
Analysis of TCR Repertoire by High-Throughput Sequencing Indicates the Feature of T Cell Immune Response after SARS-CoV-2 Infection

 ,
,
Abstract
:1. Introduction
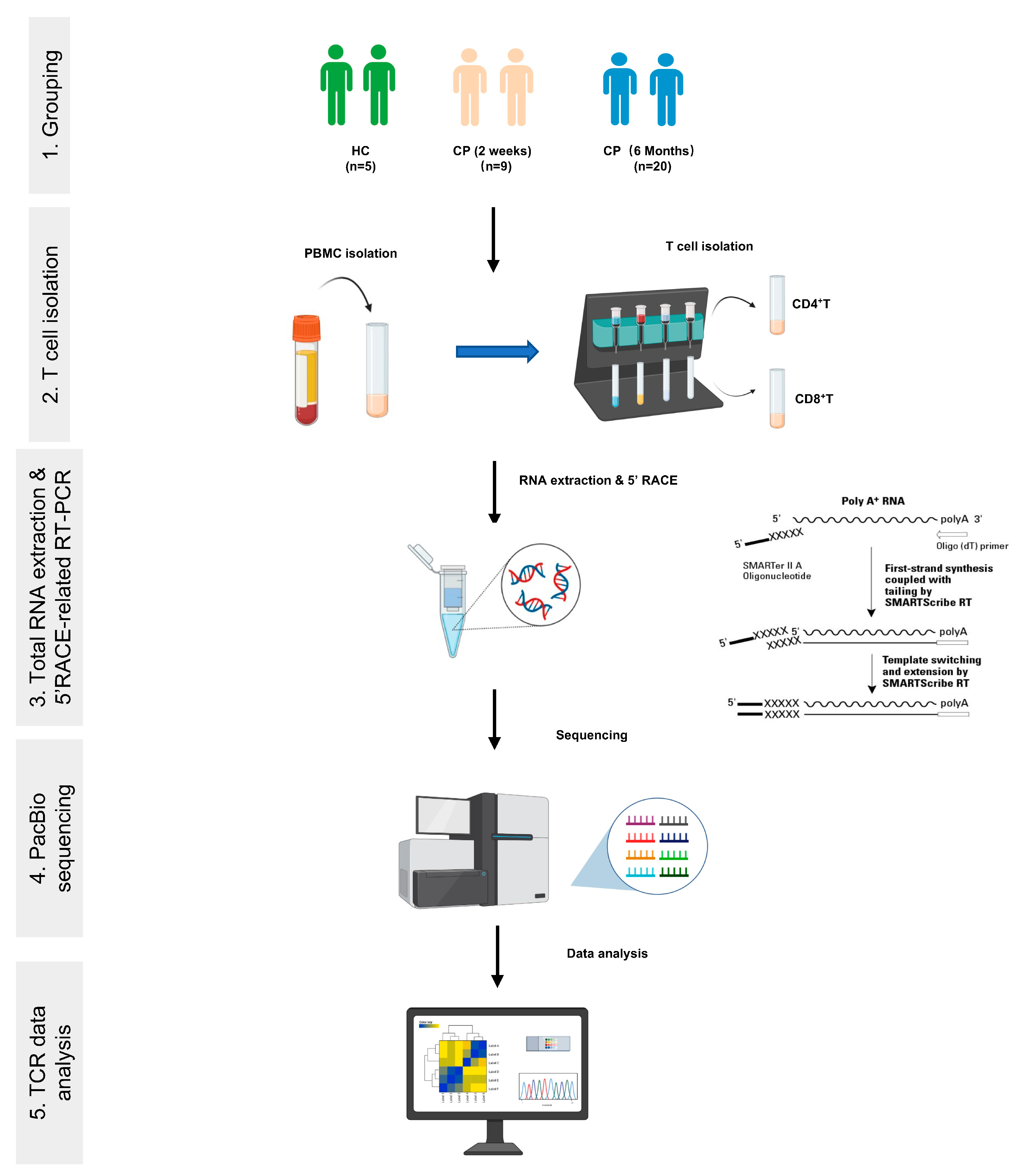
2. Materials and Methods
2.1. Donors and Blood Samples
2.2. Isolation of PBMCs and T Cell Subpopulations
2.3. TCRα and TCRβ Library Preparation and PCR Amplification
2.4. TCRα and TCRβ Sequencing and Data Preprocessing
2.5. TCR Repertoire Analysis
3. Results
3.1. Clinical Characteristics of COVID-19 Patients with Different Convalescent Stages for TCR Repertoire Profiling Analysis
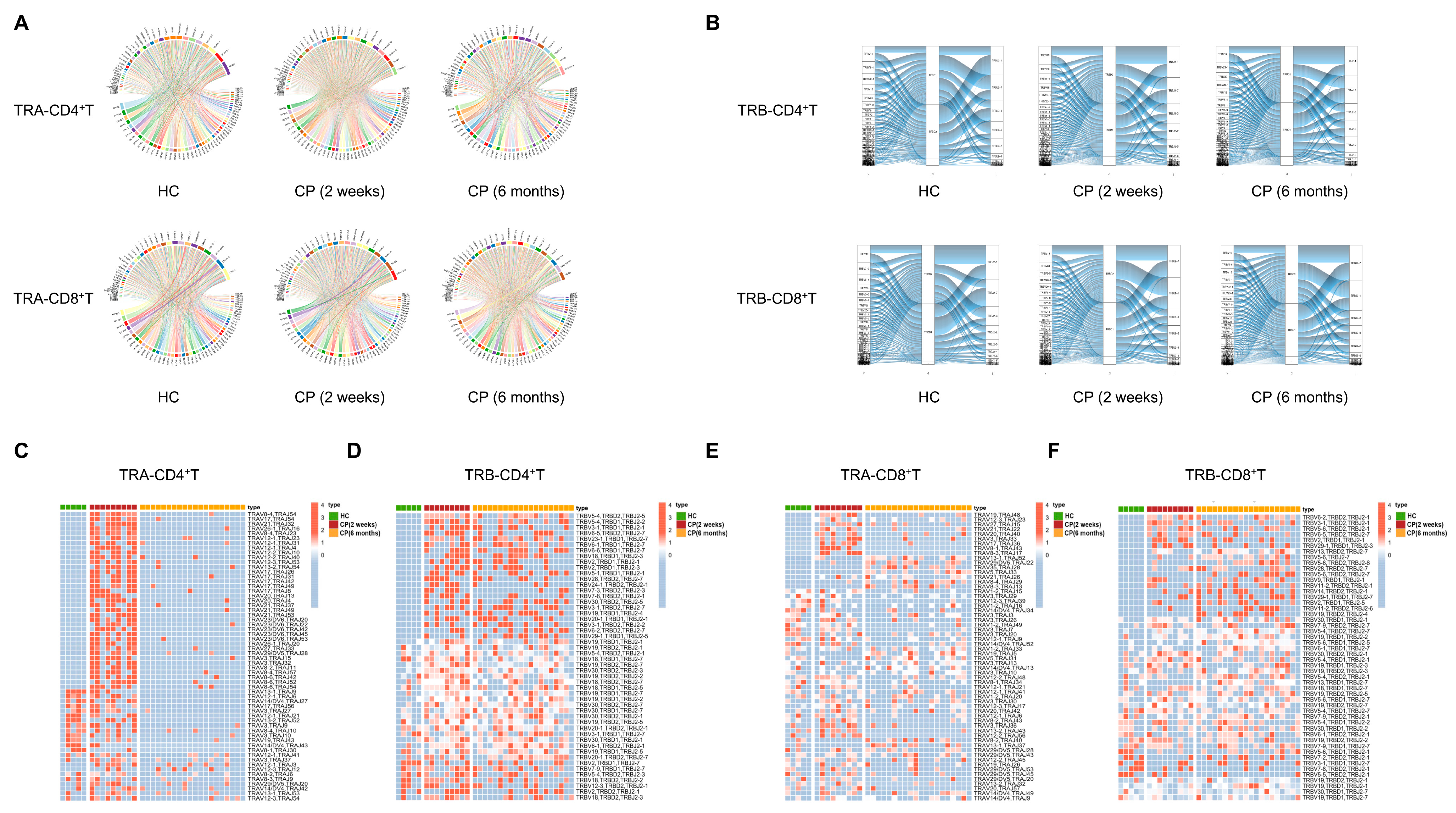
3.2. Significance of T Cell Receptor Bias during the Different Convalescent Phases of COVID-19 Patients Compared with Healthy Controls
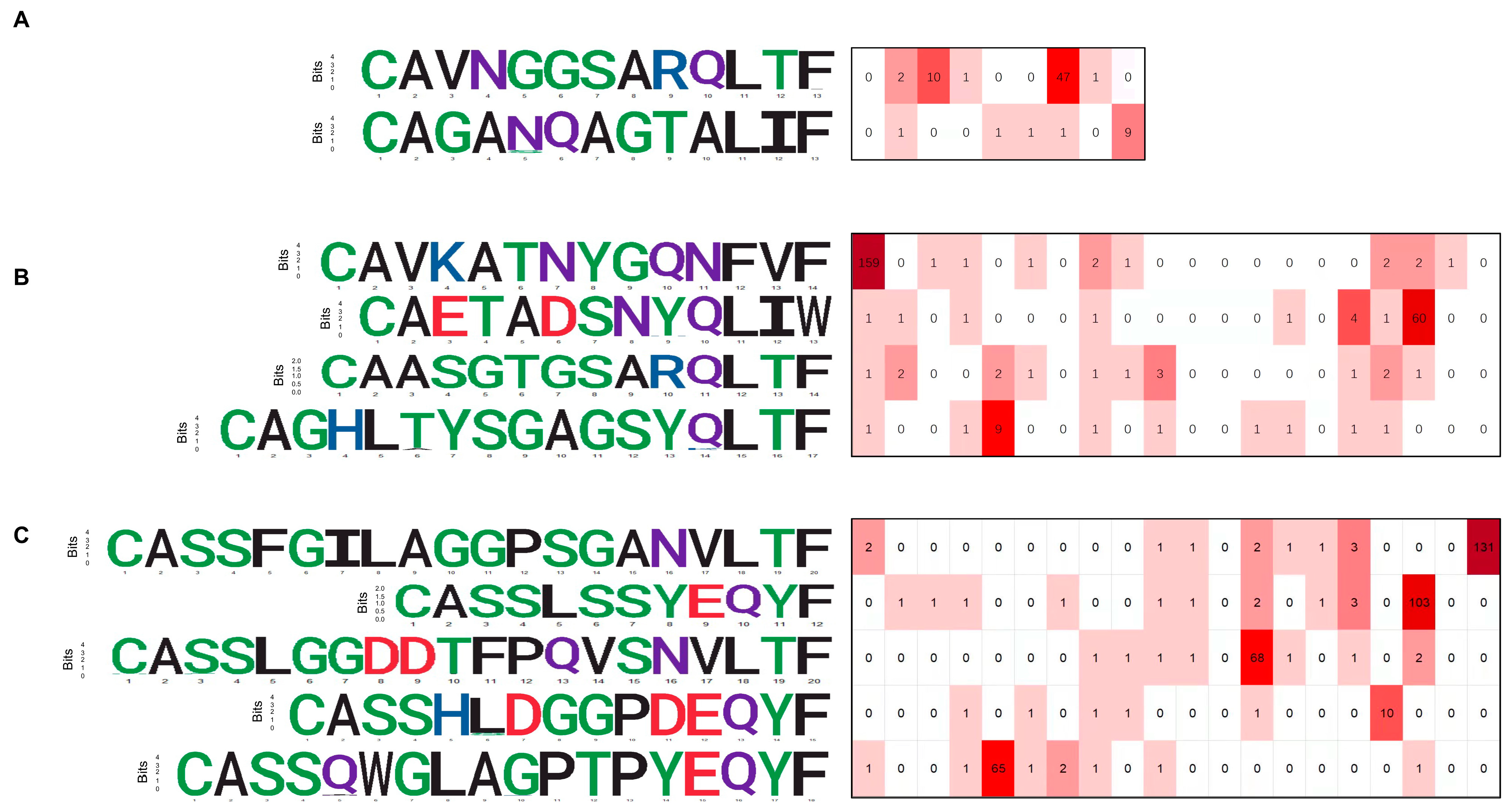
3.3. CDR3 Sequence Motifs of Responding Clones
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons from the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72 314 Cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Shomuradova, A.S.; Vagida, M.S.; Sheetikov, S.A.; Zornikova, K.V.; Kiryukhin, D.; Titov, A.; Peshkova, I.O.; Khmelevskaya, A.; Dianov, D.V.; Malasheva, M.; et al. SARS-CoV-2 Epitopes Are Recognized by a Public and Diverse Repertoire of Human T Cell Receptors. Immunity 2020, 53, 1245–1257.e5. [Google Scholar] [CrossRef] [PubMed]
- Nelde, A.; Bilich, T.; Heitmann, J.S.; Maringer, Y.; Salih, H.R.; Roerden, M.; Lübke, M.; Bauer, J.; Rieth, J.; Wacker, M.; et al. SARS-CoV-2-derived peptides define heterologous and COVID-19-induced T cell recognition. Nat. Immunol. 2021, 22, 74–85. [Google Scholar] [CrossRef]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Rosendahl Huber, S.; van Beek, J.; de Jonge, J.; Luytjes, W.; van Baarle, D. T cell responses to viral infections-opportunities for Peptide vaccination. Front. Immunol. 2014, 5, 171. [Google Scholar] [CrossRef] [PubMed]
- Rouse, B.T.; Sehrawat, S. Immunity and immunopathology to viruses: What decides the outcome? Nat. Rev. Immunol. 2010, 10, 514–526. [Google Scholar] [CrossRef]
- Altmann, D.M.; Boyton, R.J. SARS-CoV-2 T cell immunity: Specificity, function, durability, and role in protection. Sci. Immunol. 2020, 5, eabd6160. [Google Scholar] [CrossRef] [PubMed]
- Snyder, T.M.; Gittelman, R.M.; Klinger, M.; May, D.H.; Osborne, E.J.; Taniguchi, R.; Zahid, H.J.; Kaplan, I.M.; Dines, J.N.; Noakes, M.N.; et al. Magnitude and Dynamics of the T-Cell Response to SARS-CoV-2 Infection at Both Individual and Population Levels. medRxiv 2020, 20165647. [Google Scholar] [CrossRef]
- Thevarajan, I.; Nguyen, T.H.O.; Koutsakos, M.; Druce, J.; Caly, L.; van de Sandt, C.E.; Jia, X.; Nicholson, S.; Catton, M.; Cowie, B.; et al. Breadth of concomitant immune responses prior to patient recovery: A case report of non-severe COVID-19. Nat. Med. 2020, 26, 453–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Chen, D.; Yuan, D.; Lausted, C.; Choi, J.; Dai, C.L.; Voillet, V.; Duvvuri, V.R.; Scherler, K.; Troisch, P.; et al. Multi-Omics Resolves a Sharp Disease-State Shift between Mild and Moderate COVID-19. Cell 2020, 183, 1479–1495.e20. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Mentzer, A.J.; Liu, G.; Yao, X.; Yin, Z.; Dong, D.; Dejnirattisai, W.; Rostron, T.; Supasa, P.; Liu, C.; et al. Broad and strong memory CD4(+) and CD8(+) T cells induced by SARS-CoV-2 in UK convalescent individuals following COVID-19. Nat. Immunol. 2020, 21, 1336–1345. [Google Scholar] [CrossRef]
- Notarbartolo, S.; Ranzani, V.; Bandera, A.; Gruarin, P.; Bevilacqua, V.; Putignano, A.R.; Gobbini, A.; Galeota, E.; Manara, C.; Bombaci, M.; et al. Integrated longitudinal immunophenotypic, transcriptional and repertoire analyses delineate immune responses in COVID-19 patients. Sci. Immunol. 2021, 6, eabg5021. [Google Scholar] [CrossRef] [PubMed]
- Neidleman, J.; Luo, X.; Frouard, J.; Xie, G.; Gill, G.; Stein, E.S.; McGregor, M.; Ma, T.; George, A.F.; Kosters, A.; et al. SARS-CoV-2-Specific T Cells Exhibit Phenotypic Features of Helper Function, Lack of Terminal Differentiation, and High Proliferation Potential. Cell Rep. Med. 2020, 1, 100081. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Zhang, S.; Mao, Y.; Huang, J.; Ma, T.; Zhang, L.; Zhu, X.; Zheng, J.; Wu, L.; Yin, C.C.; Qiu, X. Immunoglobulin gene locus events in epithelial cells of lactating mouse mammary glands. Cell. Mol. Life Sci. 2010, 67, 985–994. [Google Scholar] [CrossRef]
- Dash, P.; Fiore-Gartland, A.J.; Hertz, T.; Wang, G.C.; Sharma, S.; Souquette, A.; Crawford, J.C.; Clemens, E.B.; Nguyen, T.H.O.; Kedzierska, K.; et al. Quantifiable predictive features define epitope-specific T cell receptor repertoires. Nature 2017, 547, 89–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furman, D.; Davis, M.M. New approaches to understanding the immune response to vaccination and infection. Vaccine 2015, 33, 5271–5281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Sanders, C.M.; Yang, Q.; Schroeder, H.W., Jr.; Wang, E.; Babrzadeh, F.; Gharizadeh, B.; Myers, R.M.; Hudson, J.R., Jr.; Davis, R.W.; et al. High throughput sequencing reveals a complex pattern of dynamic interrelationships among human T cell subsets. Proc. Natl. Acad. Sci. USA 2010, 107, 1518–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultheiß, C.; Paschold, L.; Simnica, D.; Mohme, M.; Willscher, E.; von Wenserski, L.; Scholz, R.; Wieters, I.; Dahlke, C.; Tolosa, E.; et al. Next-Generation Sequencing of T and B Cell Receptor Repertoires from COVID-19 Patients Showed Signatures Associated with Severity of Disease. Immunity 2020, 53, 442–455.e4. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Gan, R.; Zhen, Z.; Hu, X.; Li, X.; Zhou, F.; Liu, Y.; Chen, C.; Xie, S.; Zhang, B.; et al. Adaptive immune responses to SARS-CoV-2 infection in severe versus mild individuals. Signal Transduct. Target. Ther. 2020, 5, 156. [Google Scholar] [CrossRef]
- Wang, P.; Jin, X.; Zhou, W.; Luo, M.; Xu, Z.; Xu, C.; Li, Y.; Ma, K.; Cao, H.; Huang, Y.; et al. Comprehensive analysis of TCR repertoire in COVID-19 using single cell sequencing. Genomics 2020, 113, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Minervina, A.; Pogorelyy, M.; Mamedov, I. T-cell receptor and B-cell receptor repertoire profiling in adaptive immunity. Transpl. Int. 2019, 32, 1111–1123. [Google Scholar] [CrossRef]
- Rosati, E.; Dowds, C.M.; Liaskou, E.; Henriksen, E.K.K.; Karlsen, T.H.; Franke, A. Overview of methodologies for T-cell receptor repertoire analysis. BMC Biotechnol. 2017, 17, 61. [Google Scholar] [CrossRef]
- Lin, Y.H.; Hung, S.J.; Chen, Y.L.; Lin, C.H.; Kung, T.F.; Yeh, Y.C.; Tseng, J.T.; Liu, T. Dissecting efficiency of a 5′ rapid amplification of cDNA ends (5′-RACE) approach for profiling T-cell receptor beta repertoire. PLoS ONE 2020, 15, e0236366. [Google Scholar] [CrossRef]
- Yeku, O.; Frohman, M.A. Rapid amplification of cDNA ends (RACE). Methods Mol. Biol. 2011, 703, 107–122. [Google Scholar] [PubMed]
- Nishana, M.; Raghavan, S.C. Role of recombination activating genes in the generation of antigen receptor diversity and beyond. Immunology 2012, 137, 271–281. [Google Scholar] [CrossRef]
- Andersen, M.H.; Schrama, D.; thor Straten, P.; Becker, J.C. Cytotoxic T Cells. J. Investig. Dermatol. 2006, 126, 32–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sant, A.J.; McMichael, A. Revealing the role of CD4(+) T cells in viral immunity. J. Exp. Med. 2012, 209, 1391–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Gruta, N.L.; Gras, S.; Daley, S.R.; Thomas, P.G.; Rossjohn, J. Understanding the drivers of MHC restriction of T cell receptors. Nat. Rev. Immunol. 2018, 18, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Perez-Potti, A.; Rivera-Ballesteros, O.; Strålin, K.; Gorin, J.B.; Olsson, A.; Llewellyn-Lacey, S.; Kamal, H.; Bogdanovic, G.; Muschiol, S.; et al. Robust T Cell Immunity in Convalescent Individuals with Asymptomatic or Mild COVID-19. Cell 2020, 183, 158–168.e14. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC (n = 5) | CP (2 Weeks) (n = 9) | |||
|---|---|---|---|---|
| Severity | Healthy (n = 5) | Moderate (n = 3) | Severe (n = 3) | Critical (n = 3) |
| Age (years) | 65.4 ± 8.76 | 58 ± 12.78 | 67 ± 9.9 | 73 ± 5.57 |
| Sex | ||||
| Male | 2 | 1 | 1 | 2 |
| Female | 3 | 2 | 2 | 1 |
| White blood cells (109/L) | 5.83 ± 1.2 | 4.97 ± 1.35 | 7.76 ± 4.87 | 7.2 ± 3.68 |
| Percentage of neutrophil | 60.48 ± 6 | 59.3 ± 11.47 | 79.3 ± 2.4 | 90 ± 6.9 |
| Number of neutrophils (109/L) | 3.22 ± 0.6 | 3.1 ± 1.27 | 6.11 ± 3.68 | 7.29 ± 3.7 |
| Percentage of lymphocytes | 20.5 ± 10.2 | 32.7 ± 11.46 | 16.4 ± 0.7 | 5.97 ± 4.5 |
| Number of lymphocytes (109/L) | 1.6 ± 0.92 | 1.49 ± 0.23 | 1.26 ± 0.74 | 0.36 ± 0.04 |
| Percentage of monocytes | 6.27 ± 0.84 | 6.48 ± 0.45 | 4 ± 3.39 | 2.85 ± 2.69 |
| Number of monocytes (109/L) | 0.28 ± 0.03 | 0.32 ± 0.08 | 0.4 ± 0.46 | 0.24 ± 0.03 |
| CRP (mg/L) | 1.91 ± 2.4 | 2.89 ± 1.1 | 8.49 ± 10.8 | 44.89 ± 39 |
| CK (U/L) | 25 ± 5.4 | 39.8 ± 16.46 | 35.5 ± 14.8 | 133.7 ± 100 |
| Alkalosis | N | N | Y (2) | Y (3) |
| Complication | ||||
| ARDS | N | N | Y (2) | Y (3) |
| Secondary infections | N | N | N | N |
| History of smoking | N | N | N | N |
| CD4+ T | CD8+ T | Total | |||||
|---|---|---|---|---|---|---|---|
| HC | CP (2 Weeks) | CP (6 Months) | HC | CP (2 Weeks) | CP (6 Months) | ||
| TRA | 3372 | 7298 | 654 | 4125 | 6670 | 10,677 | 32,796 |
| TRB | 951 | 2066 | 3916 | 805 | 1820 | 5528 | 15,086 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Duan, F.; Zhu, Z.; Yu, M.; Jia, X.; Dai, H.; Wang, P.; Qiu, X.; Lu, Y.; Huang, J. Analysis of TCR Repertoire by High-Throughput Sequencing Indicates the Feature of T Cell Immune Response after SARS-CoV-2 Infection. Cells 2022, 11, 68. https://doi.org/10.3390/cells11010068
Wang Y, Duan F, Zhu Z, Yu M, Jia X, Dai H, Wang P, Qiu X, Lu Y, Huang J. Analysis of TCR Repertoire by High-Throughput Sequencing Indicates the Feature of T Cell Immune Response after SARS-CoV-2 Infection. Cells. 2022; 11(1):68. https://doi.org/10.3390/cells11010068
Chicago/Turabian StyleWang, Yifan, Fugang Duan, Zhu Zhu, Meng Yu, Xiaodong Jia, Hui Dai, Pingzhang Wang, Xiaoyan Qiu, Yinying Lu, and Jing Huang. 2022. "Analysis of TCR Repertoire by High-Throughput Sequencing Indicates the Feature of T Cell Immune Response after SARS-CoV-2 Infection" Cells 11, no. 1: 68. https://doi.org/10.3390/cells11010068
APA StyleWang, Y., Duan, F., Zhu, Z., Yu, M., Jia, X., Dai, H., Wang, P., Qiu, X., Lu, Y., & Huang, J. (2022). Analysis of TCR Repertoire by High-Throughput Sequencing Indicates the Feature of T Cell Immune Response after SARS-CoV-2 Infection. Cells, 11(1), 68. https://doi.org/10.3390/cells11010068