
A Comparative Analysis of Mucus Immunomodulatory Properties from Seven Marine Gastropods from the Mediterranean Sea

 ,
,  , , ,
, , ,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Morphological and Molecular Identification of Mollusca
2.3. Mucus Collection
2.4. Human Cell Lines Viability
2.5. Size and Morphology of Human Monocytes
2.6. RNA Extraction and Real Time qPCR
2.7. ELISA Assay
2.8. Immunoarray Analysis
2.9. Statistical Analyses
3. Results
3.1. Species Identification
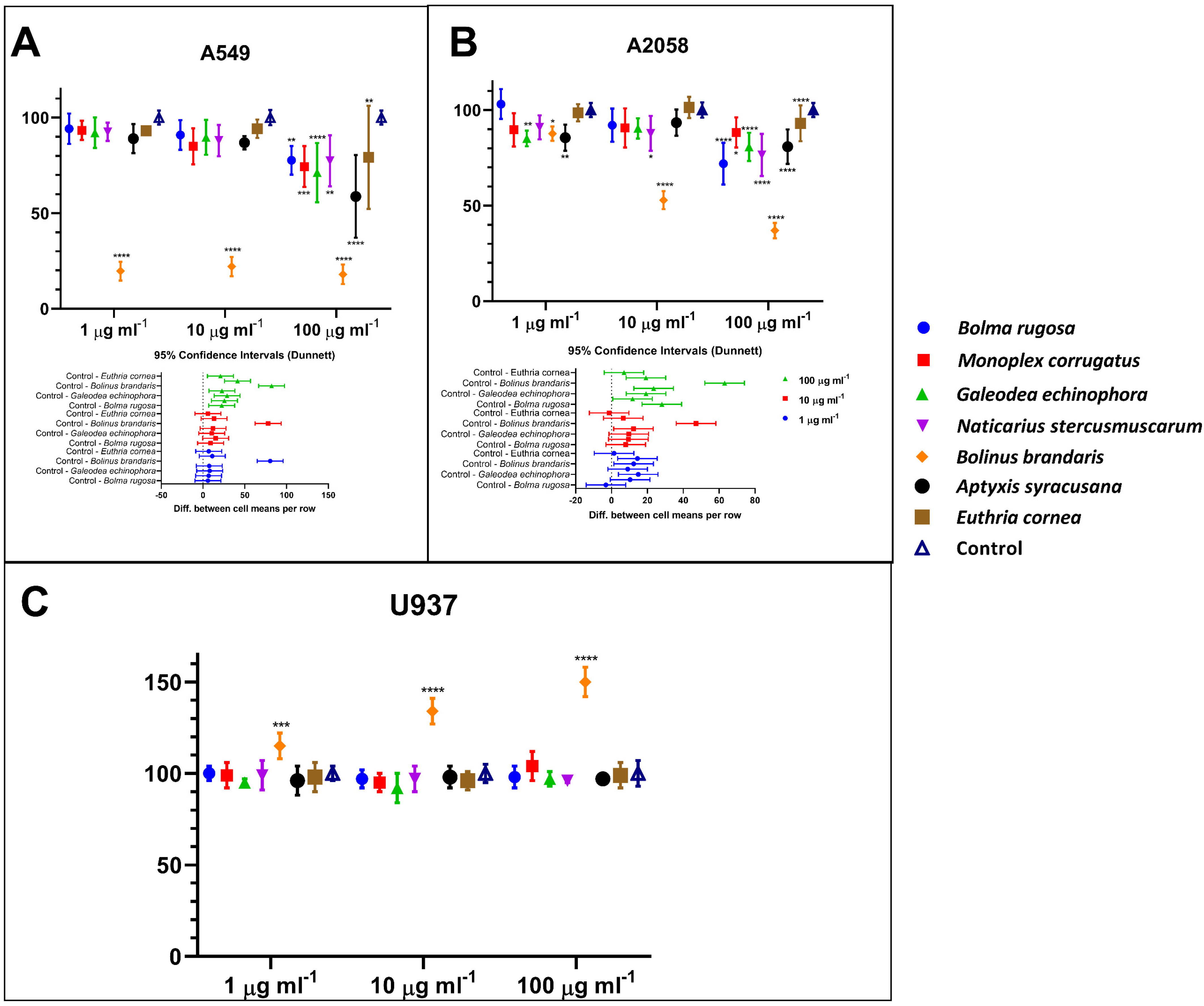
3.2. Human Cell Viability Assessment
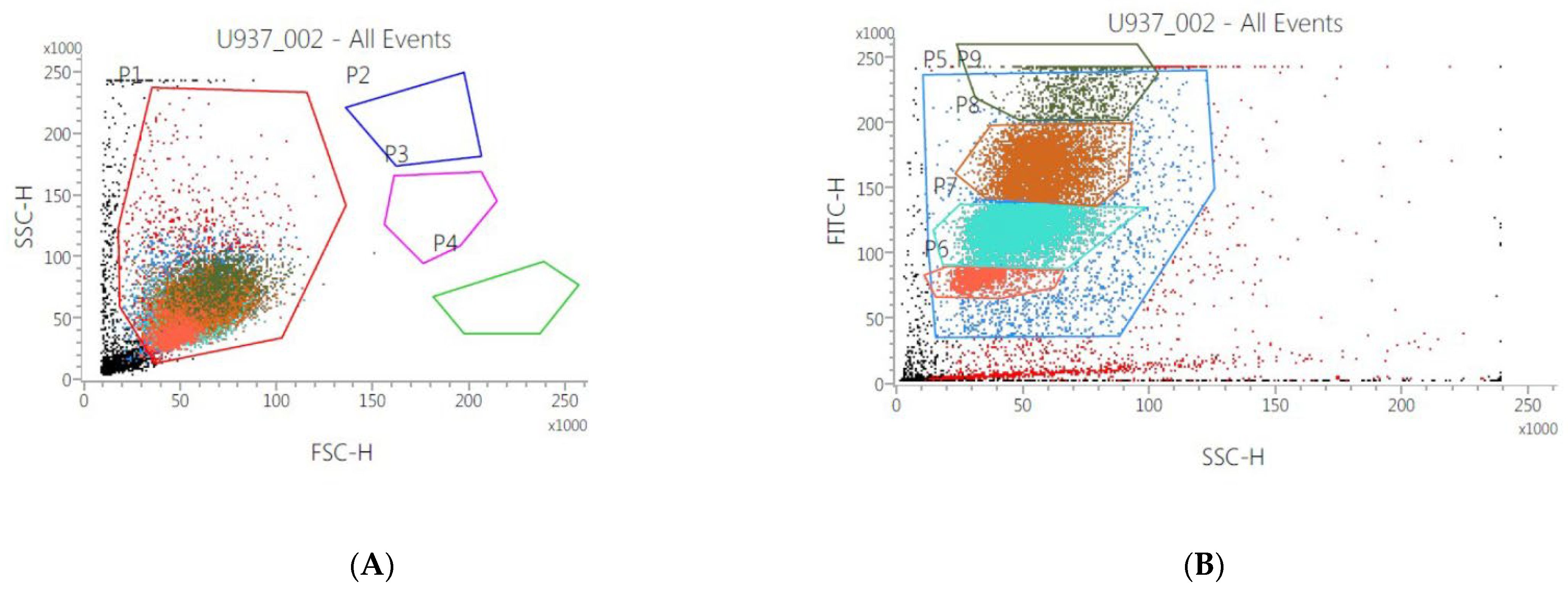
3.3. Morphological Changes of Monocytes
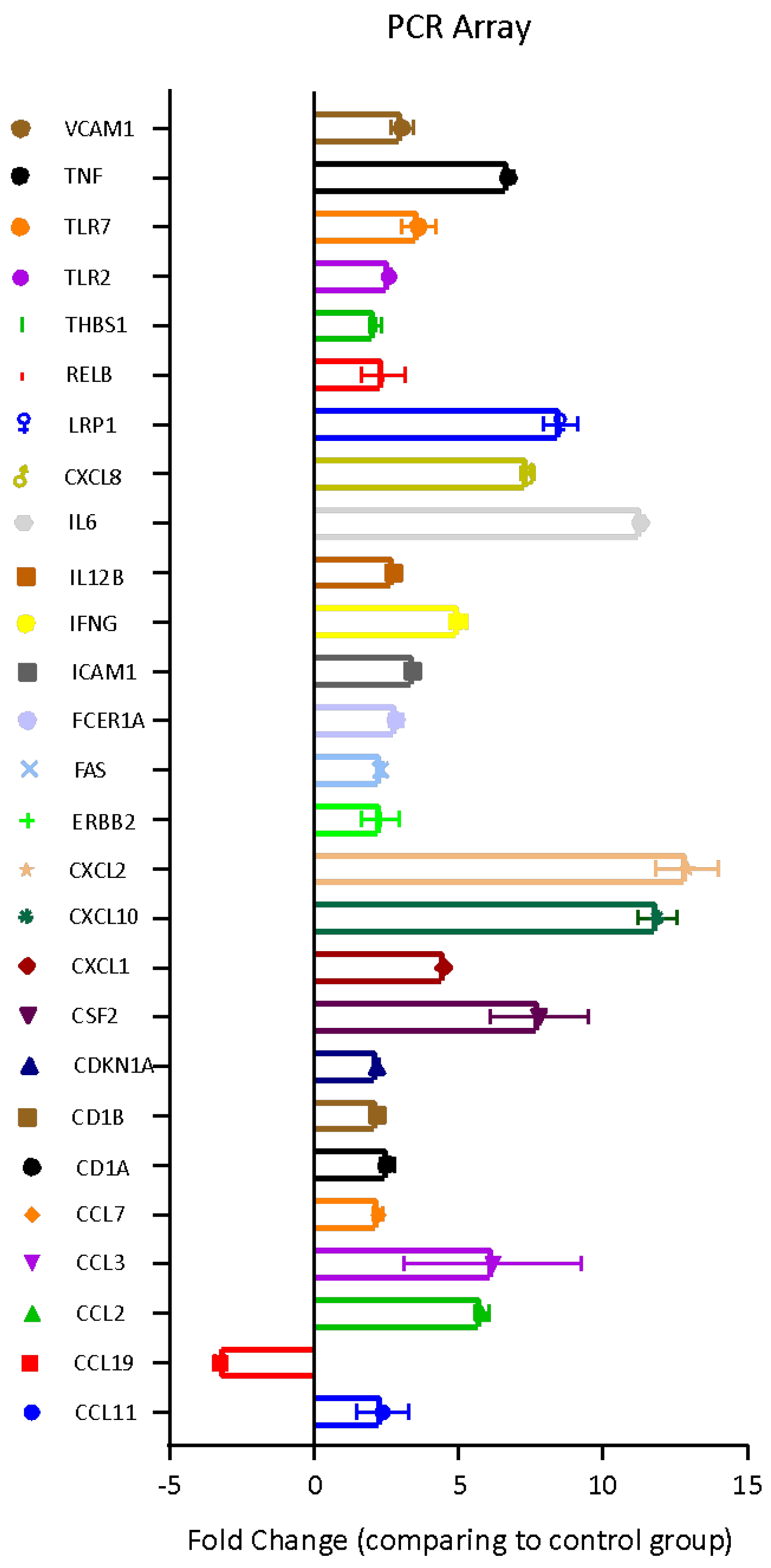
3.4. Bbm-Induced Gene Expression in U937 Cells
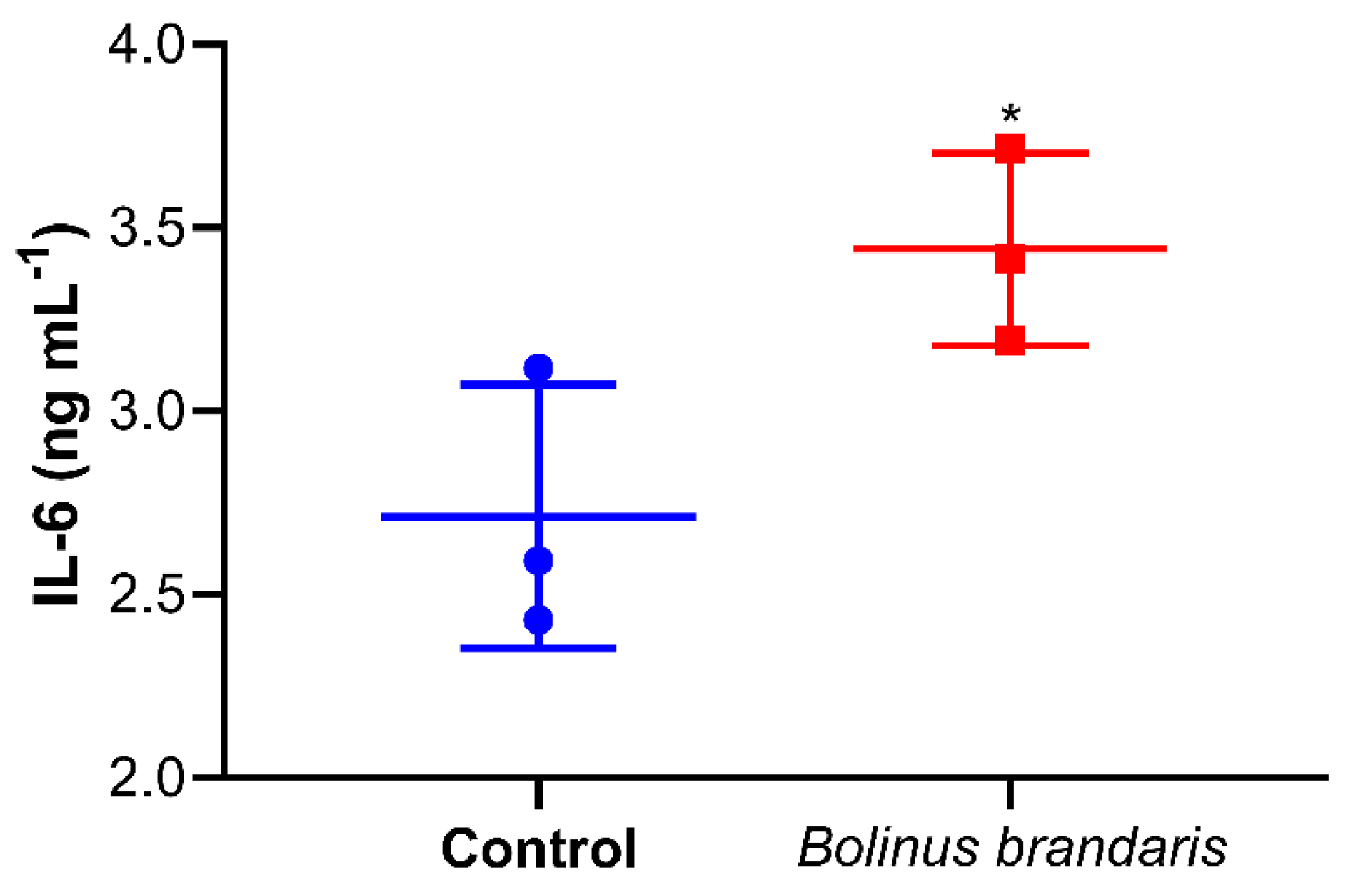
3.5. Bbm-Induced Interleukin IL-6 Release in U937 Cells
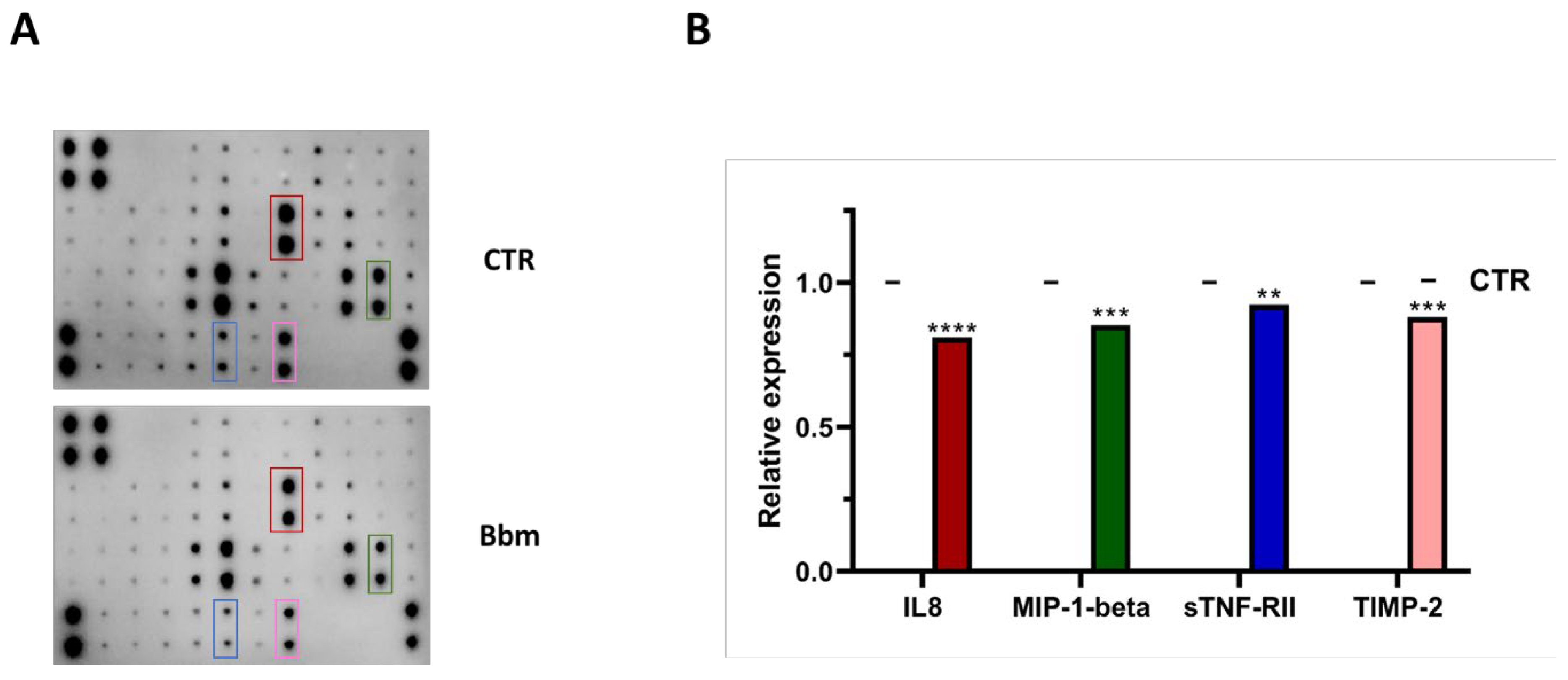
3.6. Bbm-Induced Inflammation Mediators Release in U937 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Greiner, W.; Patel, K.; Crossman-Barnes, C.-J.; Rye-Andersen, T.V.; Hvid, C.; Vandebrouck, T. High-Expenditure Disease in the EU-28: Does Drug Spend Correspond to Clinical and Economic Burden in Oncology, Autoimmune Disease and Diabetes? Pharm. Open 2021, 5, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Costello, M.; Chaudhary, C. Marine Biodiversity, Biogeography, Deep-Sea Gradients, and Conservation. Curr. Biol. 2017, 27, R511–R527. [Google Scholar] [CrossRef] [PubMed]
- Benkendorff, K.; Rudd, D.; Nongmaithem, B.D.; Liu, L.; Young, F.; Edwards, V.; Avila, C.; Abbott, C.A. Are the Traditional Medical Uses of Muricidae Molluscs Substantiated by Their Pharmacological Properties and Bioactive Compounds? Mar. Drugs 2015, 13, 5237–5275. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, B.; Shah, M.; Choi, S. Oceans as a Source of Immunotherapy. Mar. Drugs 2019, 17, 282. [Google Scholar] [CrossRef] [Green Version]
- McGettigan, S.E.; Debes, G.F. Immunoregulation by antibody secreting cells in inflammation, infection, and cancer. Immunol. Rev. 2021, 303, 103–118. [Google Scholar] [CrossRef]
- Abraham, W.M.; Bourdelais, A.J.; Sebater, J.R.; Ahmed, A.; Lee, T.A.; Serebriakov, I.; Baden, D.G. Airway responses to aerosolized brevetoxins in an animal model of asthma. Am. J. Respir. Crit. Care Med. 2005, 171, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Keeler, D.M.; Grandal, M.K.; McCall, J.R. Brevenal, a Marine Natural Product, is Anti-Inflammatory and an Immunomodulator of Macrophage and Lung Epithelial Cells. Mar. Drugs 2019, 17, 184. [Google Scholar] [CrossRef] [Green Version]
- Ponder, W.F.; Lindberg, D.R.; Ponder, J.M. Shell, Body, and Muscles. In Biology and Evolution of the Mollusca; Ponder, W.F., Lindberg, D.R., Ponder, J.M., Eds.; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Zannella, C.; Mosca, F.; Mariani, F.; Franci, G.; Folliero, V.; Galdiero, M.; Tiscar, P.G.; Galdiero, M. Microbial Diseases of Bivalve Mollusks: Infections, Immunology and Antimicrobial Defense. Mar. Drugs. 2017, 15, 182. [Google Scholar] [CrossRef]
- Gomez, D.; Sunyer, O.; Salinas, I. The mucosal immune system of fish: The evolution of tolerating commensals while fighting pathogens. Fish Shellfish Immunol. 2013, 35, 1729–1739. [Google Scholar] [CrossRef] [Green Version]
- Schröder, K.; Bosch, T.C.G. The Origin of Mucosal Immunity: Lessons from the Holobiont Hydra. mBio 2016, 7, e01184-16. [Google Scholar] [CrossRef] [Green Version]
- Dash, S.; Das, S.K.; Samal, J.; Thatoi, H.N. Epidermal mucus, a major determinant in fish health: A review. Iran. J. Vet. Res. 2018, 19, 72–81. [Google Scholar]
- Stabili, L. The mucus of marine invertebrates Cnidarians, polychaetes, and echinoderms as case studies. In Enzymatic Technologies for Marine Polysaccharides; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Salinas, I.; Fernández-Montero, Á.; Yang Ding, J.; Sunyer, O. Mucosal immunoglobulins of teleost fish: A decade of advances. Dev. Comp. Immunol. 2021, 121, 104079. [Google Scholar] [CrossRef]
- Tiralongo, F.; Messina, G.; Lombardo, B.M.; Longhitano, L.; Volti, G.L.; Tibullo, D. Skin Mucus of Marine Fish as a Source for the Development of Antimicrobial Agents. Front. Mar. Sci. 2020, 7, 541853. [Google Scholar] [CrossRef]
- Bakshani, C.R.; Morales-García, A.L.; Althaus, M.; Wilcox, M.D.; Pearson, J.P.; Bythell, J.C.; Burgess, J.G. Evolutionary conservation of the antimicrobial function of mucus: A first defence against infection. NPJ Biofilms Microb. 2020, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Bishani, A.; Chernolovskaya, E.L. Activation of Innate Immunity by Therapeutic Nucleic Acids. Int. J. Mol. Sci. 2021, 22, 13360. [Google Scholar] [CrossRef]
- Gonzalez, H.; Hagerling, C.; Werb, Z. Roles of the immune system in cancer: From tumor initiation to metastatic progression. Genes Dev. 2018, 32, 1267–1284. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Ford, S.E. Infectious diseases of marine molluscs and host responses as revealed by genomic tools. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150206. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, M.; Ueyama, D.; Kobayashi, R. The advantage of mucus for adhesive locomotion in gastropods. J. Theor Biol. 2014, 353, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Kesimer, M.; Sheehan, J.K. Mass spectrometric analysis of mucin core proteins. Methods Mol. Biol. 2012, 842, 67–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossignani, T.; Ardovini, R. Malacologia Mediterranea: Atlante Delle Conchiglie Del Mediterraneo: 7500 Foto a Colori; L’informatore Piceno: Ancona, Italy, 2011; ISBN 9788886070263. [Google Scholar]
- Gofas, S.; Moreno Lampreave, D.; Salas, C. Moluscos Marinos de Andalucía; Universidad de Málaga, Ed.; Servicio de Publicaciones e Intercambio Cientifico: Málaga, Spain, 2011; Volume 1, pp. 1–342. ISBN 9788497473569. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Meyer, C.P. Molecular Systematics of Cowries (Gastropoda: Cypraeidae) and Diversification Patterns in the Tropics. Biol. J. Linn. Soc. 2003, 79, 401–459. [Google Scholar] [CrossRef] [Green Version]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database Indexing for Production MegaBLAST Searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- World Register of Marine Species. Available online: http://www.marinespecies.org (accessed on 11 May 2022).
- Tachapuripunya, V.; Roytrakul, S.; Chumnanpuen, P.; E-kobon, T. Unveiling Putative Functions of Mucus Proteins and Their Tryptic Peptides in Seven Gastropod Species Using Comparative Proteomics and Machine Learning-Based Bioinformatics Predictions. Molecules 2021, 26, 3475. [Google Scholar] [CrossRef]
- Couto, D.R.; Bouchet, P.; Kantor, Y.I.; Simone, L.R.; Giribet, G. A Multilocus Molecular Phylogeny of Fasciolariidae (Neogastropoda: Buccinoidea). Mol. Phylogenet. Evol. 2016, 99, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Giribet, G.; Okusu, A.; Lindgren, A.R.; Huff, S.W.; Schrödl, M.; Nishiguchi, M.K. Evidence for a Clade Composed of Molluscs with Serially Repeated Structures: Monoplacophorans Are Related to Chitons. Proc. Natl. Acad. Sci. USA 2006, 103, 7723–7728. [Google Scholar] [CrossRef] [Green Version]
- Cunha, R.L.; Grande, C.; Zardoya, R. Neogastropod Phylogenetic Relationships Based on Entire Mitochondrial Genomes. BMC Evol. Biol. 2009, 9, 210. [Google Scholar] [CrossRef] [Green Version]
- Kantor, Y.I.; Fedosov, A.E.; Kosyan, A.R.; Puillandre, N.; Sorokin, P.A.; Kano, Y.; Clark, R.; Bouchet, P. Molecular Phylogeny and Revised Classification of the Buccinoidea (Neogastropoda). Zool. J. Linn. Soc. 2022, 194, 789–857. [Google Scholar] [CrossRef]
- Osca, D.; Templado, J.; Zardoya, R. Caenogastropod Mitogenomics. Mol. Phylogenet. Evol. 2015, 93, 118–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strong, E.E.; Puillandre, N.; Beu, A.G.; Castelin, M.; Bouchet, P. Frogs and Tuns and Tritons—A Molecular Phylogeny and Revised Family Classification of the Predatory Gastropod Superfamily Tonnoidea (Caenogastropoda). Mol. Phylogenet. Evol. 2019, 130, 18–34. [Google Scholar] [CrossRef]
- Williams, S.T.; Ozawa, T. Molecular Phylogeny Suggests Polyphyly of Both the Turban Shells (Family Turbinidae) and the Superfamily Trochoidea (Mollusca: Vetigastropoda). Mol. Phylogenet. Evol. 2006, 39, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Uribe, J.E.; Kano, Y.; Templado, J.; Zardoya, R. Mitogenomics of Vetigastropoda: Insights into the Evolution of Pallial Symmetry. Zool. Scr. 2016, 45, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Huelsken, T.; Marek, C.; Schreiber, S.; Schmidt, I.; Hollmann, M. The Naticidae (Mollusca: Gastropoda) of Giglio Island (Tuscany, Italy): Shell Characters, Live Animals, and a Molecular Analysis of Egg Masses. Zootaxa 2008, 1770, 1–40. [Google Scholar] [CrossRef]
- Layton, K.K.; Martel, A.L.; Hebert, P.D. Patterns of DNA Barcode Variation in Canadian Marine Molluscs. PLoS ONE 2014, 9, e95003. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Herre, J.; Brown, G.D.; Gordon, S. The potential for Toll-like receptors to collaborate with other innate immune receptors. Immunology 2004, 112, 521–530. [Google Scholar] [CrossRef]
- Raman, D.; Baugher, P.J.; Thu, Y.M.; Richmond, A. Role of chemokines in tumor growth. Cancer Lett. 2007, 256, 137–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Crijns, H.; Vanheule, V.; Proost, P. Targeting Chemokine-Glycosaminoglycan Interactions to Inhibit Inflammation. Front. Immunol. 2020, 11, 483. [Google Scholar] [CrossRef]
- Her, N.-G.; Kesari, S.; Nurmemmedov, E. Thrombospondin-1 counteracts the p97 inhibitor CB-5083 in colon carcinoma cells. Cell Cycle 2020, 19, 1590–1601. [Google Scholar] [CrossRef] [PubMed]
- Leslie, P.L.; Franklin, D.A.; Liu, Y.; Zhang, Y. p53 Regulates the Expression of LRP1 and Apoptosis through a Stress Intensity-Dependent MicroRNA Feedback Loop. Cell Rep. 2018, 24, 1484–1495. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Wang, Q.; Ke, Y.; Lin, J. Neutrophil Function in an Inflammatory Milieu of Rheumatoid Arthritis. J. Immunol. Res. 2018, 2018, 8549329. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.J.; Douglas, S.D.; Lai, J.P.; Pleasure, D.E.; Li, Y.; Williams, M.; Bannerman, P.; Song, L.; Ho, W.Z. Interleukin-1beta stimulates macrophage inflammatory protein-1alpha and -1beta expression in human neuronal cells (NT2-N). J. Neurochem. 2003, 84, 997–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gough, P.; Myles, I.A. Tumor Necrosis Factor Receptors: Pleiotropic Signaling Complexes and Their Differential Effects. Front. Immunol. 2020, 11, 585880. [Google Scholar] [CrossRef] [PubMed]
- Guedez, L.; Jensen-Taubman, S.; Bourboulia, D.; Kwityn, C.J.; Wei, B.; Caterina, J.; Stetler-Stevenson, W.G. TIMP-2 targets tumor-associated myeloid suppressor cells with effects in cancer immune dysfunction and angiogenesis. J. Immunother. 2012, 35, 502–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claesson-Welsh, L.; Dejana, E.; McDonald, D.M. Permeability of the Endothelial Barrier: Identifying and Reconciling Controversies. Trends Mol. Med. 2021, 27, 314–331. [Google Scholar] [CrossRef]
- Valacca, C.; Tassone, E.; Mignatti, P. TIMP-2 Interaction with MT1-MMP Activates the AKT Pathway and Protects Tumor Cells from Apoptosis. PLoS ONE 2015, 10, e0136797. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Depth Range | Coordinates | P | T | bp | Voucher | GenBank |
|---|---|---|---|---|---|---|---|
| Caenogastropoda | |||||||
| Aptyxis syracusana (Linnaeus, 1758) | 20–30 m | 40.8181 N, 14.1195 E | dgLCO-1490 dgHCO-2198 | 45 | 701 | 1387ML49E | ON926803 |
| Bolinus brandaris (Linnaeus, 1758) | 75–150 m | 40.8067 N, 14.1401 E | dgLCO-1490 dgHCO-2198 | 45 | 683 | 1183ML113B | ON926804 |
| Euthria cornea (Linnaeus, 1758) | 20 m | 40.8181 N, 14.1195 E | dgLCO-1490 dgHCO-2198 | 42 | 697 | 1168ML10B | ON934996 |
| Galeodea echinophora (Linnaeus, 1758) | 100–350 m | 40.7503 N, 14.1128 E | LCO-1490 HCO-2198 | 45 | 682 | 248ML107D | ON926805 |
| Monoplex corrugatus (Lamarck, 1816) | 50–100 m | 40.8067 N, 14.1401 E | LCO-1490 HCO-2198 | 45 | 608 | 464ML19A | ON930589 |
| Naticarius stercusmuscarum (Gmelin, 1791) | 100–250 m | 40.7503 N, 14.1128 E | dgLCO-1490 dgHCO-2198 | 42 | 684 | 460ML18D | ON926806 |
| Vetigastropoda | |||||||
| Bolma rugosa (Linnaeus, 1767) | 20–30 m | 40.8181 N, 14.1195 E | dgLCO-1490 dgHCO-2198 | 42 | 663 | 2894ML197A | ON926807 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sansone, C.; Balestra, C.; Pistelli, L.; Del Mondo, A.; Osca, D.; Brunet, C.; Crocetta, F. A Comparative Analysis of Mucus Immunomodulatory Properties from Seven Marine Gastropods from the Mediterranean Sea. Cells 2022, 11, 2340. https://doi.org/10.3390/cells11152340
Sansone C, Balestra C, Pistelli L, Del Mondo A, Osca D, Brunet C, Crocetta F. A Comparative Analysis of Mucus Immunomodulatory Properties from Seven Marine Gastropods from the Mediterranean Sea. Cells. 2022; 11(15):2340. https://doi.org/10.3390/cells11152340
Chicago/Turabian StyleSansone, Clementina, Cecilia Balestra, Luigi Pistelli, Angelo Del Mondo, David Osca, Christophe Brunet, and Fabio Crocetta. 2022. "A Comparative Analysis of Mucus Immunomodulatory Properties from Seven Marine Gastropods from the Mediterranean Sea" Cells 11, no. 15: 2340. https://doi.org/10.3390/cells11152340
APA StyleSansone, C., Balestra, C., Pistelli, L., Del Mondo, A., Osca, D., Brunet, C., & Crocetta, F. (2022). A Comparative Analysis of Mucus Immunomodulatory Properties from Seven Marine Gastropods from the Mediterranean Sea. Cells, 11(15), 2340. https://doi.org/10.3390/cells11152340