
Golgi Metal Ion Homeostasis in Human Health and Diseases



Abstract
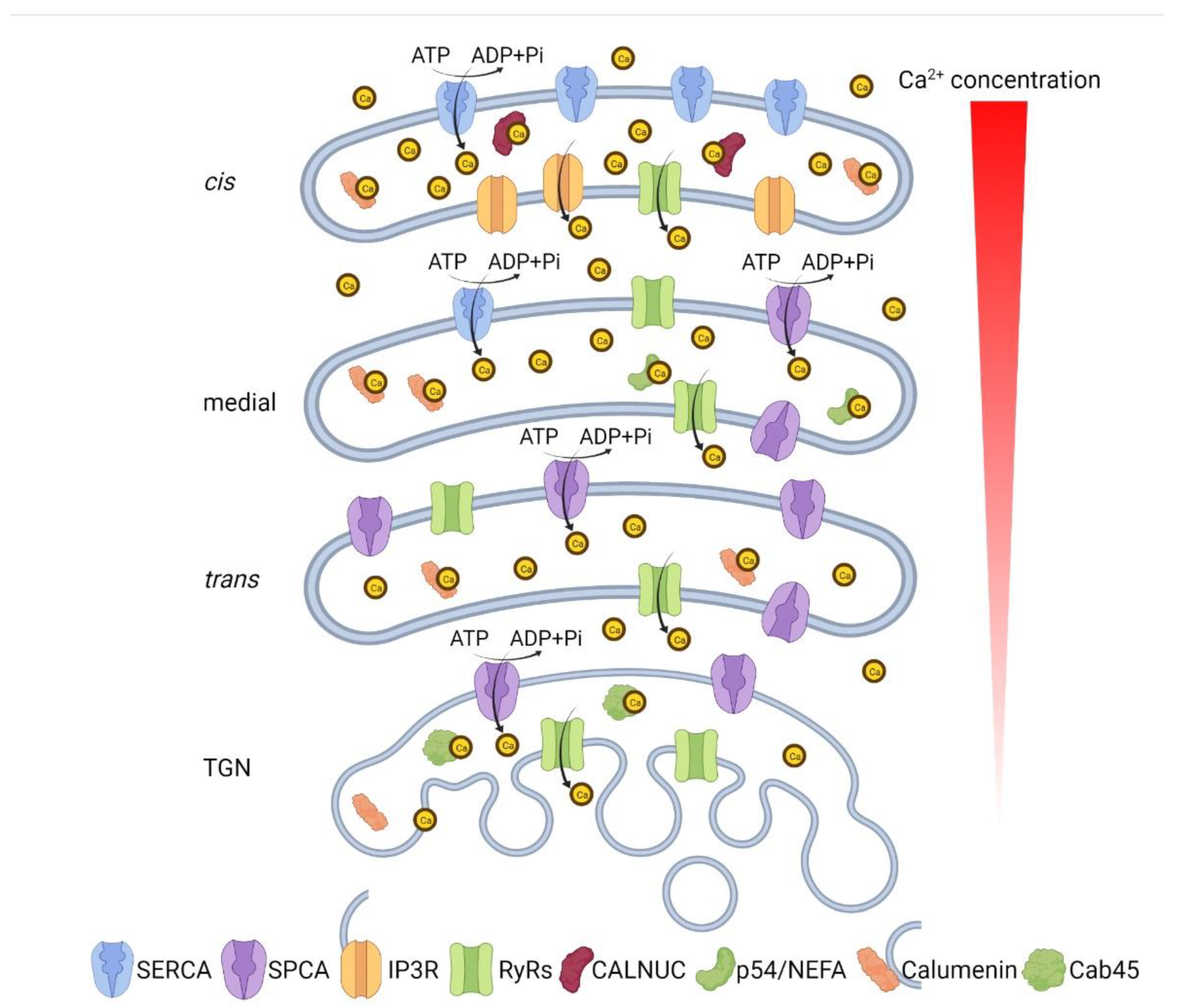
:1. Introduction

{kind=link}
| Metal Ion | Disease | OMIM | Gene | Protein | Clinical Features |
|---|---|---|---|---|---|
| Ca2+/Mn2+ | Brody myopathy (BRM) [3] | 601003 | ATP2A1 | SERCA1 | Early onset of muscle function disorder characterized by muscle cramping and post-exercise stiffening (myopathy). |
| Acrokeratosis verruciformis (AKV) [4] | 101900 | ATP2A2 | SERCA2 | Early onset keratinization disorder affecting the distal extremities. | |
| Darier disease (DD) [5] | 124200 | ATP2A2 | SERCA2 | Early onset keratinizating disorder characterized by small papules predominantly in seborrheic areas. | |
| Hailey-Hailey disease (HHD) [6] | 169600 | ATP2C1 | SPCA1 | A skin disease causing persistent blisters and suprabasal cell separation (acantholysis) of the epidermis. | |
| Spinocerebellar ataxia 15 (SCA15) [7] | 606658 | ITPR1 | IP3R 1 | A neurological condition characterized by progressive gait and limb ataxia. | |
| Spinocerebellar ataxia 29 (SCA29) [8] | 117360 | ITPR1 | IP3R 1 | Early onset cerebellar ataxia causing slowly progressive or non-progressive gait and limb ataxia. | |
| Gillespie syndrome (GLSP) [9] | 206700 | ITPR1 | IP3R 1 | A congenital neurological disorder characterized by the association of partial bilateral aniridia with non-progressive cerebellar ataxia, and intellectual disability. | |
| Anhidrosis, isolated, with normal sweat glands (ANHD) [10] | 106190 | ITPR2 | IP3R 2 | A disorder characterized by absence of perspiration and subsequent heat intolerance with normal morphology and number of sweat glands. | |
| Malignant hyperthermia 1 (MHS1) [11] | 145600 | RYR1 | RyR1 | A skeletal muscle disorder and the main causes of death due to anesthesia characterized by any combination of hyperthermia, skeletal muscle rigidity, tachycardia or arrhythmia, respiratory and metabolic acidosis, and rhabdomyolysis. | |
| Central core disease of muscle (CCD) [12,13] | 117000 | RYR1 | RyR1 | A mild congenital myopathy characterized by motor developmental delay and signs of mild proximal weakness. | |
| Multiminicore disease with external ophthalmoplegia (MMDO) [14] | 255320 | RYR1 | RyR1 | A heterogeneous neuromuscular disorder characterized by neonatal hypotonia, delayed motor development, and generalized muscle weakness and amyotrophy. | |
| Arrhythmogenic right ventricular dysplasia, familial, 2 (ARVD2) [15] | 600996 | RYR2 | RyR2 | A congenital heart disease characterized by effort-induced polymorphic ventricular tachycardias due to large areas of fatty-fibrous replacement in the subepicardial layer of the right ventricle. | |
| Ventricular tachycardia, catecholaminergic polymorphic, 1 (CPVT1) [16] | 604772 | RYR2 | RyR2 | An arrhythmogenic disorder characterized by physical activity- or stress-induced, polymorphic ventricular tachycardia that may degenerate into deteriorate into ventricular fibrillation. | |
| Congenital disorders of glycosylation, Type IIk (CDG2K) [17] | 614727 | TMEM165 | TMEM165 | An autosomal recessive disorder with a variable phenotype, characterized by growth retardation. | |
| Zn2+ | A novel syndrome with early onset agammaglobulinemia and absent B cells of unknown cause [18] | N/A | SLC39A7 | Zip7 | A novel autosomal recessive disease characterized by absent B cells, agammaglobulinemia and early-onset infections. |
| Ehlers–Danlos syndrome, Spndylodysplastic Type, 3 (SCD-EDS) [19] | 612350 | SLC39A13 | Zip13 | Postnatal growth retardation characterized by short stature, hyperelastic skin and hypermobile joints, protuberant eyes with bluish sclerae, atrophy of the thenar muscles, wrinkled palms and tapering fingers. | |
| Cu2+ | Menkes disease (MNK) [20,21,22] | 309400 | ATP7A | ATP7A | A disorder characterized by generalized copper deficiency, early retardation in growth, peculiar hair, and focal cerebral and cerebellar degeneration due to the dysfunction of several copper-dependent enzymes. |
| Occipital horn syndrome (OHS) [23] | 304150 | ATP7A | ATP7A | A rare connective tissue disorder characterized by hyperelastic and bruisable skin, hernias, bladder diverticula, hyperextensible joints, varicosities, and multiple skeletal abnormalities, sometimes accompanied by mild neurologic impairment, and bony abnormalities of the occiput. | |
| Distal spinal muscular atrophy, X-linked, 3 (DSMAX3) [24] | 300489 | ATP7A | ATP7A | Neuromuscular disorders caused by selective degeneration of motor neurons in the anterior horn of the spinal cord. | |
| Wilson disease (WD) [25,26] | 277900 | ATP7B | ATP7B | A disorder characterized by dramatic accumulation of intracellular copper with subsequent hepatic and neurologic abnormalities |
2. Ca2+/Mn2+ Transporters and Channels
2.1. Ca2+/Mn2+ Transporters in the Golgi
2.2. Ca2+-Release Channels in the Golgi
2.3. Ca2+-Binding Proteins in the Golgi Lumen
2.4. Disruption of Ca2+/Mn2+ Homeostasis Impairs Golgi Structure and Function
2.5. Golgi Lumenal Ca2+ Is Essential for Intra-Golgi Trafficking and Protein Sorting at the TGN
2.6. Golgi Ca2+/Mn2+ Homeostasis and Human Diseases
3. Zn2+ Transporters
3.1. Zn2+ Transporters
3.2. Zn2+ Function in the Earlier Secretory Pathway
3.3. Zn2+ Homeostasis and Human Diseases
4. Cu Ion (Cu2+ and Cu+) Transporters
4.1. Cu Ion Transporters in the Golgi
4.2. Cu Transporter Trafficking
4.3. Cu Homeostasis and Human Diseases
5. Summary and Perspective
Funding
Conflicts of Interest
References
- Li, J.; Ahat, E.; Wang, Y. Golgi Structure and Function in Health, Stress, and Diseases. Results Probl. Cell Differ. 2019, 67, 441–485. [Google Scholar]
- Kellokumpu, S. Golgi pH, Ion and Redox Homeostasis: How Much Do They Really Matter? Front. Cell Dev. Biol. 2019, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Odermatt, A.; Barton, K.; Khanna, V.K.; Mathieu, J.; Escolar, D.; Kuntzer, T.; Karpati, G.; MacLennan, D.H. The mutation of Pro789 to Leu reduces the activity of the fast-twitch skeletal muscle sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA1) and is associated with Brody disease. Hum. Genet. 2000, 106, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Dhitavat, J.; Macfarlane, S.; Dode, L.; Leslie, N.; Sakuntabhai, A.; MacSween, R.; Saihan, E.; Hovnanian, A. Acrokeratosis verruciformis of Hopf is caused by mutation in ATP2A2: Evidence that it is allelic to Darier’s disease. J. Investig. Dermatol. 2003, 120, 229–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuntabhai, A.; Ruiz-Perez, V.; Carter, S.; Jacobsen, N.; Burge, S.; Monk, S.; Smith, M.; Munro, C.S.; O’Donovan, M.; Craddock, N.; et al. Mutations in ATP2A2, encoding a Ca2+ pump, cause Darier disease. Nat. Genet. 1999, 21, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Bonifas, J.M.; Beech, J.; Bench, G.; Shigihara, T.; Ogawa, H.; Ikeda, S.; Mauro, T.; Epstein, E.H., Jr. Mutations in ATP2C1, encoding a calcium pump, cause Hailey-Hailey disease. Nat. Genet. 2000, 24, 61–65. [Google Scholar] [CrossRef]
- Van de Leemput, J.; Chandran, J.; Knight, M.A.; Holtzclaw, L.A.; Scholz, S.; Cookson, M.R.; Houlden, H.; Gwinn-Hardy, K.; Fung, H.C.; Lin, X.; et al. Deletion at ITPR1 underlies ataxia in mice and spinocerebellar ataxia 15 in humans. PLoS Genet. 2007, 3, e108. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Chardon, J.W.; Carter, M.T.; Friend, K.L.; Dudding, T.E.; Schwartzentruber, J.; Zou, R.; Schofield, P.W.; Douglas, S.; Bulman, D.E.; et al. Missense mutations in ITPR1 cause autosomal dominant congenital nonprogressive spinocerebellar ataxia. Orphanet J. Rare Dis. 2012, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Gerber, S.; Alzayady, K.J.; Burglen, L.; Bremond-Gignac, D.; Marchesin, V.; Roche, O.; Rio, M.; Funalot, B.; Calmon, R.; Durr, A.; et al. Recessive and Dominant De Novo ITPR1 Mutations Cause Gillespie Syndrome. Am. J. Hum. Genet. 2016, 98, 971–980. [Google Scholar] [CrossRef] [Green Version]
- Klar, J.; Hisatsune, C.; Baig, S.M.; Tariq, M.; Johansson, A.C.; Rasool, M.; Malik, N.A.; Ameur, A.; Sugiura, K.; Feuk, L.; et al. Abolished InsP3R2 function inhibits sweat secretion in both humans and mice. J. Clin. Investig. 2014, 124, 4773–4780. [Google Scholar] [CrossRef]
- MacLennan, D.H.; Duff, C.; Zorzato, F.; Fujii, J.; Phillips, M.; Korneluk, R.G.; Frodis, W.; Britt, B.A.; Worton, R.G. Ryanodine receptor gene is a candidate for predisposition to malignant hyperthermia. Nature 1990, 343, 559–561. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, H.S.; Khanna, V.K.; De Leon, S.; Phillips, M.S.; Schappert, K.; Britt, B.A.; Browell, A.K.; MacLennan, D.H. A mutation in the human ryanodine receptor gene associated with central core disease. Nat. Genet. 1993, 5, 46–50. [Google Scholar] [CrossRef]
- Quane, K.A.; Healy, J.M.; Keating, K.E.; Manning, B.M.; Couch, F.J.; Palmucci, L.M.; Doriguzzi, C.; Fagerlund, T.H.; Berg, K.; Ording, H.; et al. Mutations in the ryanodine receptor gene in central core disease and malignant hyperthermia. Nat. Genet. 1993, 5, 51–55. [Google Scholar] [CrossRef]
- Monnier, N.; Ferreiro, A.; Marty, I.; Labarre-Vila, A.; Mezin, P.; Lunardi, J. A homozygous splicing mutation causing a depletion of skeletal muscle RYR1 is associated with multi-minicore disease congenital myopathy with ophthalmoplegia. Hum. Mol. Genet. 2003, 12, 1171–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiso, N.; Stephan, D.A.; Nava, A.; Bagattin, A.; Devaney, J.M.; Stanchi, F.; Larderet, G.; Brahmbhatt, B.; Brown, K.; Bauce, B.; et al. Identification of mutations in the cardiac ryanodine receptor gene in families affected with arrhythmogenic right ventricular cardiomyopathy type 2 (ARVD2). Hum. Mol. Genet. 2001, 10, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priori, S.G.; Napolitano, C.; Tiso, N.; Memmi, M.; Vignati, G.; Bloise, R.; Sorrentino, V.; Danieli, G.A. Mutations in the cardiac ryanodine receptor gene (hRyR2) underlie catecholaminergic polymorphic ventricular tachycardia. Circulation 2001, 103, 196–200. [Google Scholar] [CrossRef] [Green Version]
- Foulquier, F.; Amyere, M.; Jaeken, J.; Zeevaert, R.; Schollen, E.; Race, V.; Bammens, R.; Morelle, W.; Rosnoblet, C.; Legrand, D.; et al. TMEM165 deficiency causes a congenital disorder of glycosylation. Am. J. Hum. Genet. 2012, 91, 15–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anzilotti, C.; Swan, D.J.; Boisson, B.; Deobagkar-Lele, M.; Oliveira, C.; Chabosseau, P.; Engelhardt, K.R.; Xu, X.; Chen, R.; Alvarez, L.; et al. An essential role for the Zn(2+) transporter ZIP7 in B cell development. Nat. Immunol. 2019, 20, 350–361. [Google Scholar] [CrossRef] [Green Version]
- Giunta, C.; Elcioglu, N.H.; Albrecht, B.; Eich, G.; Chambaz, C.; Janecke, A.R.; Yeowell, H.; Weis, M.; Eyre, D.R.; Kraenzlin, M.; et al. Spondylocheiro dysplastic form of the Ehlers-Danlos syndrome—An autosomal-recessive entity caused by mutations in the zinc transporter gene SLC39A13. Am. J. Hum. Genet. 2008, 82, 1290–1305. [Google Scholar] [CrossRef] [Green Version]
- Vulpe, C.; Levinson, B.; Whitney, S.; Packman, S.; Gitschier, J. Isolation of a candidate gene for Menkes disease and evidence that it encodes a copper-transporting ATPase. Nat. Genet. 1993, 3, 7–13. [Google Scholar] [CrossRef]
- Chelly, J.; Tumer, Z.; Tonnesen, T.; Petterson, A.; Ishikawa-Brush, Y.; Tommerup, N.; Horn, N.; Monaco, A.P. Isolation of a candidate gene for Menkes disease that encodes a potential heavy metal binding protein. Nat. Genet. 1993, 3, 14–19. [Google Scholar] [CrossRef]
- Mercer, J.F.; Livingston, J.; Hall, B.; Paynter, J.A.; Begy, C.; Chandrasekharappa, S.; Lockhart, P.; Grimes, A.; Bhave, M.; Siemieniak, D.; et al. Isolation of a partial candidate gene for Menkes disease by positional cloning. Nat. Genet. 1993, 3, 20–25. [Google Scholar] [CrossRef]
- Kaler, S.G.; Gallo, L.K.; Proud, V.K.; Percy, A.K.; Mark, Y.; Segal, N.A.; Goldstein, D.S.; Holmes, C.S.; Gahl, W.A. Occipital horn syndrome and a mild Menkes phenotype associated with splice site mutations at the MNK locus. Nat. Genet. 1994, 8, 195–202. [Google Scholar] [CrossRef]
- Kennerson, M.L.; Nicholson, G.A.; Kaler, S.G.; Kowalski, B.; Mercer, J.F.; Tang, J.; Llanos, R.M.; Chu, S.; Takata, R.I.; Speck-Martins, C.E.; et al. Missense mutations in the copper transporter gene ATP7A cause X-linked distal hereditary motor neuropathy. Am. J. Hum. Genet. 2010, 86, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Bull, P.C.; Thomas, G.R.; Rommens, J.M.; Forbes, J.R.; Cox, D.W. The Wilson disease gene is a putative copper transporting P-type ATPase similar to the Menkes gene. Nat. Genet. 1993, 5, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Tanzi, R.E.; Petrukhin, K.; Chernov, I.; Pellequer, J.L.; Wasco, W.; Ross, B.; Romano, D.M.; Parano, E.; Pavone, L.; Brzustowicz, L.M.; et al. The Wilson disease gene is a copper transporting ATPase with homology to the Menkes disease gene. Nat. Genet. 1993, 5, 344–350. [Google Scholar] [CrossRef]
- Chandra, S.; Kable, E.P.; Morrison, G.H.; Webb, W.W. Calcium sequestration in the Golgi apparatus of cultured mammalian cells revealed by laser scanning confocal microscopy and ion microscopy. J. Cell Sci. 1991, 100 Pt 4, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Pozzan, T.; Rizzuto, R. The Golgi apparatus is an inositol 1,4,5-trisphosphate-sensitive Ca2+ store, with functional properties distinct from those of the endoplasmic reticulum. EMBO J. 1998, 17, 5298–5308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pizzo, P.; Lissandron, V.; Capitanio, P.; Pozzan, T. Ca2+ signalling in the Golgi apparatus. Cell Calcium 2011, 50, 184–192. [Google Scholar] [CrossRef]
- Vangheluwe, P.; Sepulveda, M.R.; Missiaen, L.; Raeymaekers, L.; Wuytack, F.; Vanoevelen, J. Intracellular Ca2+- and Mn2+-transport ATPases. Chem. Rev. 2009, 109, 4733–4759. [Google Scholar] [CrossRef] [PubMed]
- Dally, S.; Bredoux, R.; Corvazier, E.; Andersen, J.P.; Clausen, J.D.; Dode, L.; Fanchaouy, M.; Gelebart, P.; Monceau, V.; Del Monte, F.; et al. Ca2+-ATPases in non-failing and failing heart: Evidence for a novel cardiac sarco/endoplasmic reticulum Ca2+-ATPase 2 isoform (SERCA2c). Biochem. J. 2006, 395, 249–258. [Google Scholar] [CrossRef]
- Wuytack, F.; Raeymaekers, L.; Missiaen, L. Molecular physiology of the SERCA and SPCA pumps. Cell Calcium 2002, 32, 279–305. [Google Scholar] [CrossRef] [PubMed]
- Sudbrak, R.; Brown, J.; Dobson-Stone, C.; Carter, S.; Ramser, J.; White, J.; Healy, E.; Dissanayake, M.; Larregue, M.; Perrussel, M.; et al. Hailey-Hailey disease is caused by mutations in ATP2C1 encoding a novel Ca2+ pump. Hum. Mol. Genet. 2000, 9, 1131–1140. [Google Scholar] [CrossRef]
- Xiang, M.; Mohamalawari, D.; Rao, R. A novel isoform of the secretory pathway Ca2+,Mn2+-ATPase, hSPCA2, has unusual properties and is expressed in the brain. J. Biol. Chem. 2005, 280, 11608–11614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Smaardijk, S.; Mattelaer, C.A.; Pamula, F.; Vandecaetsbeek, I.; Vanoevelen, J.; Wuytack, F.; Lescrinier, E.; Eggermont, J.; Vangheluwe, P. An N-terminal Ca2+-binding motif regulates the secretory pathway Ca2+/Mn2+-transport ATPase SPCA1. J. Biol. Chem. 2019, 294, 7878–7891. [Google Scholar] [CrossRef]
- Dode, L.; Andersen, J.P.; Vanoevelen, J.; Raeymaekers, L.; Missiaen, L.; Vilsen, B.; Wuytack, F. Dissection of the functional differences between human secretory pathway Ca2+/Mn2+-ATPase (SPCA) 1 and 2 isoenzymes by steady-state and transient kinetic analyses. J. Biol. Chem. 2006, 281, 3182–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dode, L.; Andersen, J.P.; Raeymaekers, L.; Missiaen, L.; Vilsen, B.; Wuytack, F. Functional comparison between secretory pathway Ca2+/Mn2+-ATPase (SPCA) 1 and sarcoplasmic reticulum Ca2+-ATPase (SERCA) 1 isoforms by steady-state and transient kinetic analyses. J. Biol. Chem. 2005, 280, 39124–39134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanoevelen, J.; Raeymaekers, L.; Parys, J.B.; De Smedt, H.; Van Baelen, K.; Callewaert, G.; Wuytack, F.; Missiaen, L. Inositol trisphosphate producing agonists do not mobilize the thapsigargin-insensitive part of the endoplasmic-reticulum and Golgi Ca2+ store. Cell Calcium 2004, 35, 115–121. [Google Scholar] [CrossRef]
- Aulestia, F.J.; Alonso, M.T.; Garcia-Sancho, J. Differential calcium handling by the cis and trans regions of the Golgi apparatus. Biochem. J. 2015, 466, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Behne, M.J.; Tu, C.L.; Aronchik, I.; Epstein, E.; Bench, G.; Bikle, D.D.; Pozzan, T.; Mauro, T.M. Human keratinocyte ATP2C1 localizes to the Golgi and controls Golgi Ca2+ stores. J. Investig. Dermatol. 2003, 121, 688–694. [Google Scholar] [CrossRef] [Green Version]
- Micaroni, M.; Perinetti, G.; Berrie, C.P.; Mironov, A.A. The SPCA1 Ca2+ pump and intracellular membrane trafficking. Traffic 2010, 11, 1315–1333. [Google Scholar] [CrossRef] [PubMed]
- Rosnoblet, C.; Legrand, D.; Demaegd, D.; Hacine-Gherbi, H.; de Bettignies, G.; Bammens, R.; Borrego, C.; Duvet, S.; Morsomme, P.; Matthijs, G.; et al. Impact of disease-causing mutations on TMEM165 subcellular localization, a recently identified protein involved in CDG-II. Hum. Mol. Genet. 2013, 22, 2914–2928. [Google Scholar] [CrossRef] [Green Version]
- Demaegd, D.; Foulquier, F.; Colinet, A.S.; Gremillon, L.; Legrand, D.; Mariot, P.; Peiter, E.; Van Schaftingen, E.; Matthijs, G.; Morsomme, P. Newly characterized Golgi-localized family of proteins is involved in calcium and pH homeostasis in yeast and human cells. Proc. Natl. Acad. Sci. USA 2013, 110, 6859–6864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potelle, S.; Morelle, W.; Dulary, E.; Duvet, S.; Vicogne, D.; Spriet, C.; Krzewinski-Recchi, M.A.; Morsomme, P.; Jaeken, J.; Matthijs, G.; et al. Glycosylation abnormalities in Gdt1p/TMEM165 deficient cells result from a defect in Golgi manganese homeostasis. Hum. Mol. Genet. 2016, 25, 1489–1500. [Google Scholar] [CrossRef] [Green Version]
- Potelle, S.; Dulary, E.; Climer, L.; Duvet, S.; Morelle, W.; Vicogne, D.; Lebredonchel, E.; Houdou, M.; Spriet, C.; Krzewinski-Recchi, M.A.; et al. Manganese-induced turnover of TMEM165. Biochem. J. 2017, 474, 1481–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebredonchel, E.; Houdou, M.; Hoffmann, H.H.; Kondratska, K.; Krzewinski, M.A.; Vicogne, D.; Rice, C.M.; Klein, A.; Foulquier, F. Investigating the functional link between TMEM165 and SPCA1. Biochem. J. 2019, 476, 3281–3293. [Google Scholar] [CrossRef]
- Lin, P.; Yao, Y.; Hofmeister, R.; Tsien, R.Y.; Farquhar, M.G. Overexpression of CALNUC (nucleobindin) increases agonist and thapsigargin releasable Ca2+ storage in the Golgi. J. Cell. Biol. 1999, 145, 279–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzmann, N.; Kunerth, S.; Weber, K.; Mayr, G.W.; Guse, A.H. Knock-down of the type 3 ryanodine receptor impairs sustained Ca2+ signaling via the T cell receptor/CD3 complex. J. Biol. Chem. 2002, 277, 50636–50642. [Google Scholar] [CrossRef] [Green Version]
- Surroca, A.; Wolff, D. Inositol 1,4,5-trisphosphate but not ryanodine-receptor agonists induces calcium release from rat liver Golgi apparatus membrane vesicles. J. Membr. Biol. 2000, 177, 243–249. [Google Scholar] [CrossRef]
- Cifuentes, F.; Gonzalez, C.E.; Fiordelisio, T.; Guerrero, G.; Lai, F.A.; Hernandez-Cruz, A. A ryanodine fluorescent derivative reveals the presence of high-affinity ryanodine binding sites in the Golgi complex of rat sympathetic neurons, with possible functional roles in intracellular Ca2+ signaling. Cell Signal 2001, 13, 353–362. [Google Scholar] [CrossRef]
- Lissandron, V.; Podini, P.; Pizzo, P.; Pozzan, T. Unique characteristics of Ca2+ homeostasis of the trans-Golgi compartment. Proc. Natl. Acad. Sci. USA 2010, 107, 9198–9203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallegos-Gomez, M.L.; Greotti, E.; Lopez-Mendez, M.C.; Sanchez-Vazquez, V.H.; Arias, J.M.; Guerrero-Hernandez, A. The Trans Golgi Region is a Labile Intracellular Ca2+ Store Sensitive to Emetine. Sci. Rep. 2018, 8, 17143. [Google Scholar] [CrossRef]
- Lin, P.; Le-Niculescu, H.; Hofmeister, R.; McCaffery, J.M.; Jin, M.; Hennemann, H.; McQuistan, T.; De Vries, L.; Farquhar, M.G. The mammalian calcium-binding protein, nucleobindin (CALNUC), is a Golgi resident protein. J. Cell. Biol. 1998, 141, 1515–1527. [Google Scholar] [CrossRef]
- Scherer, P.E.; Lederkremer, G.Z.; Williams, S.; Fogliano, M.; Baldini, G.; Lodish, H.F. Cab45, a novel (Ca2+)-binding protein localized to the Golgi lumen. J. Cell. Biol. 1996, 133, 257–268. [Google Scholar] [CrossRef]
- Von Blume, J.; Alleaume, A.M.; Kienzle, C.; Carreras-Sureda, A.; Valverde, M.; Malhotra, V. Cab45 is required for Ca2+-dependent secretory cargo sorting at the trans-Golgi network. J. Cell. Biol. 2012, 199, 1057–1066. [Google Scholar] [CrossRef] [Green Version]
- Hecht, T.K.; Blank, B.; Steger, M.; Lopez, V.; Beck, G.; Ramazanov, B.; Mann, M.; Tagliabracci, V.; von Blume, J. Fam20C regulates protein secretion by Cab45 phosphorylation. J. Cell. Biol. 2020, 219, e201910089. [Google Scholar] [CrossRef] [PubMed]
- Crevenna, A.H.; Blank, B.; Maiser, A.; Emin, D.; Prescher, J.; Beck, G.; Kienzle, C.; Bartnik, K.; Habermann, B.; Pakdel, M.; et al. Secretory cargo sorting by Ca2+-dependent Cab45 oligomerization at the trans-Golgi network. J. Cell. Biol. 2016, 213, 305–314. [Google Scholar] [CrossRef]
- Morel-Huaux, V.M.; Pypaert, M.; Wouters, S.; Tartakoff, A.M.; Jurgan, U.; Gevaert, K.; Courtoy, P.J. The calcium-binding protein p54/NEFA is a novel luminal resident of medial Golgi cisternae that traffics independently of mannosidase II. Eur. J. Cell. Biol. 2002, 81, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Karabinos, A.; Bhattacharya, D.; Morys-Wortmann, C.; Kroll, K.; Hirschfeld, G.; Kratzin, H.D.; Barnikol-Watanabe, S.; Hilschmann, N. The divergent domains of the NEFA and nucleobindin proteins are derived from an EF-hand ancestor. Mol. Biol. Evol. 1996, 13, 990–998. [Google Scholar] [CrossRef] [Green Version]
- Yabe, D.; Nakamura, T.; Kanazawa, N.; Tashiro, K.; Honjo, T. Calumenin, a Ca2+-binding protein retained in the endoplasmic reticulum with a novel carboxyl-terminal sequence, HDEF. J. Biol. Chem. 1997, 272, 18232–18239. [Google Scholar] [CrossRef] [Green Version]
- Vorum, H.; Hager, H.; Christensen, B.M.; Nielsen, S.; Honore, B. Human calumenin localizes to the secretory pathway and is secreted to the medium. Exp. Cell Res. 1999, 248, 473–481. [Google Scholar] [CrossRef]
- Jung, D.H.; Mo, S.H.; Kim, D.H. Calumenin, a multiple EF-hands Ca2+-binding protein, interacts with ryanodine receptor-1 in rabbit skeletal sarcoplasmic reticulum. Biochem. Biophys. Res. Commun. 2006, 343, 34–42. [Google Scholar] [CrossRef]
- Sahoo, S.K.; Kim, D.H. Calumenin interacts with SERCA2 in rat cardiac sarcoplasmic reticulum. Mol. Cells 2008, 26, 265–269. [Google Scholar]
- Sahoo, S.K.; Kim, T.; Kang, G.B.; Lee, J.G.; Eom, S.H.; Kim, D.H. Characterization of calumenin-SERCA2 interaction in mouse cardiac sarcoplasmic reticulum. J. Biol. Chem. 2009, 284, 31109–31121. [Google Scholar] [CrossRef] [Green Version]
- Ireland, S.; Ramnarayanan, S.; Fu, M.; Zhang, X.; Zhang, J.; Li, J.; Emebo, D.; Wang, Y. Cytosolic Ca2+ Modulates Golgi Structure Through PKCalpha-Mediated GRASP55 Phosphorylation. iScience 2020, 23, 100952. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y. GRASPs in Golgi Structure and Function. Front. Cell Dev. Biol. 2015, 3, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, Y. Nonredundant Roles of GRASP55 and GRASP65 in the Golgi Apparatus and Beyond. Trends Biochem. Sci. 2020, 45, 1065–1079. [Google Scholar] [CrossRef] [PubMed]
- Tempel, W.; Karaveg, K.; Liu, Z.J.; Rose, J.; Wang, B.C.; Moremen, K.W. Structure of mouse Golgi alpha-mannosidase IA reveals the molecular basis for substrate specificity among class 1 (family 47 glycosylhydrolase) alpha1,2-mannosidases. J. Biol. Chem. 2004, 279, 29774–29786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.D.; VanSlyke, J.K.; Thulin, C.D.; Jean, F.; Thomas, G. Activation of the furin endoprotease is a multiple-step process: Requirements for acidification and internal propeptide cleavage. EMBO J. 1997, 16, 1508–1518. [Google Scholar] [CrossRef] [Green Version]
- Schutzbach, J.S.; Forsee, W.T. Calcium ion activation of rabbit liver alpha 1,2-mannosidase. J. Biol. Chem. 1990, 265, 2546–2549. [Google Scholar] [CrossRef]
- Nishikawa, Y.; Pegg, W.; Paulsen, H.; Schachter, H. Control of glycoprotein synthesis. Purification and characterization of rabbit liver UDP-N-acetylglucosamine:alpha-3-D-mannoside beta-1,2-N-acetylglucosaminyltransferase I. J. Biol. Chem. 1988, 263, 8270–8281. [Google Scholar] [CrossRef]
- Fritz, T.A.; Hurley, J.H.; Trinh, L.B.; Shiloach, J.; Tabak, L.A. The beginnings of mucin biosynthesis: The crystal structure of UDP-GalNAc:polypeptide alpha-N-acetylgalactosaminyltransferase-T1. Proc. Natl. Acad. Sci. USA 2004, 101, 15307–15312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiggins, C.A.; Munro, S. Activity of the yeast MNN1 alpha-1,3-mannosyltransferase requires a motif conserved in many other families of glycosyltransferases. Proc. Natl. Acad. Sci. USA 1998, 95, 7945–7950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, A.S.; Morais, V.A.; Coelho, A.V.; Costa, J. Effect of the manganese ion on human alpha3/4 fucosyltransferase III activity. Biometals 2004, 17, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Brito, C.; Kandzia, S.; Graca, T.; Conradt, H.S.; Costa, J. Human fucosyltransferase IX: Specificity towards N-linked glycoproteins and relevance of the cytoplasmic domain in intra-Golgi localization. Biochimie 2008, 90, 1279–1290. [Google Scholar] [CrossRef]
- Missiaen, L.; Raeymaekers, L.; Dode, L.; Vanoevelen, J.; Van Baelen, K.; Parys, J.B.; Callewaert, G.; De Smedt, H.; Segaert, S.; Wuytack, F. SPCA1 pumps and Hailey-Hailey disease. Biochem. Biophys. Res. Commun. 2004, 322, 1204–1213. [Google Scholar] [CrossRef]
- Olanow, C.W. Manganese-induced parkinsonism and Parkinson’s disease. Ann. N. Y. Acad. Sci. 2004, 1012, 209–223. [Google Scholar] [CrossRef]
- Kaufman, R.J.; Swaroop, M.; Murtha-Riel, P. Depletion of manganese within the secretory pathway inhibits O-linked glycosylation in mammalian cells. Biochemistry 1994, 33, 9813–9819. [Google Scholar] [CrossRef]
- Vanoevelen, J.; Dode, L.; Van Baelen, K.; Fairclough, R.J.; Missiaen, L.; Raeymaekers, L.; Wuytack, F. The secretory pathway Ca2+/Mn2+-ATPase 2 is a Golgi-localized pump with high affinity for Ca2+ ions. J. Biol. Chem. 2005, 280, 22800–22808. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Linstedt, A.D. Identification of a gain-of-function mutation in a Golgi P-type ATPase that enhances Mn2+ efflux and protects against toxicity. Proc. Natl. Acad. Sci. USA 2011, 108, 858–863. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Linstedt, A.D. Manganese blocks intracellular trafficking of Shiga toxin and protects against Shiga toxicosis. Science 2012, 335, 332–335. [Google Scholar] [CrossRef] [Green Version]
- Fraser, M.E.; Chernaia, M.M.; Kozlov, Y.V.; James, M.N. Crystal structure of the holotoxin from Shigella dysenteriae at 2.5 A resolution. Nat. Struct. Biol. 1994, 1, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Spooner, R.A.; Lord, J.M. How ricin and Shiga toxin reach the cytosol of target cells: Retrotranslocation from the endoplasmic reticulum. Curr. Top. Microbiol. Immunol. 2012, 357, 19–40. [Google Scholar] [PubMed] [Green Version]
- Forrester, A.; Rathjen, S.J.; Daniela Garcia-Castillo, M.; Bachert, C.; Couhert, A.; Tepshi, L.; Pichard, S.; Martinez, J.; Munier, M.; Sierocki, R.; et al. Functional dissection of the retrograde Shiga toxin trafficking inhibitor Retro-2. Nat. Chem. Biol. 2020, 16, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Tewari, R.; Jarvela, T.; Linstedt, A.D. Manganese induces oligomerization to promote down-regulation of the intracellular trafficking receptor used by Shiga toxin. Mol. Biol. Cell 2014, 25, 3049–3058. [Google Scholar] [CrossRef]
- Tewari, R.; Bachert, C.; Linstedt, A.D. Induced oligomerization targets Golgi proteins for degradation in lysosomes. Mol. Biol. Cell. 2015, 26, 4427–4437. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Bachert, C.; Smith, D.R.; Linstedt, A.D. Manganese-induced trafficking and turnover of the cis-Golgi glycoprotein GPP130. Mol. Biol. Cell 2010, 21, 1282–1292. [Google Scholar] [CrossRef] [Green Version]
- Beckers, C.J.; Balch, W.E. Calcium and GTP: Essential components in vesicular trafficking between the endoplasmic reticulum and Golgi apparatus. J. Cell. Biol. 1989, 108, 1245–1256. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.I.; Beron, W.; Stahl, P.D. Calmodulin regulates endosome fusion. J. Biol. Chem. 1997, 272, 7707–7712. [Google Scholar] [CrossRef] [Green Version]
- Pryor, P.R.; Mullock, B.M.; Bright, N.A.; Gray, S.R.; Luzio, J.P. The role of intraorganellar Ca2+ in late endosome-lysosome heterotypic fusion and in the reformation of lysosomes from hybrid organelles. J. Cell. Biol. 2000, 149, 1053–1062. [Google Scholar] [CrossRef] [Green Version]
- Di Jeso, B.; Pereira, R.; Consiglio, E.; Formisano, S.; Satrustegui, J.; Sandoval, I.V. Demonstration of a Ca2+ requirement for thyroglobulin dimerization and export to the golgi complex. Eur. J. Biochem. 1998, 252, 583–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivessa, N.E.; De Lemos-Chiarandini, C.; Gravotta, D.; Sabatini, D.D.; Kreibich, G. The Brefeldin A-induced retrograde transport from the Golgi apparatus to the endoplasmic reticulum depends on calcium sequestered to intracellular stores. J. Biol. Chem. 1995, 270, 25960–25967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micaroni, M. Calcium around the Golgi apparatus: Implications for intracellular membrane trafficking. Adv. Exp. Med. Biol. 2012, 740, 439–460. [Google Scholar] [PubMed]
- Porat, A.; Elazar, Z. Regulation of intra-Golgi membrane transport by calcium. J. Biol. Chem. 2000, 275, 29233–29237. [Google Scholar] [CrossRef] [Green Version]
- Huttner, W.B.; Ohashi, M.; Kehlenbach, R.H.; Barr, F.A.; Bauerfeind, R.; Braunling, O.; Corbeil, D.; Hannah, M.; Pasolli, H.A.; Schmidt, A.; et al. Biogenesis of neurosecretory vesicles. Cold Spring Harb. Symp. Quant. Biol. 1995, 60, 315–327. [Google Scholar] [CrossRef]
- Von Blume, J.; Alleaume, A.M.; Cantero-Recasens, G.; Curwin, A.; Carreras-Sureda, A.; Zimmermann, T.; van Galen, J.; Wakana, Y.; Valverde, M.A.; Malhotra, V. ADF/cofilin regulates secretory cargo sorting at the TGN via the Ca2+ ATPase SPCA1. Dev. Cell 2011, 20, 652–662. [Google Scholar] [CrossRef] [Green Version]
- Okunade, G.W.; Miller, M.L.; Azhar, M.; Andringa, A.; Sanford, L.P.; Doetschman, T.; Prasad, V.; Shull, G.E. Loss of the Atp2c1 secretory pathway Ca2+-ATPase (SPCA1) in mice causes Golgi stress, apoptosis, and midgestational death in homozygous embryos and squamous cell tumors in adult heterozygotes. J. Biol. Chem. 2007, 282, 26517–26527. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Pakdel, M.; Blank, B.; Sundberg, E.L.; Burd, C.G.; von Blume, J. Activity of the SPCA1 Calcium Pump Couples Sphingomyelin Synthesis to Sorting of Secretory Proteins in the Trans-Golgi Network. Dev. Cell 2018, 47, 464–478.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.H.; Tian, X.R.; Jiang, Z.; Zeng, L.W.; He, W.F.; Hu, Z.P. The Golgi Apparatus: Panel Point of Cytosolic Ca2+ Regulation. Neurosignals 2013, 21, 272–284. [Google Scholar] [CrossRef]
- Hoffmann, H.H.; Schneider, W.M.; Blomen, V.A.; Scull, M.A.; Hovnanian, A.; Brummelkamp, T.R.; Rice, C.M. Diverse Viruses Require the Calcium Transporter SPCA1 for Maturation and Spread. Cell Host Microbe 2017, 22, 460–470.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Zheng, M.; Yang, Y.; Gu, X.; Yang, K.; Li, M.; Liu, Y.; Zhang, Q.; Zhang, P.; Wang, Y.; et al. Furin: A Potential Therapeutic Target for COVID-19. iScience 2020, 23, 101642. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Banci, L.; Bertini, I.; Rosato, A. Counting the zinc-proteins encoded in the human genome. J. Proteome Res. 2006, 5, 196–201. [Google Scholar] [CrossRef]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The Physiological, Biochemical, and Molecular Roles of Zinc Transporters in Zinc Homeostasis and Metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef] [PubMed]
- Bellomo, E.; Hogstrand, C.; Maret, W. Redox and zinc signalling pathways converging on protein tyrosine phosphatases. Free Radic. Biol. Med. 2014, 75 (Suppl. S1), S9. [Google Scholar] [CrossRef]
- Law, J.S.; McBride, S.A.; Graham, S.; Nelson, N.R.; Slotnick, B.M.; Henkin, R.I. Zinc deficiency decreases the activity of calmodulin regulated cyclic nucleotide phosphodiesterases in vivo in selected rat tissues. Biol. Trace Elem. Res. 1988, 16, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Truong-Tran, A.Q.; Carter, J.; Ruffin, R.E.; Zalewski, P.D. The role of zinc in caspase activation and apoptotic cell death. Biometals 2001, 14, 315–330. [Google Scholar] [CrossRef]
- Barthel, A.; Ostrakhovitch, E.A.; Walter, P.L.; Kampkotter, A.; Klotz, L.O. Stimulation of phosphoinositide 3-kinase/Akt signaling by copper and zinc ions: Mechanisms and consequences. Arch. Biochem. Biophys. 2007, 463, 175–182. [Google Scholar] [CrossRef]
- Zhang, Y.; Aizenman, E.; DeFranco, D.B.; Rosenberg, P.A. Intracellular zinc release, 12-lipoxygenase activation and MAPK dependent neuronal and oligodendroglial death. Mol. Med. 2007, 13, 350–355. [Google Scholar] [CrossRef]
- Wu, F.Y.; Huang, W.J.; Sinclair, R.B.; Powers, L. The structure of the zinc sites of Escherichia coli DNA-dependent RNA polymerase. J. Biol. Chem. 1992, 267, 25560–25567. [Google Scholar] [CrossRef]
- Wu, F.Y.; Wu, C.W. Zinc in DNA replication and transcription. Annu. Rev. Nutr. 1987, 7, 251–272. [Google Scholar] [CrossRef]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [CrossRef]
- Kirschke, C.P.; Huang, L. ZnT7, a novel mammalian zinc transporter, accumulates zinc in the Golgi apparatus. J. Biol. Chem. 2003, 278, 4096–4102. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Ishihara, K.; Migaki, H.; Ishihara, K.; Nagao, M.; Yamaguchi-Iwai, Y.; Kambe, T. Two different zinc transport complexes of cation diffusion facilitator proteins localized in the secretory pathway operate to activate alkaline phosphatases in vertebrate cells. J. Biol. Chem. 2005, 280, 30956–30962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, N.H.; Kelleher, S.L. ZnT4 provides zinc to zinc-dependent proteins in the trans-Golgi network critical for cell function and Zn export in mammary epithelial cells. Am. J. Physiol. Cell Physiol. 2012, 303, C291–C297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Kirschke, C.P.; Gitschier, J. Functional characterization of a novel mammalian zinc transporter, ZnT6. J. Biol. Chem. 2002, 277, 26389–26395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bin, B.H.; Fukada, T.; Hosaka, T.; Yamasaki, S.; Ohashi, W.; Hojyo, S.; Miyai, T.; Nishida, K.; Yokoyama, S.; Hirano, T. Biochemical characterization of human ZIP13 protein: A homo-dimerized zinc transporter involved in the spondylocheiro dysplastic Ehlers-Danlos syndrome. J. Biol. Chem. 2011, 286, 40255–40265. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Kirschke, C.P.; Zhang, Y.; Yu, Y.Y. The ZIP7 gene (Slc39a7) encodes a zinc transporter involved in zinc homeostasis of the Golgi apparatus. J. Biol. Chem. 2005, 280, 15456–15463. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, W.; Yamazaki, T.; Yamaguchi-Iwai, Y.; Masuda, S.; Nagao, M.; Andrews, G.K.; Kambe, T. SLC39A9 (ZIP9) regulates zinc homeostasis in the secretory pathway: Characterization of the ZIP subfamily I protein in vertebrate cells. Biosci. Biotechnol. Biochem. 2009, 73, 1142–1148. [Google Scholar] [CrossRef] [Green Version]
- Kelleher, S.L.; Velasquez, V.; Croxford, T.P.; McCormick, N.H.; Lopez, V.; MacDavid, J. Mapping the zinc-transporting system in mammary cells: Molecular analysis reveals a phenotype-dependent zinc-transporting network during lactation. J. Cell Physiol. 2012, 227, 1761–1770. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.; Walker, J.M.; Wang, F.; Park, J.G.; Palmer, A.E.; Giunta, C.; Rohrbach, M.; Steinmann, B.; Eide, D.J. Promotion of vesicular zinc efflux by ZIP13 and its implications for spondylocheiro dysplastic Ehlers-Danlos syndrome. Proc. Natl. Acad. Sci. USA 2012, 109, E3530–E3538. [Google Scholar] [CrossRef] [Green Version]
- Kambe, T.; Takeda, T.A.; Nishito, Y. Activation of zinc-requiring ectoenzymes by ZnT transporters during the secretory process: Biochemical and molecular aspects. Arch. Biochem. Biophys. 2016, 611, 37–42. [Google Scholar] [CrossRef]
- Suzuki, E.; Ogawa, N.; Takeda, T.A.; Nishito, Y.; Tanaka, Y.K.; Fujiwara, T.; Matsunaga, M.; Ueda, S.; Kubo, N.; Tsuji, T.; et al. Detailed analyses of the crucial functions of Zn transporter proteins in alkaline phosphatase activation. J. Biol. Chem. 2020, 295, 5669–5684. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Ishihara, K.; Migaki, H.; Matsuura, W.; Kohda, A.; Okumura, K.; Nagao, M.; Yamaguchi-Iwai, Y.; Kambe, T. Zinc transporters, ZnT5 and ZnT7, are required for the activation of alkaline phosphatases, zinc-requiring enzymes that are glycosylphosphatidylinositol-anchored to the cytoplasmic membrane. J. Biol. Chem. 2005, 280, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Van den Elsen, J.M.; Kuntz, D.A.; Rose, D.R. Structure of Golgi alpha-mannosidase II: A target for inhibition of growth and metastasis of cancer cells. EMBO J. 2001, 20, 3008–3017. [Google Scholar] [CrossRef]
- Permyakov, E.A.; Reyzer, I.L.; Berliner, L.J. Effects of Zn(II) on galactosyltransferase activity. J. Protein. Chem. 1993, 12, 633–638. [Google Scholar] [CrossRef]
- Lee, G.M.; Bin, B.H. Different Actions of Intracellular Zinc Transporters ZIP7 and ZIP13 Are Essential for Dermal Development. Int. J. Mol. Sci. 2019, 20, 3941. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Dempski, R.E.; Burdette, S.C. Zn(2+) at a cellular crossroads. Curr. Opin. Chem. Biol. 2016, 31, 120–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishihara, K.; Yamazaki, T.; Ishida, Y.; Suzuki, T.; Oda, K.; Nagao, M.; Yamaguchi-Iwai, Y.; Kambe, T. Zinc transport complexes contribute to the homeostatic maintenance of secretory pathway function in vertebrate cells. J. Biol. Chem. 2006, 281, 17743–17750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, W.; Kimura, S.; Iwanaga, T.; Furusawa, Y.; Irie, T.; Izumi, H.; Watanabe, T.; Hijikata, A.; Hara, T.; Ohara, O.; et al. Zinc Transporter SLC39A7/ZIP7 Promotes Intestinal Epithelial Self-Renewal by Resolving ER Stress. PLoS Genet. 2016, 12, e1006349. [Google Scholar] [CrossRef]
- Bafaro, E.; Liu, Y.; Xu, Y.; Dempski, R.E. The emerging role of zinc transporters in cellular homeostasis and cancer. Signal Transduct. Target. Ther. 2017, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukunaka, A.; Fujitani, Y. Role of Zinc Homeostasis in the Pathogenesis of Diabetes and Obesity. Int. J. Mol. Sci. 2018, 19, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adulcikas, J.; Sonda, S.; Norouzi, S.; Sohal, S.S.; Myers, S. Targeting the Zinc Transporter ZIP7 in the Treatment of Insulin Resistance and Type 2 Diabetes. Nutrients 2019, 11, 408. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Hong, D.K.; Choi, B.Y.; Suh, S.W. Zinc in the Brain: Friend or Foe? Int. J. Mol. Sci. 2020, 21, 8941. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.S. Zinc: An antioxidant and anti-inflammatory agent: Role of zinc in degenerative disorders of aging. J. Trace Elem. Med. Biol. 2014, 28, 364–371. [Google Scholar] [CrossRef]
- Gammoh, N.Z.; Rink, L. Zinc in Infection and Inflammation. Nutrients 2017, 9, 624. [Google Scholar] [CrossRef] [Green Version]
- Anuk, A.T.; Polat, N.; Akdas, S.; Erol, S.A.; Tanacan, A.; Biriken, D.; Keskin, H.L.; Moraloglu Tekin, O.; Yazihan, N.; Sahin, D. The Relation Between Trace Element Status (Zinc, Copper, Magnesium) and Clinical Outcomes in COVID-19 Infection During Pregnancy. Biol. Trace Elem. Res. 2021, 199, 3608–3617. [Google Scholar] [CrossRef] [PubMed]
- Lovell, M.A.; Smith, J.L.; Markesbery, W.R. Elevated zinc transporter-6 in mild cognitive impairment, Alzheimer disease, and pick disease. J. Neuropathol. Exp. Neurol. 2006, 65, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Xiao, G.; Liu, L.; Lang, M. Zinc transporters in Alzheimer’s disease. Mol. Brain 2019, 12, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, M.; Noguchi, T.; Ikegami, S.; Sakurai, T.; Kakita, A.; Toyoshima, Y.; Kambe, T.; Yamada, M.; Inden, M.; Hara, H.; et al. Zinc transporters ZnT3 and ZnT6 are downregulated in the spinal cords of patients with sporadic amyotrophic lateral sclerosis. J. Neurosci. Res. 2015, 93, 370–379. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Huo, R.; Zhi, Q.; Zhan, M.; Chen, X.; Hua, Z.C. Increased expression of zinc transporter ZIP4, ZIP11, ZnT1, and ZnT6 predicts poor prognosis in pancreatic cancer. J. Trace Elem. Med. Biol. 2021, 65, 126734. [Google Scholar] [CrossRef]
- Singh, C.K.; Malas, K.M.; Tydrick, C.; Siddiqui, I.A.; Iczkowski, K.A.; Ahmad, N. Analysis of Zinc-Exporters Expression in Prostate Cancer. Sci. Rep. 2016, 6, 36772. [Google Scholar] [CrossRef] [Green Version]
- Barresi, V.; Valenti, G.; Spampinato, G.; Musso, N.; Castorina, S.; Rizzarelli, E.; Condorelli, D.F. Transcriptome analysis reveals an altered expression profile of zinc transporters in colorectal cancer. J. Cell Biochem. 2018, 119, 9707–9719. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gordillo-Martinez, F.; Jiang, L.; He, P.; Hong, W.; Wei, X.; Staines, K.A.; Macrae, V.E.; Zhang, C.; Yu, D.; et al. Zinc ameliorates human aortic valve calcification through GPR39 mediated ERK1/2 signalling pathway. Cardiovasc. Res. 2021, 117, 820–835. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Reddy, P.K.; Kumar, B. Trace Element Zinc, a Nature’s Gift to Fight Unprecedented Global Pandemic COVID-19. Biol. Trace Elem. Res. 2021, 199, 3213–3221. [Google Scholar] [CrossRef]
- Cheng, X.; Wang, J.; Liu, C.; Jiang, T.; Yang, N.; Liu, D.; Zhao, H.; Xu, Z. Zinc transporter SLC39A13/ZIP13 facilitates the metastasis of human ovarian cancer cells via activating Src/FAK signaling pathway. J. Exp. Clin. Cancer Res. 2021, 40, 199. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, H.; Xu, Z.; Cheng, X. Zinc dysregulation in cancers and its potential as a therapeutic target. Cancer Biol. Med. 2020, 17, 612–625. [Google Scholar] [CrossRef] [PubMed]
- La Fontaine, S.; Mercer, J.F. Trafficking of the copper-ATPases, ATP7A and ATP7B: Role in copper homeostasis. Arch. Biochem. Biophys. 2007, 463, 149–167. [Google Scholar] [CrossRef]
- Dierick, H.A.; Adam, A.N.; Escara-Wilke, J.F.; Glover, T.W. Immunocytochemical localization of the Menkes copper transport protein (ATP7A) to the trans-Golgi network. Hum. Mol. Genet. 1997, 6, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Braiterman, L.; Nyasae, L.; Guo, Y.; Bustos, R.; Lutsenko, S.; Hubbard, A. Apical targeting and Golgi retention signals reside within a 9-amino acid sequence in the copper-ATPase, ATP7B. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G433–G444. [Google Scholar] [CrossRef]
- Lutsenko, S.; Barnes, N.L.; Bartee, M.Y.; Dmitriev, O.Y. Function and regulation of human copper-transporting ATPases. Physiol. Rev. 2007, 87, 1011–1046. [Google Scholar] [CrossRef]
- Francis, M.J.; Jones, E.E.; Levy, E.R.; Ponnambalam, S.; Chelly, J.; Monaco, A.P. A Golgi localization signal identified in the Menkes recombinant protein. Hum. Mol. Genet. 1998, 7, 1245–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payne, A.S.; Kelly, E.J.; Gitlin, J.D. Functional expression of the Wilson disease protein reveals mislocalization and impaired copper-dependent trafficking of the common H1069Q mutation. Proc. Natl. Acad. Sci. USA 1998, 95, 10854–10859. [Google Scholar] [CrossRef] [Green Version]
- Strausak, D.; La Fontaine, S.; Hill, J.; Firth, S.D.; Lockhart, P.J.; Mercer, J.F. The role of GMXCXXC metal binding sites in the copper-induced redistribution of the Menkes protein. J. Biol. Chem. 1999, 274, 11170–11177. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Shanbhag, V.; Hodgkinson, V.L.; Petris, M.J. Multiple di-leucines in the ATP7A copper transporter are required for retrograde trafficking to the trans-Golgi network. Metallomics 2016, 8, 993–1001. [Google Scholar] [CrossRef] [Green Version]
- Braiterman, L.; Nyasae, L.; Leves, F.; Hubbard, A.L. Critical roles for the COOH terminus of the Cu-ATPase ATP7B in protein stability, trans-Golgi network retention, copper sensing, and retrograde trafficking. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G69–G81. [Google Scholar] [CrossRef] [Green Version]
- Petris, M.J.; Voskoboinik, I.; Cater, M.; Smith, K.; Kim, B.E.; Llanos, R.M.; Strausak, D.; Camakaris, J.; Mercer, J.F. Copper-regulated trafficking of the Menkes disease copper ATPase is associated with formation of a phosphorylated catalytic intermediate. J. Biol. Chem. 2002, 277, 46736–46742. [Google Scholar] [CrossRef] [Green Version]
- Veldhuis, N.A.; Valova, V.A.; Gaeth, A.P.; Palstra, N.; Hannan, K.M.; Michell, B.J.; Kelly, L.E.; Jennings, I.; Kemp, B.E.; Pearson, R.B.; et al. Phosphorylation regulates copper-responsive trafficking of the Menkes copper transporting P-type ATPase. Int. J. Biochem. Cell Biol. 2009, 41, 2403–2412. [Google Scholar] [CrossRef]
- Holloway, Z.G.; Velayos-Baeza, A.; Howell, G.J.; Levecque, C.; Ponnambalam, S.; Sztul, E.; Monaco, A.P. Trafficking of the Menkes copper transporter ATP7A is regulated by clathrin-, AP-2-, AP-1-, and Rab22-dependent steps. Mol. Biol. Cell. 2013, 24, 1735–1748, S1–8. [Google Scholar] [CrossRef] [PubMed]
- Skjorringe, T.; Amstrup Pedersen, P.; Salling Thorborg, S.; Nissen, P.; Gourdon, P.; Birk Moller, L. Characterization of ATP7A missense mutants suggests a correlation between intracellular trafficking and severity of Menkes disease. Sci. Rep. 2017, 7, 757. [Google Scholar] [CrossRef]
- Blockhuys, S. and P. Wittung-Stafshede, Copper chaperone Atox1 plays role in breast cancer cell migration. Biochem. Biophys. Res. Commun. 2017, 483, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, A.; Dev, K.; Tanwar, R.S.; Selwal, K.K.; Tyagi, P.K. Copper mediated neurological disorder: Visions into amyotrophic lateral sclerosis, Alzheimer and Menkes disease. J. Trace Elem. Med. Biol. 2015, 29, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Telianidis, J.; Hung, Y.H.; Materia, S.; Fontaine, S.L. Role of the P-Type ATPases, ATP7A and ATP7B in brain copper homeostasis. Front. Aging Neurosci. 2013, 5, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ash, D.; Sudhahar, V.; Youn, S.W.; Okur, M.N.; Das, A.; O’Bryan, J.P.; McMenamin, M.; Hou, Y.; Kaplan, J.H.; Fukai, T.; et al. The P-type ATPase transporter ATP7A promotes angiogenesis by limiting autophagic degradation of VEGFR2. Nat. Commun. 2021, 12, 3091. [Google Scholar] [CrossRef] [PubMed]
- Arnesano, F.; Natile, G. Interference between copper transport systems and platinum drugs. Semin. Cancer Biol. 2021, 76, 173–188. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Wang, Y. Golgi Metal Ion Homeostasis in Human Health and Diseases. Cells 2022, 11, 289. https://doi.org/10.3390/cells11020289
Li J, Wang Y. Golgi Metal Ion Homeostasis in Human Health and Diseases. Cells. 2022; 11(2):289. https://doi.org/10.3390/cells11020289
Chicago/Turabian StyleLi, Jie, and Yanzhuang Wang. 2022. "Golgi Metal Ion Homeostasis in Human Health and Diseases" Cells 11, no. 2: 289. https://doi.org/10.3390/cells11020289
APA StyleLi, J., & Wang, Y. (2022). Golgi Metal Ion Homeostasis in Human Health and Diseases. Cells, 11(2), 289. https://doi.org/10.3390/cells11020289