
Membrane Properties of Human Induced Pluripotent Stem Cell-Derived Cultured Red Blood Cells

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Material
2.2. Cultivation of cRBC_iPSCs
2.3. Phenotype and Maturation Stage of cRBC_iPSCs
2.4. Electron Microscopy
2.5. Osmotic Resistance
2.6. Deformability
2.7. Immunofluorescence
2.8. Lipidomics Analysis
2.9. Blood Group Phenotyping
2.10. Blood Group Genotyping
2.11. Statistical Analysis
3. Results
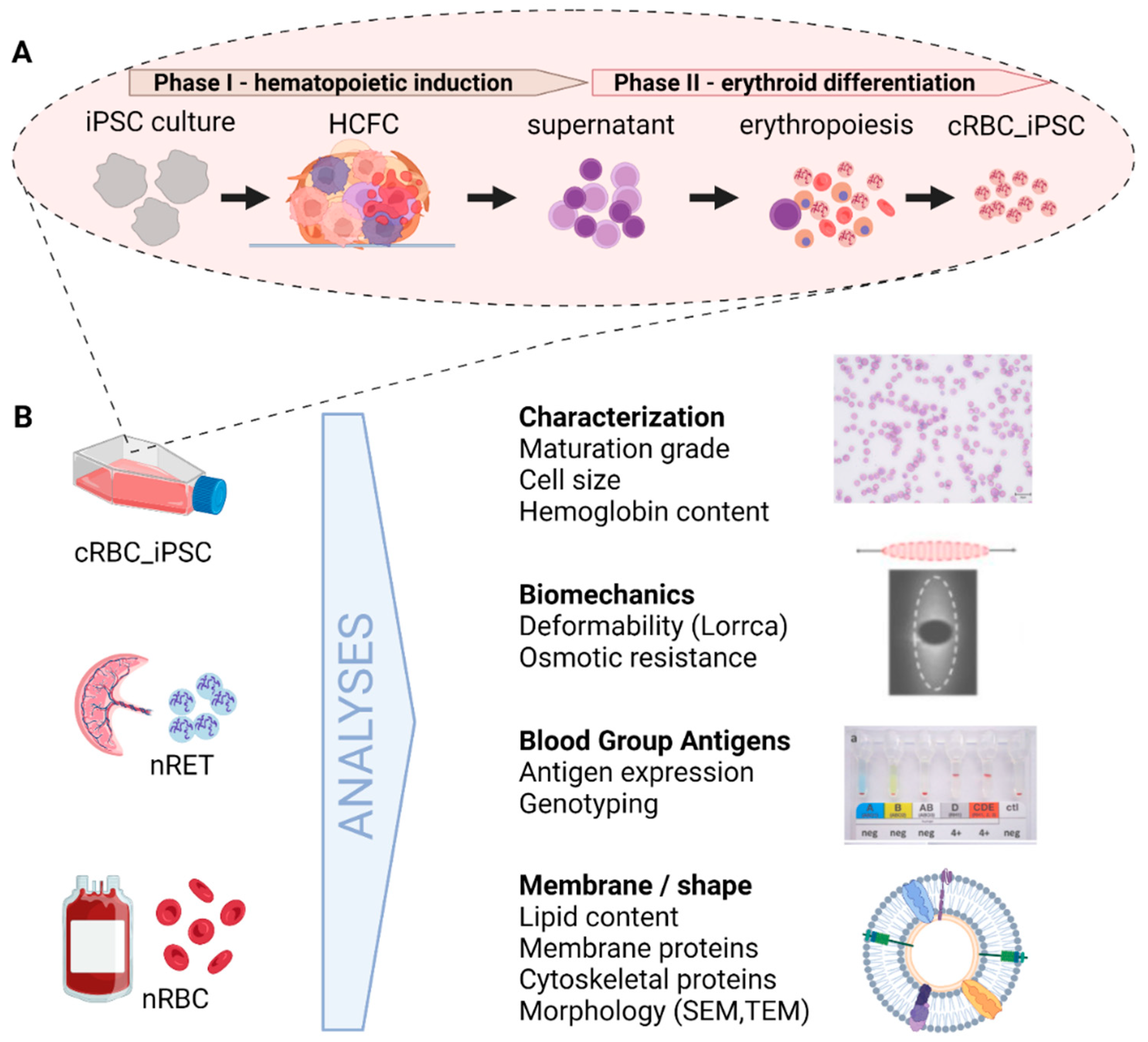
3.1. Generation of cRBC_iPSCs
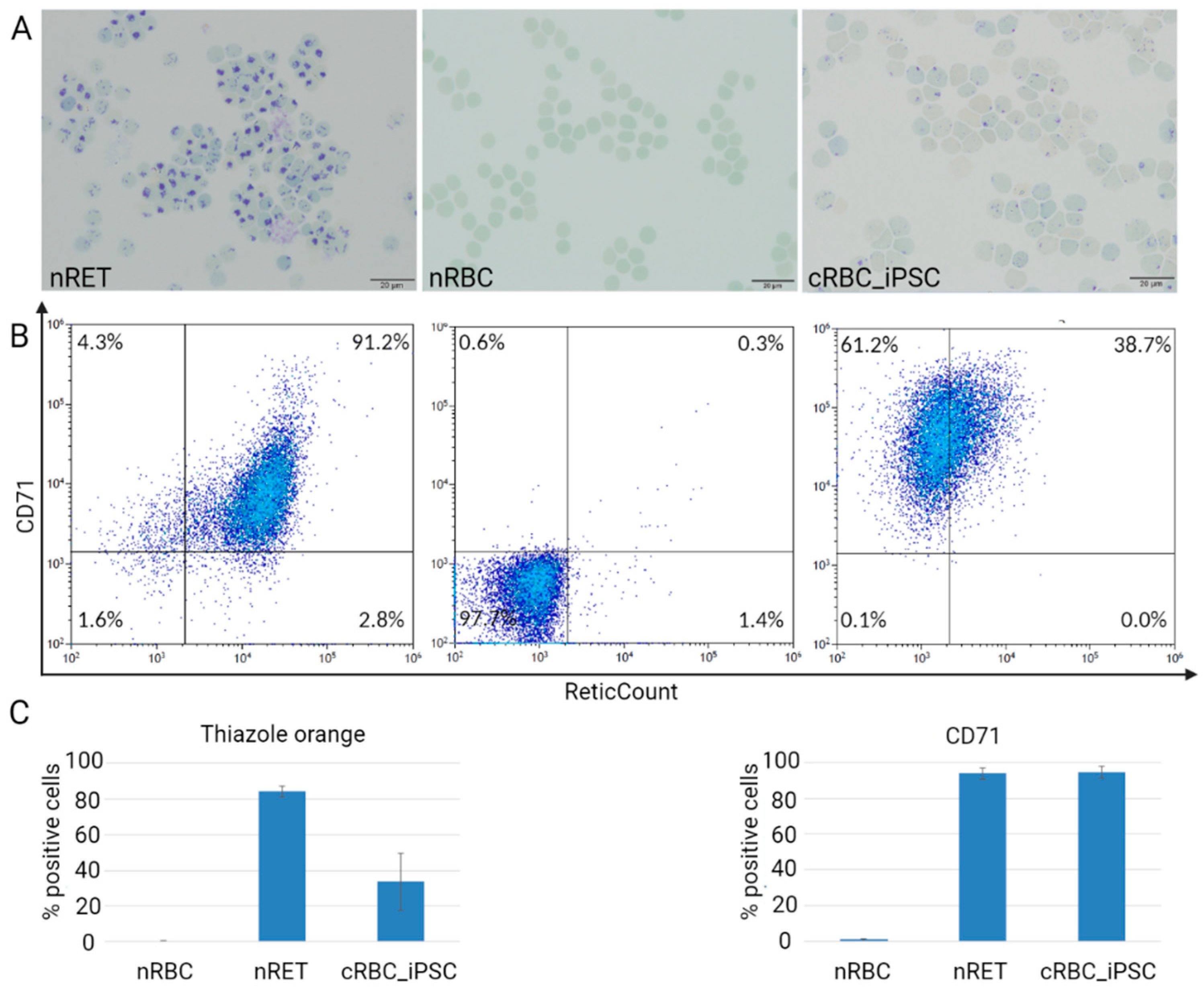
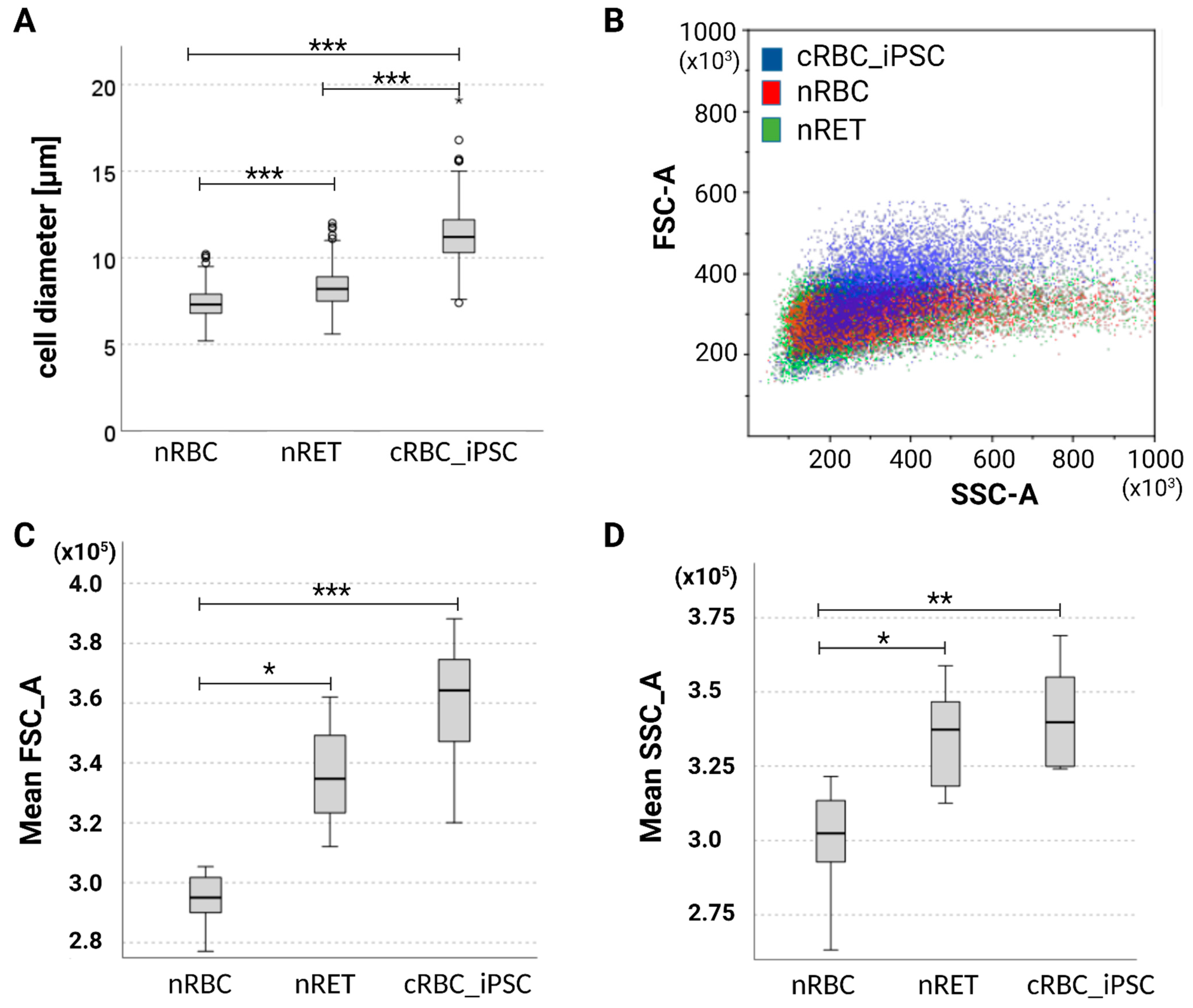
3.2. Maturation Stage of cRBC_iPSCs, Cell Size, and Hemoglobin Content
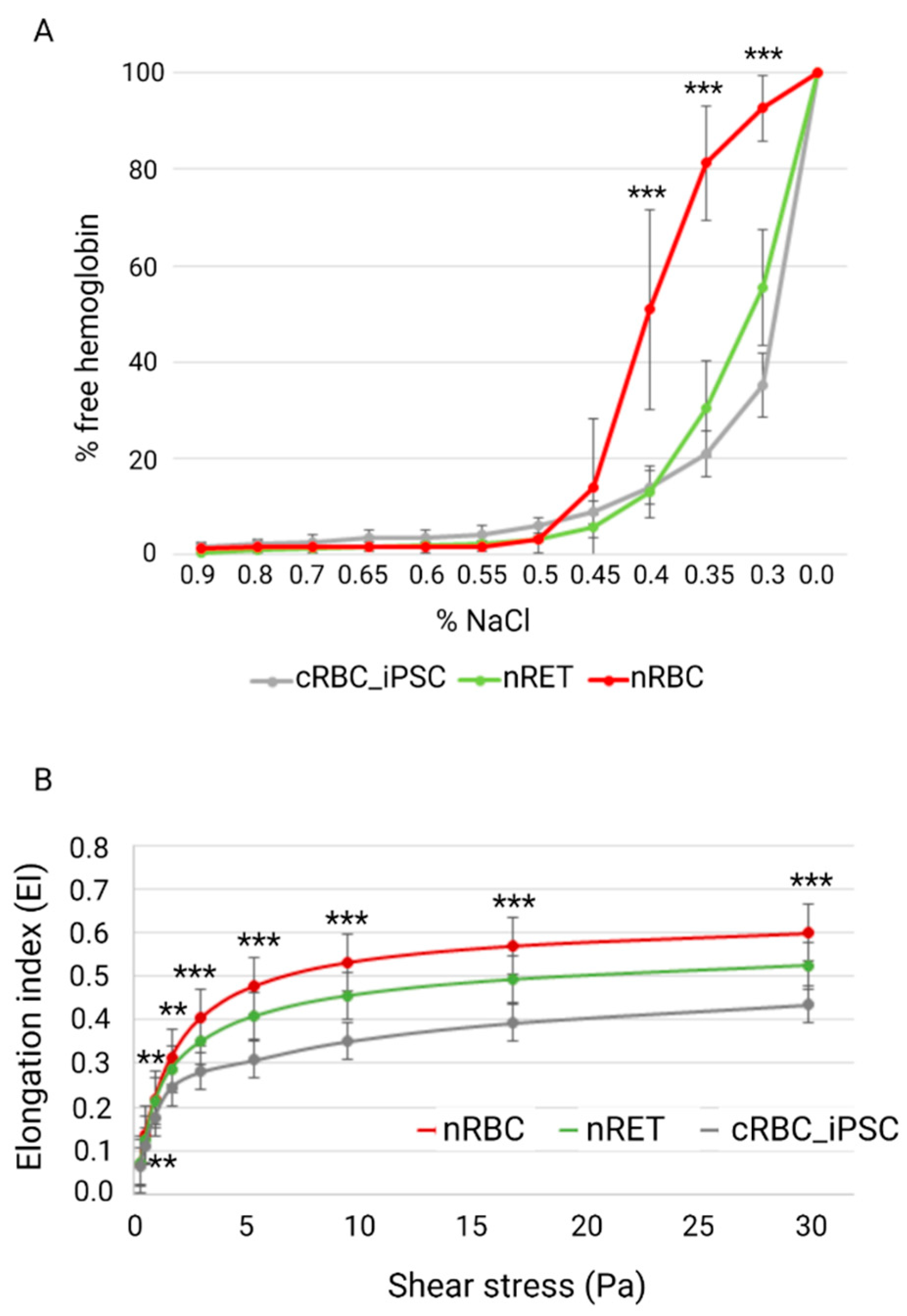
3.3. Biomechanical Characterization of cRBC_iPSCs
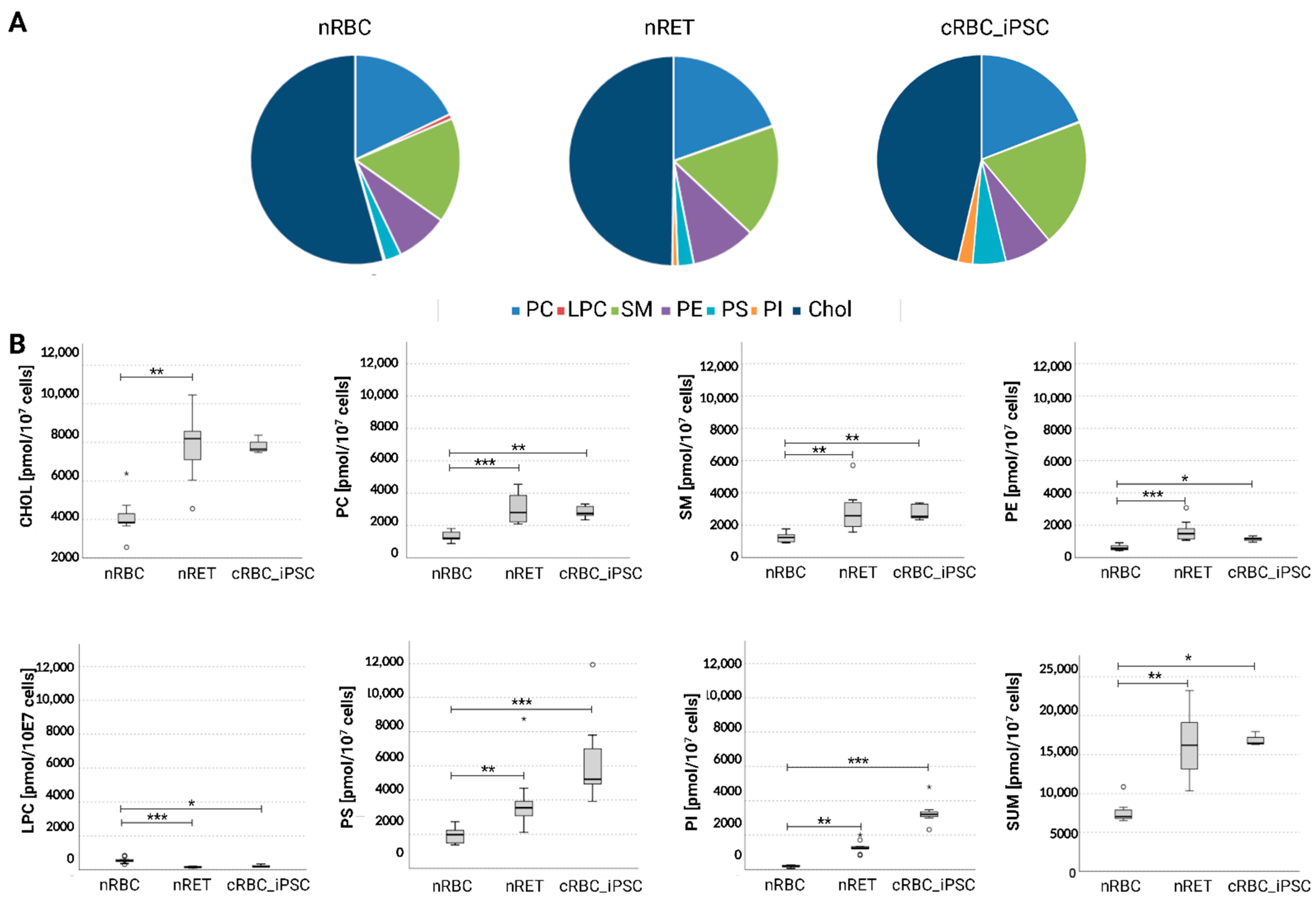
3.4. Membrane Lipid Composition of cRBC_iPSCs
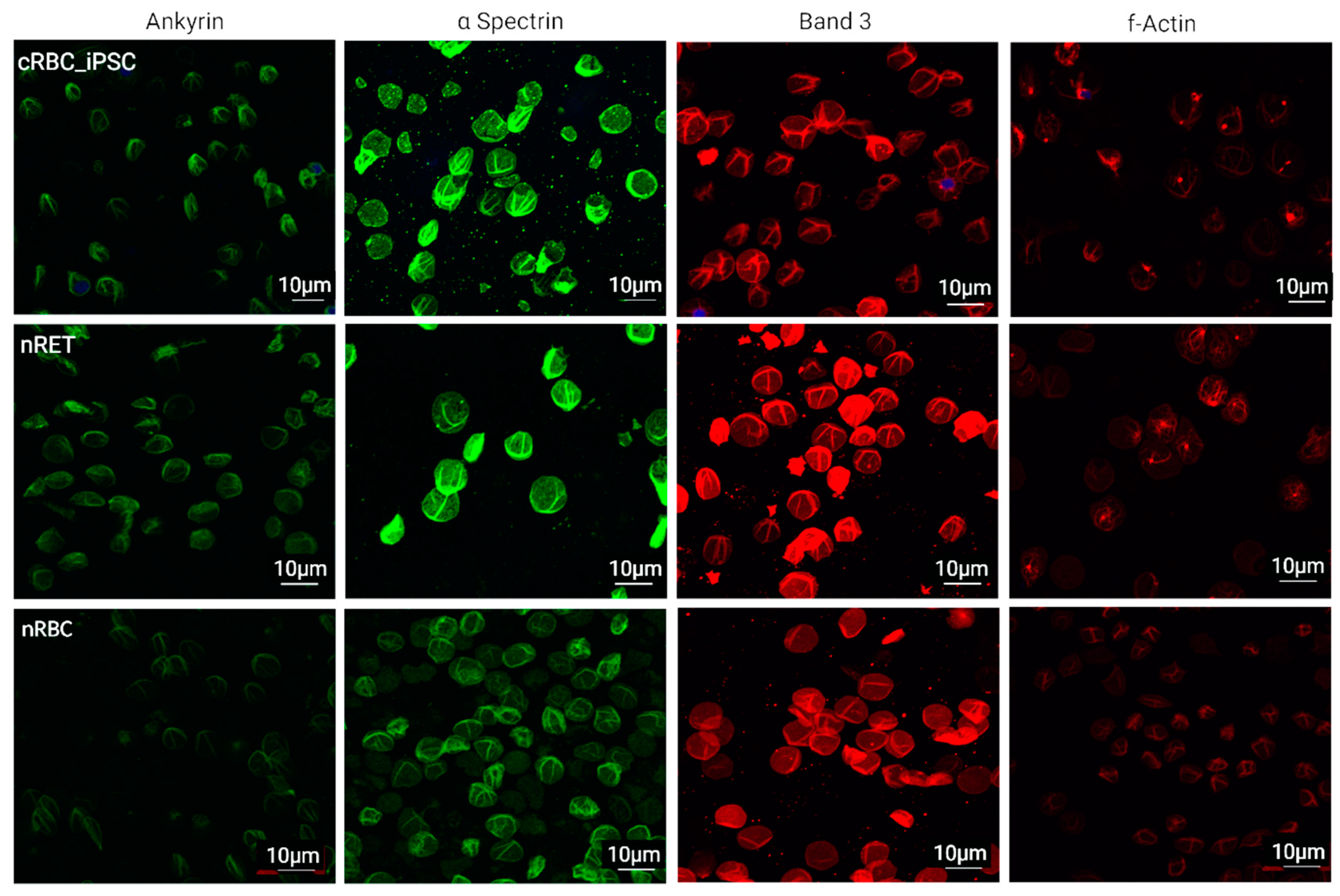
3.5. Cytoskeletal and Membrane Proteins
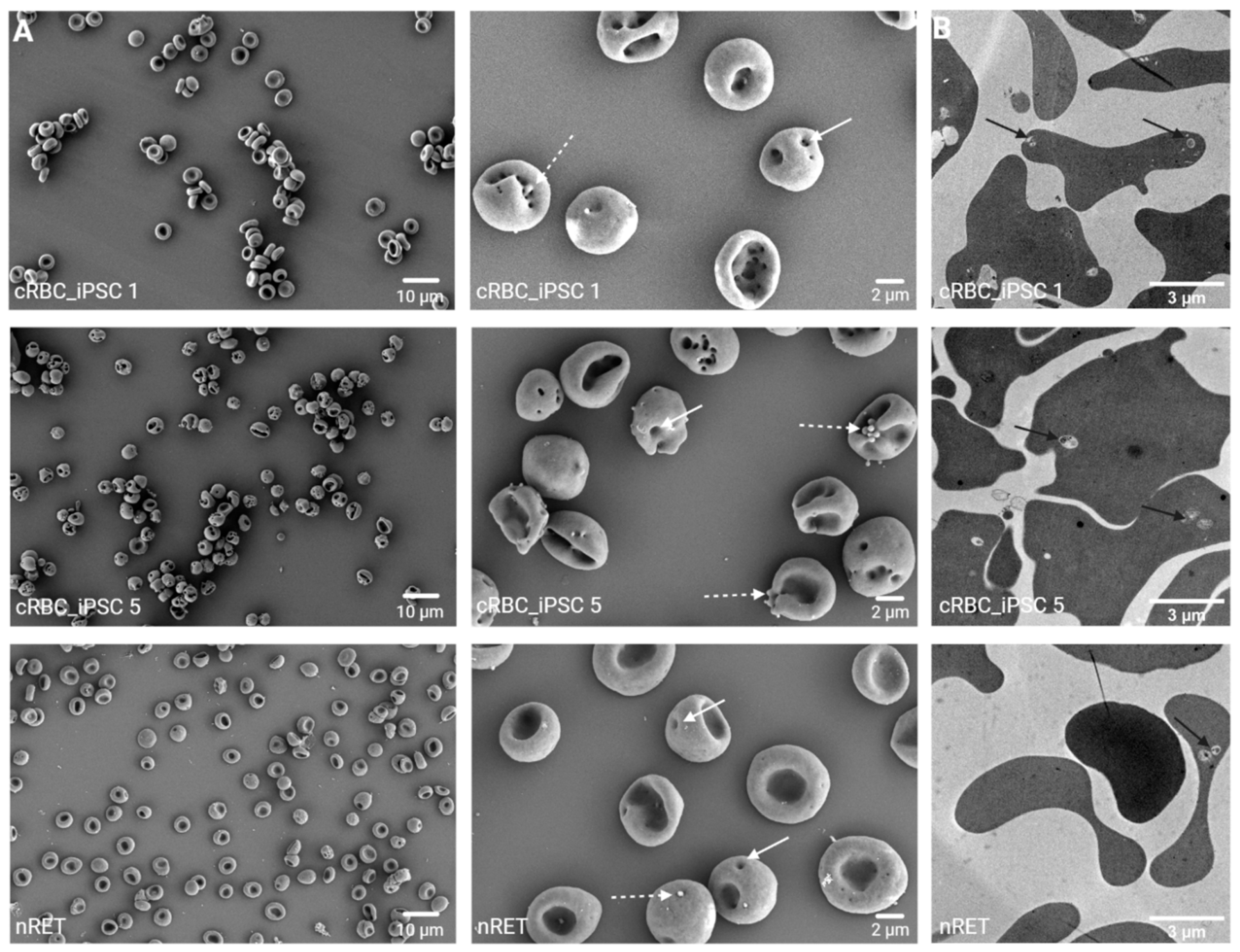
3.6. Morphology of cRBC_iPSCs
3.7. Blood Group Antigen Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, A.; Auvinen, M.K.; Rautonen, J. The aging population poses a global challenge for blood services. Transfusion 2010, 50, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Lim, Z.R.; Vassilev, S.; Leong, Y.W.; Hang, J.W.; Renia, L.; Malleret, B.; Oh, S.K. Industrially compatible transfusable ipsc-derived rbcs: Progress, challenges and prospective solutions. Int. J. Mol. Sci. 2021, 22, 9808. [Google Scholar] [CrossRef] [PubMed]
- Wahlster, L.; Daley, G.Q. Progress towards generation of human haematopoietic stem cells. Nat. Cell Biol. 2016, 18, 1111–1117. [Google Scholar] [CrossRef] [PubMed]
- Peyrard, T.; Bardiaux, L.; Krause, C.; Kobari, L.; Lapillonne, H.; Andreu, G.; Douay, L. Banking of pluripotent adult stem cells as an unlimited source for red blood cell production: Potential applications for alloimmunized patients and rare blood challenges. Transfus. Med. Rev. 2011, 25, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; Amabile, G. Induced pluripotent stem cell-derived red blood cells and platelet concentrates: From bench to bedside. Cells 2017, 7, 2. [Google Scholar] [CrossRef]
- Lee, E.; Sivalingam, J.; Lim, Z.R.; Chia, G.; Shi, L.G.; Roberts, M.; Loh, Y.H.; Reuveny, S.; Oh, S.K. Review: In vitro generation of red blood cells for transfusion medicine: Progress, prospects and challenges. Biotechnol. Adv. 2018, 36, 2118–2128. [Google Scholar] [CrossRef]
- Sivalingam, J.; Su, E.Y.; Lim, Z.R.; Lam, A.T.L.; Lee, A.P.; Lim, H.L.; Chen, H.Y.; Tan, H.K.; Warrier, T.; Hang, J.W.; et al. A scalable suspension platform for generating high-density cultures of universal red blood cells from human induced pluripotent stem cells. Stem Cell Rep. 2021, 16, 182–197. [Google Scholar] [CrossRef]
- Yu, S.; Vassilev, S.; Lim, Z.R.; Sivalingam, J.; Lam, A.T.L.; Ho, V.; Renia, L.; Malleret, B.; Reuveny, S.; Oh, S.K.W. Selection of o-negative induced pluripotent stem cell clones for high-density red blood cell production in a scalable perfusion bioreactor system. Cell Proliferation 2022, e13218. [Google Scholar] [CrossRef]
- Bernecker, C.; Ackermann, M.; Lachmann, N.; Rohrhofer, L.; Zaehres, H.; Arauzo-Bravo, M.J.; van den Akker, E.; Schlenke, P.; Dorn, I. Enhanced ex vivo generation of erythroid cells from human induced pluripotent stem cells in a simplified cell culture system with low cytokine support. Stem Cells Dev. 2019, 28, 1540–1551. [Google Scholar] [CrossRef]
- Bernecker, C.; Kofeler, H.; Pabst, G.; Trotzmuller, M.; Kolb, D.; Strohmayer, K.; Trajanoski, S.; Holzapfel, G.A.; Schlenke, P.; Dorn, I. Cholesterol deficiency causes impaired osmotic stability of cultured red blood cells. Front. Physiol. 2019, 10, 1529. [Google Scholar] [CrossRef]
- Chen, L.; Wang, J.; Liu, J.; Wang, H.; Hillyer, C.D.; Blanc, L.; An, X.; Mohandas, N. Dynamic changes in murine erythropoiesis from birth to adulthood: Implications for the study of murine models of anemia. Blood Adv. 2021, 5, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Palis, J. Primitive and definitive erythropoiesis in mammals. Front. Physiol. 2014, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Dorn, I.; Klich, K.; Arauzo-Bravo, M.J.; Radstaak, M.; Santourlidis, S.; Ghanjati, F.; Radke, T.F.; Psathaki, O.E.; Hargus, G.; Kramer, J.; et al. Erythroid differentiation of human induced pluripotent stem cells is independent of donor cell type of origin. Haematologica 2015, 100, 32–41. [Google Scholar] [CrossRef]
- Jamil, M.A.; Singer, H.; Al-Rifai, R.; Nüsgen, N.; Rath, M.; Strauss, S.; Andreou, I.; Oldenburg, J.; El-Maarri, O. Molecular analysis of fetal and adult primary human liver sinusoidal endothelial cells: A comparison to other endothelial cells. Int. J. Mol. Sci. 2020, 21, 7776. [Google Scholar] [CrossRef]
- Varga, E.; Hansen, M.; Wust, T.; von Lindern, M.; van den Akker, E. Generation of human erythroblast-derived ipsc line using episomal reprogramming system. Stem Cell Res. 2017, 25, 30–33. [Google Scholar] [CrossRef] [PubMed]
- Harboe, M. A method for determination of hemoglobin in plasma by near-ultraviolet spectrophotometry. Scand. J. Clin. Lab. Investig. 1959, 11, 66–70. [Google Scholar] [CrossRef]
- Baskurt, O.K.; Hardeman, M.R.; Uyuklu, M.; Ulker, P.; Cengiz, M.; Nemeth, N.; Shin, S.; Alexy, T.; Meiselman, H.J. Parameterization of red blood cell elongation index--shear stress curves obtained by ektacytometry. Scand. J. Clin. Lab. Investig. 2009, 69, 777–788. [Google Scholar] [CrossRef]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef]
- Triebl, A.; Trotzmuller, M.; Hartler, J.; Stojakovic, T.; Kofeler, H.C. Lipidomics by ultrahigh performance liquid chromatography-high resolution mass spectrometry and its application to complex biological samples. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1053, 72–80. [Google Scholar] [CrossRef]
- Hartler, J.; Triebl, A.; Ziegl, A.; Trotzmuller, M.; Rechberger, G.N.; Zeleznik, O.A.; Zierler, K.A.; Torta, F.; Cazenave-Gassiot, A.; Wenk, M.R.; et al. Deciphering lipid structures based on platform-independent decision rules. Nat. Methods 2017, 14, 1171–1174. [Google Scholar] [CrossRef]
- Hartler, J.; Trotzmuller, M.; Chitraju, C.; Spener, F.; Kofeler, H.C.; Thallinger, G.G. Lipid data analyzer: Unattended identification and quantitation of lipids in lc-ms data. Bioinformatics 2011, 27, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Kundu, M.; Lindsten, T.; Yang, C.Y.; Wu, J.; Zhao, F.; Zhang, J.; Selak, M.A.; Ney, P.A.; Thompson, C.B. Ulk1 plays a critical role in the autophagic clearance of mitochondria and ribosomes during reticulocyte maturation. Blood 2008, 112, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Ney, P.A. Normal and disordered reticulocyte maturation. Curr. Opin. Hematol. 2011, 18, 152–157. [Google Scholar] [CrossRef]
- Kono, M.; Kondo, T.; Takagi, Y.; Wada, A.; Fujimoto, K. Morphological definition of cd71 positive reticulocytes by various staining techniques and electron microscopy compared to reticulocytes detected by an automated hematology analyzer. Clin. Chim. Acta Int. J. Clin. Chem. 2009, 404, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Malleret, B.; Xu, F.; Mohandas, N.; Suwanarusk, R.; Chu, C.; Leite, J.A.; Low, K.; Turner, C.; Sriprawat, K.; Zhang, R.; et al. Significant biochemical, biophysical and metabolic diversity in circulating human cord blood reticulocytes. PLoS ONE 2013, 8, e76062. [Google Scholar] [CrossRef] [PubMed]
- Mohandas, N.; Gallagher, P.G. Red cell membrane: Past, present, and future. Blood 2008, 112, 3939–3948. [Google Scholar] [CrossRef]
- Minetti, G.; Achilli, C.; Perotti, C.; Ciana, A. Continuous change in membrane and membrane-skeleton organization during development from proerythroblast to senescent red blood cell. Front. Physiol. 2018, 9, 286. [Google Scholar] [CrossRef]
- Ovchynnikova, E.; Aglialoro, F.; von Lindern, M.; van den Akker, E. The shape shifting story of reticulocyte maturation. Front. Physiol. 2018, 9, 829. [Google Scholar] [CrossRef]
- Waugh, R.E.; McKenney, J.B.; Bauserman, R.G.; Brooks, D.M.; Valeri, C.R.; Snyder, L.M. Surface area and volume changes during maturation of reticulocytes in the circulation of the baboon. J. Lab. Clin. Med. 1997, 129, 527–535. [Google Scholar] [CrossRef]
- Mortensen, M.; Ferguson, D.J.; Edelmann, M.; Kessler, B.; Morten, K.J.; Komatsu, M.; Simon, A.K. Loss of autophagy in erythroid cells leads to defective removal of mitochondria and severe anemia in vivo. Proc. Natl. Acad. Sci. USA 2010, 107, 832–837. [Google Scholar] [CrossRef]
- Griffiths, R.E.; Kupzig, S.; Cogan, N.; Mankelow, T.J.; Betin, V.M.; Trakarnsanga, K.; Massey, E.J.; Lane, J.D.; Parsons, S.F.; Anstee, D.J. Maturing reticulocytes internalize plasma membrane in glycophorin a-containing vesicles that fuse with autophagosomes before exocytosis. Blood 2012, 119, 6296–6306. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, P.D.; Greenfest-Allen, E.; Frame, J.M.; Bushnell, T.P.; Malik, J.; McGrath, K.E.; Stoeckert, C.J.; Palis, J. Ontogeny of erythroid gene expression. Blood 2013, 121, e5–e13. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Hale, J.; Jaffray, J.; Li, J.; Wang, Y.; Huang, Y.; An, X.; Hillyer, C.; Wang, N.; Kinet, S.; et al. Developmental differences between neonatal and adult human erythropoiesis. Am. J. Hematol. 2018, 93, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, L.; Mohandas, N.; Sorette, M.; Grange, M.J.; Tchernia, G.; Cynober, T. Temporal differences in membrane loss lead to distinct reticulocyte features in hereditary spherocytosis and in immune hemolytic anemia. Blood 2001, 98, 2894–2899. [Google Scholar] [CrossRef]
- Da Costa, L.; Suner, L.; Galimand, J.; Bonnel, A.; Pascreau, T.; Couque, N.; Fenneteau, O.; Mohandas, N.; The Société Française d’Hématologie (SFH). Diagnostic tool for red blood cell membrane disorders: Assessment of a new generation ektacytometer. Blood Cells Mol. Dis. 2016, 56, 9–22. [Google Scholar] [CrossRef]
- Fischbach, F.; Frances, T.; Dunning, M.B. A Manual of Laboratory and Diagnostic Tests, 8th ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2009; p. 1317. [Google Scholar]
- Chasis, J.A.; Prenant, M.; Leung, A.; Mohandas, N. Membrane assembly and remodeling during reticulocyte maturation. Blood 1989, 74, 1112–1120. [Google Scholar] [CrossRef]
- Kobari, L.; Yates, F.; Oudrhiri, N.; Francina, A.; Kiger, L.; Mazurier, C.; Rouzbeh, S.; El-Nemer, W.; Hebert, N.; Giarratana, M.C.; et al. Human induced pluripotent stem cells can reach complete terminal maturation: In vivo and in vitro evidence in the erythropoietic differentiation model. Haematologica 2012, 97, 1795–1803. [Google Scholar] [CrossRef]
- Renoux, C.; Faivre, M.; Bessaa, A.; Da Costa, L.; Joly, P.; Gauthier, A.; Connes, P. Impact of surface-area-to-volume ratio, internal viscosity and membrane viscoelasticity on red blood cell deformability measured in isotonic condition. Sci. Rep. 2019, 9, 6771. [Google Scholar] [CrossRef]
- Giarratana, M.C.; Rouard, H.; Dumont, A.; Kiger, L.; Safeukui, I.; Le Pennec, P.Y.; Francois, S.; Trugnan, G.; Peyrard, T.; Marie, T.; et al. Proof of principle for transfusion of in vitro-generated red blood cells. Blood 2011, 118, 5071–5079. [Google Scholar] [CrossRef]
- Timmins, N.E.; Athanasas, S.; Gunther, M.; Buntine, P.; Nielsen, L.K. Ultra-high-yield manufacture of red blood cells from hematopoietic stem cells. Tissue Eng. Part C Methods 2011, 17, 1131–1137. [Google Scholar] [CrossRef]
- Xiang, J.; Wu, D.C.; Chen, Y.; Paulson, R.F. In vitro culture of stress erythroid progenitors identifies distinct progenitor populations and analogous human progenitors. Blood 2015, 125, 1803–1812. [Google Scholar] [CrossRef] [PubMed]
- Heshusius, S.; Heideveld, E.; Burger, P.; Thiel-Valkhof, M.; Sellink, E.; Varga, E.; Ovchynnikova, E.; Visser, A.; Martens, J.H.A.; von Lindern, M.; et al. Large-scale in vitro production of red blood cells from human peripheral blood mononuclear cells. Blood Adv. 2019, 3, 3337–3350. [Google Scholar] [CrossRef]
- Huang, Y.S.; Delgadillo, L.F.; Cyr, K.H.; Kingsley, P.D.; An, X.; McGrath, K.E.; Mohandas, N.; Conboy, J.G.; Waugh, R.E.; Wan, J.; et al. Circulating primitive erythroblasts establish a functional, protein 4.1r-dependent cytoskeletal network prior to enucleating. Sci. Rep. 2017, 7, 5164. [Google Scholar] [CrossRef]
- Fraser, S.T. The modern primitives: Applying new technological approaches to explore the biology of the earliest red blood cells. ISRN Hematol. 2013, 2013, 568928. [Google Scholar] [CrossRef]
- Olivier, E.N.; Zhang, S.; Yan, Z.; Suzuka, S.; Roberts, K.; Wang, K.; Bouhassira, E.E. Psc-red and mnc-red: Albumin-free and low-transferrin robust erythroid differentiation protocols to produce human enucleated red blood cells. Exp. Hematol. 2019, 75, 31–52 e15. [Google Scholar] [CrossRef] [PubMed]
- Waugh, R.E.; Huang, Y.S.; Arif, B.J.; Bauserman, R.; Palis, J. Development of membrane mechanical function during terminal stages of primitive erythropoiesis in mice. Exp. Hematol. 2013, 41, 398–408.e392. [Google Scholar] [CrossRef] [PubMed]
- Ponnusamy, S.; Zhang, H.; Kadam, P.; Lin, Q.; Lim, T.K.; Sandhu, J.S.; Kothandaraman, N.; Mahyuddin, A.P.; Biswas, A.; Venkat, A.; et al. Membrane proteins of human fetal primitive nucleated red blood cells. J. Proteom. 2012, 75, 5762–5773. [Google Scholar] [CrossRef]
- Faivre, M.; Renoux, C.; Bessaa, A.; Da Costa, L.; Joly, P.; Gauthier, A.; Connes, P. Mechanical signature of red blood cells flowing out of a microfluidic constriction is impacted by membrane elasticity, cell surface-to-volume ratio and diseases. Front. Physiol. 2020, 11, 576. [Google Scholar] [CrossRef]
- Bosman, G.J.; Stappers, M.; Novotny, V.M. Changes in band 3 structure as determinants of erythrocyte integrity during storage and survival after transfusion. Blood Transfus. Trasfus. Del. Sangue 2010, 8 (Suppl. S3), s48–s52. [Google Scholar] [CrossRef]
- Satchwell, T.J.; Hawley, B.R.; Bell, A.J.; Ribeiro, M.L.; Toye, A.M. The cytoskeletal binding domain of band 3 is required for multiprotein complex formation and retention during erythropoiesis. Haematologica 2015, 100, 133–142. [Google Scholar] [CrossRef]
- Mei, Y.; Liu, Y.; Ji, P. Understanding terminal erythropoiesis: An update on chromatin condensation, enucleation, and reticulocyte maturation. Blood Rev. 2021, 46, 100740. [Google Scholar] [CrossRef] [PubMed]
- Daniels, G. Duffy blood group system. In Human Blood Groups, 3rd ed.; Blackwell Publishing Ltd.: Oxford, UK, 2013; pp. 306–319. [Google Scholar]
- Hoher, G.; Fiegenbaum, M.; Almeida, S. Molecular basis of the duffy blood group system. Blood Transfus. Trasfus. Del Sangue 2018, 16, 93–100. [Google Scholar] [CrossRef]
- Deng, J.; Lancelot, M.; Jajosky, R.; Deng, Q.; Deeb, K.; Saakadze, N.; Gao, Y.; Jaye, D.; Liu, S.; Stowell, S.R.; et al. Erythropoietic properties of human induced pluripotent stem cells-derived red blood cells in immunodeficient mice. Am. J. Hematol. 2022, 97, 194–202. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | Blood group phenotypes cRBC_iPSCs | |||||
| Samples | ABO | Rhesus | Kell | Kidd | Duffy | MNS |
| cRBC_iPSC1 | O | D, c, E, e, Cw− | K−, k+, Kp(a−b+) | Jk(a+b−) | Fy(a−b−) | M+N+S+s+ |
| cRBC_iPSC3 | O | D, C, c, e, Cw− | K−, k+, Kp(a−b+) | Jk(a+b+) | Fy(a+b−) | M−N+S−s+ |
| cRBC_iPSC5 | O | D, C, c, e, Cw− | K−, k+, Kp(a-b+) | Jk(a+b+) | Fy(a+b−) | M+N−S−s+ |
| B | Blood group genotypes undifferentiated iPSCs | |||||
| Samples | ABO | Rhesus | Kell | Kidd | Duffy | MNS |
| iPSC1 | ABO*O.01.01/ABO*O.01.02 | ccD.Ee | KEL*02, KEL*02.04 | JK*01 | FY*02 | GYPA*01/GYPA*02 GYPB*03/GYPB*04 |
| iPSC3 | ABO*O.01.01/ABO*O.01.01 | CcD.ee | KEL*02, KEL*02.04 | JK*01/JK*02 | FY*01/FY*02 | GYPA*02/GYPA*04 |
| iPSC5 | ABO*O.01.01/ABO*O.01.02 | CcD.ee | KEL*02 KEL*02.04 | JK*01/JK*02 | FY*01/FY*02 | GYPA*01/GYPA*04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernecker, C.; Matzhold, E.M.; Kolb, D.; Avdili, A.; Rohrhofer, L.; Lampl, A.; Trötzmüller, M.; Singer, H.; Oldenburg, J.; Schlenke, P.; et al. Membrane Properties of Human Induced Pluripotent Stem Cell-Derived Cultured Red Blood Cells. Cells 2022, 11, 2473. https://doi.org/10.3390/cells11162473
Bernecker C, Matzhold EM, Kolb D, Avdili A, Rohrhofer L, Lampl A, Trötzmüller M, Singer H, Oldenburg J, Schlenke P, et al. Membrane Properties of Human Induced Pluripotent Stem Cell-Derived Cultured Red Blood Cells. Cells. 2022; 11(16):2473. https://doi.org/10.3390/cells11162473
Chicago/Turabian StyleBernecker, Claudia, Eva Maria Matzhold, Dagmar Kolb, Afrim Avdili, Lisa Rohrhofer, Annika Lampl, Martin Trötzmüller, Heike Singer, Johannes Oldenburg, Peter Schlenke, and et al. 2022. "Membrane Properties of Human Induced Pluripotent Stem Cell-Derived Cultured Red Blood Cells" Cells 11, no. 16: 2473. https://doi.org/10.3390/cells11162473
APA StyleBernecker, C., Matzhold, E. M., Kolb, D., Avdili, A., Rohrhofer, L., Lampl, A., Trötzmüller, M., Singer, H., Oldenburg, J., Schlenke, P., & Dorn, I. (2022). Membrane Properties of Human Induced Pluripotent Stem Cell-Derived Cultured Red Blood Cells. Cells, 11(16), 2473. https://doi.org/10.3390/cells11162473