
The Influence of MicroRNA-31 on Oxidative Stress and Radiosensitivity in Pancreatic Ductal Adenocarcinoma




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Radiation Treatment
2.3. MiR Transfection
2.4. Reverse Transcription and qPCR
2.5. Silencing GPx8 in PDAC Cells
2.6. Clonogenic Assay
2.7. Cumulative Proliferation Assay
2.8. Measurement of Intracellular ROS
2.9. Measurement of Intracellular GSH/GSSG Levels
2.10. Measurement of Caspase 3/7 Levels
2.11. Protein Extraction and Western Blot Analysis
2.12. Statistical Analysis
3. Results
3.1. Establishing a Stable miR-31 Model in PDAC Cell Lines
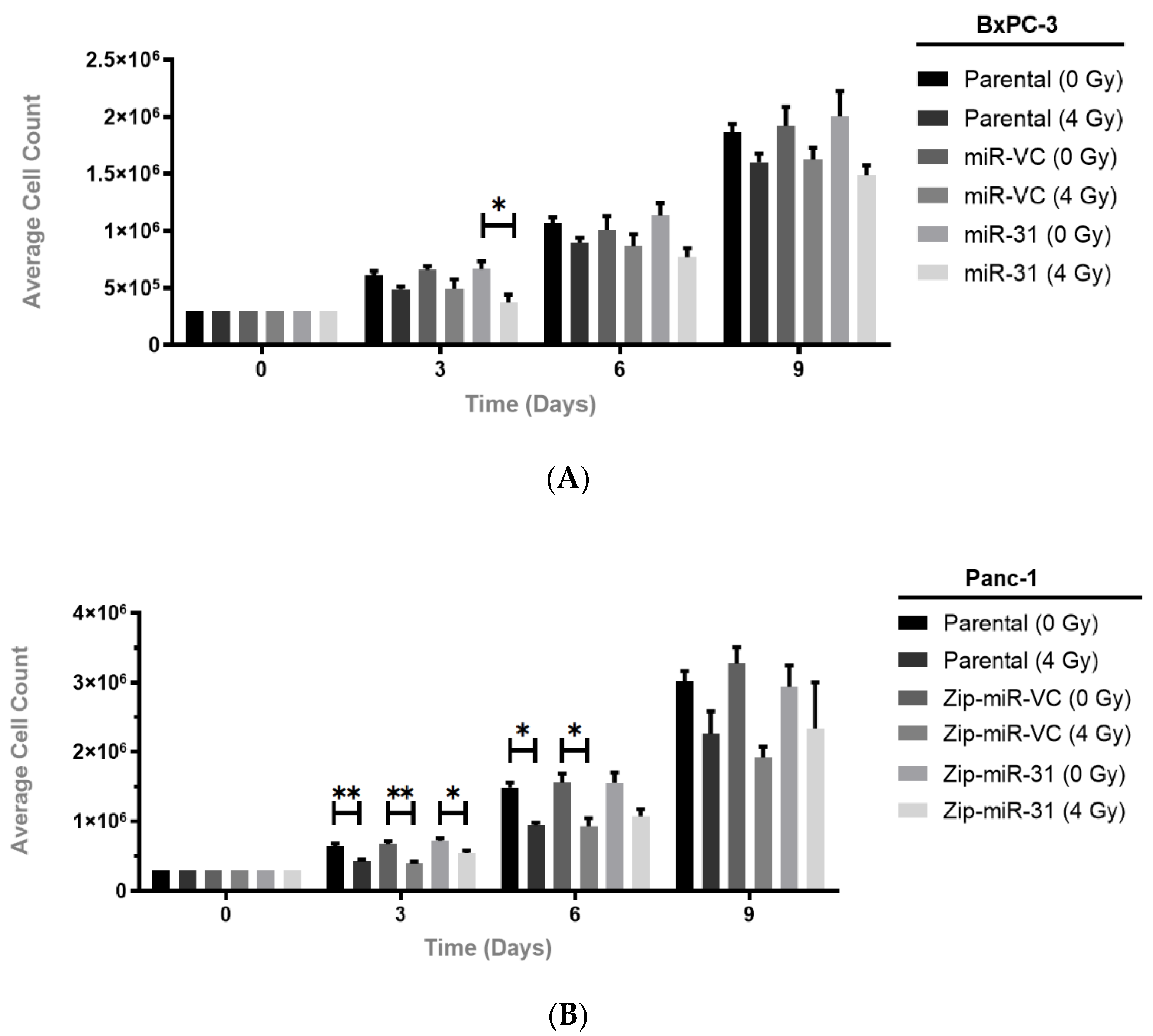
3.2. Manipulating miR-31 Regulates Radiosensitivity in PDAC Cell Lines
3.3. Manipulating miR-31 Alters DNA Damage Induction and Repair in PDAC Cell Lines
3.4. Manipulating miR-31 Alters Radiation-Induced Apoptosis in PDAC Cell Lines
3.5. Manipulating miR-31 Alters Reactive Oxygen Species (ROS) Levels in PDAC Cell Lines
3.6. Manipulating miR-31 Does Not Alter Glutathione (GSH) Levels in PDAC Cell Lines
3.7. Overexpressing miR-31 Alters Glutathione Peroxidase 8 (GPx8) in PDAC Cell Lines
3.8. Silencing GPx8 Enhances Radiosensitivity in BxPC-3 Cells
3.9. Silencing GPx8 Alters Reactive Oxygen Species (ROS) in BxPC-3 Cells
3.10. GPx8 Protects BxPC-3 Cells against DNA-Damage Post-Radiation Treatment
4. Discussion
5. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of Pancreatic Cancer: Global Trends, Etiology and Risk Factors. World J. Oncol. 2019, 10, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Varghese, A.M.; Lowery, M.A.; Yu, K.H.; O’Reilly, E.M. Current management and future directions in metastatic pancreatic adenocarcinoma. Cancer 2016, 122, 3765–3775. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Yang, X.; He, W.; Ye, W. Burden of pancreatic cancer along with attributable risk factors in Europe between 1990 and 2019, and projections until 2039. Int. J. Cancer 2021, 149, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Kikuyama, M.; Kamisawa, T.; Kuruma, S.; Chiba, K.; Kawaguchi, S.; Terada, S.; Satoh, T. Early diagnosis to improve the poor prognosis of pancreatic cancer. Cancers 2018, 10, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, J.F.; Poruk, K.E.; Wolfgang, C.L. Pancreatic cancer surgery: Past, present, and future. Chin. J. Cancer Res. 2015, 27, 332–348. [Google Scholar] [CrossRef]
- Yu, S.; Zhang, C.; Xie, K.P. Therapeutic resistance of pancreatic cancer: Roadmap to its reversal. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188461. [Google Scholar] [CrossRef]
- Quiñonero, F.; Mesas, C.; Doello, K.; Cabeza, L.; Perazzoli, G.; Jimenez-Luna, C.; Rama, A.R.; Melguizo, C.; Prados, J. The challenge of drug resistance in pancreatic ductal adenocarcinoma: A current overview. Cancer Biol. Med. 2019, 16, 688–699. [Google Scholar] [CrossRef]
- Maity, A.; Kao, G.D.; Muschel, R.J.; Gillies Mckenna, W. Potential Molecular Targets for Manipulating The Radiation Response. Int. J. Radiat. Oncol. Biol. Phys 1997, 37, 639–653. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–62. [Google Scholar] [CrossRef] [Green Version]
- Cadet, J.; Richard Wagner, J. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559. [Google Scholar] [CrossRef]
- Turesson, I.; Carlsson, J.; Brahme, A.; Glimelius, B.; Zackrisson, B.; Stenerlöw, B.; Forssell-Aronsson, E.; Hell-Ström, M.; Johansson, K.A.; Kjellén, E.; et al. Biological response to radiation therapy. Acta Oncol. 2003, 42, 1. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Chen, Y.; Li, M.; Ge, Z. Role of antioxidant enzymes on ionizing radiation resistance. Free. Radic. Biol. Med. 1998, 24, 586–593. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Smalheiser, N.R.; Torvik, V.I. Alu elements within human mRNAs are probable microRNA targets. Trends Genet. 2006, 22, 532–536. [Google Scholar] [CrossRef]
- Deer, E.L.; González-Hernández, J.; Coursen, J.D.; Shea, J.E.; Ngatia, J.; Scaife, C.L.; Firpo, M.A.; Mulvihill, S.J. Phenotype and genotype of pancreatic cancer cell lines. Pancreas 2010, 39, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Yan, C.; Gao, Y.; Du, J.; Zhu, X.; Yu, F.; Huang, T.; Dai, J.; Ma, H.; Jiang, Y.; et al. Genetic variants at 9p21.3 are associated with risk of esophageal squamous cell carcinoma in a Chinese population. Cancer Sci. 2017, 108, 250–255. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, S.; Goparaju, C.M.V.; Lopez, P.; Zavadil, J.; Toren-Haritan, G.; Rosenwald, S.; Hoshen, M.; Chajut, A.; Cohen, D.; Pass, H.I. Pro-tumorigenic effects of miR-31 loss in mesothelioma. J. Biol. Chem. 2010, 285, 22809–22817. [Google Scholar] [CrossRef] [Green Version]
- Moody, H.L.; Lind, M.J.; Maher, S.G. MicroRNA-31 Regulates Chemosensitivity in Malignant Pleural Mesothelioma. Mol. Ther. Nucleic Acids 2017, 8, 317–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynam-Lennon, N.; Reynolds, J.V.; Marignol, L.; Sheils, O.M.; Pidgeon, G.P.; Maher, S.G. MicroRNA-31 modulates tumour sensitivity to radiation in oesophageal adenocarcinoma. J. Mol. Med. 2012, 90, 1449–1458. [Google Scholar] [CrossRef] [PubMed]
- Chin, V.; Nagrial, A.; Sjoquist, K.; O’Connor, C.A.; Chantrill, L.; Biankin, A.V.; Scholten, R.J.P.M.; Yip, D. Chemotherapy and radiotherapy for advanced pancreatic cancer. Cochrane Database Syst. Rev. 2018, 3, CD011044. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Fruehauf, J.P.; Meyskens, F.L. Reactive oxygen species: A breath of life or death? Clin. Cancer Res. 2007, 13, 789–794. [Google Scholar] [CrossRef] [Green Version]
- Srinivas, U.S.; Tan BW, Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef]
- Kawamura, K.; Qi, F.; Kobayashi, J. Potential relationship between the biological effects of low-dose irradiation and mitochondrial ROS production. J. Radiat. Res. 2018, 59, ii91–ii97. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin MT, D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Thyagarajan, A.; Sahu, R.P. Potential Contributions of Antioxidants to Cancer Therapy: Immunomodulation and Radiosensitization. Integr. Cancer Ther. 2018, 17, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Ammar, H.O.; Shamma, R.N.; Elbatanony RS, E.; Khater, B. Antioxidants in cancer therapy: Recent trends in application of nanotechnology for enhanced delivery. Sci. Pharm. 2020, 88, 5. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Kim, U.; Kim, C.Y.; Lee, J.M.; Ryu, B.; Kim, J.; Bang, J.; Ahn, N.; Park, J.H. Loss of glutathione peroxidase 3 induces ROS and contributes to prostatic hyperplasia in Nkx3.1 knockout mice. Andrology 2020, 5, 1486–1493. [Google Scholar] [CrossRef]
- Tian, R.; Geng, Y.; Yang, Y.; Seim, I.; Yang, G. Oxidative stress drives divergent evolution of glutathione peroxidase (GPX) gene family in mammals. Integr. Zool. 2021, 5, 696–711. [Google Scholar] [CrossRef]
- Hayes, J.D.; McLellan, L.I. Glutathione and glutathione-dependent enzymes represent a coordinately regulated defence against oxidative stress. Free. Radic. Res. 1999, 31, 273–300. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Guo, S.; Wang, G. Glutathione peroxidases as oncotargets. Oncotarget 2017, 8, 80093–80102. [Google Scholar] [CrossRef] [Green Version]
- Behnisch-Cornwell, S.; Bandaru, S.S.M.; Napierkowski, M.; Wolff, L.; Zubair, M.; Urbainsky, C.; Lillig, C.; Schulzke, C.; Bednarski, P.J. Pentathiepins: A Novel Class of Glutathione Peroxidase 1 Inhibitors that Induce Oxidative Stress, Loss of Mitochondrial Membrane Potential and Apoptosis in Human Cancer Cells. ChemMedChem 2020, 15, 1515–1528. [Google Scholar] [CrossRef]
- Ramming, T.; Hansen, H.G.; Nagata, K.; Ellgaard, L.; Appenzeller-Herzog, C. GPx8 peroxidase prevents leakage of H2O2 from the endoplasmic reticulum. Free. Radic. Biol. Med. 2014, 70, 106–116. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Y.; Guo, Y.; Zhao, Q. GPX8 promotes migration and invasion by regulating epithelial characteristics in non-small cell lung cancer. Thorac. Cancer 2020, 11, 3299–3308. [Google Scholar] [CrossRef]
- Khatib, A.; Solaimuthu, B.; Yosef, M.B.; Rmaileh, A.A.; Tanna, M.; Oren, G.; Frisch, M.S.; Axelrod, J.H.; Lichtenstein, M.; Shaul, Y.D. The glutathione peroxidase 8 (GPX8)/IL-6/STAT3 axis is essential in maintaining an aggressive breast cancer phenotype. Proc. Natl. Acad. Sci. USA 2020, 117, 21420–21431. [Google Scholar] [CrossRef]
- Cellini, F.; Morganti, A.G.; Genovesi, D.; Silvestris, N.; Valentini, V. Role of microRNA in response to ionizing radiations: Evidences and potential impact on clinical practice for radiotherapy. Molecules 2014, 19, 5374–5401. [Google Scholar] [CrossRef]
- Yu, T.; Ma, P.; Wu, D.; Shu, Y.; Gao, W. Functions and mechanisms of microRNA-31 in human cancers. Biomed. Pharmacother. 2018, 108, 1162–1169. [Google Scholar] [CrossRef]
- Papaconstantinou, I.G.; Manta, A.; Gazouli, M.; Lyberopoulou, A.; Lykoudis, P.M.; Polymeneas, G.; Voros, D. Expression of micrornas in patients with pancreatic cancer and its prognostic significance. Pancreas 2013, 42, 9380–9404. [Google Scholar] [CrossRef]
- Peng, H.; Wang, L.; Su, Q.; Yi, K.; Du, J.; Wang, Z. MiR-31-5p promotes the cell growth, migration and invasion of colorectal cancer cells by targeting NUMB. Biomed. Pharmacother. 2019, 109, 208–216. [Google Scholar] [CrossRef]
- Luo, L.; Yang, F.; Ding, J.; Yan, D.; Wang, D.; Yang, S.; Ding, L.; Li, J.; Chen, D.; Ma, R.; et al. MiR-31 inhibits migration and invasion by targeting SATB2 in triple negative breast cancer. Gene 2016, 594, 47–58. [Google Scholar] [CrossRef]
- Estrela, J.M.; Ortega, A.; Obrador, E. Glutathione in cancer biology and therapy. Crit. Rev. Clin. Lab. Sci. 2006, 43, 143–181. [Google Scholar] [CrossRef]
- Pajic, M.; Froio, D.; Daly, S.; Doculara, L.; Millar, E.; Graham, P.H.; Drury, A.; Steinmann, A.; De Bock, C.E.; Boulghourjian, A.; et al. miR-139-5p modulates radiotherapy resistance in breast cancer by repressing multiple gene networks of DNA repair and ROS defense. Cancer Res. 2018, 78, 501–515. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhang, Y.; Ding, J.; Hu, W.; Tan, C.; Wang, M.; Tang, J.; Xu, Y. miR-17-3p Downregulates Mitochondrial Antioxidant Enzymes and Enhances the Radiosensitivity of Prostate Cancer Cells. Mol. Ther. Nucleic Acids 2018, 13, 64–77. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Shen, Y.; Wei, J.; Liu, F. MicroRNA-153/Nrf-2/GPx1 pathway regulates radiosensitivity and stemness of glioma stem cells via reactive oxygen species. Oncotarget 2015, 6, 22006–22027. [Google Scholar] [CrossRef] [Green Version]



















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGrath, J.; Kane, L.E.; Maher, S.G. The Influence of MicroRNA-31 on Oxidative Stress and Radiosensitivity in Pancreatic Ductal Adenocarcinoma. Cells 2022, 11, 2294. https://doi.org/10.3390/cells11152294
McGrath J, Kane LE, Maher SG. The Influence of MicroRNA-31 on Oxidative Stress and Radiosensitivity in Pancreatic Ductal Adenocarcinoma. Cells. 2022; 11(15):2294. https://doi.org/10.3390/cells11152294
Chicago/Turabian StyleMcGrath, Jason, Laura E. Kane, and Stephen G. Maher. 2022. "The Influence of MicroRNA-31 on Oxidative Stress and Radiosensitivity in Pancreatic Ductal Adenocarcinoma" Cells 11, no. 15: 2294. https://doi.org/10.3390/cells11152294
APA StyleMcGrath, J., Kane, L. E., & Maher, S. G. (2022). The Influence of MicroRNA-31 on Oxidative Stress and Radiosensitivity in Pancreatic Ductal Adenocarcinoma. Cells, 11(15), 2294. https://doi.org/10.3390/cells11152294