
Circular RNA as a Novel Biomarker and Therapeutic Target for HCC


Abstract
:1. Introduction
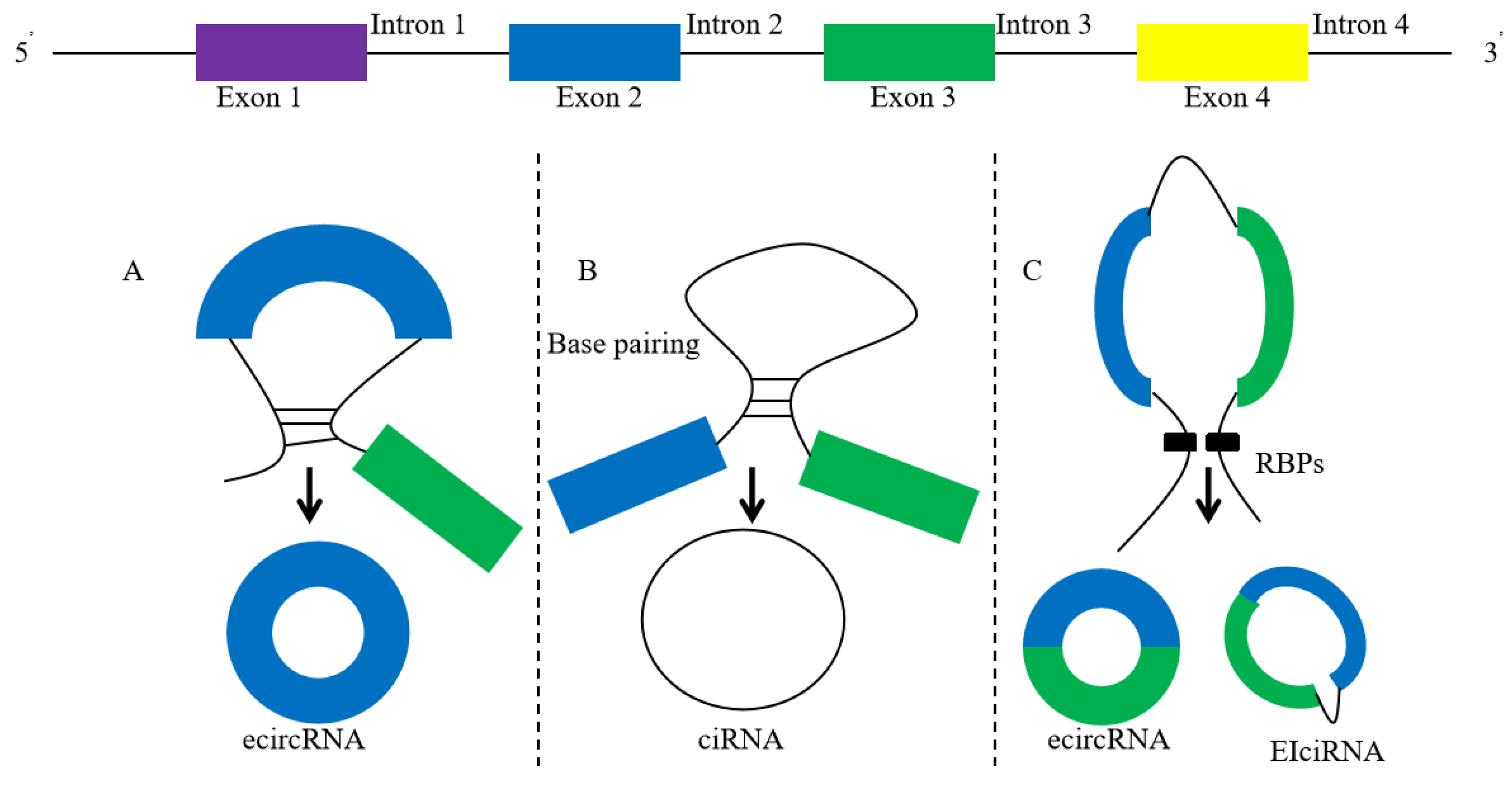
2. Biogenesis of circRNA
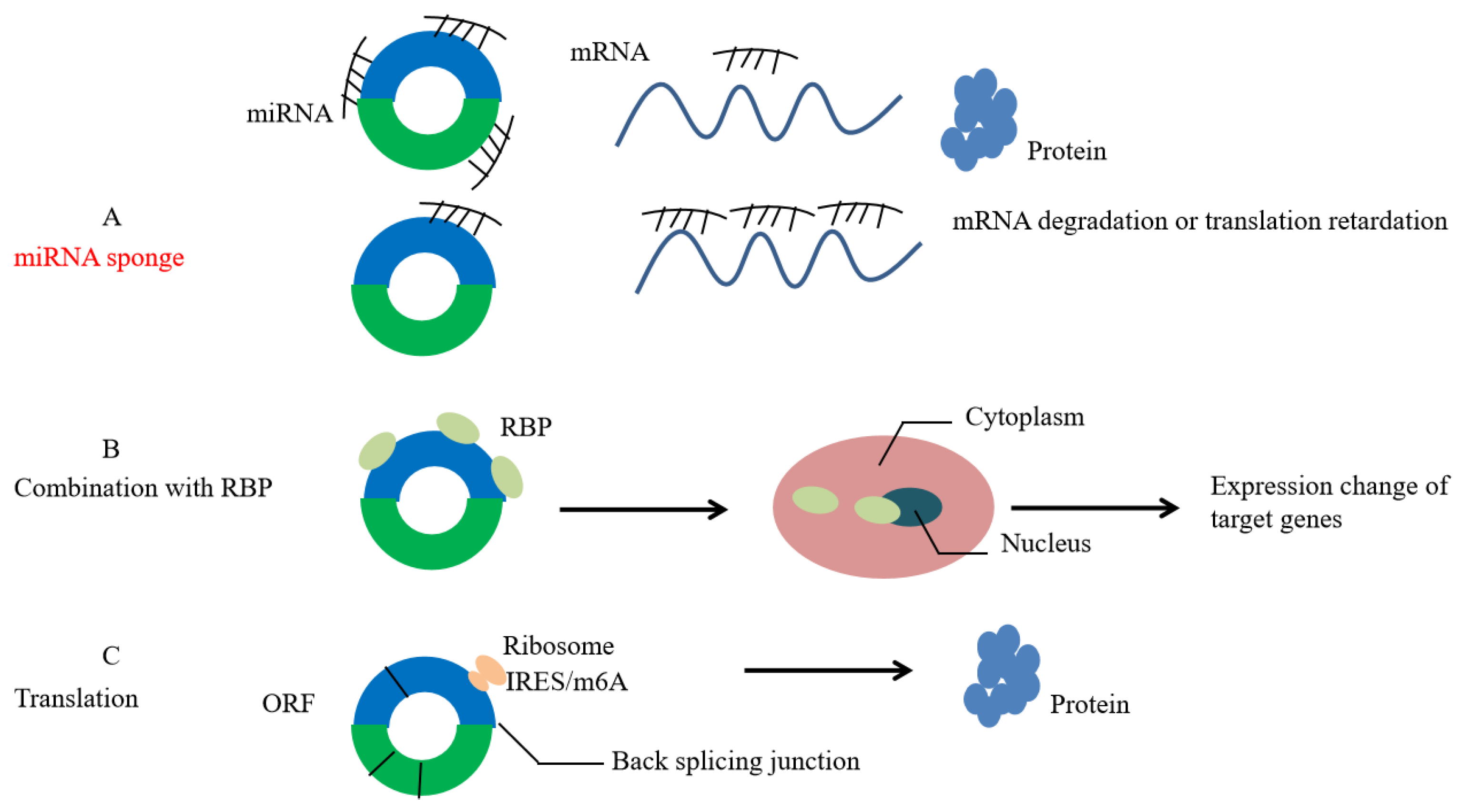
3. Biological Functions of circRNA
4. CircRNA and Liver Diseases
4.1. CircRNA and Alcoholic Liver Injury
4.2. CircRNA and Hepatic Fibrosis
4.3. CircRNA and Hepatocellular Carcinoma
4.3.1. Carcinogenic circRNAs
4.3.2. Tumor Suppressor circRNAs
4.4. Exosome circRNA and Hepatocellular Carcinoma
5. Summary and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, M.T.; Coca-Prados, M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature 1979, 280, 339–340. [Google Scholar] [CrossRef] [PubMed]
- Cocquerelle, C.; Mascrez, B.; Hétuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. FASEB J. 1993, 7, 155–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.G. Epidemiology of alcoholic and nonalcoholic fatty liver disease in China. J. Gastroenterol. Hepatol. 2013, 28, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.S.; Fan, J.G.; Zhang, Z.; Gao, B.; Wang, H.Y. The global burden of liver disease: The major impact of China. Hepatology 2014, 60, 2099–2108. [Google Scholar] [CrossRef]
- Guo, X.Y.; Chen, J.N.; Sun, F.; Wang, Y.Q.; Pan, Q.; Fan, J.G. circRNA_0046367 Prevents Hepatoxicity of Lipid Peroxidation: An Inhibitory Role against Hepatic Steatosis. Oxid. Med. Cell. Longev. 2017, 2017, 3960197. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, K.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Zhang, X.O.; Dong, R.; Zhang, Y.; Zhang, J.L.; Luo, Y.; Zhang, J.; Chen, L.L.; Yang, L. Diverse alternative back-splicing and alternative splicing landscape of circular RNAs. Genome Res. 2016, 26, 1277–1287. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Corrigendum: Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2017, 24, 194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Xu, Q.G.; Wang, Z.G.; Yang, Y.; Zhang, L.; Ma, J.Z.; Sun, S.H.; Yang, F.; Zhou, W.P. Circular RNA cSMARCA5 inhibits growth and metastasis in hepatocellular carcinoma. J. Hepatol. 2018, 68, 1214–1227. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, F.; Li, P. Circular RNAs: Characteristics, Function and Clinical Significance in Hepatocellular Carcinoma. Cancers 2018, 10, 258. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.G.; Chen, R.; Ahmad, S.; Verma, R.; Kasturi, S.P.; Amaya, L.; Broughton, J.P.; Kim, J.; Cadena, C.; Pulendran, B.; et al. Chang, N6-Methyladenosine Modification Controls Circular RNA Immunity. Mol. Cell 2019, 76, 96–109.e9. [Google Scholar] [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA that Can Be Translated and Functions in Myogenesis. Mol. Cell 2017, 66, 22–37.e9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Wang, Z. Efficient backsplicing produces translatable circular mRNAs. RNA 2015, 21, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.L.; Wang, Y.; et al. Extensive translation of circular RNAs driven by N(6)-methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [Green Version]
- Ki, S.H.; Park, O.; Zheng, M.; Morales-Ibanez, O.; Kolls, J.K.; Bataller, R.; Gao, B. Interleukin-22 treatment ameliorates alcoholic liver injury in a murine model of chronic-binge ethanol feeding: Role of signal transducer and activator of transcription 3. Hepatology 2010, 52, 1291–1300. [Google Scholar] [CrossRef] [Green Version]
- Meng, H.; Wang, L.; You, H.; Huang, C.; Li, J. Circular RNA expression profile of liver tissues in an EtOH-induced mouse model of alcoholic hepatitis. Eur. J. Pharmacol. 2019, 862, 172642. [Google Scholar] [CrossRef]
- Dou, X.; Feng, L.; Ying, N.; Ding, Q.; Song, Q.; Jiang, F.; Wang, C.; Li, S. RNA Sequencing Reveals a Comprehensive Circular RNA Expression Profile in a Mouse Model of Alcoholic Liver Disease. Alcohol Clin. Exp. Res. 2020, 44, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.A.; Rossi, S.J.; Paredes, A.H.; Trotter, J.F.; Bashir, M.R.; Guy, C.D.; Banerjee, R.; Jaros, M.J.; Owers, S.; Baxter, B.A.; et al. NGM282 Improves Liver Fibrosis and Histology in 12 Weeks in Patients With Nonalcoholic Steatohepatitis. Hepatology 2020, 71, 1198–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Li, W.X.; Chen, Y.; Li, X.F.; Li, H.D.; Huang, H.M.; Bu, F.T.; Pan, X.Y.; Yang, Y.; Huang, C.; et al. Suppression of SUN2 by DNA methylation is associated with HSCs activation and hepatic fibrosis. Cell Death Dis. 2018, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Feng, R.; Li, X.; Li, D.; Zhai, W. TGF-beta- and lipopolysaccharide-induced upregulation of circular RNA PWWP2A promotes hepatic fibrosis via sponging miR-203 and miR-223. Aging 2019, 11, 9569–9580. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.D.; Bu, F.T.; Li, X.F.; Chen, Y.; Zhu, S.; Wang, J.N.; Chen, S.Y.; Sun, Y.Y.; Pan, X.Y.; et al. Circular RNA circFBXW4 suppresses hepatic fibrosis via targeting the miR-18b-3p/FBXW7 axis. Theranostics 2020, 10, 4851–4870. [Google Scholar] [CrossRef]
- Zhou, Y.; Lv, X.; Qu, H.; Zhao, K.; Fu, L.; Zhu, L.; Ye, G.; Guo, J. Differential expression of circular RNAs in hepatic tissue in a model of liver fibrosis and functional analysis of their target genes. Hepatol. Res. 2019, 49, 324–334. [Google Scholar] [CrossRef]
- Chen, Y.; Yuan, B.; Wu, Z.; Dong, Y.; Zhang, L.; Zeng, Z. Microarray profiling of circular RNAs and the potential regulatory role of hsa_circ_0071410 in the activated human hepatic stellate cell induced by irradiation. Gene 2017, 629, 35–42. [Google Scholar] [CrossRef]
- Jin, H.; Li, C.; Dong, P.; Huang, J.; Yu, J.; Zheng, J. Circular RNA cMTO1 Promotes PTEN Expression Through Sponging miR-181b-5p in Liver Fibrosis. Front. Cell Dev. Biol. 2020, 8, 714. [Google Scholar] [CrossRef]
- Xu, Z.X.; Li, J.Z.; Li, Q.; Xu, M.Y.; Li, H.Y. CircRNA608-microRNA222-PINK1 axis regulates the mitophagy of hepatic stellate cells in NASH related fibrosis. Biochem. Biophys. Res. Commun. 2022, 610, 35–42. [Google Scholar] [CrossRef]
- Ji, D.; Chen, G.F.; Wang, J.C.; Ji, S.H.; Wu, X.W.; Lu, X.J.; Chen, J.L.; Li, J.Y. Hsa_circ_0070963 inhibits liver fibrosis via regulation of miR-223-3p and LEMD3. Aging 2020, 12, 1643–1655. [Google Scholar] [CrossRef]
- Li, S.; Song, F.; Lei, X.; Li, J.; Li, F.; Tan, H. Hsa_circ_0004018 suppresses the progression of liver fibrosis through regulating the hsa-miR-660-3p/TEP1 axis. Aging 2020, 12, 11517–11529. [Google Scholar] [CrossRef] [PubMed]
- Bai, N.; Peng, E.; Qiu, X.; Lyu, N.; Zhang, Z.; Tao, Y.; Li, X.; Wang, Z. circFBLIM1 act as a ceRNA to promote hepatocellular cancer progression by sponging miR-346. J. Exp. Clin. Cancer Res. 2018, 37, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.C.; Wong, C.W.; Liang, P.P.; Shi, M.; Cao, Y.; Rao, S.T.; Tsui, S.K.; Waye, M.M.; Zhang, Q.; Fu, W.M.; et al. Translation of the circular RNA circbeta-catenin promotes liver cancer cell growth through activation of the Wnt pathway. Genome Biol. 2019, 20, 84. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Sun, X.; Huang, Q. Circ_0000105 promotes liver cancer by regulating miR-498/PIK3R1. J. Gene Med. 2020, 22, e3256. [Google Scholar] [CrossRef]
- Wang, M.; Gu, B.; Yao, G.; Li, P.; Wang, K. Circular RNA Expression Profiles and the Pro-tumorigenic Function of CircRNA_10156 in Hepatitis B Virus-Related Liver Cancer. Int. J. Med. Sci. 2020, 17, 1351–1365. [Google Scholar] [CrossRef]
- Niu, W.Y.; Chen, L.; Zhang, P.; Zang, H.; Zhu, B.; Shao, W.B. Circ_0091579 promotes proliferative ability and metastasis of liver cancer cells by regulating microRNA-490-3p. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10264–10273. [Google Scholar]
- Qi, S.X.; Sun, H.; Liu, H.; Yu, J.; Jiang, Z.Y.; Yan, P. Role and mechanism of circ-PRKCI in hepatocellular carcinoma. World J. Gastroenterol. 2019, 25, 1964–1974. [Google Scholar] [CrossRef]
- Fu, Y.; Cai, L.; Lei, X.; Wang, D. Circular RNA ABCB10 promotes hepatocellular carcinoma progression by increasing HMG20A expression by sponging miR-670-3p. Cancer Cell Int. 2019, 19, 338. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Y.; Huang, Z.L.; Xu, Y.H.; Zheng, Q.; Chen, Z.; Song, W.; Zhou, J.; Tang, Z.Y.; Huang, X.Y. Comprehensive circular RNA profiling reveals the regulatory role of the circRNA-100338/miR-141-3p pathway in hepatitis B-related hepatocellular carcinoma. Sci. Rep. 2017, 7, 5428. [Google Scholar] [CrossRef]
- Hu, Z.Q.; Zhou, S.L.; Li, J.; Zhou, Z.J.; Wang, P.C.; Xin, H.Y.; Mao, L.; Luo, C.B.; Yu, S.Y.; Huang, X.W.; et al. Circular RNA Sequencing Identifies CircASAP1 as a Key Regulator in Hepatocellular Carcinoma Metastasis. Hepatology 2020, 72, 906–922. [Google Scholar] [CrossRef]
- Wang, L.; Long, H.; Zheng, Q.; Bo, X.; Xiao, X.; Li, B. Circular RNA circRHOT1 promotes hepatocellular carcinoma progression by initiation of NR2F6 expression. Mol. Cancer 2019, 18, 119. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Pan, X.; Zhu, D.; Deng, Z.; Jiang, R.; Wang, X. Circular RNA MAT2B Promotes Glycolysis and Malignancy of Hepatocellular Carcinoma Through the miR-338-3p/PKM2 Axis Under Hypoxic Stress. Hepatology 2019, 70, 1298–1316. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Huang, G.; Jin, H.; Jian, Z. Circular RNA Gprc5a Promotes HCC Progression by Activating YAP1/TEAD1 Signalling Pathway by Sponging miR-1283. Onco. Targets Ther. 2020, 13, 4509–4521. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Yu, Z.; Zhou, Y.; Tu, J.; Lou, J.; Wang, Y. Circular RNA Circ100084 functions as sponge of miR23a5p to regulate IGF2 expression in hepatocellular carcinoma. Mol. Med. Rep. 2020, 21, 2395–2404. [Google Scholar] [PubMed] [Green Version]
- Zhao, Y.; Song, J.; Tang, B.; Fang, S.; Zhang, D.; Zheng, L.; Wu, F.; Gao, Y.; Chen, C.; Hu, X.; et al. CircSOD2 induced epigenetic alteration drives hepatocellular carcinoma progression through activating JAK2/STAT3 signaling pathway. J. Exp. Clin. Cancer Res. 2020, 39, 259. [Google Scholar] [CrossRef]
- Jiang, Q.L.; Feng, S.J.; Yang, Z.Y.; Xu, Q.; Wang, S.Z. CircHECTD1 up-regulates mucin 1 expression to accelerate hepatocellular carcinoma development by targeting microRNA-485-5p via a competing endogenous RNA mechanism. Chin. Med. J. 2020, 133, 1774–1785. [Google Scholar] [CrossRef]
- Zhou, S.; Wei, J.; Wang, Y.; Liu, X. Cisplatin resistance-associated circRNA_101237 serves as a prognostic biomarker in hepatocellular carcinoma. Exp. Ther. Med. 2020, 19, 2733–2740. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z.; Tan, J.; Gao, W.; Li, X.; Yang, Y.; Li, X.; Li, Y.; Wang, Q. Circular RNA hsa_circ_0016788 regulates hepatocellular carcinoma tumorigenesis through miR-486/CDK4 pathway. J. Cell. Physiol. 2018, 234, 500–508. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Y.; Zhang, P.F.; Wei, C.Y.; Peng, R.; Lu, J.C.; Gao, C.; Cai, J.B.; Yang, X.; Fan, J.; Ke, A.W.; et al. Circular RNA circMET drives immunosuppression and anti-PD1 therapy resistance in hepatocellular carcinoma via the miR-30-5p/snail/DPP4 axis. Mol. Cancer 2020, 19, 92. [Google Scholar] [CrossRef]
- Li, S.; Gu, H.; Huang, Y.; Peng, Q.; Zhou, R.; Yi, P.; Chen, R.; Huang, Z.; Hu, X.; Huang, Y.; et al. Circular RNA 101368/miR-200a axis modulates the migration of hepatocellular carcinoma through HMGB1/RAGE signaling. Cell Cycle 2018, 17, 2349–2359. [Google Scholar] [CrossRef] [Green Version]
- Zhan, W.; Liao, X.; Chen, Z.; Li, L.; Tian, T.; Yu, L.; Wang, W.; Hu, Q. Circular RNA hsa_circRNA_103809 promoted hepatocellular carcinoma development by regulating miR-377-3p/FGFR1/ERK axis. J. Cell. Physiol. 2020, 235, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Shi, Y.; Liu, M.; Sun, J. circHIPK3 regulates cell proliferation and migration by sponging miR-124 and regulating AQP3 expression in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xu, Y.; Qian, Z.; Zheng, W.; Wu, Q.; Chen, Y.; Zhu, G.; Liu, Y.; Bian, Z.; Xu, W.; et al. circRNA_104075 stimulates YAP-dependent tumorigenesis through the regulation of HNF4a and may serve as a diagnostic marker in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Weng, J.; Song, F.; Li, L.; Xiao, C.; Yang, W.; Xu, J. Circular RNA circZNF566 promotes hepatocellular carcinoma progression by sponging miR-4738-3p and regulating TDO2 expression. Cell Death Dis. 2020, 11, 452. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Jing, B.; Bai, Y.; Zhang, Y.; Yu, H. Circular RNA circTMEM45A Acts as the Sponge of MicroRNA-665 to Promote Hepatocellular Carcinoma Progression. Mol. Ther. Nucleic Acids 2020, 22, 285–297. [Google Scholar] [CrossRef]
- Zhang, P.F.; Wei, C.Y.; Huang, X.Y.; Peng, R.; Yang, X.; Lu, J.C.; Zhang, C.; Gao, C.; Cai, J.B.; Gao, P.T.; et al. Circular RNA circTRIM33-12 acts as the sponge of MicroRNA-191 to suppress hepatocellular carcinoma progression. Mol. Cancer 2019, 18, 105. [Google Scholar] [CrossRef]
- Li, X.; Shen, M. Circular RNA hsa_circ_103809 suppresses hepatocellular carcinoma proliferation and invasion by sponging miR-620. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 555–566. [Google Scholar]
- Zhong, L.; Wang, Y.; Cheng, Y.; Wang, W.; Lu, B.; Zhu, L.; Ma, Y. Circular RNA circC3P1 suppresses hepatocellular carcinoma growth and metastasis through miR-4641/PCK1 pathway. Biochem. Biophys. Res. Commun. 2018, 499, 1044–1049. [Google Scholar] [CrossRef]
- Song, C.; Li, D.; Liu, H.; Sun, H.; Liu, Z.; Zhang, L.; Hu, Y. The competing endogenous circular RNA ADAMTS14 suppressed hepatocellular carcinoma progression through regulating microRNA-572/regulator of calcineurin 1. J. Cell. Physiol. 2019, 234, 2460–2470. [Google Scholar] [CrossRef]
- Qiu, L.; Huang, Y.; Li, Z.; Dong, X.; Chen, G.; Xu, H.; Zeng, Y.; Cai, Z.; Liu, X.; Liu, J. Circular RNA profiling identifies circADAMTS13 as a miR-484 sponge which suppresses cell proliferation in hepatocellular carcinoma. Mol. Oncol. 2019, 13, 441–455. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.J.; Zheng, B.; Luo, G.J.; Ma, X.K.; Lu, X.Y.; Lin, X.M.; Yang, S.; Zhao, Q.; Wu, T.; Li, Z.X.; et al. Circular RNAs negatively regulate cancer stem cells by physically binding FMRP against CCAR1 complex in hepatocellular carcinoma. Theranostics 2019, 9, 3526–3540. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xiong, Q.; Wu, Y.; Li, S.; Ge, F. Quantitative Proteomics Reveals the Regulatory Networks of Circular RNA CDR1as in Hepatocellular Carcinoma Cells. J. Proteome Res. 2017, 16, 3891–3902. [Google Scholar] [CrossRef]
- Lin, Q.; Ling, Y.B.; Chen, J.W.; Zhou, C.R.; Chen, J.; Li, X.; Huang, M.S. Circular RNA circCDK13 suppresses cell proliferation, migration and invasion by modulating the JAK/STAT and PI3K/AKT pathways in liver cancer. Int. J. Oncol. 2018, 53, 246–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Chen, T.; Li, C.; Xu, C.; Ding, C.; Chen, J.; Ju, S.; Zhang, Z.; Liang, Z.; Cui, Z.; et al. A circular RNA hsa_circ_0079929 inhibits tumor growth in hepatocellular carcinoma. Cancer Manag. Res. 2019, 11, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Dai, J.; Shu, M. Circular RNA-0072309 has antitumor influences in Hep3B cell line by targeting microRNA-665. Biofactors 2020. [Google Scholar] [CrossRef]
- Han, D.; Li, J.; Wang, H.; Su, X.; Hou, J.; Gu, Y.; Qian, C.; Lin, Y.; Liu, X.; Huang, M.; et al. Circular RNA circMTO1 acts as the sponge of microRNA-9 to suppress hepatocellular carcinoma progression. Hepatology 2017, 66, 1151–1164. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.; Lv, X.; Yin, W.; Zhou, L.; Hu, Y.; Zhou, A.; Qi, F. CircRNA Cdr1as functions as a competitive endogenous RNA to promote hepatocellular carcinoma progression. Aging 2019, 11, 8182–8203. [Google Scholar] [CrossRef]
- Chen, W.; Quan, Y.; Fan, S.; Wang, H.; Liang, J.; Huang, L.; Chen, L.; Liu, Q.; He, P.; Ye, Y. Exosome-transmitted circular RNA hsa_circ_0051443 suppresses hepatocellular carcinoma progression. Cancer Lett. 2020, 475, 119–128. [Google Scholar] [CrossRef]
- Lai, Z.; Wei, T.; Li, Q.; Wang, X.; Zhang, Y.; Zhang, S. Exosomal circFBLIM1 Promotes Hepatocellular Carcinoma Progression and Glycolysis by Regulating the miR-338/LRP6 Axis. Cancer Biother. Radiopharm. 2020. [Google Scholar] [CrossRef]
- Huang, X.Y.; Huang, Z.L.; Huang, J.; Xu, B.; Huang, X.Y.; Xu, Y.H.; Zhou, J.; Tang, Z.Y. Exosomal circRNA-100338 promotes hepatocellular carcinoma metastasis via enhancing invasiveness and angiogenesis. J. Exp. Clin. Cancer Res. 2020, 39, 20. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Deng, T.; Ge, S.; Liu, Y.; Bai, M.; Zhu, K.; Fan, Q.; Li, J.; Ning, T.; Tian, F.H.; et al. Exosome circRNA secreted from adipocytes promotes the growth of hepatocellular carcinoma by targeting deubiquitination-related USP7. Oncogene 2019, 38, 2844–2859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.H.; Wang, Y.T.; Li, G.F.; Zhang, N.; Fan, L. Serum-derived three-circRNA signature as a diagnostic biomarker for hepatocellular carcinoma. Cancer Cell Int. 2020, 20, 226. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| CircRNAs | Expression Change | Function | Possible Mechanism | References |
|---|---|---|---|---|
| hsa_circ_0074837 | Up | HSC activation(+); proliferation(+). | miR-203/miR-223/FSTL1/TLR4 | [24] |
| circFBXW4 | Down | HSC activation(−); proliferation(−); apoptosis(+); Anti-inflammation. | miR-18b-3p/FBXW7 | [25] |
| mmu_circ_34116 | Down | HSC activation(−). | BMP7 | [26] |
| hsa_circ_0071410 | Up | HSC activation(+). | miR-9-5p/MRP1/ABCC1 | [27] |
| hsa_circ_0007874/cMTO1 | Down | HSC activation(−); proliferation(−). | miR-181b-5p/PTEN | [28] |
| circ608 | Down | HSC mitophagy(+). | miR222/PINK1 | [29] |
| hsa_circ_0070963 | Down | HSC activation(−); proliferation(−); cell cycle(−). | miR-223-3p/LEMD3 | [30] |
| hsa_circ_0004018 | Down | HSC activation(−); proliferation(−). | miR-660-3p/TEP1 | [31] |
| CircRNAs | Expression Change | Function | Possible Mechanism | References |
|---|---|---|---|---|
| circβ-catenin | Up | Proliferation(+); migration(+). | β-catenin370aa/Wnt/β-catenin | [33] |
| circ_0000105 | Up | Proliferation(+); apoptosis(−). | miR-498/PIK3R1 | [34] |
| circ_10156 | Up | Proliferation(+) | miR-149-3P/AKT1 pathway | [35] |
| circ_0091579 | Up | Proliferation(+); migration(+). | miRNA-490-3p | [36] |
| circ-PRKCI | Up | Apoptosis(−); invasion(+). | miRNA-545/AKT3 and E2F7 | [37] |
| circABCB10 | Up | Proliferation(+); invasion(+). | miR-670-3p/HMG20A | [38] |
| hsa_circRNA_100338 | Up | Biomarker for clinical diagnosis | miR-141-3p | [39] |
| circASAP1 | Up | Proliferation(+); migration(+); invasion(+). | miR-326/miR-532-5p-MAPK1/CSF-1 signaling | [40] |
| circRHOT1 | Up | Proliferation(+); migration(+); invasion(+); apoptosis(−). | TIP60-dependent NR2F6 | [41] |
| circMAT2B | Up | Tumor-promoting; Glycolysis(+). | miR-338-3p/PKM2 axis | [42] |
| circGprc5a | Up | Proliferation(+); apoptosis(−). | miR-1283/ YAP1/TEAD1 | [43] |
| hsa_circrna_100084 | Up | Proliferation(+); migration(+); invasion(+). | miR-23a-5p/IGF2 | [44] |
| circSOD2 | Up | Cell growth(+); migration(+); cell cycle(+). | miR-502-5p/JAK2/STAT3 | [45] |
| circHECTD1 | Up | Proliferation(+); migration(+); invasion(+); apoptosis(−). | miR-485-5p/MUC1 | [46] |
| circRNA_101237 | Up | Diagnostic biomarker | Not shown | [47] |
| hsa_circ_0016788 | Up | Proliferation(+); invasion(+); apoptosis(−). | miR-486/CDK4 | [48] |
| circMET | Up | Invasion(+); metastasis(+); epithelial to mesenchymal transition (+); Immunosuppression(+). | miR-30-5p/snail/DPP4 | [49] |
| circRNA-101368 | Up | Migration(+). | miR-200a/ HMGB1/RAGE | [50] |
| hsa_circ_103809 | Up | Proliferation(+); cell cycle(+); migration(+). | miR-377-3P/FGFR1 | [51] |
| circHIPK3 | Up | Proliferation(+); migration(+). | miR-124/AQP3 | [52] |
| circ_104075 | Up | Diagnostic biomarker and a therapeutic target. | HNF4a/circ_104075/ miR-582-3p/YAP | [53] |
| circZNF566 | Up | Proliferation(+); migration(+); invasion(+). | miR-4738-3p/ TDO2 | [54] |
| circTMEM45A | Up | tumor growth(+) | miR-665/IGF2 | [55] |
| CircRNAs | Expression Change | Function | Possible Mechanism | References |
|---|---|---|---|---|
| circTRIM33–12 | Down | Proliferation(−); migration(−); invasion(−); immune evasion abilities(−). | miR-191/TET1 | [56] |
| hsa_circ_103809 | Down | Proliferation(−); migration(−); invasion(−). | miR-620 | [57] |
| circC3P1 | Down | Proliferation(−); migration(−); invasion(−). | miR-4641/PCK1 | [58] |
| circADAMTS14 | Down | Proliferation(−); migration(−); invasion(−). | miR-572/RCAN1 | [59] |
| circADAMTS13 | Down | Proliferation(−) | miR-484 | [60] |
| circZKSCAN1 | Down | cancer stem cells(−) | Qki5/circZKSCAN1/FMRP/CCAR1/Wnt signaling axis | [61] |
| circSETD2 | Down | Proliferation(−); invasion(−); migration(−). | E-cadherin, N-cadherin and Vimentin. | [62] |
| circCDK13 | Down | Migration(−); cell cycle(−); invasion(−). | JAK/STAT and PI3K/AKT | [63] |
| hsa_circ_0079299 | Down | Proliferation(−); cell cycle(−). | PI3K/AKT/mTOR | [64] |
| circRNA-0072309 | Down | Tumor suppressor | miR-665/PI3K/AKT and Wnt/β-catenin pathways | [65] |
| hsa_circ_0001445 | Down | Proliferation(−); migration(−). | miR-17-3p and miR-181b-5p/TIMP3 | [13] |
| circMTO1 | Down | Proliferation(−); invasion(−). | miR-9/p21 | [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, H.; Niu, R.; Huang, C.; Li, J. Circular RNA as a Novel Biomarker and Therapeutic Target for HCC. Cells 2022, 11, 1948. https://doi.org/10.3390/cells11121948
Meng H, Niu R, Huang C, Li J. Circular RNA as a Novel Biomarker and Therapeutic Target for HCC. Cells. 2022; 11(12):1948. https://doi.org/10.3390/cells11121948
Chicago/Turabian StyleMeng, Hongwu, Ruowen Niu, Cheng Huang, and Jun Li. 2022. "Circular RNA as a Novel Biomarker and Therapeutic Target for HCC" Cells 11, no. 12: 1948. https://doi.org/10.3390/cells11121948
APA StyleMeng, H., Niu, R., Huang, C., & Li, J. (2022). Circular RNA as a Novel Biomarker and Therapeutic Target for HCC. Cells, 11(12), 1948. https://doi.org/10.3390/cells11121948