
Axl Expression in Renal Mesangial Cells Is Regulated by Sp1, Ap1, MZF1, and Ep300, and the IL-6/miR-34a Pathway


Abstract
:
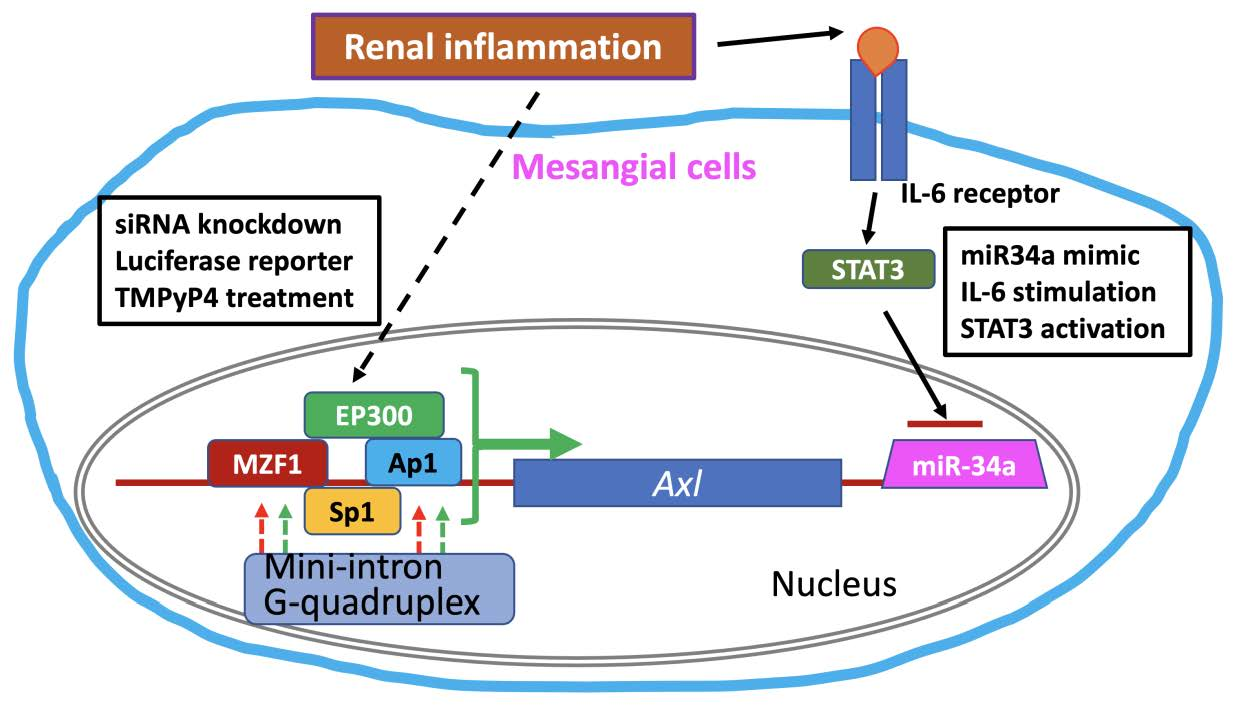{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents
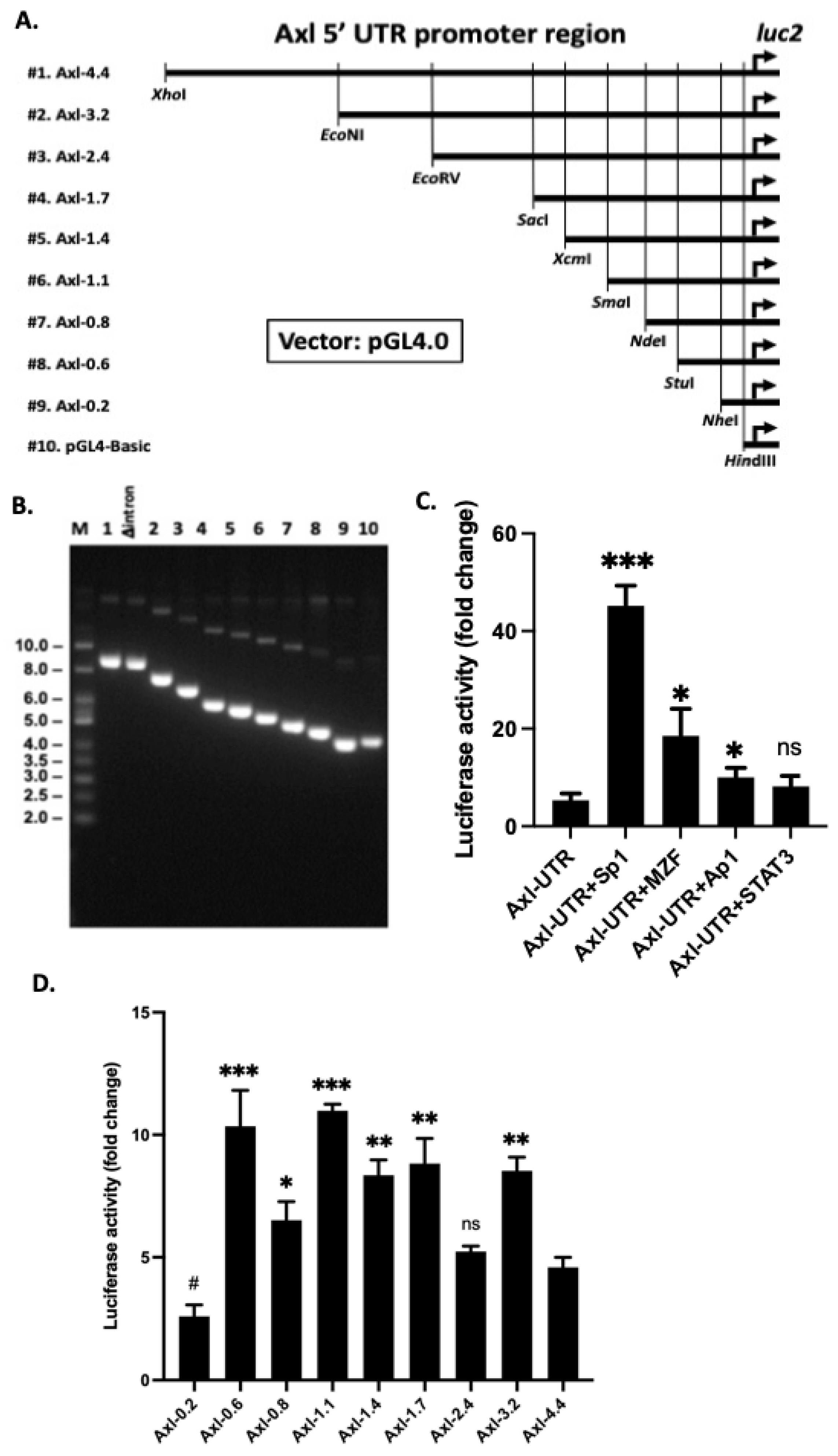
2.2. Analysis of the Axl Promoter and 5′-UTR
2.3. SiRNA Mediated TF Knockdown and Western Blot Analysis
2.4. Generation of Luciferase Reporter Constructs
2.5. Luciferase Assays in Hela Cells
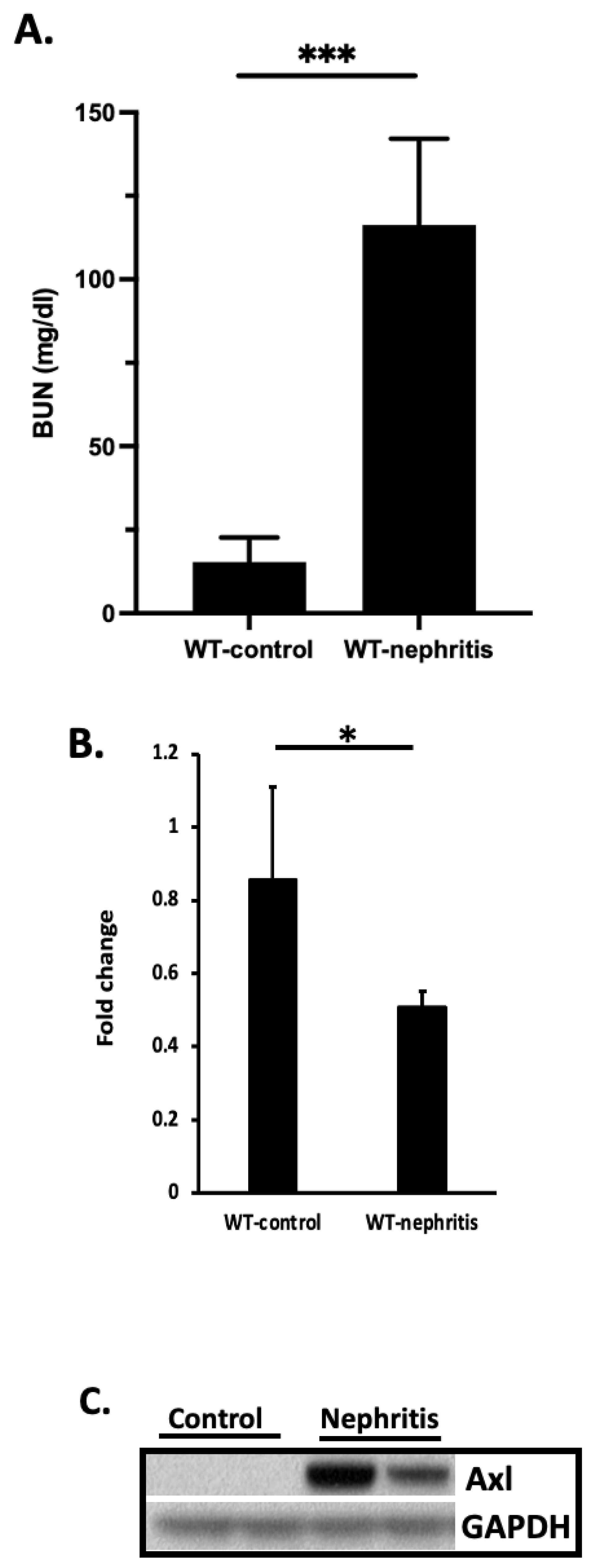
2.6. MicroRNA-34a (miR-34a) Analysis in the Kidney of Nephritic Mice and in Cell Cultures
2.7. IL-6 Treatment
2.8. Statistical Analysis
3. Results
3.1. Multiple TFs Regulate Axl Expression in Mesangial Cells
3.2. Proximal 600 bp Is Sufficient for Basal Axl Expression
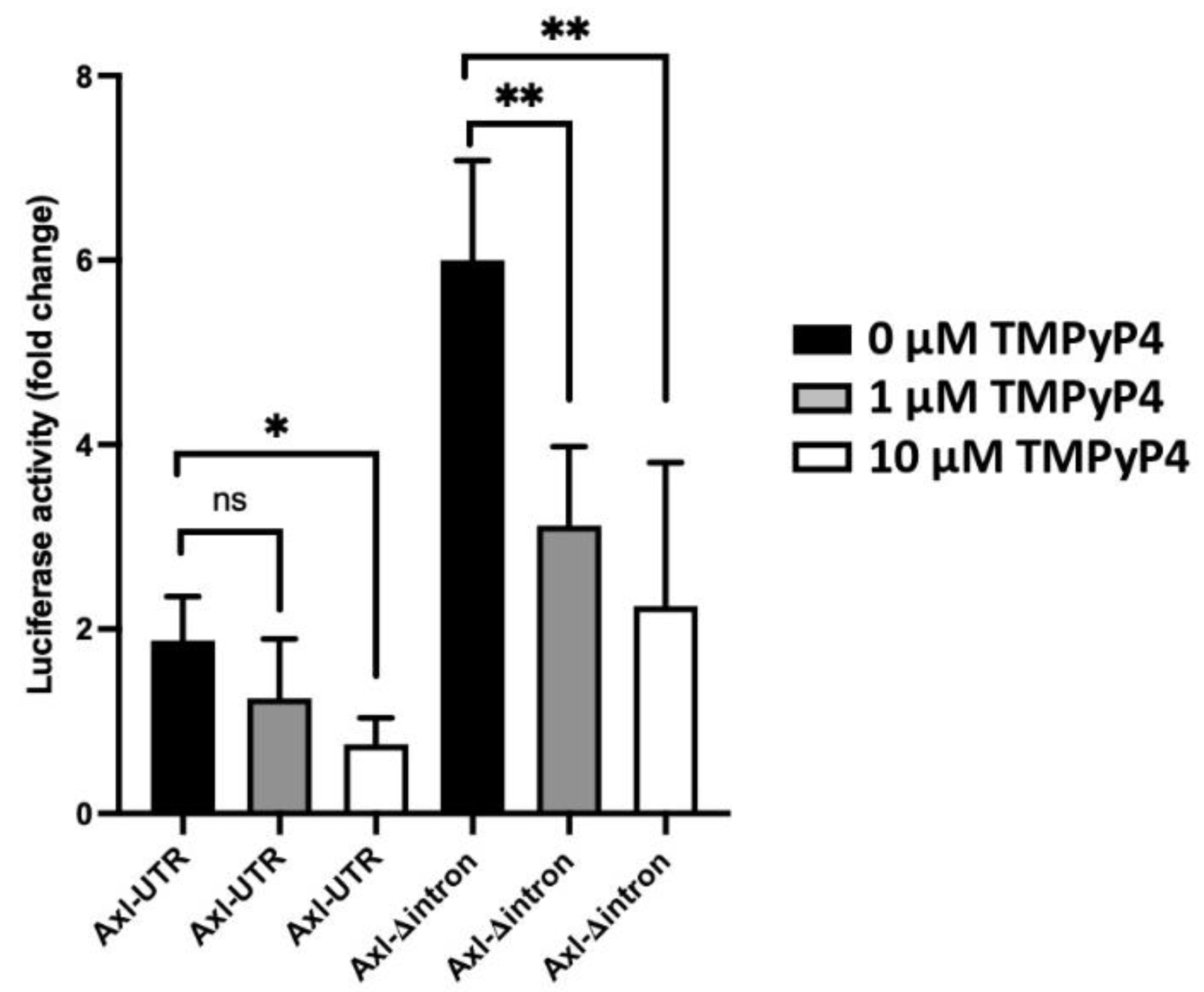
3.3. An Inhibitory Role of a Mini-Intron in the 5′-UTR of the Axl Promoter Region
3.4. Transcriptional and Translational Inhibition of Axl by the G-Quadruplex Structure
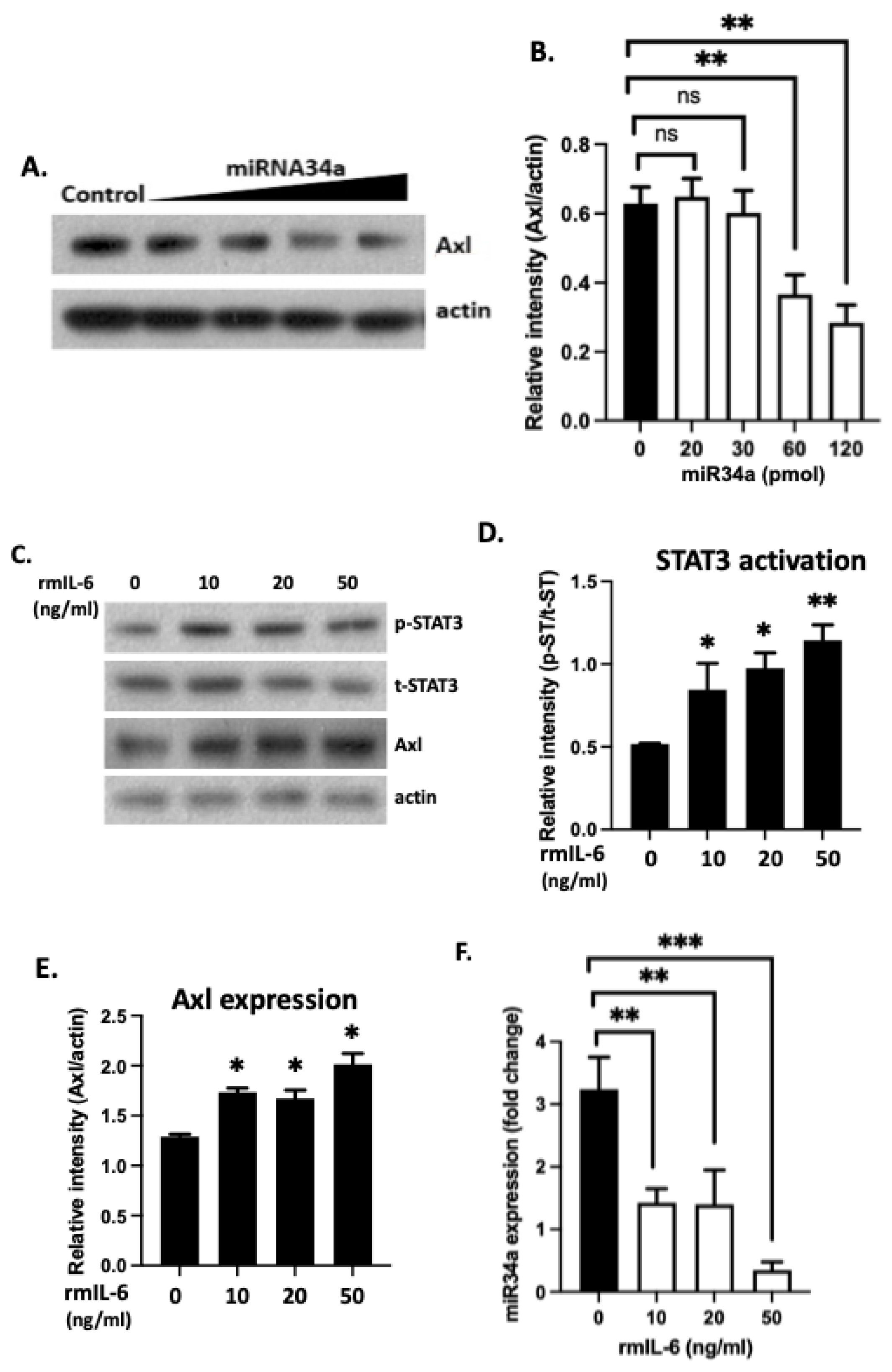
3.5. 3′-UTR Binding by miRNA Inhibits Axl Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Cohen, P.L.; Shao, W.H. Gas6/TAM receptors in systemic lupus erythematosus. Dis. Markers 2019, 2019, 7838195. [Google Scholar] [CrossRef] [PubMed]
- Lemke, G. Biology of the TAM receptors. Cold Spring Harb. Perspect. Biol. 2013, 5, a009076. [Google Scholar] [CrossRef] [PubMed]
- Linger, R.M.; Keating, A.K.; Earp, H.S.; Graham, D.K. TAM receptor tyrosine kinases: Biologic functions, signaling, and potential therapeutic targeting in human cancer. Adv. Cancer Res. 2008, 100, 35–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagita, M.; Ishimoto, Y.; Arai, H.; Nagai, K.; Ito, T.; Nakano, T.; Salant, D.J.; Fukatsu, A.; Doi, T.; Kita, T. Essential role of Gas6 for glomerular injury in nephrotoxic nephritis. J. Clin. Investig. 2002, 110, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.; Lee, I.J.; Finkelman, F.D.; Shao, W.H. Targeted inhibition of Axl receptor tyrosine kinase ameliorates anti-GBM-induced lupus-like nephritis. J. Autoimmun. 2018, 93, 37–44. [Google Scholar] [CrossRef]
- Zhen, Y.; Priest, S.O.; Shao, W.H. Opposing roles of tyrosine kinase receptors Mer and Axl determine clinical outcomes in experimental immune-mediated nephritis. J. Immunol. 2016, 197, 2187–2194. [Google Scholar] [CrossRef] [Green Version]
- Giles, K.M.; Kalinowski, F.C.; Candy, P.A.; Epis, M.R.; Zhang, P.M.; Redfern, A.D.; Stuart, L.M.; Goodall, G.J.; Leedman, P.J. Axl mediates acquired resistance of head and neck cancer cells to the epidermal growth factor receptor inhibitor erlotinib. Mol. Cancer Ther. 2013, 12, 2541–2558. [Google Scholar] [CrossRef] [Green Version]
- Mudduluru, G.; Allgayer, H. The human receptor tyrosine kinase Axl gene--Promoter characterization and regulation of constitutive expression by Sp1, Sp3 and CpG methylation. Biosci. Rep. 2008, 28, 161–176. [Google Scholar] [CrossRef] [Green Version]
- Dhakal, S.; Yu, Z.; Konik, R.; Cui, Y.; Koirala, D.; Mao, H. G-quadruplex and i-motif are mutually exclusive in ILPR double-stranded DNA. Biophys. J. 2012, 102, 2575–2584. [Google Scholar] [CrossRef] [Green Version]
- Farre, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Rosello, L.; Alba, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef] [Green Version]
- Messeguer, X.; Escudero, R.; Farre, D.; Nunez, O.; Martinez, J.; Alba, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef]
- Doi, T.; Vlassara, H.; Kirstein, M.; Yamada, Y.; Striker, G.E.; Striker, L.J. Receptor-specific increase in extracellular matrix production in mouse mesangial cells by advanced glycosylation end products is mediated via platelet-derived growth factor. Proc. Natl. Acad. Sci. USA 1992, 89, 2873–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, W.H.; Zhen, Y.; Rosenbaum, J.; Eisenberg, R.A.; McGaha, T.L.; Birkenbach, M.; Cohen, P.L. A protective role of Mer receptor tyrosine kinase in nephrotoxic serum-induced nephritis. Clin. Immunol. 2010, 136, 236–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crane, M.M.; Sands, B.; Battaglia, C.; Johnson, B.; Yun, S.; Kaeberlein, M.; Brent, R.; Mendenhall, A. In vivo measurements reveal a single 5’-intron is sufficient to increase protein expression level in Caenorhabditis elegans. Sci. Rep. 2019, 9, 9192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiegel, J.; Cuesta, S.M.; Adhikari, S.; Hansel-Hertsch, R.; Tannahill, D.; Balasubramanian, S. G-quadruplexes are transcription factor binding hubs in human chromatin. Genome Biol. 2021, 22, 117. [Google Scholar] [CrossRef]
- Agarwala, P.; Pandey, S.; Ekka, M.K.; Chakraborty, D.; Maiti, S. Combinatorial role of two G-quadruplexes in 5′ UTR of transforming growth factor beta2 (TGFbeta2). Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 129416. [Google Scholar] [CrossRef]
- Morris, M.J.; Negishi, Y.; Pazsint, C.; Schonhoft, J.D.; Basu, S. An RNA G-quadruplex is essential for cap-independent translation initiation in human VEGF IRES. J. Am. Chem. Soc. 2010, 132, 17831–17839. [Google Scholar] [CrossRef]
- Grand, C.L.; Han, H.; Munoz, R.M.; Weitman, S.; Von Hoff, D.D.; Hurley, L.H.; Bearss, D.J. The cationic porphyrin TMPyP4 down-regulates c-MYC and human telomerase reverse transcriptase expression and inhibits tumor growth in vivo. Mol. Cancer Ther. 2002, 1, 565–573. [Google Scholar]
- Mudduluru, G.; Ceppi, P.; Kumarswamy, R.; Scagliotti, G.V.; Papotti, M.; Allgayer, H. Regulation of Axl receptor tyrosine kinase expression by miR-34a and miR-199a/b in solid cancer. Oncogene 2011, 30, 2888–2899. [Google Scholar] [CrossRef]
- Cash, H.; Relle, M.; Menke, J.; Brochhausen, C.; Jones, S.A.; Topley, N.; Galle, P.R.; Schwarting, A. Interleukin 6 (IL-6) deficiency delays lupus nephritis in MRL-Faslpr mice: The IL-6 pathway as a new therapeutic target in treatment of autoimmune kidney disease in systemic lupus erythematosus. J. Rheumatol. 2010, 37, 60–70. [Google Scholar] [CrossRef]
- Rokavec, M.; Oner, M.G.; Li, H.; Jackstadt, R.; Jiang, L.; Lodygin, D.; Kaller, M.; Horst, D.; Ziegler, P.K.; Schwitalla, S.; et al. IL-6R/STAT3/miR-34a feedback loop promotes EMT-mediated colorectal cancer invasion and metastasis. J. Clin. Investig. 2014, 124, 1853–1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.J.; Hilliard, B.; Swami, A.; Madara, J.C.; Rao, S.; Patel, T.; Gaughan, J.P.; Lee, J.; Gadegbeku, C.A.; Choi, E.T.; et al. Growth arrest-specific gene 6 (Gas6) levels are elevated in patients with chronic renal failure. Nephrol. Dial. Transplant. 2012, 27, 4166–4172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, K.; Matsubara, T.; Mima, A.; Sumi, E.; Kanamori, H.; Iehara, N.; Fukatsu, A.; Yanagita, M.; Nakano, T.; Ishimoto, Y.; et al. Gas6 induces Akt/mTOR-mediated mesangial hypertrophy in diabetic nephropathy. Kidney Int. 2005, 68, 552–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagita, M. Gas6, warfarin, and kidney diseases. Clin. Exp. Nephrol. 2004, 8, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Fiebeler, A.; Park, J.K.; Muller, D.N.; Lindschau, C.; Mengel, M.; Merkel, S.; Banas, B.; Luft, F.C.; Haller, H. Growth arrest specific protein 6/Axl signaling in human inflammatory renal diseases. Am. J. Kidney Dis. 2004, 43, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Mudduluru, G.; Leupold, J.H.; Stroebel, P.; Allgayer, H. PMA up-regulates the transcription of Axl by AP-1 transcription factor binding to TRE sequences via the MAPK cascade in leukaemia cells. Biol. Cell 2010, 103, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Axelrod, H.; Pienta, K.J. Axl as a mediator of cellular growth and survival. Oncotarget 2014, 5, 8818–8852. [Google Scholar] [CrossRef] [Green Version]
- Horii, Y.; Muraguchi, A.; Iwano, M.; Matsuda, T.; Hirayama, T.; Yamada, H.; Fujii, Y.; Dohi, K.; Ishikawa, H.; Ohmoto, Y.; et al. Involvement of IL-6 in mesangial proliferative glomerulonephritis. J. Immunol. 1989, 143, 3949–3955. [Google Scholar]
- Zagorska, A.; Traves, P.G.; Lew, E.D.; Dransfield, I.; Lemke, G. Diversification of TAM receptor tyrosine kinase function. Nat. Immunol. 2014, 15, 920–928. [Google Scholar] [CrossRef] [Green Version]
- Lorincz, M.C.; Groudine, M. C(m)C(a/t)GG methylation: A new epigenetic mark in mammalian DNA? Proc. Natl. Acad Sci. USA 2001, 98, 10034–10036. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adams, D.E.; Zhen, Y.; Qi, X.; Shao, W.-H. Axl Expression in Renal Mesangial Cells Is Regulated by Sp1, Ap1, MZF1, and Ep300, and the IL-6/miR-34a Pathway. Cells 2022, 11, 1869. https://doi.org/10.3390/cells11121869
Adams DE, Zhen Y, Qi X, Shao W-H. Axl Expression in Renal Mesangial Cells Is Regulated by Sp1, Ap1, MZF1, and Ep300, and the IL-6/miR-34a Pathway. Cells. 2022; 11(12):1869. https://doi.org/10.3390/cells11121869
Chicago/Turabian StyleAdams, David E., Yuxuan Zhen, Xiaoyang Qi, and Wen-Hai Shao. 2022. "Axl Expression in Renal Mesangial Cells Is Regulated by Sp1, Ap1, MZF1, and Ep300, and the IL-6/miR-34a Pathway" Cells 11, no. 12: 1869. https://doi.org/10.3390/cells11121869
APA StyleAdams, D. E., Zhen, Y., Qi, X., & Shao, W. -H. (2022). Axl Expression in Renal Mesangial Cells Is Regulated by Sp1, Ap1, MZF1, and Ep300, and the IL-6/miR-34a Pathway. Cells, 11(12), 1869. https://doi.org/10.3390/cells11121869