
Auxin Regulates Apical Stem Cell Regeneration and Tip Growth in the Marine Red Alga Neopyropia yezoensis



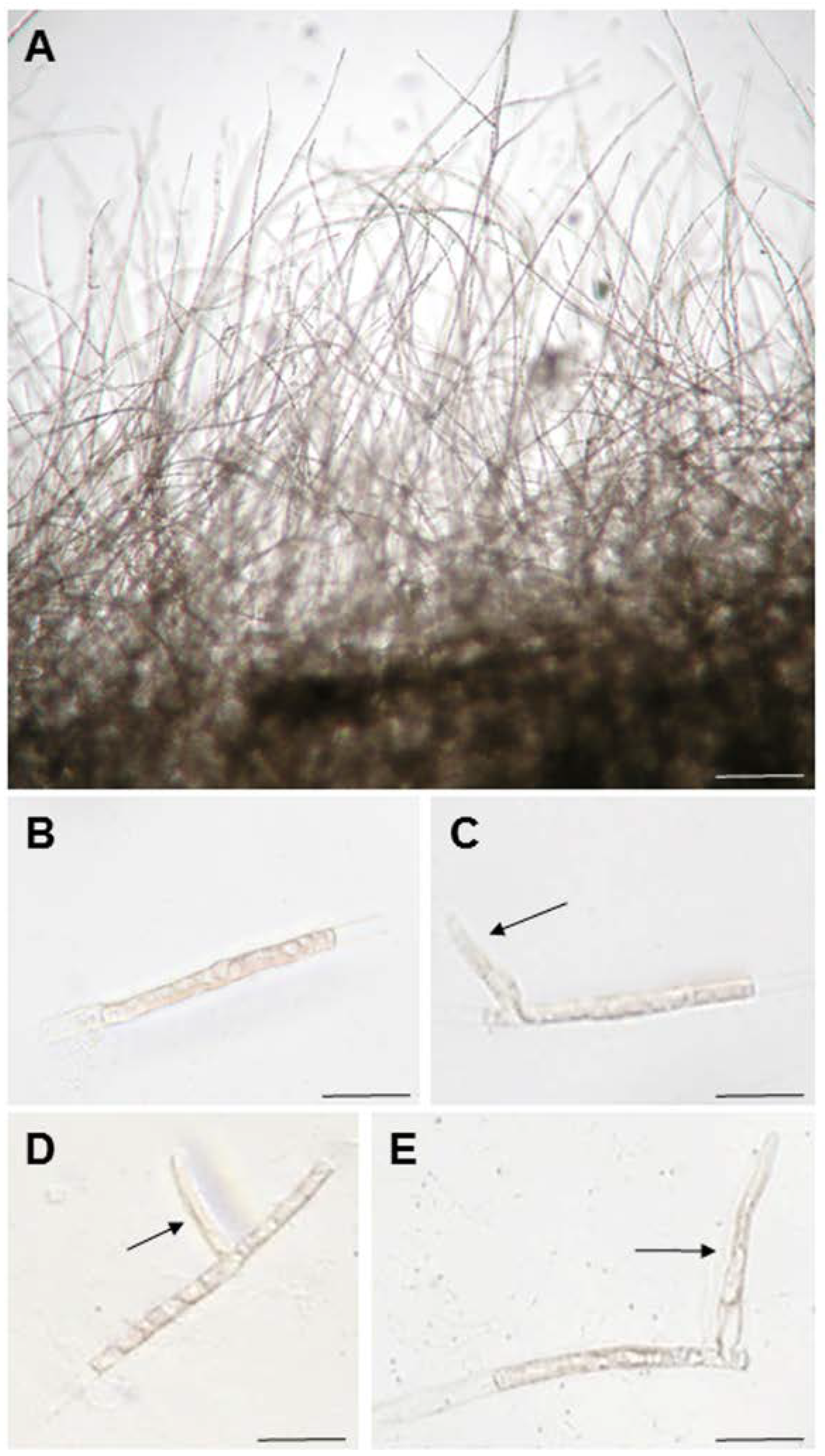{kind=link}
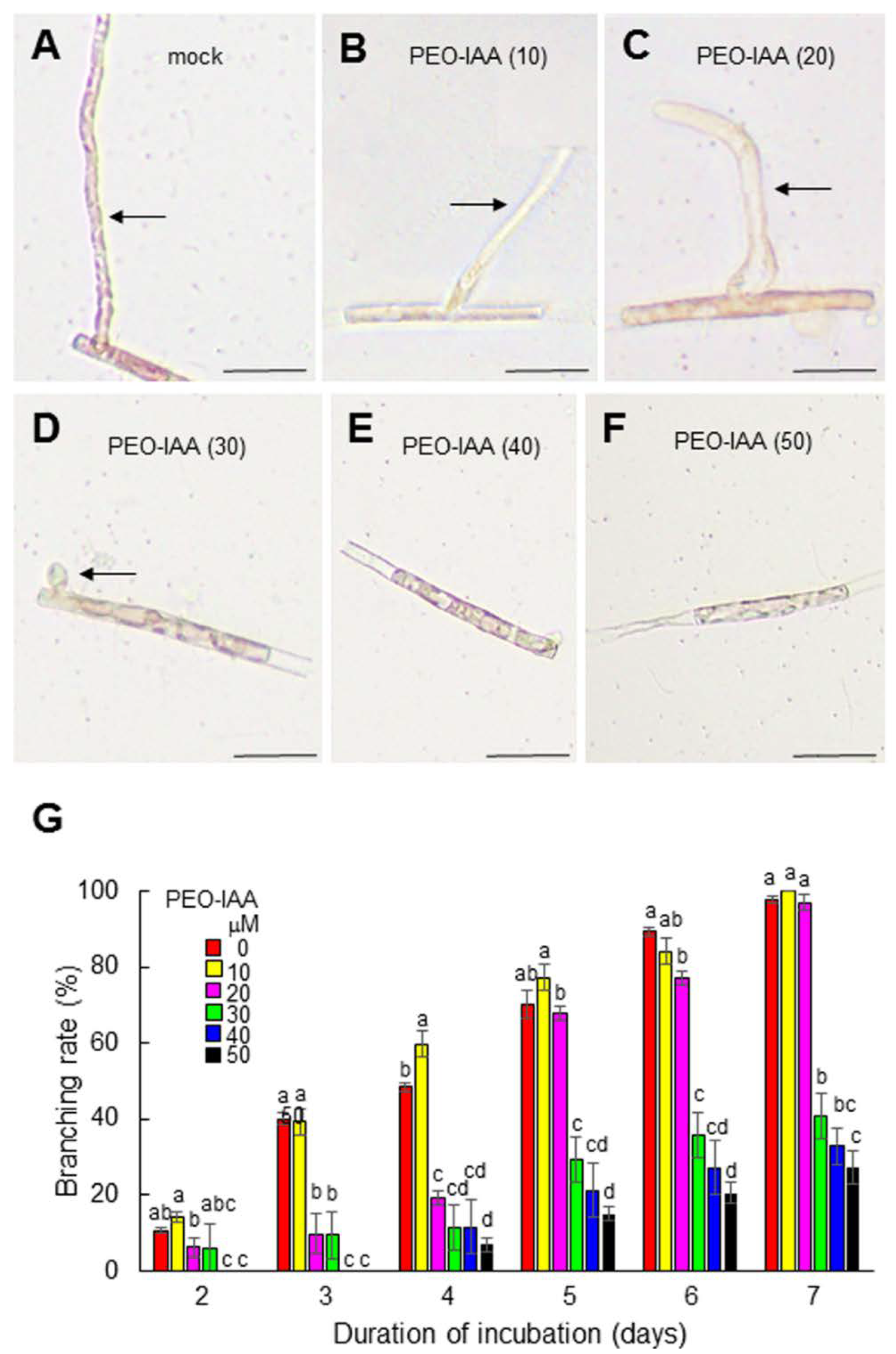{kind=link}
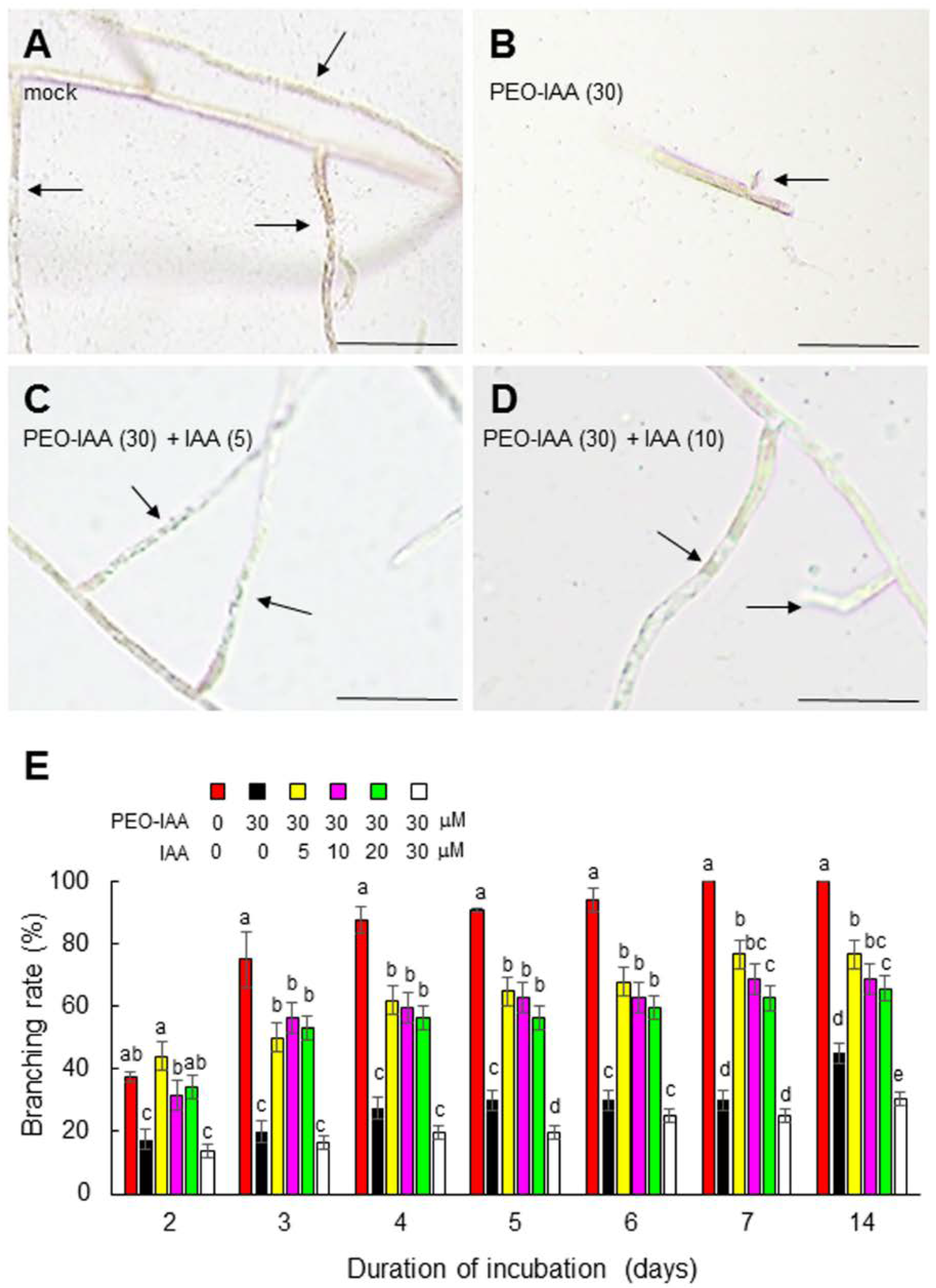{kind=link}
{kind=link}
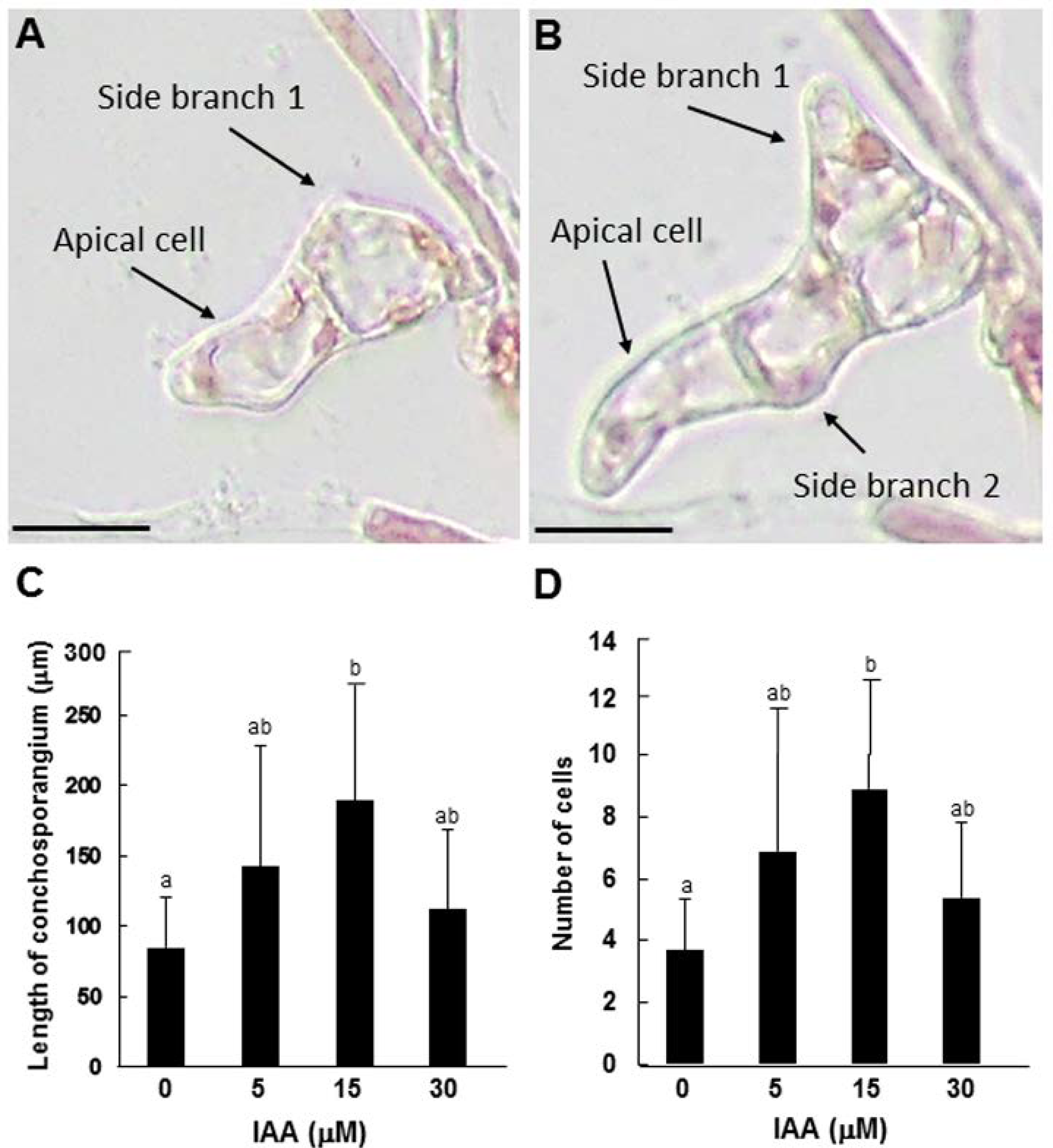{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Materials and Culture Conditions
2.2. Synthesis of Chemical Compounds
2.3. Preparation of Single-Celled Conchocelis and Conchosporangium
2.4. Observation of Naturally Produced Conchosporangia
2.5. Chemical Treatment of Isolated Cells and Naturally Produced Conchosporangia
2.6. Statistical Analysis
3. Results
3.1. Generation of Apical Stem Cells from Single-Celled Conchocelis to Observe Tip Growth
3.2. The Role of Auxin in the Generation and Tip Growth of Side Branches in Conchocelis
3.3. Role of Auxin in Tip Growth and Side Branch Production in Conchosporangia
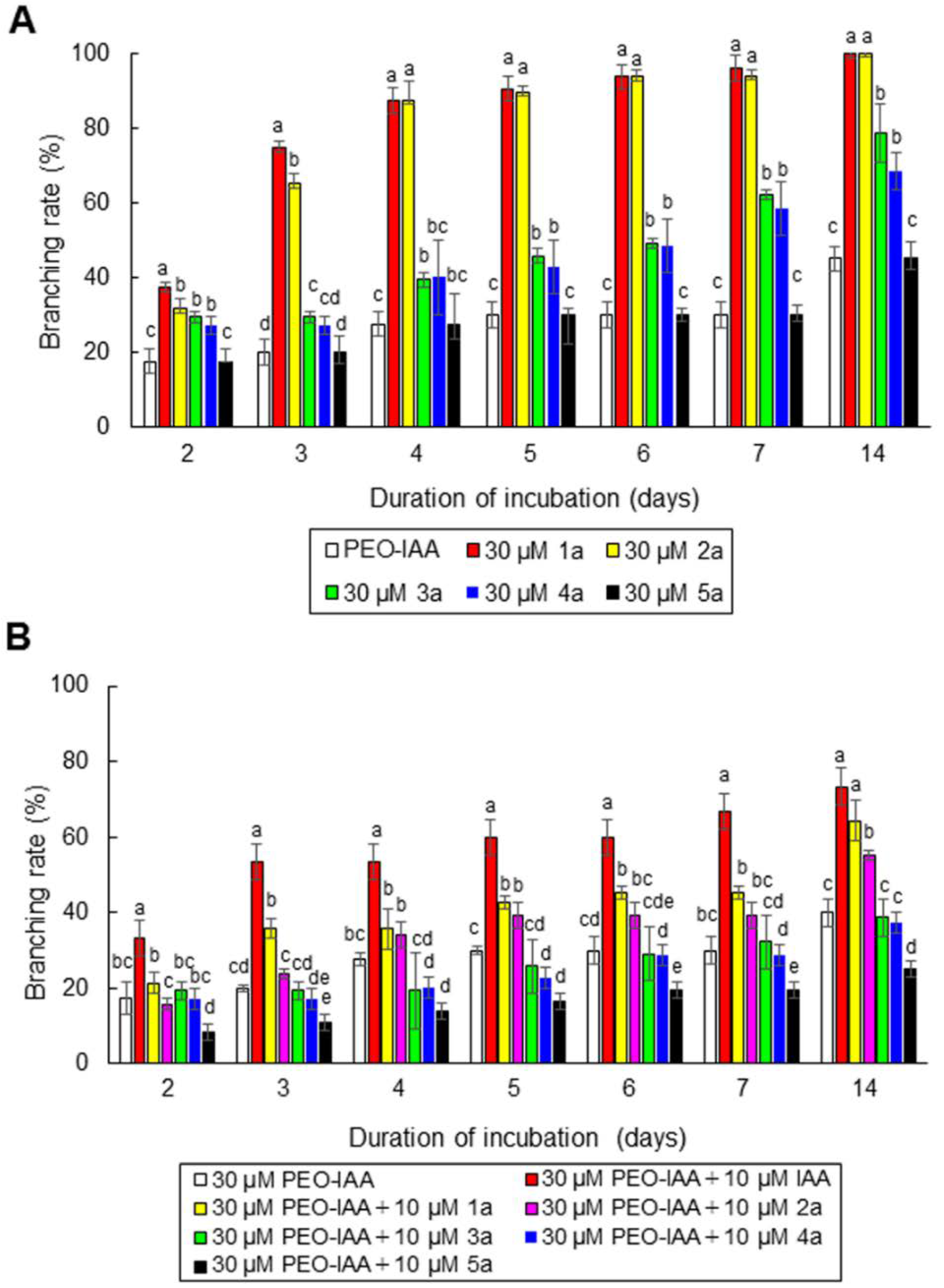
3.4. Characterization of a Unique Auxin Receptor in N. yezoensis Using IAA Derivatives
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heath, I.B. Tip Growth in Plant and Fungal Cells; Academic Press: San Diego, CA, USA, 1990. [Google Scholar] [CrossRef]
- Hepler, P.K.; Vidali, L.; Cheung, A.Y. Polarized cell growth in higher plants. Annu. Rev. Cell Dev. Biol. 2001, 17, 159–187. [Google Scholar] [CrossRef] [PubMed]
- Campàs, O.; Rojas, E.; Dumais, J.; Mahadevan, L. Strategies for cell shape control in tip-growing cells. Am. J. Bot. 2012, 99, 1577–1582. [Google Scholar] [CrossRef] [PubMed]
- Coudert, Y.; Harris, S.; Charrier, B. Design principles of branching morphogenesis in filamentous organisms. Curr. Biol. 2019, 29, R1149. [Google Scholar] [CrossRef] [PubMed]
- Rabille, H.; Billoud, B.; Tesson, B.; Le Panse, S.; Rolland, E.; Charrier, B. The brown algal mode of tip growth: Keeping stress under control. PLoS Biol. 2019, 17, e2005258. [Google Scholar] [CrossRef] [PubMed]
- Orr, R.G.; Cheng, X.; Vidali, L.; Bezanilla, M. Orchestrating cell morphology from the inside out–using polarized cell expansion in plants as a model. Curr. Opin. Cell Biol. 2020, 62, 46–53. [Google Scholar] [CrossRef]
- Steer, M.W.; Steer, J.M. Pollen tube tip growth. New Phytol. 1989, 111, 323–358. [Google Scholar] [CrossRef]
- Yang, Z. Signaling tip growth in plants. Curr. Opin. Plant Biol. 1998, 1, 525–530. [Google Scholar] [CrossRef]
- Carol, R.J.; Dolan, L. Building a hair: Tip growth in Arabidopsis thaliana root hairs. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2002, 357, 815–821. [Google Scholar] [CrossRef]
- Lee, Y.J.; Yang, Z. Tip growth: Signaling in the apical dome. Curr. Opin. Plant Biol. 2008, 11, 662–671. [Google Scholar] [CrossRef] [Green Version]
- Jones, V.A.S.; Dolan, L. The evolution of root hairs and rhizoids. Ann. Bot. 2012, 110, 205–212. [Google Scholar] [CrossRef]
- Kofuji, R.; Hasebe, M. Eight types of stem cells in the life cycle of the moss Physcomitrella patens. Curr. Opin. Plant Biol. 2014, 17, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Suo, J.; Zhao, Q.; Zhang, Z.; Chen, S.; Cao, J.; Liu, G.; Wei, X.; Wang, T.; Yang, C.; Dai, S. Cytological and proteomic analyses of Osmunda cinnamomea germinating spores reveal characteristics of fern spore germination and rhizoid tip growth. Mol. Cell Proteom. 2015, 14, 2510–2534. [Google Scholar] [CrossRef] [PubMed]
- Dehors, J.; Mareck, A.; Kiefer-Meyer, M.C.; Menu-Bouaouiche, L.; Lehner, A.; Mollet, J.C. Evolution of cell wall polymers in tip-growing land plant gametophytes: Composition, distribution, functional aspects and their remodeling. Front. Plant Sci. 2019, 10, 441. [Google Scholar] [CrossRef] [PubMed]
- Geitmann, A.; Emons, A.M.C. The cytoskeleton in plant and fungal cell tip growth. J. Microsc. 2000, 198, 218–245. [Google Scholar] [CrossRef] [PubMed]
- Campanoni, P.; Blatt, M.R. Membrane trafficking and polar growth in root hairs and pollen tubes. J. Exp. Bot. 2007, 58, 65–74. [Google Scholar] [CrossRef]
- Ischebeck, T.; Seiler, S.; Heilmann, I. At the poles across kingdoms: Phosphoinositides and polar tip growth. Protoplasma 2010, 240, 13–31. [Google Scholar] [CrossRef]
- Pei, W.; Du, F.; Zhang, Y.; He, T.; Ren, H. Control of the actin cytoskeleton in root hair development. Plant Sci. 2012, 187, 10–18. [Google Scholar] [CrossRef]
- Malhó, R.; Serrazina, S.; Saavedra, L.; Dias, F.V.; Rehman, R.U. Ion and lipid signaling in apical growth—a dynamic machinery responding to extracellular cues. Front. Plant Sci. 2015, 6, 816. [Google Scholar] [CrossRef]
- Mangano, S.; Juárez, S.P.D.; Estevez, J.M. ROS regulation of polar growth in plant cells. Plant Physiol. 2016, 171, 1593–1605. [Google Scholar] [CrossRef] [Green Version]
- Mangano, S.; Denita-Juarez, S.P.; Choi, H.S.; Marzol, E.; Hwang, Y.; Ranocha, P.; Velasquez, S.M.; Borassi, C.; Barberini, M.L.; Aptekmann, A.A.; et al. Molecular link between auxin and ROS-mediated polar growth. Proc. Natl. Acad. Sci. USA 2017, 114, 5289–5294. [Google Scholar] [CrossRef]
- Velasquez, S.M.; Barbez, E.; Kleine-Vehn, J.; Estevez, J.M. Auxin and cellular elongation. Plant Physiol. 2016, 170, 1206–1215. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Konno, H.; Takeda, S.; Dolan, L.; Kato, M.; Aoyama, T.; Higaki, T.; Takigawa-Imamura, H.; Sato, M.H. PtdIns(3,5)P2 mediates root hair shank hardening in Arabidopsis. Nat. Plants 2018, 4, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Sugimoto, K. A gene regulatory network for root hair development. J. Plant Res. 2019, 132, 301–309. [Google Scholar] [CrossRef]
- Hayashi, K.; Arai, K.; Aoi, Y.; Tanaka, Y.; Hira, H.; Guo, R.; Hu, Y.; Ge, C.; Zhao, Y.; Kasahara, H.; et al. The main oxidative inactivation pathway of the plant hormone auxin. Nat. Commun. 2021, 12, 6752. [Google Scholar] [CrossRef] [PubMed]
- Rounds, C.M.; Bezanilla, M. Growth mechanisms in tip-growing plant cells. Annu. Rev. Plant Biol. 2013, 64, 243–265. [Google Scholar] [CrossRef]
- Riquelme, M.; Aguirre, J.; Bartnicki-García, S.; Braus, G.H.; Feldbrügge, M.; Fleig, U.; Hansberg, W.; Herrera-Estrella, A.; Kämper, J.; Kück, U.; et al. Fungal morphogenesis, from the polarized growth of hyphae to complex reproduction and infection structures. Microbiol. Mol. Biol. Rev. 2018, 82, e00068-17. [Google Scholar] [CrossRef]
- Bittisnich, D.J.; Williamson, R.E. Tip-localised H+-fluxes and the applicability of the acid-growth hypothesis to tip-growing cells: Control of chloronemal extension in Funaria hygrometrica by auxin and light. Planta 1989, 178, 96–102. [Google Scholar] [CrossRef]
- Wu, J.Z.; Lin, Y.; Zhang, X.L.; Pang, D.W.; Zhao, J. IAA stimulates pollen tube growth and mediates the modification of its wall composition and structure in Torenia fournieri. J. Exp. Bot. 2008, 59, 2529–2543. [Google Scholar] [CrossRef]
- Wu, J.Z.; Qin, Y.; Zhao, J. Pollen tube growth is affected by exogenous hormones and correlated with hormone changes in styles before and after pollination in Torenia fournieri L. Plant Growth Regul. 2008, 55, 137–148. [Google Scholar] [CrossRef]
- Tao, S.; Estelle, M. Mutational studies of the Aux/IAA proteins in Physcomitrella reveal novel insights into their function. New Phytol. 2018, 218, 1534–1542. [Google Scholar] [CrossRef]
- Thelander, M.; Landberg, K.; Sundberg, E. Auxin-mediated developmental control in the moss Physcomitrella patens. J. Exp. Bot. 2018, 69, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Neve, J.; Hirose, M.; Kuboki, A.; Shimada, Y.; Kepinski, S.; Nozaki, H. Rational design of an auxin antagonist of the SCF (TIR1) auxin receptor complex. ACS Chem. Biol. 2012, 7, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Fukui, K.; Hayashi, K. Manipulation and sensing of auxin metabolism, transport and signaling. Plant Cell Physiol. 2018, 59, 1500–1510. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. Auxin signaling. Plant Physiol. 2018, 176, 465–479. [Google Scholar] [CrossRef]
- Bhosale, R.; Giri, J.; Pandey, B.K.; Giehl, R.F.H.; Hartmann, A.; Traini, R.; Truskina, J.; Leftley, N.; Hanlon, M.; Swarup, K.; et al. A mechanistic framework for auxin dependent Arabidopsis root hair elongation to low external phosphate. Nat. Commun. 2018, 9, 1409. [Google Scholar] [CrossRef]
- Schoenaers, S.; Balcerowicz, D.; Breen, G.; Hill, K.; Zdanio, M.; Mouille, G.; Holman, T.J.; Oh, J.; Wilson, M.H.; Nikonorova, N.; et al. The auxin-regulated CrRLK1L kinase ERULUS controls cell wall composition during root hair tip growth. Curr. Biol. 2018, 28, 722–732.e6. [Google Scholar] [CrossRef]
- Blouin, N.A.; Brodie, J.A.; Grossman, A.C.; Xu, P.; Brawley, S.H. Porphyra: A marine crop shaped by stress. Trends Plant Sci. 2011, 16, 29–37. [Google Scholar] [CrossRef]
- Takahashi, M.; Mikami, K. Oxidative stress promotes asexual reproduction and apogamy in the red seaweed Pyropia yezoensis. Front. Plant Sci. 2017, 8, 62. [Google Scholar] [CrossRef]
- Mikami, K.; Li, C.; Irie, R.; Hama, Y. A unique life cycle transition in the red seaweed Pyropia yezoensis depends on apospory. Commun. Biol. 2019, 2, 299. [Google Scholar] [CrossRef] [Green Version]
- Hiwatashi, Y.; Shimada, M.; Mikami, K.; Takada, N. Establishment of a live-imaging analysis for polarized growth of conchocelis in the multicellular red alga Neopyropia yezoensis. Front. Plant Sci. 2022, 12, 716011. [Google Scholar] [CrossRef]
- Mikami, K.; Takahashi, M. Life cycle and reproduction dynamics of Bangiales in response to environmental stresses. Semin. Cell Dev. Biol. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Tan, X.; Zheng, N.; Hatate, T.; Kimura, Y.; Kepinski, S.; Nozaki, H. Small-molecule agonists and antagonists of F-box protein-substrate interactions in auxin perception and signaling. Proc. Natl. Acad. Sci. USA 2008, 105, 5632–5637. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K. Chemical biology in auxin research. Cold Spring Harb. Perspect. Biol. 2021, 13, a040105. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, E.; Yang, H.; Nishimura, T.; Uehara, Y.; Sakai, T.; Furutani, M.; Koshiba, T.; Hirose, M.; Nozaki, H.; Murphy, A.S.; et al. Alkoxy-auxins are selective inhibitors of auxin transport mediated by PIN, ABCB, and AUX1 transporters. J. Biol. Chem. 2011, 286, 2354–2364. [Google Scholar] [CrossRef]
- Li, C.; Ariga, I.; Mikami, K. Difference in nitrogen starvation-inducible expression patterns among phylogenetically diverse ammonium transporter genes in the red seaweed Pyropia yezoensis. Am. J. Plant Sci. 2019, 10, 1325–1349. [Google Scholar] [CrossRef]
- Takahashi, M.; Mikami, K. Phototropism in the marine red macroalga Pyropia yezoensis. Am. J. Plant Sci. 2016, 7, 2412–2428. [Google Scholar] [CrossRef]
- Cleland, R.; Bennet-Clark, T.A. The effect of high concentrations of auxin on the extension of Avena coleoptile sections. J. Exp. Bot. 1961, 12, 65–74. [Google Scholar] [CrossRef]
- Chadwick, A.V.; Burg, S.P. An explanation of the inhibition of root growth caused by indole-3-acetic Acid. Plant Physiol. 1967, 42, 415–420. [Google Scholar] [CrossRef]
- Sauer, M.; Robert, S.; Kleine-Vehn, J. Auxin: Simply complicated. J. Exp. Bot. 2013, 64, 2565–2577. [Google Scholar] [CrossRef] [Green Version]
- Küplemez, H.; Yildirım, M.U. Effects of cytokinin and auxin on plant development and vascular tissues in Lens culinaris. Comm. J. Biol. 2020, 4, 16–21. [Google Scholar] [CrossRef]
- Adams, E.; Mikami, K.; Shin, R. Selection and functional analysis of a Pyropia yezoensis ammonium transporter PyAMT1 in potassium deficiency. J. Appl. Phycol. 2017, 29, 2617–2626. [Google Scholar] [CrossRef]
- Mikami, K.; Mori, I.C.; Matsuura, T.; Ikeda, Y.; Kojima, M.; Sakakibara, H.; Hirayama, T. Comprehensive quantification and genome survey reveal the presence of novel phytohormone action modes in red seaweeds. J. Appl. Phycol. 2016, 28, 2539–2548. [Google Scholar] [CrossRef]
- Mori, I.C.; Ikeda, Y.; Matsuura, T.; Hirayama, T.; Mikami, K. Phytohormones in red seaweeds: A technical review of methods for analysis and a consideration of genomic data. Bot. Mar. 2017, 60, 153–170. [Google Scholar] [CrossRef]
- Uenaka, H.; Wada, M.; Kadota, A. Four distinct photoreceptors contribute to light-induced side branch formation in the moss Physcomitrella Patens. Planta 2005, 222, 623–631. [Google Scholar] [CrossRef]
- Menand, B.; Calder, G.; Dolan, L. Both chloronemal and caulonemal cells expand by tip growth in the moss Physcomitrella patens. J. Exp. Bot. 2007, 58, 1843–1849. [Google Scholar] [CrossRef]
- Gray, W.M.; Kepinski, S.; Rouse, D.; Leyser, O.; Estelle, M. Auxin regulates SCFTIR1-dependent degradation of AUX/IAA proteins. Nature 2001, 414, 271–276. [Google Scholar] [CrossRef]
- Parry, G.; Calderon-Villalobos, L.I.; Prigge, M.; Peret, B.; Dharmasiri, S.; Itoh, H.; Lechner, E.; Gray, M.; Bennett, M.; Estelle, M. Complex regulation of the TIR1/AFB family of auxin receptors. Proc. Natl. Acad. Sci. USA 2009, 106, 22540–22545. [Google Scholar] [CrossRef]
- Salehin, M.; Bagchi, R.; Estelle, M. SCFTIR1/AFB-based auxin perception: Mechanism and role in plant growth and development. Plant Cell 2015, 27, 9–19. [Google Scholar] [CrossRef]
- Le Bail, A.; Billoud, B.; Kowalczyk, N.; Kowalczyk, M.; Gicquel, M.; Le Panse, S.; Stewart, S.; Scornet, D.; Cock, J.M.; Ljung, K.; et al. Auxin metabolism and function in the multicellular brown alga Ectocarpus siliculosus. Plant Physiol. 2010, 153, 128–144. [Google Scholar] [CrossRef] [Green Version]
- Namba, A.; Shigenobu, Y.; Kobayashi, M.; Kobayashi, T.; Oohara, I. A new primer for 16S rDNA analysis of microbial communities associated with Porphyra yezoensis. Fish. Sci. 2010, 76, 873–878. [Google Scholar] [CrossRef]
- Fukui, Y.; Abe, M.; Kobayashi, M.; Ishihara, K.; Oikawa, H.; Yano, Y.; Satomi, M. Maritalea porphyrae sp. nov., isolated from a red alga (Porphyra yezoensis), and transfer of Zhangella mobilis to Maritalea mobilis comb. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Fukui, Y.; Abe, M.; Kobayashi, M.; Yano, Y.; Satomi, M. Isolation of Hyphomonas strains that induce normal morphogenesis in protoplasts of the marine red alga Pyropia yezoensis. Microb. Ecol. 2014, 68, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Handayani, M.; Sasaki, H.; Matsuda, R.; Takechi, K.; Takano, H.; Takio, S. Characterization of an epiphytic bacterium Neptunomonas sp. BPy-1 on the gametophytes of a red alga Pyropia yezoensis. Am. J. Plant Sci. 2014, 5, 3652–3661. [Google Scholar] [CrossRef]
- Yan, Y.W.; Yang, H.C.; Tang, L.; Li, J.; Mao, Y.X.; Mo, Z.L. Compositional shifts of bacterial communities associated with Pyropia yezoensis and surrounding seawater co-occurring with red rot disease. Front. Microbiol. 2019, 10, 1666. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Khurshid, A.; Tang, X.; Wang, J.; Khan, T.U.; Mao, Y. Structural and functional impacts of microbiota on Pyropia yezoensis and surrounding seawater in cultivation farms along coastal areas of the Yellow Sea. Microorganisms 2021, 9, 1291. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Bijo, A.J.; Baghel, R.S.; Reddy, C.R.; Jha, B. Role of bacterial isolates in enhancing the bud induction in the industrially important red alga Gracilaria dura. FEMS Microbiol. Ecol. 2011, 76, 381–392. [Google Scholar] [CrossRef]
- Singh, R.P.; Reddy, C.R. Seaweed-microbial interactions: Key functions of seaweed-associated bacteria. FEMS Microbiol. Ecol. 2014, 88, 213–230. [Google Scholar] [CrossRef]
- Xiong, Y.; Yang, R.; Sun, X.; Yang, H.; Chen, H. Effect of the epiphytic bacterium Bacillus sp. WPySW2 on the metabolism of Pyropia haitanensis. J. Appl. Phycol. 2018, 30, 1225–1237. [Google Scholar] [CrossRef]
- Wang, J.; Tang, X.; Mo, Z.; Mao, Y. Metagenome-assembled genomes from Pyropia haitanensis microbiome provide insights into the potential metabolic functions to the seaweed. Front. Microbiol. 2022, 13, 857901. [Google Scholar] [CrossRef]
- Matsuda, R.; Handayani, M.L.; Sasaki, H.; Takechi, K.; Takano, H.; Takio, S. Production of indole acetic acid by strains of the epiphytic bacteria Neptunomonas spp. isolated from the red alga Pyropia yezoensis and the seagrass Zostera marina. Arch. Microbiol. 2018, 200, 255–265. [Google Scholar] [CrossRef]
- Tapia, J.E.; González, B.; Goulitquer, S.; Potin, P.; Correa, J.A. Microbiota influences morphology and reproduction of the brown alga Ectocarpus sp. Front. Microbiol. 2016, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- KleinJan, H.; Jeanthon, C.; Boyen, C.; Dittami, S.M. Exploring the cultivable Ectocarpus microbiome. Front. Microbiol. 2017, 8, 2456. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taya, K.; Takeuchi, S.; Takahashi, M.; Hayashi, K.-i.; Mikami, K. Auxin Regulates Apical Stem Cell Regeneration and Tip Growth in the Marine Red Alga Neopyropia yezoensis. Cells 2022, 11, 2652. https://doi.org/10.3390/cells11172652
Taya K, Takeuchi S, Takahashi M, Hayashi K-i, Mikami K. Auxin Regulates Apical Stem Cell Regeneration and Tip Growth in the Marine Red Alga Neopyropia yezoensis. Cells. 2022; 11(17):2652. https://doi.org/10.3390/cells11172652
Chicago/Turabian StyleTaya, Kensuke, Shunzei Takeuchi, Megumu Takahashi, Ken-ichiro Hayashi, and Koji Mikami. 2022. "Auxin Regulates Apical Stem Cell Regeneration and Tip Growth in the Marine Red Alga Neopyropia yezoensis" Cells 11, no. 17: 2652. https://doi.org/10.3390/cells11172652
APA StyleTaya, K., Takeuchi, S., Takahashi, M., Hayashi, K. -i., & Mikami, K. (2022). Auxin Regulates Apical Stem Cell Regeneration and Tip Growth in the Marine Red Alga Neopyropia yezoensis. Cells, 11(17), 2652. https://doi.org/10.3390/cells11172652