
Transcriptomes of Injured Lamprey Axon Tips: Single-Cell RNA-Seq Suggests Differential Involvement of MAPK Signaling Pathways in Axon Retraction and Regeneration after Spinal Cord Injury

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Spinal Cord Transection and Retrograde Labeling of Axons
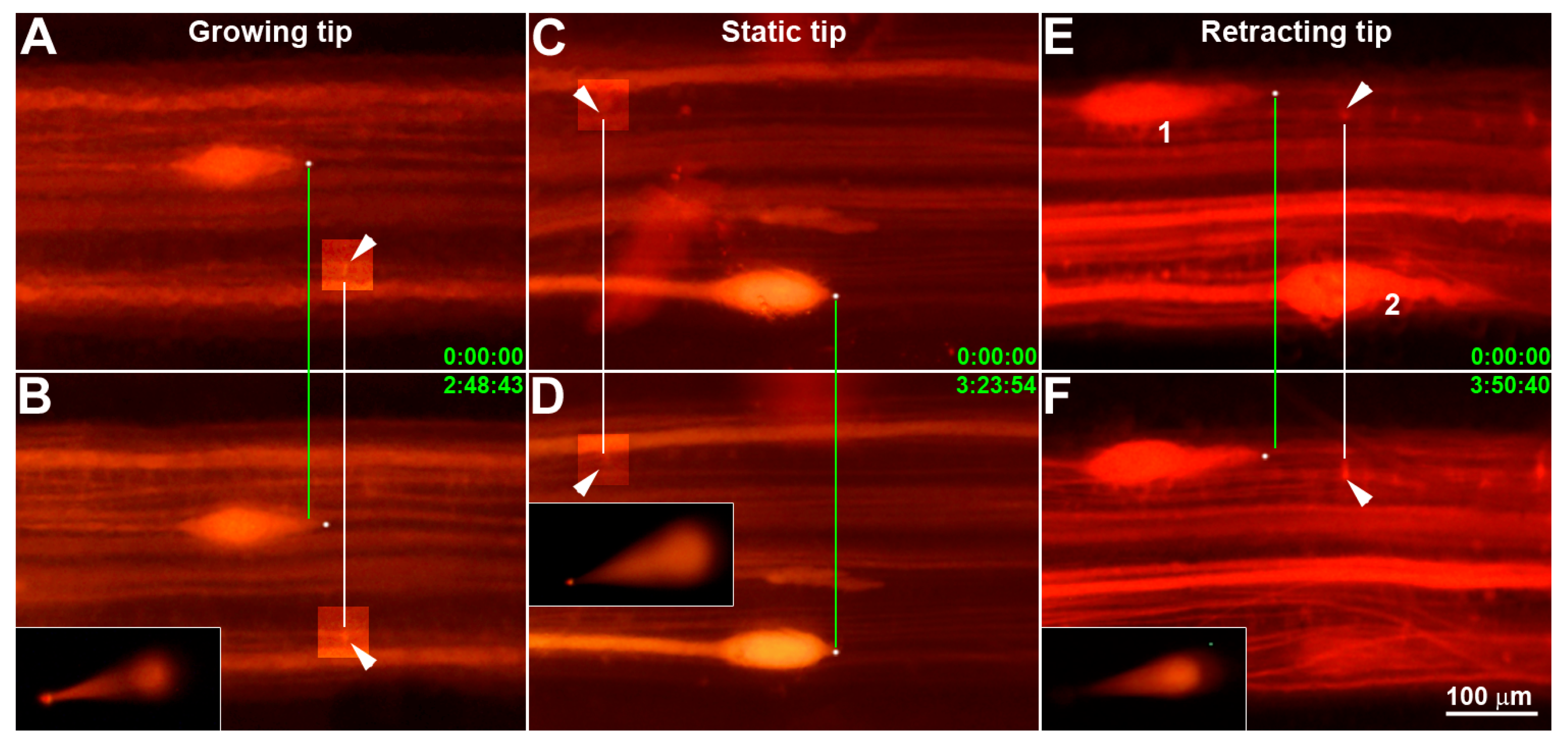
2.2. Determination of Axon Tip Growth Status
2.3. Isolation of the Axoplasmic Contents from Individual Axon Tips
2.4. Construction of Libraries
2.5. scRNA-seq and Data Analysis
2.6. Cryostat Sectioning
2.7. Immunohistochemistry (IHC)
2.8. Real-Time Quantitative Polymerase Chain Reaction (q-PCR)
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. Determination of Growth Status and the Micro-Aspiration of Axoplasms
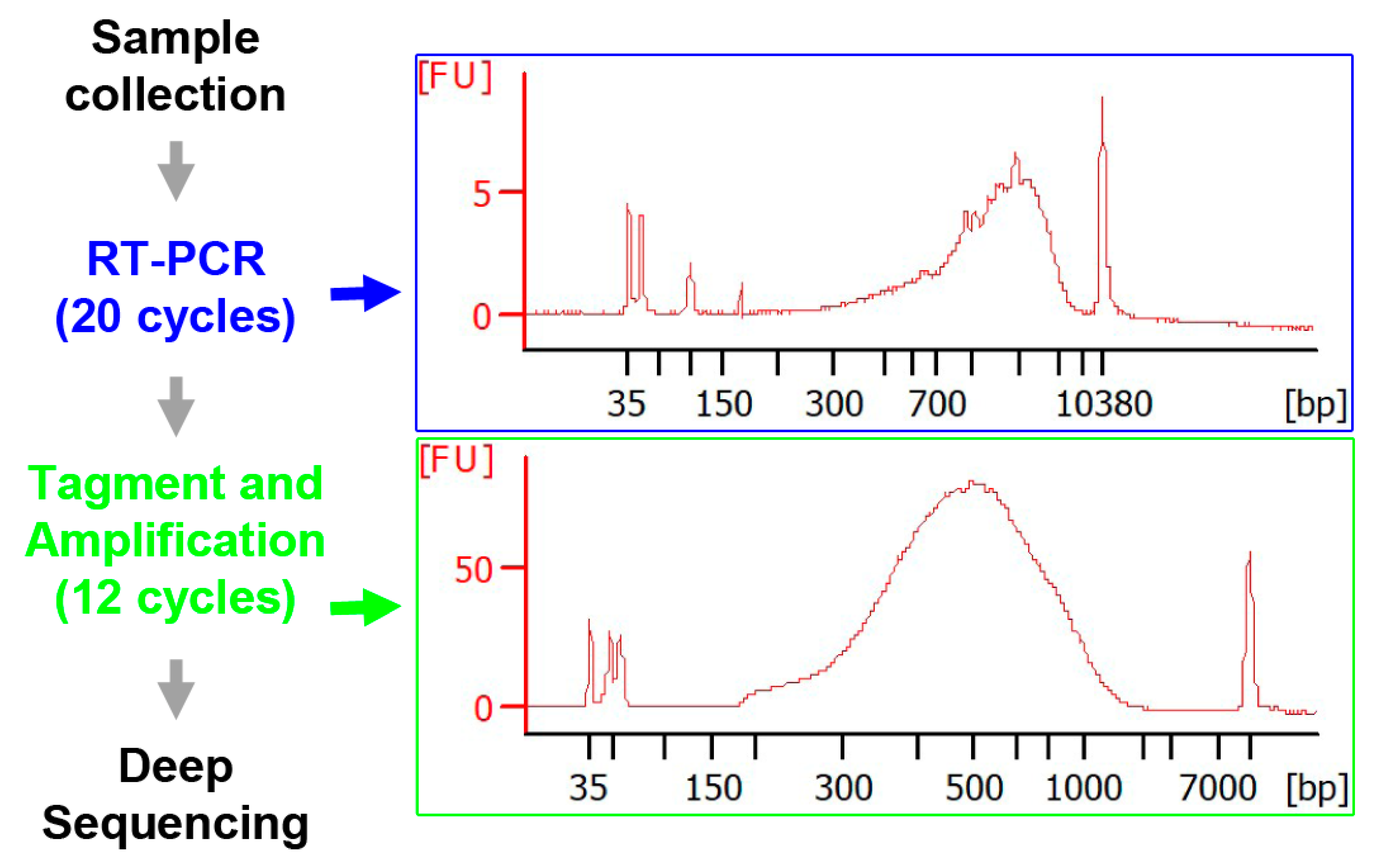
3.2. Construction of scRNA-seq Libraries
3.3. RNA-Seq, Alignments and Assemblies
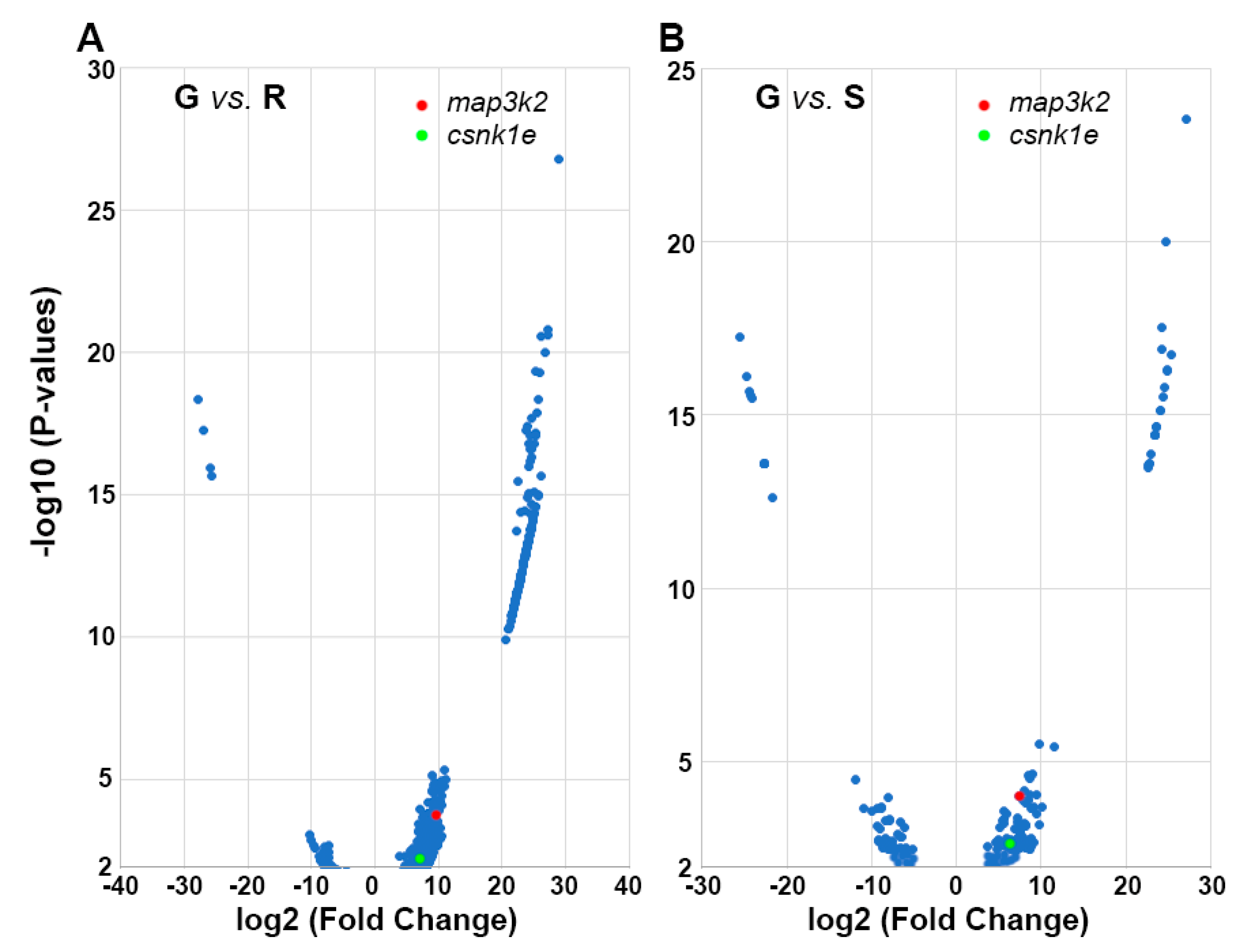
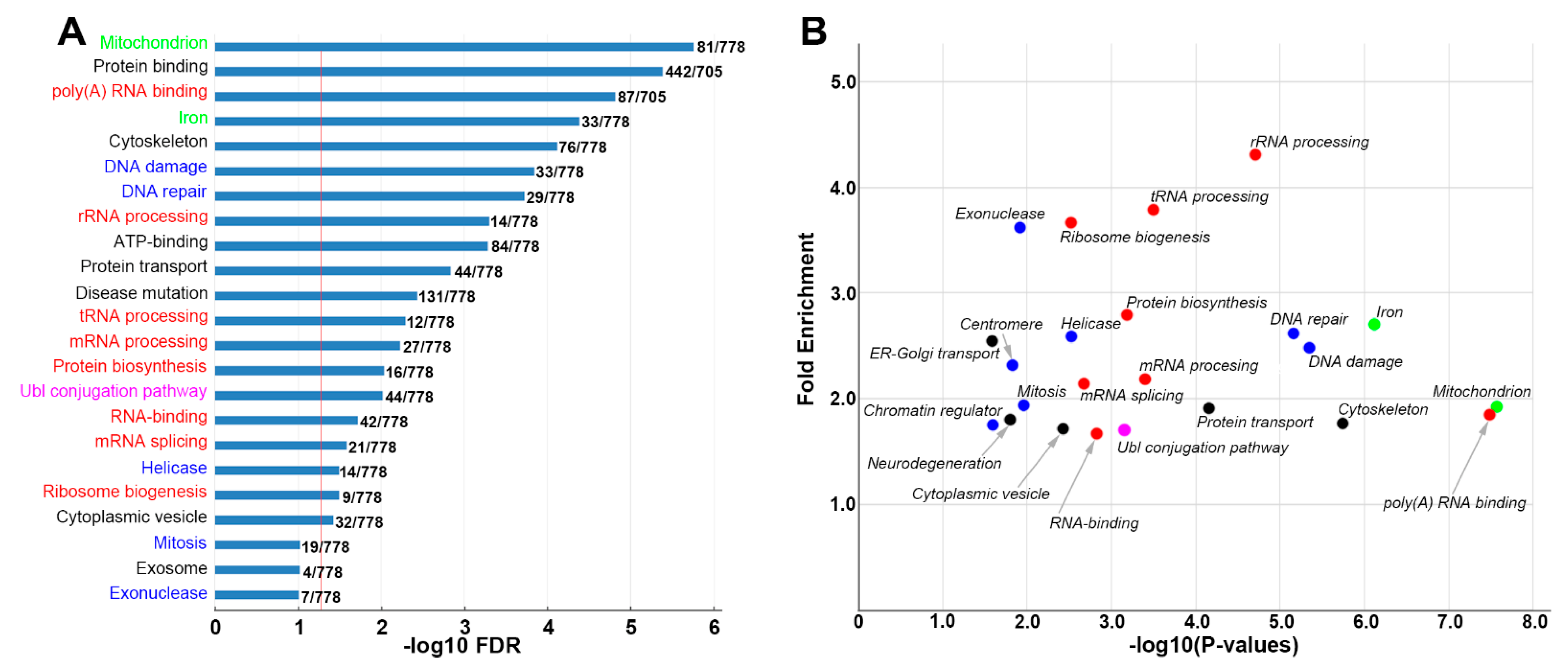
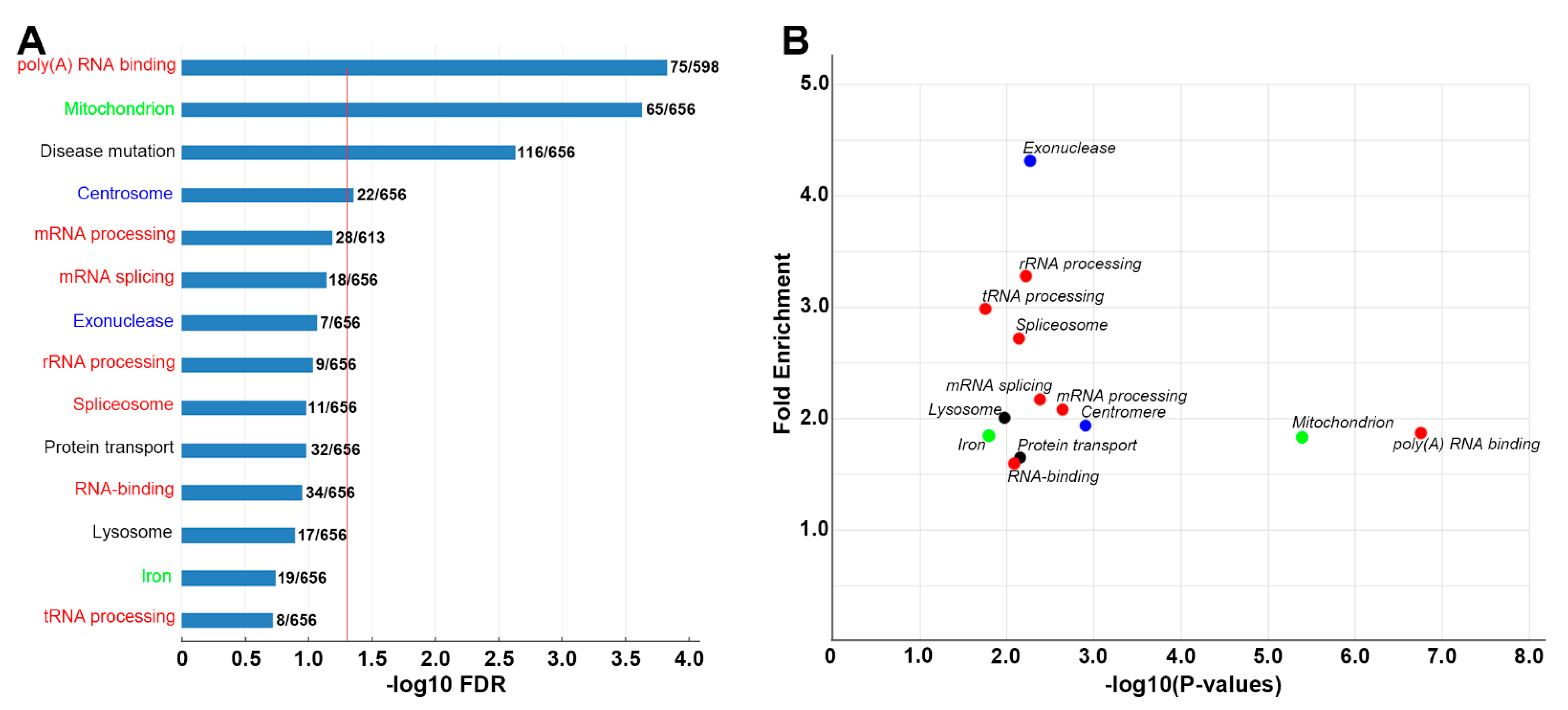
3.4. DEG Identification and DAVID Enrichment Analysis
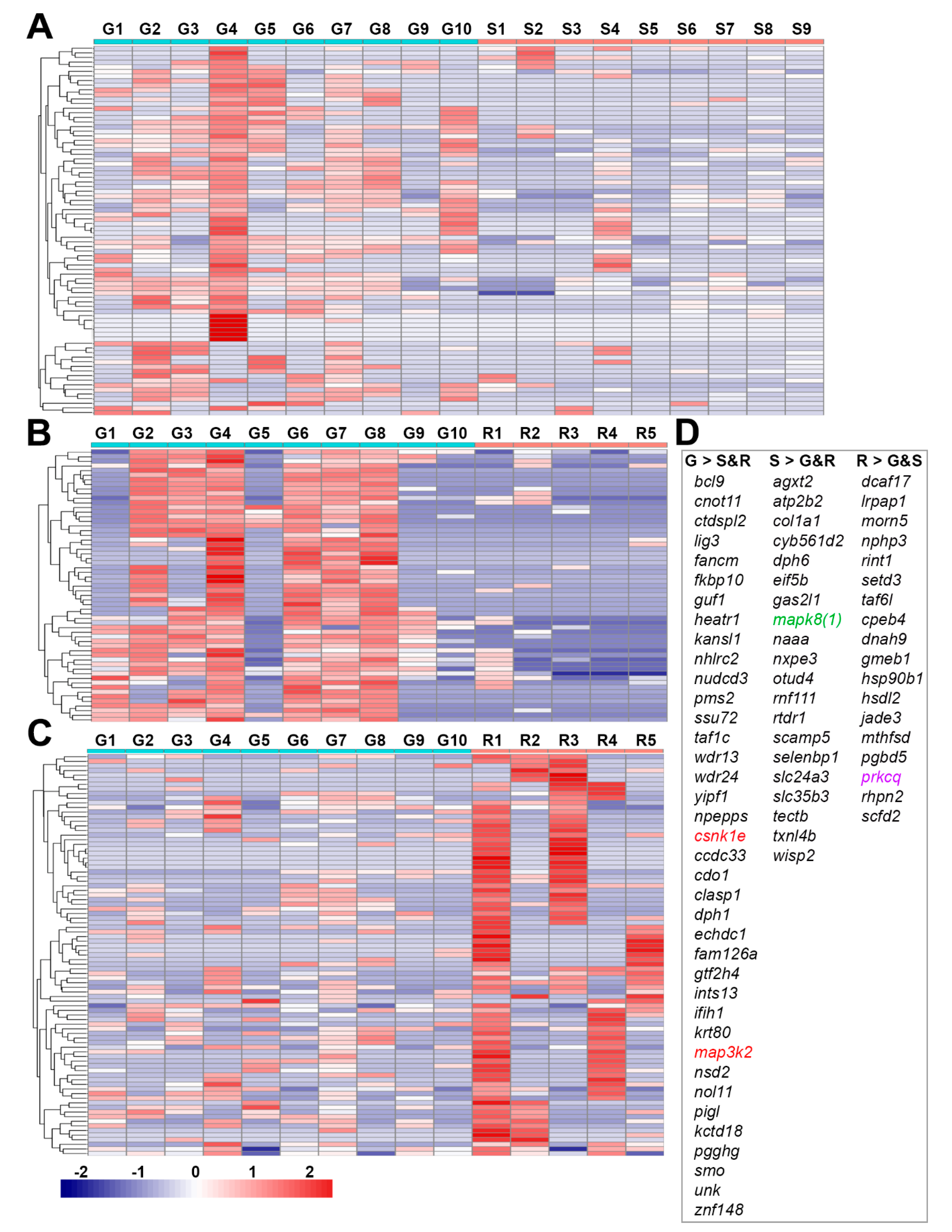
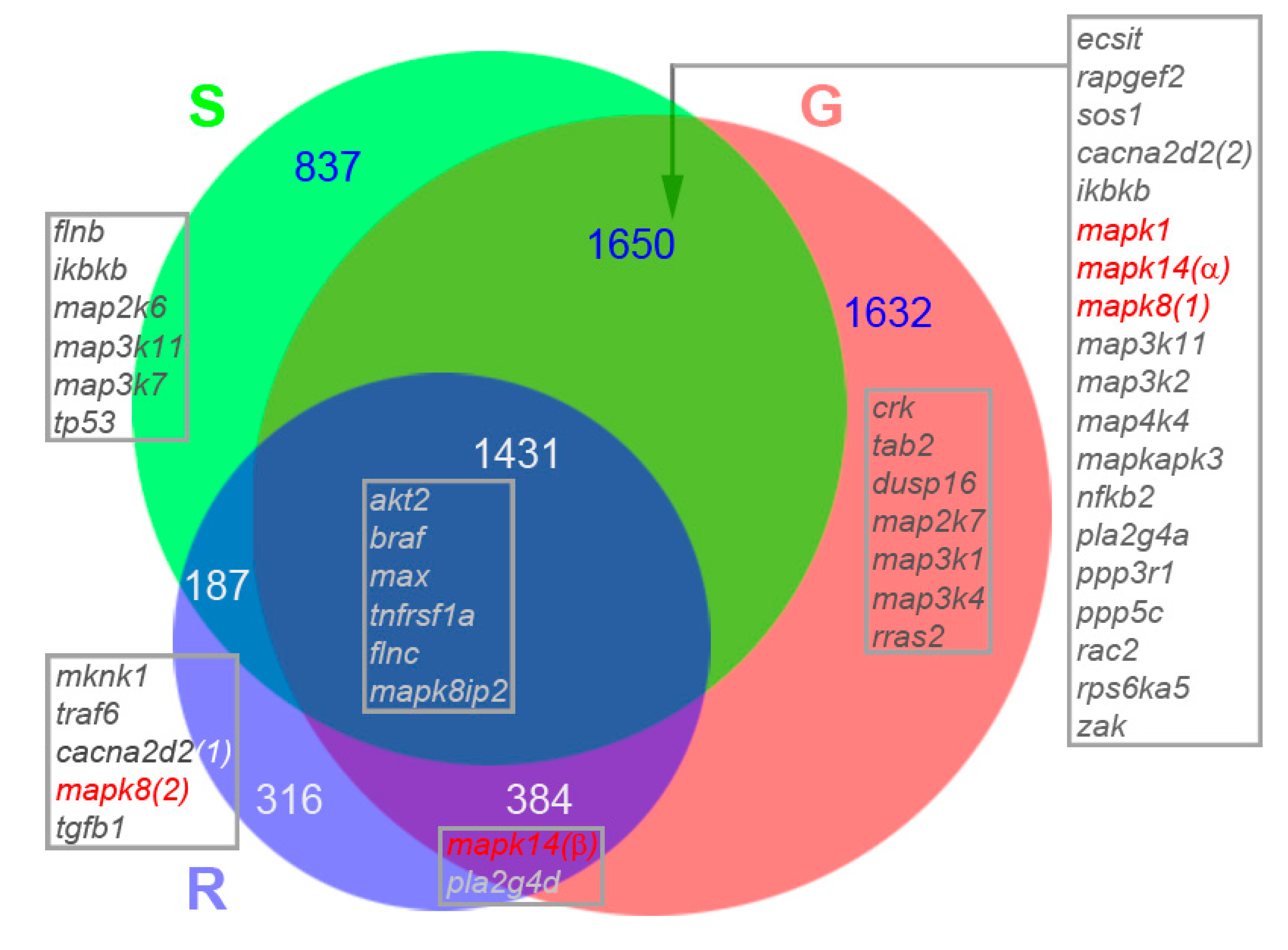
3.5. Venn Diagram Analysis of Genes Participating in MAPK Pathways and Regulating Histone Functions
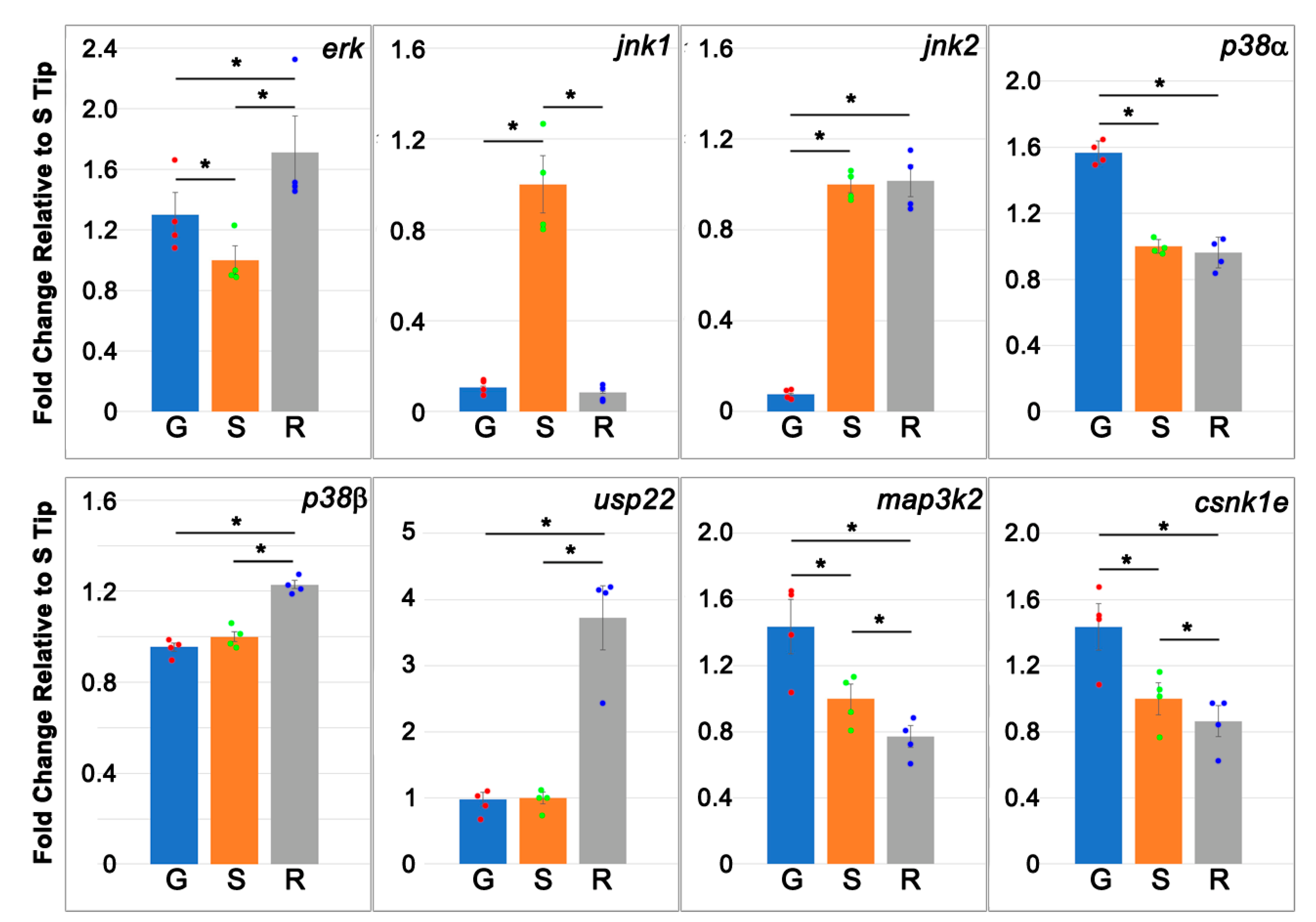
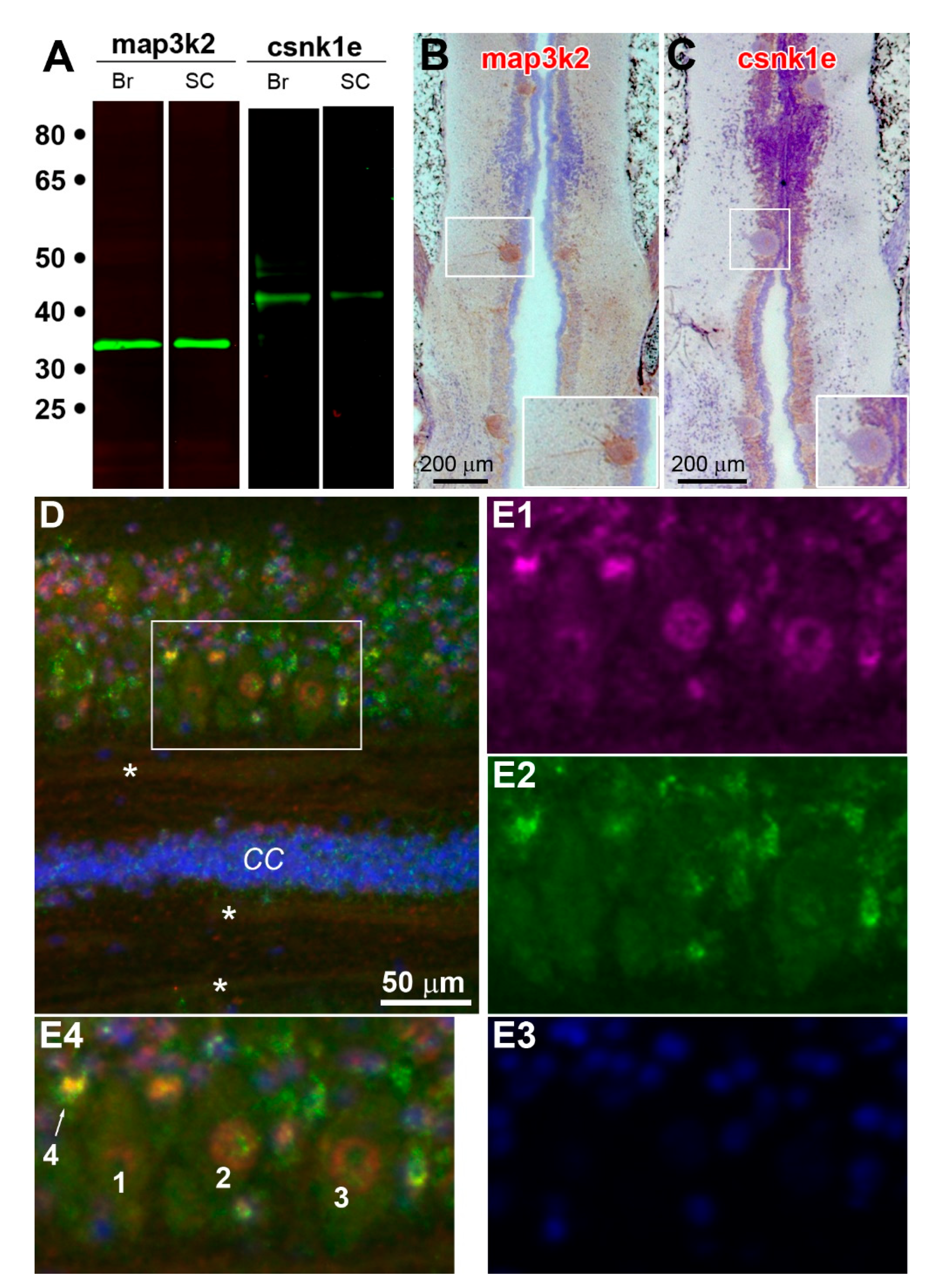
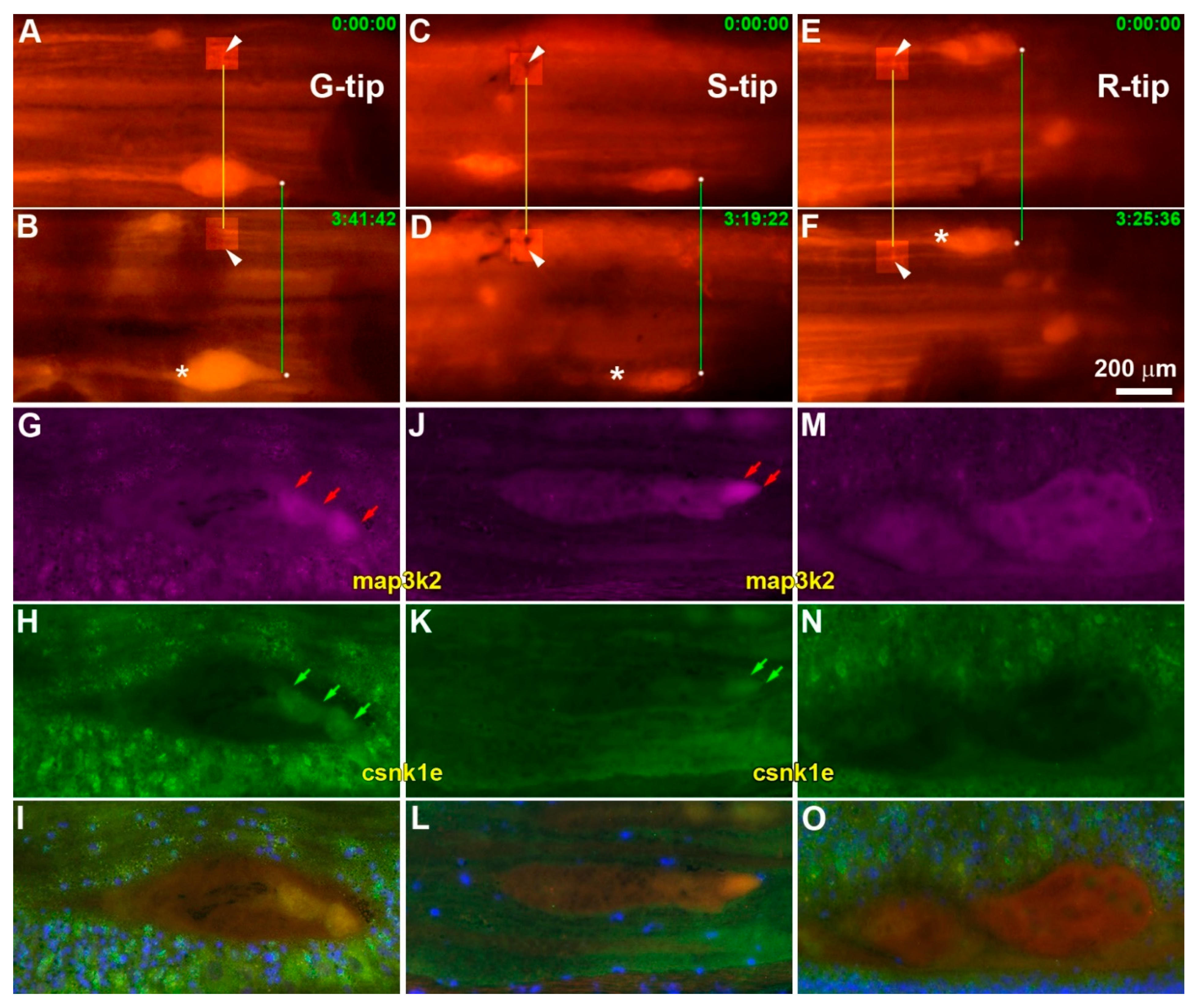
3.6. Validation of Genes by Real-Time q-PCR, IHC and Fluorescence Labeling
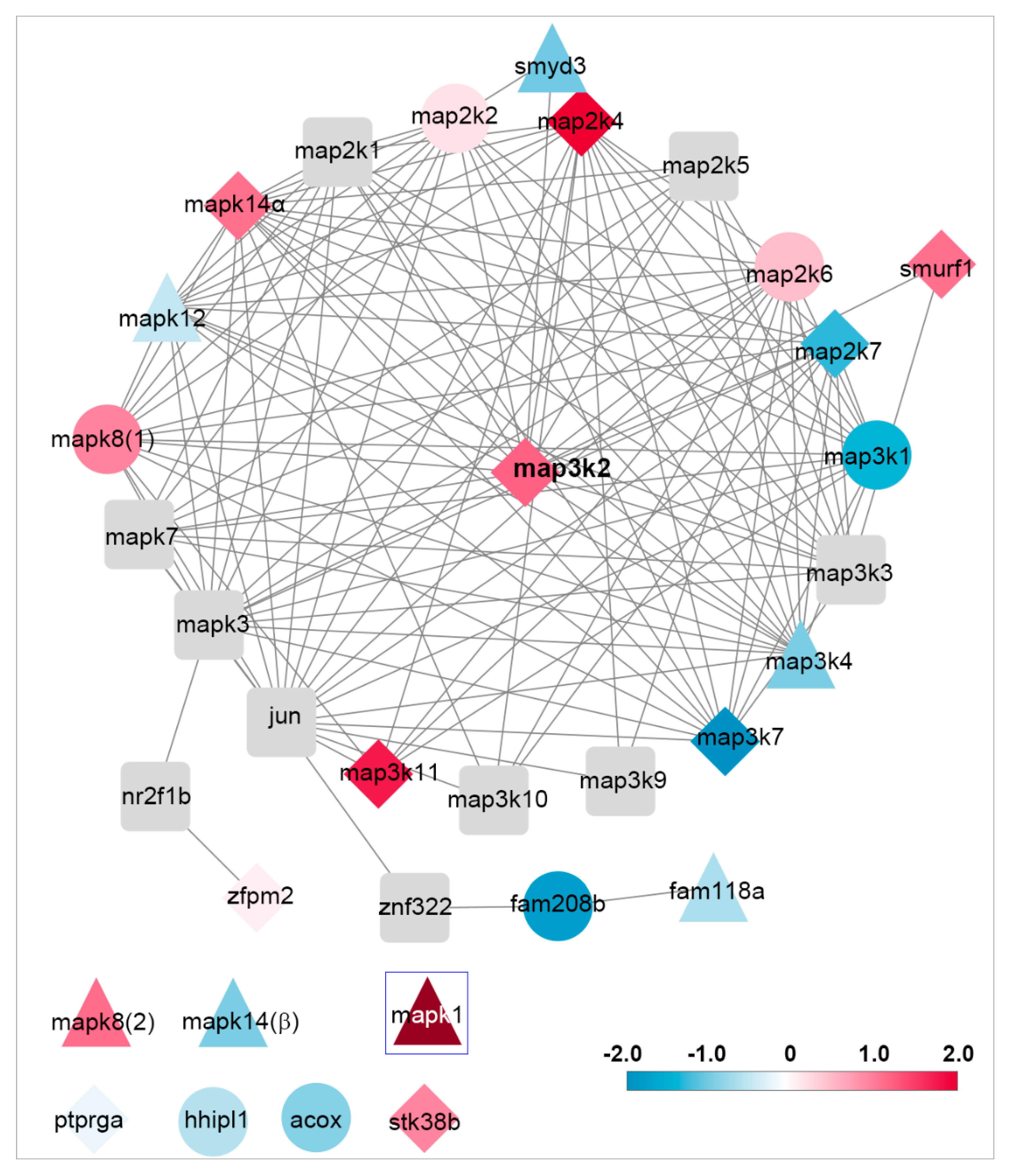
3.7. Network Analysis of Genes Related to map3k2 and Csnk1e
4. Discussion
4.1. Transcriptomes Prepared from Individual Axon Tips in Three Stages of Axon Growth
4.2. Genes Directly Related to Axon Regeneration
- (1)
- map3k2 and csnk1e: From the G > S and R list in hierarchical cluster analysis (Figure 3D), the protein encoded by map3k2 is involved in the MAPK pathway, and the protein encoded by csnk1e, casein kinase 1 epsilon, is involved in circadian rhythms. Results from qPCR (Figure 8) and IHC studies (Figure 10) confirmed their presence in axon tips. The colocalization of map3k2 and csnk1e in growing tips, and their parallel expression levels in three different tips, suggests cross-talk between the MAPK pathway and circadian rhythm proteins in the regulation of axon elongation. Indeed, such cross-talk has been described previously for MAPK in the regulation of circadian rhythms [50]. Their roles in axon regeneration are discussed below.
- (2)
- Genes involved in ribosomal function: Approximately 300 of the genes found in axon tips are believed to be directly involved in RNA processing, ribosome biogenesis and protein synthesis (Figure 5).
- (3)
- G > R genes: Among the top 10 genes upregulated more in growing tips than in retracting tips (Table 2), eif2s1a and eif4b are translation initiation factors. The protein encoded by alas2 catalyzes the first step in the heme biosynthetic pathway.
- (4)
- Genes related to histone modification: Genes, including hadac1, hadac3 and hdac8, encode histone deacetylase isoforms 1, 3 and 8, respectively, found in all three types of tips (G, S and R) are thought to act in maintaining the stability of chromatin structures (Figure S5). The gene hat1 encodes histone acetyltransferase-1, which modifies DNA transcription, and is found in G and S tips.
- (5)
- Genes related to RNA binding.
4.3. Genes Related to Axon Retraction
- (1)
- Genes found in the R > S and G list (Figure 3D): Among the genes expressed selectively in retracting axon tips, dcaf17 encodes a protein associated with cullin 4A/damaged DNA binding protein 1 (DDB1), which is involved in protein degradation. The gene prkcq encodes protein kinase C theta, which is involved in T cell activation and other functions.
- (2)
- Genes found in the G < R or R > S lists: Among the top 10 genes expressed at higher levels in retracting than in growing tips (G < R, Table 2) or in retracting than in static tips (R > S, Table 4) are cdk7, rxfp3, oprm1, grm7,and sdc2. Proteins encoded belong to the “Gα(i) signaling events” family (STRING, FDR = 0.035). They inhibit the cAMP-dependent pathway through the inhibition of adenylate cyclase [51]. Genes prkcq and sdc2 encode protein kinase C theta (above) and syndecan 2, and both participate in apoptosis of osteoblasts [52].
- (3)
4.4. Genes Involved in Local Protein Synthesis and Mitochondrial Function
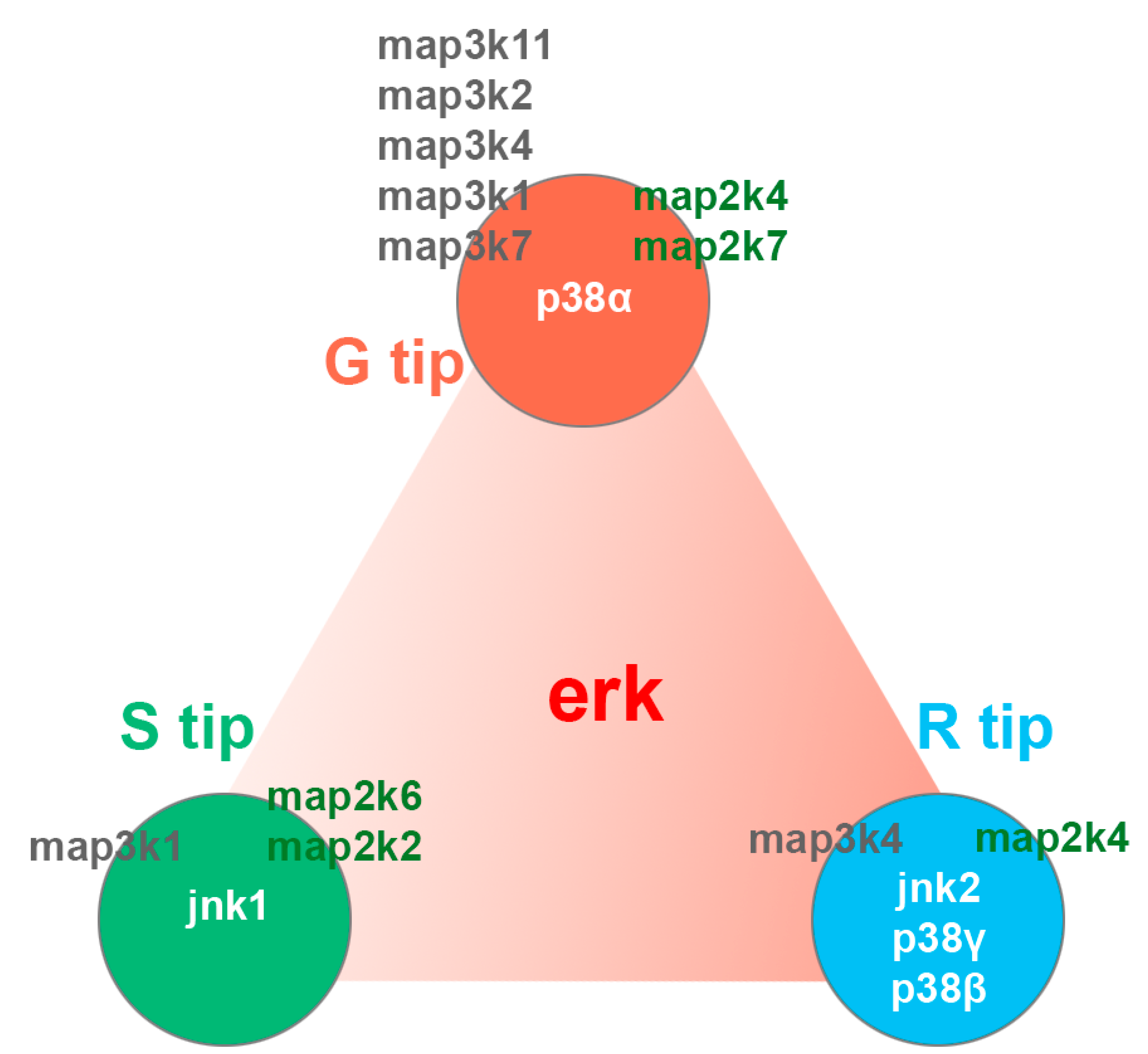
4.5. MAPK Pathway in Axon Regeneration
4.6. Csnk1e/Circadian Rhythm Network in Axon Regeneration
4.7. Genes Smo and Prkcq in Axon Regeneration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Perlson, E.; Hanz, S.; Ben-Yaakov, K.; Segal-Ruder, Y.; Seger, R.; Fainzilber, M. Vimentin-Dependent Spatial Translocation of an Activated MAP Kinase in Injured Nerve. Neuron 2005, 45, 715–726. [Google Scholar] [CrossRef] [Green Version]
- Lindwall, C.; Kanje, M. Retrograde axonal transport of JNK signaling molecules influence injury induced nuclear changes in p-c-Jun and ATF3 in adult rat sensory neurons. Mol. Cell Neurosci. 2005, 29, 269–282. [Google Scholar] [CrossRef]
- Jin, L.-Q.; John, B.H.; Hu, J.; Selzer, M.E. Activated Erk Is an Early Retrograde Signal After Spinal Cord Injury in the Lamprey. Front. Neurosci. 2020, 14, 580692. [Google Scholar] [CrossRef]
- Dalla Costa, I.; Buchanan, C.N.; Zdradzinski, M.D.; Sahoo, P.K.; Smith, T.P.; Thames, E.; Kar, A.N.; Twiss, J.L. The Functional Organization of Axonal Mrna Transport and Translation. Nat. Rev. Neurosci. 2021, 22, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Twiss, J.L.; Kalinski, A.L.; Sachdeva, R.; Houle, J.D. Intra-Axonal Protein Synthesis—A New Target for Neural Repair? Neural Regen. Res. 2016, 11, 1365–1367. [Google Scholar] [CrossRef]
- Twiss, J.L.; van Minnen, J. New Insights into Neuronal Regeneration: The Role of Axonal Protein Synthesis in Pathfinding and Axonal Extension. J. Neurotrauma 2006, 23, 295–308. [Google Scholar] [CrossRef]
- Gumy, L.F.; Tan, C.L.; Fawcett, J.W. The Role of Local Protein Synthesis and Degradation in Axon Regeneration. Exp. Neurol. 2010, 223, 28–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tobias, G.S.; Koenig, E. Axonal Protein Synthesizing Activity During the Early Outgrowth Period Following Neurotomy. Exp. Neurol. 1975, 49, 221–234. [Google Scholar] [CrossRef]
- Zheng, J.Q.; Kelly, T.K.; Chang, B.; Ryazantsev, S.; Rajasekaran, A.K.; Martin, K.C.; Twiss, J.L. A Functional Role for Intra-Axonal Protein Synthesis During Axonal Regeneration from Adult Sensory Neurons. J. Neurosci. 2001, 21, 9291–9303. [Google Scholar] [CrossRef]
- Gumy, L.F.; Yeo, G.S.; Tung, Y.C.; Zivraj, K.H.; Willis, D.; Coppola, G.; Lam, B.Y.; Twiss, J.L.; Holt, C.E.; Fawcett, J.W. Transcriptome Analysis of Embryonic and Adult Sensory Axons Reveals Changes in Mrna Repertoire Localization. RNA 2011, 17, 85–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zivraj, K.H.; Tung, Y.C.; Piper, M.; Gumy, L.; Fawcett, J.W.; Yeo, G.S.; Holt, C.E. Subcellular Profiling Reveals Distinct and Developmentally Regulated Repertoire of Growth Cone Mrnas. J. Neurosci. 2010, 30, 15464–15478. [Google Scholar] [CrossRef] [Green Version]
- Piper, M.; Holt, C. Rna Translation in Axons. Annu. Rev. Cell Dev. Biol. 2004, 20, 505–523. [Google Scholar] [CrossRef] [Green Version]
- Farias, J.; Sotelo, J.R.; Sotelo-Silveira, J. Toward Axonal System Biology: Genome Wide Views of Local Mrna Translation. Proteomics 2019, 19, e1900054. [Google Scholar] [CrossRef]
- Minis, A.; Dahary, D.; Manor, O.; Leshkowitz, D.; Pilpel, Y.; Yaron, A. Subcellular Transcriptomics-Dissection of the Mrna Composition in the Axonal Compartment of Sensory Neurons. Dev. Neurobiol. 2014, 74, 365–381. [Google Scholar] [CrossRef]
- Selzer, M.E. Mechanisms of Functional Recovery and Regeneration after Spinal Cord Transection in Larval Sea Lamprey. J. Physiol. 1978, 277, 395–408. [Google Scholar] [CrossRef] [Green Version]
- Cohen, A.H.; Mackler, S.A.; Selzer, M.E. Functional Regeneration Following Spinal Transection Demonstrated in the Isolated Spinal Cord of the Larval Sea Lamprey. Proc. Natl. Acad. Sci. USA 1986, 83, 2763–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovainen, C.M. Regeneration of Müller and Mauthner Axons after Spinal Transection in Larval Lampreys. J. Comp. Neurol. 1976, 168, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.H.; Mackler, S.A.; Selzer, M.E. Behavioral Recovery Following Spinal Transection: Functional Regeneration in the Lamprey Cns. Trends Neurosci. 1988, 11, 227–231. [Google Scholar] [CrossRef]
- Davis, G.R., Jr.; McClellan, A.D. Time Course of Anatomical Regeneration of Descending Brainstem Neurons and Behavioral Recovery in Spinal-Transected Lamprey. Brain Res. 1993, 602, 131–137. [Google Scholar] [CrossRef]
- Yin, H.S.; Selzer, M.E. Axonal Regeneration in Lamprey Spinal Cord. J. Neurosci. 1983, 3, 1135–1144. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, A.J.; Swain, G.P.; Snedeker, J.A.; Pijak, D.S.; Gladstone, L.J.; Selzer, M.E. Recovery of Neurofilament Expression Selectively in Regenerating Reticulospinal Neurons. J. Neurosci. 1997, 17, 5206–5220. [Google Scholar] [CrossRef]
- Davis, G.R., Jr.; McClellan, A.D. Long Distance Axonal Regeneration of Identified Lamprey Reticulospinal Neurons. Exp. Neurol. 1994, 127, 94–105. [Google Scholar] [CrossRef]
- Shifman, M.I.; Zhang, G.; Selzer, M.E. Delayed Death of Identified Reticulospinal Neurons after Spinal Cord Injury in Lampreys. J. Comp. Neurol. 2008, 510, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Jin, L.Q.; Sul, J.Y.; Haydon, P.G.; Selzer, M.E. Live Imaging of Regenerating Lamprey Spinal Axons. Neurorehabilit. Neural Repair 2005, 19, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Rodemer, W.; Lee, T.; Hu, J.; Selzer, M.E. The Effect of Axon Resealing on Retrograde Neuronal Death after Spinal Cord Injury in Lamprey. Brain Sci. 2018, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Rodemer, W.; Sinitsa, I.; Hu, J.; Selzer, M.E. Source of Early Regenerating Axons in Lamprey Spinal Cord Revealed by Wholemount Optical Clearing with Babb. Cells 2020, 9, 2427. [Google Scholar] [CrossRef]
- Busch, D.J.; Morgan, J.R. Synuclein Accumulation Is Associated with Cell-Specific Neuronal Death after Spinal Cord Injury. J. Comp. Neurol. 2012, 520, 1751–1771. [Google Scholar] [CrossRef]
- Jin, L.Q.; Zhang, G.; Jamison, C.J.; Takano, H.; Haydon, P.G.; Selzer, M.E. Axon Regeneration in the Absence of Growth Cones; Acceleration by Cyclic Amp. J. Comp. Neurol. 2009, 515, 295–312. [Google Scholar] [CrossRef]
- Jin, L.Q.; Pennise, C.R.; Rodemer, W.; Jahn, K.S.; Selzer, M.E. Protein Synthetic Machinery and Mrna in Regenerating Tips of Spinal Cord Axons in Lamprey. J. Comp. Neurol. 2016, 524, 3614–3640. [Google Scholar] [CrossRef] [Green Version]
- Rovainen, C.M.; Johnson, P.A.; Roach, E.A.; Mankovsky, J.A. Projections of Individual Axons in Lamprey Spinal Cord Determined by Tracings through Serial Sections. J. Comp. Neurol. 1973, 149, 193–202. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. Star: Ultrafast Universal Rna-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. Htseq—A Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential Expression Analysis for Sequence Count Data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using David Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. String V11: Protein-Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, S.; Sarkar, R.R. Comparison of Human Cell Signaling Pathway Databases--Evolution, Drawbacks and Challenges. Database 2015, 2015, bau126. [Google Scholar] [CrossRef] [Green Version]
- Bhullar, K.S.; Lagaron, N.O.; McGowan, E.M.; Parmar, I.; Jha, A.; Hubbard, B.P.; Rupasinghe, H.P.V. Kinase-Targeted Cancer Therapies: Progress, Challenges and Future Directions. Mol. Cancer 2018, 17, 48. [Google Scholar] [CrossRef]
- Ruby, C.L.; Major, R.J.; Hinrichsen, R.D. Regulation of Tissue Regeneration by the Circadian Clock. Eur. J. Neurosci. 2021, 53, 3576–3597. [Google Scholar] [CrossRef]
- Palmisano, I.; Danzi, M.C.; Hutson, T.H.; Zhou, L.; McLachlan, E.; Serger, E.; Shkura, K.; Srivastava, P.K.; Hervera, A.; Neill, N.O.; et al. Epigenomic Signatures Underpin the Axonal Regenerative Ability of Dorsal Root Ganglia Sensory Neurons. Nat. Neurosci. 2019, 22, 1913–1924. [Google Scholar] [CrossRef]
- Wang, S.; Smith, G.M.; Selzer, M.E.; Li, S. Emerging Molecular Therapeutic Targets for Spinal Cord Injury. Expert Opin. Targets 2019, 23, 787–803. [Google Scholar] [CrossRef]
- Becker, T.; Lieberoth, B.C.; Becker, C.G.; Schachner, M. Differences in the Regenerative Response of Neuronal Cell Populations and Indications for Plasticity in Intraspinal Neurons after Spinal Cord Transection in Adult Zebrafish. Mol. Cell Neurosci. 2005, 30, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Laramore, C.; Shifman, M.I. The Expression of Chemorepulsive Guidance Receptors and the Regenerative Abilities of Spinal-Projecting Neurons after Spinal Cord Injury. Neuroscience 2017, 341, 95–111. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, G.; Rodemer, W.; Jin, L.Q.; Shifman, M.; Selzer, M.E. The Role of Rhoa in Retrograde Neuronal Death and Axon Regeneration after Spinal Cord Injury. Neurobiol. Dis. 2017, 98, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogerson, S.M.; van Brummen, A.J.; Busch, D.J.; Allen, S.R.; Roychaudhuri, R.; Banks, S.M.; Klarner, F.G.; Schrader, T.; Bitan, G.; Morgan, J.R. Reducing Synuclein Accumulation Improves Neuronal Survival after Spinal Cord Injury. Exp. Neurol. 2016, 278, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Rodemer, W.; Hu, J.; Selzer, M.E.; Shifman, M.I. Heterogeneity in the Regenerative Abilities of Central Nervous System Axons within Species: Why Do Some Neurons Regenerate Better Than Others? Neural Regen. Res. 2020, 15, 996–1005. [Google Scholar] [PubMed]
- Willis, D.E.; van Niekerk, E.A.; Sasaki, Y.; Mesngon, M.; Merianda, T.T.; Williams, G.G.; Kendall, M.; Smith, D.S.; Bassell, G.J.; Twiss, J.L. Extracellular Stimuli Specifically Regulate Localized Levels of Individual Neuronal Mrnas. J. Cell Biol. 2007, 178, 965–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.M.; Berchtold, N.C.; Perreau, V.M.; Tu, C.H.; Jeon, N.L.; Cotman, C.W. Axonal Mrna in Uninjured and Regenerating Cortical Mammalian Axons. J. Neurosci. 2009, 29, 4697–4707. [Google Scholar] [CrossRef] [PubMed]
- Rotem, N.; Magen, I.; Ionescu, A.; Gershoni-Emek, N.; Altman, T.; Costa, C.J.; Gradus, T.; Pasmanik-Chor, M.; Willis, D.E.; Ben-Dov, I.Z.; et al. Als Along the Axons—Expression of Coding and Noncoding Rna Differs in Axons of Als Models. Sci. Rep. 2017, 7, 44500. [Google Scholar] [CrossRef] [Green Version]
- Mathur, C.; Johnson, K.R.; Tong, B.A.; Miranda, P.; Srikumar, D.; Basilio, D.; Latorre, R.; Bezanilla, F.; Holmgren, M. Demonstration of Ion Channel Synthesis by Isolated Squid Giant Axon Provides Functional Evidence for Localized Axonal Membrane Protein Translation. Sci. Rep. 2018, 8, 2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldsmith, C.S.; Bell-Pedersen, D. Diverse Roles for Mapk Signaling in Circadian Clocks. Adv. Genet. 2013, 84, 1–39. [Google Scholar]
- Dessauer, C.W.; Chen-Goodspeed, M.; Chen, J. Mechanism of Galpha I-Mediated Inhibition of Type V Adenylyl Cyclase. J. Biol. Chem. 2002, 277, 28823–28829. [Google Scholar] [CrossRef] [Green Version]
- Orosco, A.; Fromigue, O.; Hay, E.; Marie, P.J.; Modrowski, D. Dual Involvement of Protein Kinase C Delta in Apoptosis Induced by Syndecan-2 in Osteoblasts. J. Cell Biochem. 2006, 98, 838–850. [Google Scholar] [CrossRef]
- Roedig, J.; Kowald, L.; Juretschke, T.; Karlowitz, R.; Abhari, B.A.; Roedig, H.; Fulda, S.; Beli, P.; van Wijk, S.J. Usp22 Controls Necroptosis by Regulating Receptor-Interacting Protein Kinase 3 Ubiquitination. EMBO Rep. 2021, 22, e50163. [Google Scholar] [CrossRef]
- Anderson, S.; Bankier, A.T.; Barrell, B.G.; de Bruijn, M.H.; Coulson, A.R.; Drouin, J.; Eperon, I.C.; Nierlich, D.P.; Roe, B.A.; Sanger, F.; et al. Sequence and Organization of the Human Mitochondrial Genome. Nature 1981, 290, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Kocher, T.D. Complete Sequence of a Sea Lamprey (Petromyzon Marinus) Mitochondrial Genome: Early Establishment of the Vertebrate Genome Organization. Genetics 1995, 139, 873–887. [Google Scholar] [CrossRef]
- Fox, T.D. Mitochondrial Protein Synthesis, Import, and Assembly. Genetics 2012, 192, 1203–1234. [Google Scholar] [CrossRef] [Green Version]
- Rangaraju, V.; Lauterbach, M.; Schuman, E.M. Spatially Stable Mitochondrial Compartments Fuel Local Translation During Plasticity. Cell 2019, 176, 73–84.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cioni, J.M.; Lin, J.Q.; Holtermann, A.V.; Koppers, M.; Jakobs, M.A.H.; Azizi, A.; Turner-Bridger, B.; Shigeoka, T.; Franze, K.; Harris, W.A.; et al. Late Endosomes Act as Mrna Translation Platforms and Sustain Mitochondria in Axons. Cell 2019, 176, 56–72.e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cargnello, M.; Roux, P.P. Activation and Function of the Mapks and Their Substrates, the Mapk-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.J.; Timoshevskaya, N.; Ye, C.; Holt, C.; Keinath, M.C.; Parker, H.J.; Cook, M.E.; Hess, J.E.; Narum, S.R.; Lamanna, F.; et al. The Sea Lamprey Germline Genome Provides Insights into Programmed Genome Rearrangement and Vertebrate Evolution. Nat. Genet. 2018, 50, 270–277. [Google Scholar] [CrossRef]
- Johnson, G.L.; Nakamura, K. The C-Jun Kinase/Stress-Activated Pathway: Regulation, Function and Role in Human Disease. Biochim. Biophys. Acta 2007, 1773, 1341–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canovas, B.; Nebreda, A.R. Diversity and Versatility of P38 Kinase Signalling in Health and Disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 346–366. [Google Scholar] [CrossRef]
- Yue, J.; Lopez, J.M. Understanding Mapk Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bode, A.M.; Dong, Z. The Functional Contrariety of Jnk. Mol. Carcinog. 2007, 46, 591–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammarlund, M.; Nix, P.; Hauth, L.; Jorgensen, E.M.; Bastiani, M. Axon Regeneration Requires a Conserved Map Kinase Pathway. Science 2009, 323, 802–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, D.; Wu, Z.; Chisholm, A.D.; Jin, Y. The Dlk-1 Kinase Promotes Mrna Stability and Local Translation in C. Elegans Synapses and Axon Regeneration. Cell 2009, 138, 1005–1018. [Google Scholar] [CrossRef] [Green Version]
- Nix, P.; Hisamoto, N.; Matsumoto, K.; Bastiani, M. Axon Regeneration Requires Coordinate Activation of P38 and Jnk Mapk Pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 10738–10743. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ritchie, E.M.; Steinke, C.L.; Qi, C.; Chen, L.; Zheng, B.; Jin, Y. Activation of Map3k Dlk and Lzk in Purkinje Cells Causes Rapid and Slow Degeneration Depending on Signaling Strength. eLife 2021, 10, e63509. [Google Scholar] [CrossRef]
- Cheung, E.C.; Slack, R.S. Emerging Role for Erk as a Key Regulator of Neuronal Apoptosis. Sci. STKE 2004, 2004, PE45. [Google Scholar] [CrossRef]
- Huang, H.T.; Sun, Z.G.; Liu, H.W.; Ma, J.T.; Hu, M. Erk/Mapk and Pi3k/Akt Signal Channels Simultaneously Activated in Nerve Cell and Axon after Facial Nerve Injury. Saudi J. Biol. Sci. 2017, 24, 1853–1858. [Google Scholar]
- Zhang, W.; Liu, H.T. Mapk Signal Pathways in the Regulation of Cell Proliferation in Mammalian Cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Subramaniam, S.; Unsicker, K. Erk and Cell Death: Erk1/2 in Neuronal Death. FEBS J. 2010, 277, 22–29. [Google Scholar] [CrossRef]
- Yao, W.; Frie, M.; Pan, J.; Pak, K.; Webster, N.; Wasserman, S.I.; Ryan, A.F. C-Jun N-Terminal Kinase (Jnk) Isoforms Play Differing Roles in Otitis Media. BMC Immunol. 2014, 45, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, N.; Matsumoto, M.; Kogawa, M.; Atkins, G.J.; Findlay, D.M.; Fujikawa, T.; Oda, H.; Ogata, M. Critical Role of P38 Mapk for Regeneration of the Sciatic Nerve Following Crush Injury in Vivo. J. Neuroinflamm. 2013, 10, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchi, K.N.; Moore, J.G.; Hrushesky, W.J.; Sothern, R.B.; Rubin, N.H. Circadian Rhythm of Cellular Proliferation in the Human Rectal Mucosa. Gastroenterology 1991, 101, 410–415. [Google Scholar] [CrossRef]
- Garcia, M.N.; Barbeito, C.G.; Andrini, L.A.; Badran, A.F. Circadian Rhythm of DNA Synthesis and Mitotic Activity in Tongue Keratinocytes. Cell Biol. Int. 2001, 25, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, T.; Yamaguchi, S.; Mitsui, S.; Emi, A.; Shimoda, F.; Okamura, H. Control Mechanism of the Circadian Clock for Timing of Cell Division in Vivo. Science 2003, 302, 255–259. [Google Scholar] [CrossRef] [Green Version]
- Fagiani, F.; di Marino, D.; Romagnoli, A.; Travelli, C.; Voltan, D.; Mannelli, L.d.; Racchi, M.; Govoni, S.; Lanni, C. Molecular Regulations of Circadian Rhythm and Implications for Physiology and Diseases. Signal Transduct. Target. Ther. 2022, 7, 41. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Kar, A.N.; Samra, N.; Terenzio, M.; Patel, P.; Lee, S.J.; Miller, S.; Thames, E.; Jones, B.; Kawaguchi, R.; et al. A Ca(2+)-Dependent Switch Activates Axonal Casein Kinase 2alpha Translation and Drives G3bp1 Granule Disassembly for Axon Regeneration. Curr. Biol. 2020, 30, 4882–4895.e6. [Google Scholar] [CrossRef]
- Carballo, G.B.; Honorato, J.R.; de Lopes, G.P.F.; Spohr, T. A Highlight on Sonic Hedgehog Pathway. Cell Commun. Signal. 2018, 16, 11. [Google Scholar] [CrossRef]
- Dobbs, R.; Kalmanek, E.; Choe, S.; Harrington, D.A.; Stupp, S.I.; McVary, K.T.; Podlasek, C.A. Sonic Hedgehog Regulation of Cavernous Nerve Regeneration and Neurite Formation in Aged Pelvic Plexus. Exp. Neurol. 2019, 312, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.V.; Shanmugasundaram, J.; Sundaram, C.; Anandaraj, M.P. Activity of Novel Protein Kinase C and Distribution of Protein Kinase C Theta in Subcellular Fractions of Normal and Duchenne Muscular Dystrophic Muscle. Indian J. Biochem. Biophys. 2002, 39, 377–381. [Google Scholar]
- Takeda, K.; Harada, Y.; Watanabe, R.; Inutake, Y.; Ogawa, S.; Onuki, K.; Kagaya, S.; Tanabe, K.; Kishimoto, H.; Abe, R. Cd28 Stimulation Triggers Nf-Kappab Activation through the Carma1-Pkctheta-Grb2/Gads Axis. Int. Immunol. 2008, 20, 1507–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, R.H.; Wang, C.Y.; Yang, C.M. Nf-Kappab Signaling Pathways in Neurological Inflammation: A Mini Review. Front. Mol. Neurosci. 2015, 8, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dresselhaus, E.C.; Meffert, M.K. Cellular Specificity of Nf-Kappab Function in the Nervous System. Front. Immunol. 2019, 10, 1043. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, N.; Feng, Y.; Cai, M.; Li, Y.; Xu, X.; Zhang, H.; Yao, D. Protein Kinase C Theta (Prkcq) Affects Nerve Degeneration and Regeneration through the C-Fos and C-Jun Pathways in Injured Rat Sciatic Nerves. Exp. Neurol. 2021, 346, 113843. [Google Scholar] [CrossRef]
- Hu, J.; Rodemer, W.; Zhang, G.; Jin, L.Q.; Li, S.; Selzer, M.E. Chondroitinase Abc Promotes Axon Regeneration and Reduces Retrograde Apoptosis Signaling in Lamprey. Front. Cell Dev. Biol. 2021, 9, 653638. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank Accession No. | Forward Primer 5′–3′ | Reverse Primer 5′–3′ |
|---|---|---|---|
| mapk1 (erk) | XR_004402373 | GAGGGCGCCTACGGCATGGT | GCTGGTCGAGATAATGCTTC |
| mapk81 (jnk1) | XM_032949959 | GCATCTACATTCAGCTGGCA | TGTGCTAGTGCCTGATCGAC |
| mapk82 (jnk2) | XM_032969277 | TCACTTCTTCACCATGAGCG | GCAGTCAGACTTCACCACGA |
| mapk141 (p381) | XM_032969733 | GTAATTGGGCTGCTGGATGT | ATCAGCTTCAGCTGGTCGA |
| mapk142 (p382) | XM_032954129 | GACGTGAAGGAAGGCTTCTG | CAAATTGCTGGGTTTCAGGT |
| usp22 | XM_032970308 | CATGCATAGCGGAGGAAAAT | TTACACTTGCGTTCACGGAC |
| map3k2 | XM_032951177 | CTGAAAGGAACTCGCCTACG | TATTTGACAAACAGCGCCAC |
| csnk1e | XM_032967217 | CCTACCGCGAGAACAAAAAC | GGCTGGTGTGTGGGAGAGAG |
| Symbol | Description | Fold Change (log2FC) | FDR | GO-Terms |
|---|---|---|---|---|
| G > R | ||||
| alas2 | 5′-aminolevulinate synthase 2 | 27.2 | 2.05 × 10−18 | M., porphyrin biosynthesis |
| gpatch11 | G-patch domain containing 11 | 27.2 | 2.05 × 10−18 | chromosome, protein binding |
| eif2s1a | eukaryotic translation initiation factor 2 subunit alpha | 26.8 | 6.05 × 10−18 | C., protein synthesis |
| dnaja4 | Dnaj heat shock protein family (hsp40) member A4 | 26.1 | 2.05 × 10−18 | C., Hsp70 protein binding |
| eif4b (1 of 2) | eukaryotic translation initiation factor 4b | 25.8 | 2.03 × 10−17 | C., protein synthesis |
| actn4 (2 of 2) | actinin alpha 4 | 25.7 | 1.42 × 10−16 | N./C., F-actin cross-link |
| pigq | phosphatidylinositol glycan anchor biosynthesis class Q | 25.6 | 9.83 × 10−14 | P., PIGQ activity |
| leo1 | LEO1 homolog, Paf1/RNA polymerase II complex component | 25.6 | 9.36 × 10−14 | N./C., RNA transcription |
| sdha (2 of 2) | succinate dehydrogenase complex flavoprotein subunit a | 25.6 | 4.03 × 10−16 | M., electron transfer |
| mrpl12 | mitochondrial ribosomal protein L12 | 25.3 | 2.16 × 10−13 | M., RNA binding |
| G < R | ||||
| dcaf17 | DDB1- and CUL4-associated factor 17 | −27.9 | 1.42 × 10−16 | N./C., protein binding |
| hsdl2 | hydroxysteroid dehydrogenase-like 2 | −25.9 | 1.40 × 10−14 | M., oxidoreductase activity |
| cdk7 | cyclin-dependent kinase 7 | −25.7 | 2.47 × 10−14 | N./C., cell cycle, RNA transcription |
| oprm1 (2 of 3) | opioid receptor mu 1 | −10.4 | 1.05 × 10−2 | P./ER, receptor for endogenous opioids |
| mul1a | mitochondrial ubiquitin ligase activator of nfkb 1-A | −10.0 | 1.48 × 10−2 | M., protein ubiquitination |
| scfd2 | sec1 family domain containing 2 | −9.6 | 2.06 × 10−2 | P., protein transport |
| btk | Bruton tyrosine kinase | −9.4 | 2.33 × 10−2 | P./C./N., B lymphocyte signaling |
| prkcq | protein kinase C theta | −8.7 | 3.53 × 10−2 | P./C., T cell differentiation |
| rxfp3(1 of 2) | relaxin family peptide receptor 3 | −8.5 | 4.58 × 10−2 | P., inhibit cAMP accumulation |
| gpank1 | G-patch domain and ankyrin repeats 1 | −8.4 | 4.87 × 10−2 | N./C., nucleic acid binding |
| Symbol | Description | Fold Change (log2FC) | FDR | GO-Terms |
|---|---|---|---|---|
| G > S | ||||
| mb21d2a | Mab-21 domain containing 2 | 27.1 | 2.29 × 10−20 | C., cadherin binding |
| slc9a3 | solute carrier family 9 member A3 | 25.3 | 2.36 × 10−14 | P., solute:proton antiporter activity |
| stk19 | eukaryotic translation initiation factor 2 subunit alpha | 24.9 | 5.31 × 10−14 | N., N-ras signaling |
| kcna4 (1 of 2) | potassium voltage-gated channel subfamily A member 4 | 24.9 | 5.31 × 10−14 | P., transmembrane K+ transport |
| krt80 (11 of 11) | keratin 80 | 24.7 | 3.90 × 10−17 | C., protein binding |
| dph1 | diphthamide biosynthesis 1 | 24.5 | 1.25 × 10−13 | N./C., histidine modification |
| smchd1 | structural maintenance of chromosomes flexible hinge domain containing 1 | 24.3 | 1.89 × 10−13 | N., structural maintenance of chromosomes |
| rtca | RNA 3′-terminal phosphate cyclase | 24.3 | 2.10 × 10−14 | N., RNA processing |
| tmem167b | transmembrane protein 167B | 24.1 | 8.30 × 10−15 | Golgi, secretory pathway |
| inhbb | inhibin subunit beta B | 24.0 | 3.85 × 10−13 | N., growth factor activity |
| G < S | ||||
| ldlrad1 | low density lipoprotein receptor class A domain containing 1 | −25.6 | 1.17 × 10−14 | P./Ex., protein binding |
| selenbp1 | selenium binding protein 1 | −24.7 | 7.14 × 10−14 | N., intra-Golgi protein transport |
| f2r | coagulation factor II thrombin receptor | −24.2 | 1.95 × 10−13 | P./Ex./En, phosphoinositide hydrolysis |
| atp2b2 (2 of 3) | ATPase plasma membrane Ca2+ transporting 2 | −22.7 | 5.84 × 10−12 | P./Ex., intracellular Ca2+ levels |
| tectb | tectorin Beta | −22.7 | 5.84 × 10−12 | Ex./extracellular matrix |
| gskip | GSK3B interacting protein | −22.7 | 5.84 × 10−12 | N., anchoring for GSK3B and PKA |
| cyb561d1 | cytochrome B561 family member D1 | −22.7 | 5.84 × 10−12 | P., heme binding |
| wisp2 | cellular communication network factor 5 | −22.7 | 5.84 × 10−12 | N./Ex., insulin-like GF binding |
| ptrh2 | peptidyl-tRNA hydrolase 2 | −22.7 | 5.84 × 10−12 | M., hydrolase activity |
| scamp5 | secretory carrier membrane protein 5 | −11.9 | 6.33 × 10−3 | P,/En./Golgi, exocytosis |
| Symbol | Description | Fold Change (log2FC) | FDR | GO-Terms |
|---|---|---|---|---|
| R > S | ||||
| ptpra | protein tyrosine phosphatase receptor type A | 26.9 | 2.42 × 10−15 | P./Ex., focal adhesion |
| dnah9 | dynein axonemal heavy chain 9 | 26.8 | 2.43 × 10−15 | CS., force-producing |
| grm7 | glutamate metabotropic receptor 7 | 26.3 | 6.54 × 10−15 | P., axon movement |
| slc5a3a | solute carrier family 5 member 3a | 26.1 | 1.08 × 10−14 | P., inositol transport |
| morn5 | MORN repeat containing 5 | 25.5 | 3.64 × 10−14 | CS., protein binding |
| sdc2 | syndecan 2 | 25.3 | 6.75 × 10−14 | CS., protein binding |
| prkcq | protein kinase C theta | 14.2 | 8.18 × 10−5 | P./CS., protein kinase C activity |
| txnl4b | Thioredoxin-like 4B | 10.3 | 1.34 × 10−2 | N./C., pre-mRNA splicing |
| fam160b1 | FHF complex subunit hook interacting protein 2A | 10.2 | 1.42 × 10−2 | N./C., vesicle trafficking and/or fusion |
| prrc1 | proline rich coiled-coil 1 | 9.9 | 1.88 × 10−2 | Golgi, NA |
| R < S | ||||
| gmppab | GDP-mannose pyrophosphorylase A | −26.5 | 1.13 × 10−14 | Ex./N., inhibition of GMPPB |
| efcab6 | EF-hand calcium binding domain 6 | −25.6 | 8.32 × 10−14 | N./M., regulates the androgen receptor |
| col19a1 | collagen type XIX alpha 1 chain | −25.3 | 1.29 × 10−13 | Ex./En., cross-bridge between fibrils |
| raf1a | raf-1 proto-oncogene, serine/threonine kinase | −25.1 | 2.34 × 10−13 | P./M./N./C., MAPK/ERK ptw |
| gclc (1 of 2) | glutamate-cysteine ligase catalytic subunit | −25.0 | 2.34 × 10−13 | C./N., glutamate-cysteine ligase activity |
| siae (1 of 3) | sialic acid acetylesterase | −25.0 | 2.34 × 10−13 | Ex./Lys, acetyl ester removal |
| wdr48b | WD repeat domain 48 | −25.0 | 1.57 × 10−15 | C./N., DNA repair |
| trak1 | trafficking kinesin protein 1 | −25.0 | 2.73 × 10−13 | M./En., En.-to-Lys. trafficking |
| wdr37 | WD repeat domain 37 | −24.9 | 3.16 × 10−13 | N./CS., NA |
| ptprk | protein tyrosine phosphatase receptor type K | −24.8 | 3.30 × 10−13 | P., cell contact and adhesion |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, L.-Q.; Zhou, Y.; Li, Y.-S.; Zhang, G.; Hu, J.; Selzer, M.E. Transcriptomes of Injured Lamprey Axon Tips: Single-Cell RNA-Seq Suggests Differential Involvement of MAPK Signaling Pathways in Axon Retraction and Regeneration after Spinal Cord Injury. Cells 2022, 11, 2320. https://doi.org/10.3390/cells11152320
Jin L-Q, Zhou Y, Li Y-S, Zhang G, Hu J, Selzer ME. Transcriptomes of Injured Lamprey Axon Tips: Single-Cell RNA-Seq Suggests Differential Involvement of MAPK Signaling Pathways in Axon Retraction and Regeneration after Spinal Cord Injury. Cells. 2022; 11(15):2320. https://doi.org/10.3390/cells11152320
Chicago/Turabian StyleJin, Li-Qing, Yan Zhou, Yue-Sheng Li, Guixin Zhang, Jianli Hu, and Michael E. Selzer. 2022. "Transcriptomes of Injured Lamprey Axon Tips: Single-Cell RNA-Seq Suggests Differential Involvement of MAPK Signaling Pathways in Axon Retraction and Regeneration after Spinal Cord Injury" Cells 11, no. 15: 2320. https://doi.org/10.3390/cells11152320
APA StyleJin, L. -Q., Zhou, Y., Li, Y. -S., Zhang, G., Hu, J., & Selzer, M. E. (2022). Transcriptomes of Injured Lamprey Axon Tips: Single-Cell RNA-Seq Suggests Differential Involvement of MAPK Signaling Pathways in Axon Retraction and Regeneration after Spinal Cord Injury. Cells, 11(15), 2320. https://doi.org/10.3390/cells11152320