New concepts and implications for homeostasis and disease. Int. J. Mol. Sci. 2020, 21, 851. [Google Scholar] [CrossRef] [Green Version]
- Hartl, D.; Krauss-Etschmann, S.; Koller, B.; Hordijk, P.L.; Kuijpers, T.W.; Hoffmann, F.; Hector, A.; Eber, E.; Marcos, V.; Bittmann, I.; et al. Infiltrated Neutrophils Acquire Novel Chemokine Receptor Expression and Chemokine Responsiveness in Chronic Inflammatory Lung Diseases. J. Immunol. 2008, 181, 8053–8067. [Google Scholar] [CrossRef]
- Bongers, S.H.; Chen, N.; van Grinsven, E.; van Staveren, S.; Hassani, M.; Spijkerman, R.; Hesselink, L.; Lo Tam Loi, A.T.; van Aalst, C.; Leijte, G.P.; et al. Kinetics of Neutrophil Subsets in Acute, Subacute, and Chronic Inflammation. Front. Immunol. 2021, 12, 2386. [Google Scholar] [CrossRef]
- Hardisty, G.R.; Llanwarne, F.; Minns, D.; Gillan, J.L.; Davidson, D.J.; Gwyer Findlay, E.; Gray, R.D. High Purity Isolation of Low Density Neutrophils Casts Doubt on Their Exceptionality in Health and Disease. Front. Immunol. 2021, 12, 2057. [Google Scholar] [CrossRef]
- Ng, L.G.; Ostuni, R.; Hidalgo, A. Heterogeneity of neutrophils. Nat. Rev. Immunol. 2019, 19, 255–265. [Google Scholar] [CrossRef]
- Manley, H.R.; Keightley, M.C.; Lieschke, G.J. The Neutrophil Nucleus: An Important Influence on Neutrophil Migration and Function. Front. Immunol. 2018, 9, 2867. [Google Scholar] [CrossRef]
- Thiam, H.-R.R.; Vargas, P.; Carpi, N.; Crespo, C.L.; Raab, M.; Terriac, E.; King, M.C.; Jacobelli, J.; Alberts, A.S.; Stradal, T.; et al. Perinuclear Arp2/3-driven actin polymerization enables nuclear deformation to facilitate cell migration through complex environments. Nat. Commun. 2016, 7, 10997. [Google Scholar] [CrossRef]
- Fingerhut, L.; Dolz, G.; de Buhr, N. What is the evolutionary fingerprint in neutrophil granulocytes? Int. J. Mol. Sci. 2020, 21, 4523. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.A.; Bass, D.A.; Trillo, A.A.; Snyder, M.S.; DeChatelet, L.R. Functional and metabolic studies of polymorphonuclear leukocytes in the congenital Pelger-Huet anomaly. Blood 1980, 55, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Rowat, A.C.; Jaalouk, D.E.; Zwerger, M.; Ung, W.L.; Eydelnant, I.A.; Olins, D.E.; Olins, A.L.; Herrmann, H.; Weitz, D.A.; Lammerding, J. Nuclear envelope composition determines the ability of neutrophil-type cells to passage through micron-scale constrictions. J. Biol. Chem. 2013, 288, 8610–8618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, L.J.; Maher, D.W.; Tay, D.L.; Boyd, A.W.; Rockman, S.; McGrath, K.; Fox, R.M.; Morstyn, G. Marrow proliferation and the appearance of giant neutrophils in response to recombinant human granulocyte colony stimulating factor (rhG-CSF). Br. J. Haematol. 1992, 80, 298–304. [Google Scholar] [CrossRef]
- Kaferle, J.; Strzoda, C.E. Evaluation of macrocytosis. Am. Fam. Physician 2009, 79, 203–208. [Google Scholar]
- Whitmore, L.C.; Weems, M.N.; Allen, L.-A.H. Cutting Edge: Helicobacter pylori Induces Nuclear Hypersegmentation and Subtype Differentiation of Human Neutrophils In Vitro. J. Immunol. 2017, 198, 1793–1797. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, S.; Kim, S.-Y.; Yun, Y.-J.; Kim, J.-K.; Lee, J.M.; Shin, M.; Song, D.-K.; Hong, C.-W. Retinoic acid induces hypersegmentation and enhances cytotoxicity of neutrophils against cancer cells. Immunol. Lett. 2017, 182, 24–29. [Google Scholar] [CrossRef]
- Keenan, C.R.; Mlodzianoski, M.J.; Coughlan, H.D.; Bediaga, N.G.; Naselli, G.; Lucas, E.C.; Wang, Q.; de Graaf, C.A.; Hilton, D.J.; Harrison, L.C.; et al. Chromosomes distribute randomly to, but not within, human neutrophil nuclear lobes. iScience 2021, 24, 102161. [Google Scholar] [CrossRef]
- Hübner, B.; Lomiento, M.; Mammoli, F.; Illner, D.; Markaki, Y.; Ferrari, S.; Cremer, M.; Cremer, T. Remodeling of nuclear landscapes during human myelopoietic cell differentiation maintains co-aligned active and inactive nuclear compartments. Epigenetics Chromatin 2015, 8, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Gong, K.; Denholtz, M.; Chandra, V.; Kamps, M.P.; Alber, F.; Murre, C. Comprehensive characterization of neutrophil genome topology. Genes Dev. 2017, 31, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Tamassia, N.; Zimmermann, M.; Castellucci, M.; Ostuni, R.; Bruderek, K.; Schilling, B.; Brandau, S.; Bazzoni, F.; Natoli, G.; Cassatella, M.A. Cutting Edge: An Inactive Chromatin Configuration at the IL-10 Locus in Human Neutrophils. J. Immunol. 2013, 190, 1921–1925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yvan-Charvet, L.; Ng, L.G. Granulopoiesis and Neutrophil Homeostasis: A Metabolic, Daily Balancing Act. Trends Immunol. 2019, 40, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.J.L.; Ritchie, W.; Ebner, O.A.; Selbach, M.; Wong, J.W.H.; Huang, Y.; Gao, D.; Pinello, N.; Gonzalez, M.; Baidya, K.; et al. Orchestrated intron retention regulates normal granulocyte differentiation. Cell 2013, 154, 583–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaines, P.; Tien, C.W.; Olins, A.L.; Olins, D.E.; Shultz, L.D.; Carney, L.; Berliner, N. Mouse neutrophils lacking lamin B-receptor expression exhibit aberrant development and lack critical functional responses. Exp. Hematol. 2008, 36, 965–976. [Google Scholar] [CrossRef] [Green Version]
- Martewicz, S.; Luni, C.; Zhu, X.; Cui, M.; Hu, M.; Qu, S.; Buratto, D.; Yang, G.; Grespan, E.; Elvassore, N. Nuclear Morphological Remodeling in Human Granulocytes Is Linked to Prenylation Independently from Cytoskeleton. Cells 2020, 9, 2509. [Google Scholar] [CrossRef]
- Rydzynska, Z.; Pawlik, B.; Krzyzanowski, D.; Mlynarski, W.; Madzio, J. Neutrophil Elastase Defects in Congenital Neutropenia. Front. Immunol. 2021, 12, 1382. [Google Scholar] [CrossRef]
- Tidwell, T.; Wechsler, J.; Nayak, R.C.; Trump, L.; Salipante, S.J.; Cheng, J.C.; Donadieu, J.; Glaubach, T.; Corey, S.J.; Grimes, H.L.; et al. Neutropenia-associated ELANE mutations disrupting translation initiation produce novel neutrophil elastase isoforms. Blood 2014, 123, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, M.S.; Laurino, M.Y.; Keel, S.B. Normal peripheral blood neutrophil numbers accompanying ELANE whole gene deletion mutation. Blood Adv. 2019, 3, 2470–2473. [Google Scholar] [CrossRef] [Green Version]
- Fischer, J.; Walter, C.; Tönges, A.; Aleth, H.; Jordão, M.J.C.; Leddin, M.; Gröning, V.; Erdmann, T.; Lenz, G.; Roth, J.; et al. Safeguard function of PU.1 shapes the inflammatory epigenome of neutrophils. Nat. Immunol. 2019, 20, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Watt, S.; Vasquez, L.; Walter, K.; Mann, A.L.; Kundu, K.; Chen, L.; Sims, Y.; Ecker, S.; Burden, F.; Farrow, S.; et al. Genetic perturbation of PU.1 binding and chromatin looping at neutrophil enhancers associates with autoimmune disease. Nat. Commun. 2021, 12, 2298. [Google Scholar] [CrossRef] [PubMed]
- Adrover, J.M.; del Fresno, C.; Crainiciuc, G.; Cuartero, M.I.; Casanova-Acebes, M.; Weiss, L.A.; Huerga-Encabo, H.; Silvestre-Roig, C.; Rossaint, J.; Cossío, I.; et al. A Neutrophil Timer Coordinates Immune Defense and Vascular Protection. Immunity 2019, 50, 390–402. [Google Scholar] [CrossRef] [Green Version]
- Ella, K.; Csépányi-Kömi, R.; Káldi, K. Circadian regulation of human peripheral neutrophils. Brain. Behav. Immun. 2016, 57, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Chen, G.; Manwani, D.; Mortha, A.; Xu, C.; Faith, J.J.; Burk, R.D.; Kunisaki, Y.; Jang, J.E.; Scheiermann, C.; et al. Neutrophil ageing is regulated by the microbiome. Nature 2015, 525, 528–532. [Google Scholar] [CrossRef]
- Chen, L.; Ge, B.; Casale, F.P.; Vasquez, L.; Kwan, T.; Garrido-Martín, D.; Watt, S.; Yan, Y.; Kundu, K.; Ecker, S.; et al. Genetic Drivers of Epigenetic and Transcriptional Variation in Human Immune Cells. Cell 2016, 167, 1398–1414. [Google Scholar] [CrossRef] [Green Version]
- Ecker, S.; Chen, L.; Pancaldi, V.; Bagger, F.O.; Fernández, J.M.; Carrillo de Santa Pau, E.; Juan, D.; Mann, A.L.; Watt, S.; Casale, F.P.; et al. Genome-wide analysis of differential transcriptional and epigenetic variability across human immune cell types. Genome Biol. 2017, 18, 18. [Google Scholar] [CrossRef] [Green Version]
- Radom-Aizik, S.; Zaldivar, F.; Leu, S.-Y.; Galassetti, P.; Cooper, D.M. Effects of 30 min of aerobic exercise on gene expression in human neutrophils. J. Appl. Physiol. 2008, 104, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, E.C.; Perry, J.K.; Long, D.S.; Olins, A.L.; Olins, D.E.; Wright, B.E.; Vickers, M.H.; O’Sullivan, J.M. Migration through a small pore disrupts inactive chromatin organization in neutrophil-like cells. BMC Biol. 2018, 16, 142. [Google Scholar] [CrossRef] [Green Version]
- Wright, H.L.; Thomas, H.B.; Moots, R.J.; Edwards, S.W. RNA-seq reveals activation of both common and cytokine-specific pathways following neutrophil priming. PLoS ONE 2013, 8, e58598. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Kluger, Y.; Nakayama, Y.; Poddar, R.; Whitney, C.; Detora, A.; Weissman, S.M.; Newburger, P.E. Gene expression in mature neutrophils: Early responses to inflammatory stimuli. J. Leukoc. Biol. 2004, 75, 358–372. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.D.; Voyich, J.M.; Whitney, A.R.; DeLeo, F.R. Spontaneous neutrophil apoptosis and regulation of cell survival by granulocyte macrophage-colony stimulating factor. J. Leukoc. Biol. 2005, 78, 1408–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemiec, M.J.; Grumaz, C.; Ermert, D.; Desel, C.; Shankar, M.; Lopes, J.P.; Mills, I.G.; Stevens, P.; Sohn, K.; Urban, C.F. Dual transcriptome of the immediate neutrophil and Candida albicans interplay. BMC Genom. 2017, 18, 696. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Palaniyar, N. Transcriptional firing helps to drive NETosis. Sci. Rep. 2017, 7, 41749. [Google Scholar] [CrossRef] [PubMed]
- Tatsiy, O.; Mayer, T.Z.; de Carvalho Oliveira, V.; Sylvain-Prévost, S.; Isabel, M.; Dubois, C.M.; McDonald, P.P. Cytokine Production and NET Formation by Monosodium Urate-Activated Human Neutrophils Involves Early and Late Events, and Requires Upstream TAK1 and Syk. Front. Immunol. 2020, 10, 299. [Google Scholar] [CrossRef] [Green Version]
- Radom-Aizik, S.; Zaldivar, F.; Oliver, S.; Galassetti, P.; Cooper, D.M. Evidence for microRNA involvement in exercise-associated neutrophil gene expression changes. J. Appl. Physiol. 2010, 109, 252–261. [Google Scholar] [CrossRef]
- Khan, S.; Masood, M.; Gaur, H.; Ahmad, S.; Syed, M.A. Long non-coding RNA: An immune cells perspective. Life Sci. 2021, 271, 119152. [Google Scholar] [CrossRef]
- Kotzin, J.J.; Spencer, S.P.; McCright, S.J.; Kumar, D.B.U.; Collet, M.A.; Mowel, W.K.; Elliott, E.N.; Uyar, A.; Makiya, M.A.; Dunagin, M.C.; et al. The long non-coding RNA Morrbid regulates Bim and short-lived myeloid cell lifespan. Nature 2016, 537, 239–243. [Google Scholar] [CrossRef]
- Jiang, N.; Zhang, X.; He, Y.; Luo, B.; He, C.; Liang, Y.; Zeng, J.; Li, W.; Xian, Y.; Zheng, X. Identification of key protein-coding genes and lncRNAs in spontaneous neutrophil apoptosis. Sci. Rep. 2019, 9, 15106. [Google Scholar] [CrossRef] [Green Version]
- Forrest, O.A.; Ingersoll, S.A.; Preininger, M.K.; Laval, J.; Limoli, D.H.; Brown, M.R.; Lee, F.E.; Bedi, B.; Sadikot, R.T.; Goldberg, J.B.; et al. Frontline Science: Pathological conditioning of human neutrophils recruited to the airway milieu in cystic fibrosis. J. Leukoc. Biol. 2018, 104, 665–675. [Google Scholar] [CrossRef]
- Margaroli, C.; Moncada-Giraldo, D.; Gulick, D.A.; Dobosh, B.; Giacalone, V.D.; Forrest, O.A.; Sun, F.; Gu, C.; Gaggar, A.; Kissick, H.; et al. Transcriptional firing represses bactericidal activity in cystic fibrosis airway neutrophils. Cell Rep. Med. 2021, 100239. [Google Scholar] [CrossRef]
- Casanova-Acebes, M.; Nicolás-Ávila, J.A.; Yao Li, J.L.; García-Silva, S.; Balachander, A.; Rubio-Ponce, A.; Weiss, L.A.; Adrover, J.M.; Burrows, K.; A-González, N.; et al. Neutrophils instruct homeostatic and pathological states in naive tissues. J. Exp. Med. 2018, 215, 2778–2795. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.L.; Demers, M.; Martinod, K.; Gallant, M.; Wang, Y.; Goldfine, A.B.; Kahn, C.R.; Wagner, D.D. Diabetes primes neutrophils to undergo NETosis, which impairs wound healing. Nat. Med. 2015, 21, 815–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puga, I.; Cols, M.; Barra, C.M.; He, B.; Cassis, L.; Gentile, M.; Comerma, L.; Chorny, A.; Shan, M.; Xu, W.; et al. B cell-helper neutrophils stimulate the diversification and production of immunoglobulin in the marginal zone of the spleen. Nat. Immunol. 2011, 13, 170–180. [Google Scholar] [CrossRef] [Green Version]
- Nagelkerke, S.Q.; aan de Kerk, D.J.; Jansen, M.H.; van den Berg, T.K.; Kuijpers, T.W. Failure To Detect Functional Neutrophil B Helper Cells in the Human Spleen. PLoS ONE 2014, 9, e88377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker Harp, C.R.; Archambault, A.S.; Cheung, M.; Williams, J.W.; Czepielewski, R.S.; Duncker, P.C.; Kilgore, A.J.; Miller, A.T.; Segal, B.M.; Kim, A.H.J.J.; et al. Neutrophils promote VLA-4–dependent B cell antigen presentation and accumulation within the meninges during neuroinflammation. Proc. Natl. Acad. Sci. USA 2019, 116, 24221–24230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hertwig, L.; Pache, F.; Romero-Suarez, S.; Stürner, K.H.; Borisow, N.; Behrens, J.; Bellmann-Strobl, J.; Seeger, B.; Asselborn, N.; Ruprecht, K.; et al. Distinct functionality of neutrophils in multiple sclerosis and neuromyelitis optica. Mult. Scler. 2016, 22, 160–173. [Google Scholar] [CrossRef]
- Giordano, D.; Kuley, R.; Draves, K.E.; Roe, K.; Holder, U.; Giltiay, N.V.; Clark, E.A. BAFF Produced by Neutrophils and Dendritic Cells Is Regulated Differently and Has Distinct Roles in Antibody Responses and Protective Immunity against West Nile Virus. J. Immunol. 2020, 204, 1508–1520. [Google Scholar] [CrossRef]
- Stasulli, N.M.; Eichelberger, K.R.; Price, P.A.; Pechous, R.D.; Montgomery, S.A.; Parker, J.S.; Goldman, W.E. Spatially distinct neutrophil responses within the inflammatory lesions of pneumonic plague. MBio 2015, 6, e01530-15. [Google Scholar] [CrossRef] [Green Version]
- Gomez, J.C.; Dang, H.; Kanke, M.; Hagan, R.S.; Mock, J.R.; Kelada, S.N.P.; Sethupathy, P.; Doerschuk, C.M. Predicted effects of observed changes in the mRNA and microRNA transcriptome of lung neutrophils during S. pneumoniae pneumonia in mice. Sci. Rep. 2017, 7, 11258. [Google Scholar] [CrossRef]
- Ivan, F.X.; Tan, K.S.; Phoon, M.C.; Engelward, B.P.; Welsch, R.E.; Rajapakse, J.C.; Chow, V.T. Neutrophils infected with highly virulent influenza H3N2 virus exhibit augmented early cell death and rapid induction of type I interferon signaling pathways. Genomics 2013, 101, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Baßler, K.; Schlickeiser, S.; Zhang, B.; Krämer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L.; et al. Severe COVID-19 Is Marked by a Dysregulated Myeloid Cell Compartment. Cell 2020, 182, 1419–1440.e23. [Google Scholar] [CrossRef] [PubMed]
- Silvin, A.; Chapuis, N.; Dunsmore, G.; Goubet, A.; Dubuisson, A.; Derosa, L.; Almire, C.; Hénon, C.; Kosmider, O.; Droin, N.; et al. Elevated calprotectin and abnormal myeloid cell subsets discriminate severe from mild COVID-19. Cell 2020, 182, 1401–1418.e18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Sun, X.; Chen, Y.; Shen, Y.; Jarvis, J.N. RNA sequencing from human neutrophils reveals distinct transcriptional differences associated with chronic inflammatory states. BMC Med. Genom. 2015, 8, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Jiang, K.; Frank, M.B.; Chen, Y.; Jarvis, J.N. Complexity and Specificity of the Neutrophil Transcriptomes in Juvenile Idiopathic Arthritis. Sci. Rep. 2016, 6, 27453. [Google Scholar] [CrossRef] [Green Version]
- Yanaoka, H.; Nagafuchi, Y.; Hanata, N.; Takeshima, Y.; Ota, M.; Suwa, Y.; Shirai, H.; Sugimori, Y.; Okubo, M.; Kobayashi, S.; et al. Identifying the most influential gene expression profile in distinguishing ANCA-associated vasculitis from healthy controls. J. Autoimmun. 2021, 119, 102617. [Google Scholar] [CrossRef] [PubMed]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of Tumor-Associated Neutrophil Phenotype by TGF-β: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Fridlender, Z.G.; Sun, J.; Mishalian, I.; Singhal, S.; Cheng, G.; Kapoor, V.; Horng, W.; Fridlender, G.; Bayuh, R.; Worthen, G.S.; et al. Transcriptomic analysis comparing tumor-associated neutrophils with granulocytic myeloid-derived suppressor cells and normal neutrophils. PLoS ONE 2012, 7, e31524. [Google Scholar] [CrossRef] [PubMed]
- Shaul, M.E.; Levy, L.; Sun, J.; Mishalian, I.; Singhal, S.; Kapoor, V.; Horng, W.; Fridlender, G.; Albelda, S.M.; Fridlender, Z.G. Tumor-associated neutrophils display a distinct N1 profile following TGFβ modulation: A transcriptomics analysis of pro- vs. antitumor TANs. Oncoimmunology 2016, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Zhang, J.; Mao, Z.; Jiang, H.; Liu, W.; Shi, H.; Ji, R.; Xu, W.; Qian, H.; Zhang, X. Extracellular Vesicles From Gastric Cancer Cells Induce PD-L1 Expression on Neutrophils to Suppress T-Cell Immunity. Front. Oncol. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Zilionis, R.; Engblom, C.; Pfirschke, C.; Savova, V.; Zemmour, D.; Saatcioglu, H.D.; Krishnan, I.; Maroni, G.; Meyerovitz, C.V.; Kerwin, C.M.; et al. Single-Cell Transcriptomics of Human and Mouse Lung Cancers Reveals Conserved Myeloid Populations across Individuals and Species. Immunity 2019, 50, 1317–1334.e10. [Google Scholar] [CrossRef]
- Schupp, J.C.; Khanal, S.; Gomez, J.L.; Sauler, M.; Adams, T.S.; Chupp, G.L.; Yan, X.; Poli, S.; Zhao, Y.; Montgomery, R.R.; et al. Single-cell transcriptional archetypes of airway inflammation in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2020, 202, 1419–1429. [Google Scholar] [CrossRef]
- Subrahmanyam, Y.V.B.K.; Yamaga, S.; Prashar, Y.; Lee, H.H.; Hoe, N.P.; Kluger, Y.; Gerstein, M.; Goguen, J.D.; Newburger, P.E.; Weissman, S.M. RNA expression patterns change dramatically in human neutrophils exposed to bacteria. Blood 2001, 97, 2457–2468. [Google Scholar] [CrossRef] [PubMed]
- Malcolm, K.C.; Arndt, P.G.; Manos, E.J.; Jones, D.A.; Worthen, G.S. Microarray analysis of lipopolysaccharide-treated human neutrophils. Am. J. Physiol.—Lung Cell. Mol. Physiol. 2003, 284, 663–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukahara, Y.; Lian, Z.; Zhang, X.; Whitney, C.; Kluger, Y.; Tuck, D.; Yamaga, S.; Nakayama, Y.; Weissman, S.M.; Newburger, P.E. Gene expression in human neutrophils during activation and priming by bacterial lipopolysaccharide. J. Cell. Biochem. 2003, 89, 848–861. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, B.; Carlyon, J.A.; Cai, J.L.; Berliner, N.; Fikrig, E. Early transcriptional response of human neutrophils to Anaplasma phagocytophilum infection. Infect. Immun. 2005, 73, 8089–8099. [Google Scholar] [CrossRef] [Green Version]
- Águila, S.; de los Reyes-García, A.M.; Fernández-Pérez, M.P.; Reguilón-Gallego, L.; Zapata-Martínez, L.; Ruiz-Lorente, I.; Vicente, V.; González-Conejero, R.; Martínez, C. Micrornas as new regulators of neutrophil extracellular trap formation. Int. J. Mol. Sci. 2021, 22, 2116. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; Romero, R.; Varrey, A.; Leng, Y.; Miller, D.; Done, B.; Xu, Y.; Bhatti, G.; Motomura, K.; Gershater, M.; et al. RNA Sequencing Reveals Diverse Functions of Amniotic Fluid Neutrophils and Monocytes/Macrophages in Intra-Amniotic Infection. J. Innate Immun. 2021, 13, 63–82. [Google Scholar] [CrossRef]
- Fradin, C.; Mavor, A.L.; Weindl, G.; Schaller, M.; Hanke, K.; Kaufmann, S.H.E.; Mollenkopf, H.; Hube, B. The early transcriptional response of human granulocytes to infection with Candida albicans is not essential for killing but reflects cellular communications. Infect. Immun. 2007, 75, 1493–1501. [Google Scholar] [CrossRef] [Green Version]
- Camp, J.V.; Jonsson, C.B. A role for neutrophils in viral respiratory disease. Front. Immunol. 2017, 8, 550. [Google Scholar] [CrossRef] [Green Version]
- Hartshorn, K.L.; Liou, L.S.; White, M.R.; Kazhdan, M.M.; Tauber, J.L.; Tauber, A.I. Neutrophil deactivation by influenza A virus: Role of hemagglutinin binding to specific sialic acid-bearing cellular proteins. J. Immunol. 1995, 154, 3952–3960. [Google Scholar]
- Hufford, M.M.; Richardson, G.; Zhou, H.; Manicassamy, B.; García-Sastre, A.; Enelow, R.I.; Braciale, T.J. Influenza-Infected Neutrophils within the Infected Lungs Act as Antigen Presenting Cells for Anti-Viral CD8+ T Cells. PLoS ONE 2012, 7, e46581. [Google Scholar] [CrossRef]
- Zhu, B.; Zhang, R.; Li, C.; Jiang, L.; Xiang, M.; Ye, Z.; Kita, H.; Melnick, A.M.; Dent, A.L.; Sun, J. BCL6 modulates tissue neutrophil survival and exacerbates pulmonary inflammation following influenza virus infection. Proc. Natl. Acad. Sci. USA 2019, 116, 11888–11893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangalmurti, N.; Hunter, C.A. Cytokine Storms: Understanding COVID-19. Immunity 2020, 53, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Didangelos, A. COVID-19 Hyperinflammation: What about Neutrophils? mSphere 2020, 5, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Overmyer, K.A.; Shishkova, E.; Miller, I.J.; Balnis, J.; Bernstein, M.N.; Peters-Clarke, T.M.; Meyer, J.G.; Quan, Q.; Muehlbauer, L.K.; Trujillo, E.A.; et al. Large-Scale Multi-omic Analysis of COVID-19 Severity. Cell Syst. 2021, 12, 23–40.e7. [Google Scholar] [CrossRef]
- Plassmeyer, M.; Alpan, O.; Corley, M.J.; Premeaux, T.A.; Lillard, K.; Coatney, P.; Vaziri, T.; Michalsky, S.; Pang, A.P.S.; Bukhari, Z.; et al. Caspases and therapeutic potential of caspase inhibitors in moderate-severe SARS CoV2 infection and long COVID. Allergy 2021. [Google Scholar] [CrossRef]
- Wauters, E.; Van Mol, P.; Garg, A.D.; Jansen, S.; Van Herck, Y.; Vanderbeke, L.; Bassez, A.; Boeckx, B.; Malengier-Devlies, B.; Timmerman, A.; et al. Discriminating mild from critical COVID-19 by innate and adaptive immune single-cell profiling of bronchoalveolar lavages. Cell Res. 2021, 31, 272–290. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, H.K. Re-analysis of Single Cell Transcriptome Reveals That the NR3C1-CXCL8-Neutrophil Axis Determines the Severity of COVID-19. Front. Immunol. 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Shaath, H.; Vishnubalaji, R.; Elkord, E.; Alajez, N.M. Single-Cell Transcriptome Analysis Highlights a Role for Neutrophils and Inflammatory Macrophages in the Pathogenesis of Severe COVID-19. Cells 2020, 9, 2374. [Google Scholar] [CrossRef] [PubMed]
- Aschenbrenner, A.C.; Mouktaroudi, M.; Krämer, B.; Oestreich, M.; Antonakos, N.; Nuesch-Germano, M.; Gkizeli, K.; Bonaguro, L.; Reusch, N.; Baßler, K.; et al. Disease severity-specific neutrophil signatures in blood transcriptomes stratify COVID-19 patients. Genome Med. 2021, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Ko, T.M.; Chang, J.S.; Chen, S.P.; Liu, Y.M.; Chang, C.J.; Tsai, F.J.; Lee, Y.C.; Chen, C.H.; Chen, Y.T.; Wu, J.Y. Genome-wide transcriptome analysis to further understand neutrophil activation and lncRNA transcript profiles in Kawasaki disease. Sci. Rep. 2019, 9, 328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, K.; Poppenberg, K.E.; Wong, L.; Chen, Y.; Borowitz, D.; Goetz, D.; Sheehan, D.; Frederick, C.; Tutino, V.M.; Meng, H.; et al. RNA sequencing data from neutrophils of patients with cystic fibrosis reveals potential for developing biomarkers for pulmonary exacerbations. J. Cyst. Fibros. 2019, 18, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Geng, S.; Zhang, Y.; Lee, C.; Li, L. Novel reprogramming of neutrophils modulates inflammation resolution during atherosclerosis. Sci. Adv. 2019, 5, eaav2309. [Google Scholar] [CrossRef] [Green Version]
- Wright, H.L.; Thomas, H.B.; Moots, R.J.; Edwards, S.W. Interferon gene expression signature in rheumatoid arthritis neutrophils correlates with a good response to TNFi therapy. Rheumatology 2014, 54, 188–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirouvanziam, R.; Gernez, Y.; Conrad, C.K.; Moss, R.B.; Schrijver, I.; Dunn, C.E.; Davies, Z.A.; Herzenberg, L.A.L.A.; Herzenberg, L.A.L.A. Profound functional and signaling changes in viable inflammatory neutrophils homing to cystic fibrosis airways. Proc. Natl. Acad. Sci. USA 2008, 105, 4335–4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margaroli, C.; Garratt, L.W.; Horati, H.; Dittrich, A.S.; Rosenow, T.; Montgomery, S.T.; Frey, D.L.; Brown, M.R.; Schultz, C.; Guglani, L.; et al. Elastase Exocytosis by Airway Neutrophils Is Associated with Early Lung Damage in Children with Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2019, 199, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Sly, P.D.; Gangell, C.L.; Chen, L.; Ware, R.S.; Ranganathan, S.; Mott, L.S.; Murray, C.P.; Stick, S.M. Risk factors for bronchiectasis in children with cystic fibrosis. N. Engl. J. Med. 2013, 368, 1963–1970. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.; Giladi, A.; Gorki, A.-D.; Solodkin, D.G.; Zada, M.; Hladik, A.; Miklosi, A.; Salame, T.-M.; Halpern, K.B.; David, E.; et al. Lung Single-Cell Signaling Interaction Map Reveals Basophil Role in Macrophage Imprinting. Cell 2018, 175, 1031–1044.e18. [Google Scholar] [CrossRef] [Green Version]
- Byrne, A.J.; Powell, J.E.; O’Sullivan, B.J.; Ogger, P.P.; Hoffland, A.; Cook, J.; Bonner, K.L.; Hewitt, R.J.; Wolf, S.; Ghai, P.; et al. Dynamics of human monocytes and airway macrophages during healthy aging and after transplant. J. Exp. Med. 2020, 217, e20191236. [Google Scholar] [CrossRef]
- Forrest, O.A.; Chopyk, D.M.; Gernez, Y.; Brown, M.R.; Conrad, C.K.; Moss, R.B.; Tangpricha, V.; Peng, L.; Tirouvanziam, R. Resistin is elevated in cystic fibrosis sputum and correlates negatively with lung function. J. Cyst. Fibros. 2019, 18, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Lecot, P.; Sarabi, M.; Pereira Abrantes, M.; Mussard, J.; Koenderman, L.; Caux, C.; Bendriss-Vermare, N.; Michallet, M.-C. Neutrophil Heterogeneity in Cancer: From Biology to Therapies. Front. Immunol. 2019, 10, 2155. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, A.; Sharma, S.; Wu, K.; Wu, S.Y.; Xing, F.; Liu, Y.; Zhao, D.; Deshpande, R.P.; D’Agostino, R.B.; Watabe, K. Nicotine promotes breast cancer metastasis by stimulating N2 neutrophils and generating pre-metastatic niche in lung. Nat. Commun. 2021, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Turnis, M.E.; Sawant, D.V.; Szymczak-Workman, A.L.; Andrews, L.P.; Delgoffe, G.M.; Yano, H.; Beres, A.J.; Vogel, P.; Workman, C.J.; Vignali, D.A.A. Interleukin-35 Limits Anti-Tumor Immunity. Immunity 2016, 44, 316–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.-M.; Qin, J.; Li, Y.-C.; Wang, Y.; Li, D.; Shu, Y.; Luo, C.; Wang, S.-S.; Chi, G.; Guo, F.; et al. IL-35 induces N2 phenotype of neutrophils to promote tumor growth. Oncotarget 2017, 8, 33501–33514. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Hu, L.P.; Qin, W.T.; Yang, Q.; Chen, D.Y.; Li, Q.; Zhou, K.X.; Huang, P.Q.; Xu, C.J.; Li, J.; et al. Identification of a subset of immunosuppressive P2RX1-negative neutrophils in pancreatic cancer liver metastasis. Nat. Commun. 2021, 12, 174. [Google Scholar] [CrossRef] [PubMed]
- Alshetaiwi, H.; Pervolarakis, N.; McIntyre, L.L.; Ma, D.; Nguyen, Q.; Rath, J.A.; Nee, K.; Hernandez, G.; Evans, K.; Torosian, L.; et al. Defining the emergence of myeloid-derived suppressor cells in breast cancer using single-cell transcriptomics. Sci. Immunol. 2020, 5, eaay6017. [Google Scholar] [CrossRef]
- Veglia, F.; Hashimoto, A.; Dweep, H.; Sanseviero, E.; de Leo, A.; Tcyganov, E.; Kossenkov, A.; Mulligan, C.; Nam, B.; Masters, G.; et al. Analysis of classical neutrophils and polymorphonuclear myeloid-derived suppressor cells in cancer patients and tumor-bearing mice. J. Exp. Med. 2021, 218, e20201803. [Google Scholar] [CrossRef]
- Zhao, S. Alternative splicing, RNA-seq and drug discovery. Drug Discov. Today 2019, 24, 1258–1267. [Google Scholar] [CrossRef]
- Yang, X.; Kui, L.; Tang, M.; Li, D.; Wei, K.; Chen, W.; Miao, J.; Dong, Y. High-Throughput Transcriptome Profiling in Drug and Biomarker Discovery. Front. Genet. 2020, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.B.; Moots, R.J.; Edwards, S.W.; Wright, H.L. Whose gene is it anyway? the effect of preparation purity on neutrophil transcriptome studies. PLoS ONE 2015, 10, e0138982. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, E.M.; Pestonjamasp, K.N.; Bohl, B.P.; Chamberlain, C.; Hahn, K.M.; Bokoch, G.M. Spatial and temporal analysis of Rac activation during live neutrophil chemotaxis. Curr. Biol. 2002, 12, 2029–2034. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.L.; Ellis, B.A.; Munafo, D.B.; Brzezinska, A.A.; Catz, S.D. Gene transfer and expression in human neutrophils. The phox homology domain of p47phox translocates to the plasma membrane but not to the membrane of mature phagosomes. BMC Immunol. 2006, 7, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Biological Scenario | Analysis Methods | Neutrophil Gene Upregulation Observed | Citations |
|---|---|---|---|
| Diurnal Homeostasis | RNAseq | Fluctuations in Cry2, Arntl, Clock, Per1, Cxcr5, Vav2, Cxcl2, Icam1, Cxcr4, Sell, Cxcr2, Tlr4, IL17A, Csf3r, Mcl1, Fgr, Il1b, Il13ra1, Lmnb1, Atm, Dbp, Rev-erbα, CYBB | [43,44] |
| Aerobic Exercise | Microarray | GZMA, GZMH, PRF1, GZMB, HSPA1B, AREG, FYN, PDGFD, KSP37, SPON2, STAT4, TGFBR3, LDLR, SOCS2, RUNX3, KLRC4, ZAP70, SLAMf7, CTSW, GNLY | [48] |
| Small pore migration | RNAseq | EGR3, SERPINE1, CCL3, CCL4, EGR2, EGR1, TNF, FOSB, CCL4L2, CXCL8, BCL3, IER3, NFKBIA, TREM1, FOS, BTG2, NAB2, NR4A2, NFKBID, DUSP2, CD69 | [49] |
| Chemoattractant Priming (TNFα/GM-CSF/fMLF/LPS stimulation) | Microarray, RNAseq | CCL3, CCL4, CD69, CISH, CXCL1, CXCL2, DUSP2, EDN1, EGR1, EGR2, GADD45B, GPR84, HBEGF, HCAR2, HCAR3, HRH4, ICAM1, IL1A, IL1B, IL1RN, KCNJ2, MFSD2A, NFKBIA, NFKBIE, OLR1, PDE4B, PLAU, PNPLA1, PPP1R15A, RHOH, SLC35B2, SOCS3, TARP, TIFA, TNF, TNFAIP3, TNFAIP6, TRAF1, ZFP36, BIRC3, CCR1, IL3RA, C3AR1, CD83, CYBB | [50,51,52] |
| Microbial killing (bacteria or yeast) | Microarray, RNAseq | NR4A3, OLR1, TRAF1, CCRL2, PLAU, JMY, HS3STB1, LIF, IL1A, CXCL2, CREM, IRAK2, NR1D1, SFMBT2, SAMSN1, PNP, EIF2AK3, ZNF331, FRMD4B, TGIF1, VEGFA, EGR1, ILR1, C3AR1, C5AR1, TREM1, NLRP3, IRAK2, TICAM1, BFAR, CXCL1, BIRC3, IL1B, CXCL8, RIPK2, ADORA2A, GPR65, MAPK6, DUSP2, PTPRE, ICAM1, PLAUR, ACTG, TRIF, CD83, OSR1, TNF | [51,53] |
| NETosis | Microarray | GLA, RP11, ANXA1, EGR1, MAP4K5, SEMA7A, C3AR1, CYBB, H3F3C, IL18RAP, KLF2, IL1B, JUNB, MAP3K8, TNF, CCL4, CXCL8 | [54,55] |
| Disease | Analysis Methods | Neutrophil Gene Expression Observed | Citations |
|---|---|---|---|
| Bacterial/Fungal infection | RNAseq | Cxcl9, Gbp5, Ifit2, Cxcl11, Ifi205, Il1a, Ikbke, Csf3, Cxcl10, Ifnar1, Ifnar2, Tlr9, Il12a, Mx2, Ccl4, Il10, Nlrc5, Il1f6, Gbp3, Tnfrsf9, Phynin1, IL15ra, IL6, Ccl2, Tnf, Nos2, Cln3, Zfp64, Fundc2, Brix1, Slfn3, Yipf6, Dlg1, Smek1, Prr12, 2410016O06Rik, Ebna1bp2, Cdc37l1, Impa1, Gclc, Atp2c1, Las1l, Ahi1, Zfp192, Nt5e, Piga | [69,70] |
| Viral Infection | RNAseq, scRNAseq | IFNβ1, ISG15, IRF1, CXCL10, CASP4, NOD1, PARP14, EIF2AK2, XAF1, PARP9, PARP10, BCL6, Krt18, Prf1, IFITM1/3, RSAD2, CD274 (PD-L1), ZC3H12A, CD177, ARG1, S100A8, S100A9, CXCR4, SELL, SPI1 | [71,72,73] |
| Chronic Inflammation/Auto-immune | RNAseq | PAM, ADARB2, C5orf56, ICAM1, IL1B, CCR1, IFIH1, SOCS1, TNFAIP3, TNFSF13B, OAS1, OAS2, IFI35, IFI44, IFI44L, IFI6, IFIT2, IFIT3, IFIT5, IFITM1/3, APOBEC3B, AASS, ELF5, COL4A3, ZNF772, RNU5A-1, CEACAM19, OAS3, PGM5, TIAF1, LY6E, LILRB5, FAM21B, TBC1D15, HMGN3, PRKAR2B, PLXNB2, APMAP, PGM1, ACAP1, PYGL, S100A12, SRPK1, ACSL1, CLEC4D, MAP4K4, MAPK14, ACTB, PXK, TP53I11, ZDHHC3, BMX, PGD, SLC37A3, SLC26A8, ALOX5, KIF1B, PLP2, S100A9 | [74,75,76] |
| Cancer | Microarray, RNAseq, scRNAseq | CD206, CCL2, ARG2, Nos2, CALR, LGMN, CXCL13, CXCL10, TNF, CASS4, CTSC, CCL3, CD274 (PD-L1), TLR1, TLR4, IRAK2, IEX1, SOD2, GADD45b, BCL2A1, CD74, Cd1d-1, Psme1, H2-DMa, DMb1, Eb1, CCL17, CXCL9, CXCL16, CXCL1, CXCL2, IL1a, IL1b, IL12 | [77,78,79,80,81] |
| Cystic Fibrosis | RNAseq, scRNAseq | FAM107A, SPOCK2, GZMH, GPRC5B, EDG1, TMEM35, A_24_P920715, AGT, ACVR1C, HBS1L, TRBV5-4, HBA2, PKN1, AEBP1, EPB41L2, GFAP, HBB, RNASEN, AK090499, HBA1, CXCR4, CD83, IRAK2, IRAK3, TRAF3, PLAU, CREM, HES4, KRAS, TRPM7, CCL3, CCL4, TLR2, IGF2R, KLRG1, CCR5, C4A, CCR4, CFD, SPP1, IFITM2, MSH2, FCGR2C, TREM1, CFD, CDKN1A, PLK3, ZC3HC1, MSH2, CYP26B1, KIF20B, CENPV, G0S2, CDK6, AHR, MYH6, PPP4R4, AIM2, NLRP3, S100A12 | [59,61,74,82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garratt, L.W. Current Understanding of the Neutrophil Transcriptome in Health and Disease. Cells 2021, 10, 2406. https://doi.org/10.3390/cells10092406
Garratt LW. Current Understanding of the Neutrophil Transcriptome in Health and Disease. Cells. 2021; 10(9):2406. https://doi.org/10.3390/cells10092406
Chicago/Turabian StyleGarratt, Luke W. 2021. "Current Understanding of the Neutrophil Transcriptome in Health and Disease" Cells 10, no. 9: 2406. https://doi.org/10.3390/cells10092406
APA StyleGarratt, L. W. (2021). Current Understanding of the Neutrophil Transcriptome in Health and Disease. Cells, 10(9), 2406. https://doi.org/10.3390/cells10092406