

Exosomal Plasma Gelsolin Is an Immunosuppressive Mediator in the Ovarian Tumor Microenvironment and a Determinant of Chemoresistance

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Ovarian Cancer (OVCA) and Extracellular Vesicles (EVs)
1.2. Structure, Function, and Regulation of Gelsolin (GSN)
2. pGSN and Chemoresistance
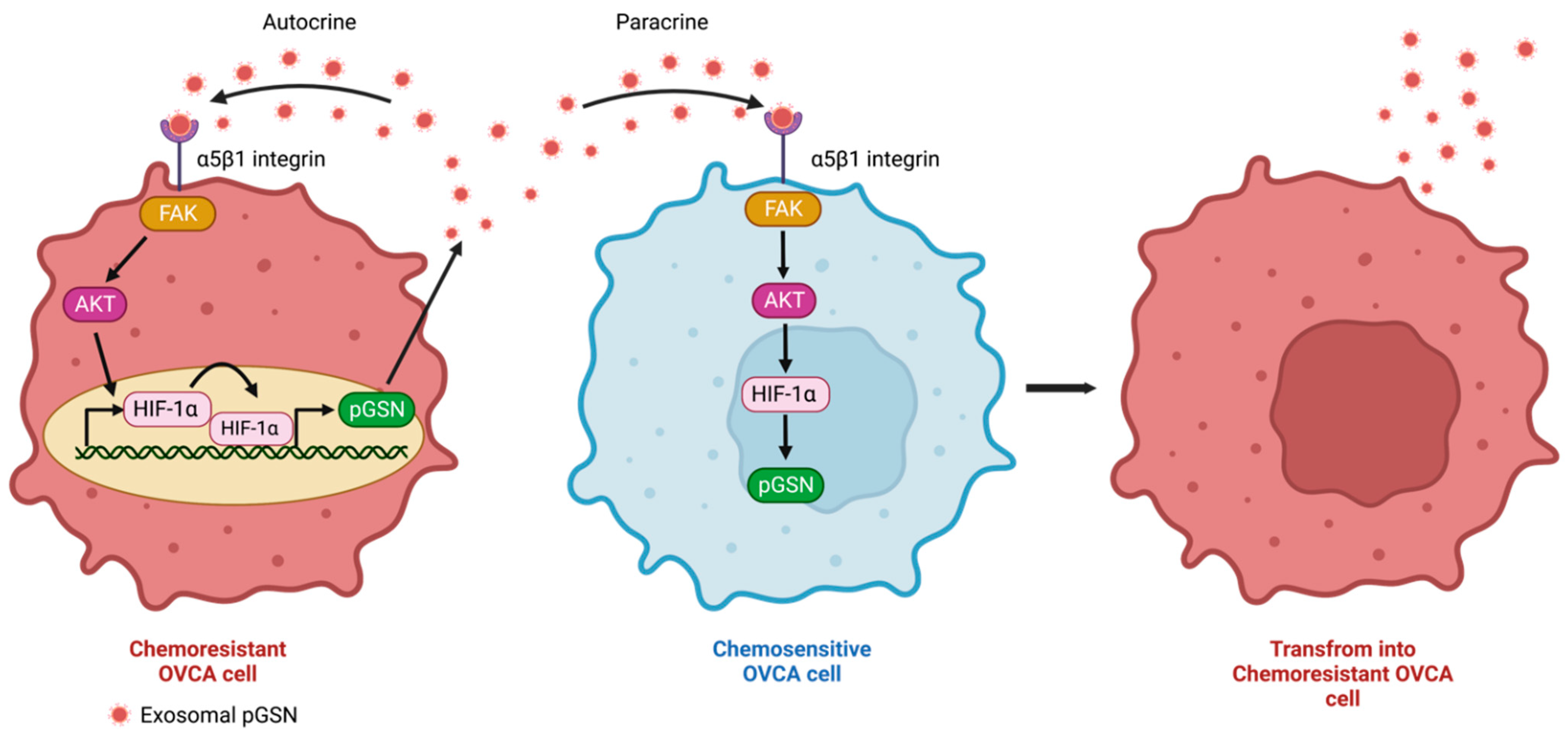
2.1. Autocrine and Paracrine Mechanisms of pGSN-Mediated Chemoresistance
2.2. Apoptosis Regulation by pGSN
3. pGSN and Immune Dysfunction
3.1. T Cell Dysfunction and Increased Glutathione (GSH) Production in OVCA through Decreased Interferon (IFN)γ Production
3.2. pGSN and Macrophage Dysfunction
3.3. pGSN and Other Immune Cells
4. Clinical Significance of pGSN
4.1. Early Diagnosis
4.2. Residual Disease Prediction
4.3. Prognostic Marker
5. Summary and Future Research Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prat, J. Ovarian Carcinomas: Five Distinct Diseases with Different Origins, Genetic Alterations, and Clinicopathological Features. Virchows Arch. 2012, 460, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Surveillance, Epidemiology, and End Results (SEER) Program. SEER*Stat Database: Incidence—SEER Research Data, 8 Registries, Nov 2021 Sub (1975–2019)—Linked to County Attributes—Time Dependent (1990–2019) Income/Rurality, 1969–2020 Counties, National Cancer Institute, DCCPS, Surveillance Research Program, Released April 2022, Based on the November 2021 Submission. Available online: https://seer.cancer.gov/statfacts/html/cervix.html (accessed on 26 August 2022).
- Surveillance, Epidemiology, and End Results (SEER) Program. SEER*Stat Database: Incidence—SEER Research Data, 8 Registries, Nov 2021 Sub (1975–2019)—Linked to County Attributes—Time Dependent (1990–2019) Income/Rurality, 1969–2020 Counties, National Cancer Institute, DCCPS, Surveillance Research Program, Released April 2022, Based on the November 2021 Submission. Available online: https://seer.cancer.gov/statfacts/html/corp.html (accessed on 26 August 2022).
- Surveillance, Epidemiology, and End Results (SEER) Program. SEER*Stat Database: Incidence—SEER Research Data, 8 Registries, Nov 2021 Sub (1975–2019)—Linked to County Attributes—Time Dependent (1990–2019) Income/Rurality, 1969–2020 Counties, National Cancer Institute, DCCPS, Surveillance Research Program, Released April 2022, Based on the November 2021 Submission. Available online: https://seer.cancer.gov/statfacts/html/ovary.html (accessed on 26 August 2022).
- Goff, B.A.; Mandel, L.S.; Melancon, C.H.; Muntz, H.G. Frequency of Symptoms of Ovarian Cancer in Women Presenting to Primary Care Clinics. JAMA 2004, 291, 2705–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menon, U.; Karpinskyj, C.; Gentry-Maharaj, A. Ovarian Cancer Prevention and Screening. Obstet. Gynecol. 2018, 131, 909–927. [Google Scholar] [CrossRef] [Green Version]
- Hennessy, B.T.; Coleman, R.L.; Markman, M. Ovarian Cancer. Lancet 2009, 374, 1371–1382. [Google Scholar] [CrossRef]
- Kuroki, L.; Guntupalli, S.R. Treatment of Epithelial Ovarian Cancer. BMJ 2020, 371, m3773. [Google Scholar] [CrossRef]
- Guo, C.; Song, C.; Zhang, J.; Gao, Y.; Qi, Y.; Zhao, Z.; Yuan, C. Revisiting Chemoresistance in Ovarian Cancer: Mechanism, Biomarkers, and Precision Medicine. Genes Dis. 2022, 9, 668–681. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug Resistance in Cancer: Role of ATP–Dependent Transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.I.; Williams, R.T.; Henderson, M.J.; Norris, M.D.; Haber, M. ABC Transporters as Mediators of Drug Resistance and Contributors to Cancer Cell Biology. Drug Resist. Updat. 2016, 26, 1–9. [Google Scholar] [CrossRef]
- Barbato, L.; Bocchetti, M.; Di Biase, A.; Regad, T. Cancer Stem Cells and Targeting Strategies. Cells 2019, 8, 926. [Google Scholar] [CrossRef]
- Tam, C.; Rao, S.; Waye, M.M.Y.; Ng, T.B.; Wang, C.C. Autophagy Signals Orchestrate Chemoresistance of Gynecological Cancers. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188525. [Google Scholar] [CrossRef] [PubMed]
- Akman, M.; Belisario, D.C.; Salaroglio, I.C.; Kopecka, J.; Donadelli, M.; De Smaele, E.; Riganti, C. Hypoxia, Endoplasmic Reticulum Stress and Chemoresistance: Dangerous Liaisons. J. Exp. Clin. Cancer Res. 2021, 40, 28. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Sharma, S.; Obermair, A.; Salomon, C. Extracellular Vesicle-Associated MiRNAs and Chemoresistance: A Systematic Review. Cancers 2021, 13, 4608. [Google Scholar] [CrossRef]
- Zhong, Y.; Li, H.; Li, P.; Chen, Y.; Zhang, M.; Yuan, Z.; Zhang, Y.; Xu, Z.; Luo, G.; Fang, Y.; et al. Exosomes: A New Pathway for Cancer Drug Resistance. Front. Oncol. 2021, 11, 743556. [Google Scholar] [CrossRef]
- Asare-Werehene, M.; Communal, L.; Carmona, E.; Han, Y.; Song, Y.S.; Burger, D.; Mes-Masson, A.-M.; Tsang, B.K. Plasma Gelsolin Inhibits CD8+ T-Cell Function and Regulates Glutathione Production to Confer Chemoresistance in Ovarian Cancer. Cancer Res. 2020, 80, 3959–3971. [Google Scholar] [CrossRef] [PubMed]
- Di Vizio, D.; Morello, M.; Dudley, A.C.; Schow, P.W.; Adam, R.M.; Morley, S.; Mulholland, D.; Rotinen, M.; Hager, M.H.; Insabato, L.; et al. Large Oncosomes in Human Prostate Cancer Tissues and in the Circulation of Mice with Metastatic Disease. Am. J. Pathol. 2012, 181, 1573–1584. [Google Scholar] [CrossRef]
- Margolis, L.; Sadovsky, Y. The Biology of Extracellular Vesicles: The Known Unknowns. PLoS Biol. 2019, 17, e3000363. [Google Scholar] [CrossRef]
- Raju, D.; Bathini, S.; Badilescu, S.; Ghosh, A.; Packirisamy, M. Microfluidic Platforms for the Isolation and Detection of Exosomes: A Brief Review. Micromachines 2022, 13, 730. [Google Scholar] [CrossRef]
- Milane, L.; Singh, A.; Mattheolabakis, G.; Suresh, M.; Amiji, M.M. Exosome Mediated Communication within the Tumor Microenvironment. J. Control Release 2015, 219, 278–294. [Google Scholar] [CrossRef]
- Safaei, R.; Larson, B.J.; Cheng, T.C.; Gibson, M.A.; Otani, S.; Naerdemann, W.; Howell, S.B. Abnormal Lysosomal Trafficking and Enhanced Exosomal Export of Cisplatin in Drug-Resistant Human Ovarian Carcinoma Cells. Mol. Cancer Ther. 2005, 4, 1595–1604. [Google Scholar] [CrossRef]
- Qian, L.; Yang, X.; Li, S.; Zhao, H.; Gao, Y.; Zhao, S.; Lv, X.; Zhang, X.; Li, L.; Zhai, L.; et al. Reduced O-GlcNAcylation of SNAP-23 Promotes Cisplatin Resistance by Inducing Exosome Secretion in Ovarian Cancer. Cell Death Discov. 2021, 7, 112. [Google Scholar] [CrossRef] [PubMed]
- Samuel, P.; Mulcahy, L.A.; Furlong, F.; McCarthy, H.O.; Brooks, S.A.; Fabbri, M.; Pink, R.C.; Carter, D.R.F. Cisplatin Induces the Release of Extracellular Vesicles from Ovarian Cancer Cells That Can Induce Invasiveness and Drug Resistance in Bystander Cells. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20170065. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Gao, X.; Dong, Y.; Zhao, Y.; Chen, X.; Yang, G.; Liu, L. Exosome-Mediated Cell-Cell Communication in Tumor Progression. Am. J. Cancer Res. 2018, 8, 1661–1673. [Google Scholar] [PubMed]
- Xie, S.; Zhang, Q.; Jiang, L. Current Knowledge on Exosome Biogenesis, Cargo-Sorting Mechanism and Therapeutic Implications. Membranes 2022, 12, 498. [Google Scholar] [CrossRef] [PubMed]
- Weiner-Gorzel, K.; Dempsey, E.; Milewska, M.; McGoldrick, A.; Toh, V.; Walsh, A.; Lindsay, S.; Gubbins, L.; Cannon, A.; Sharpe, D.; et al. Overexpression of the MicroRNA MiR-433 Promotes Resistance to Paclitaxel through the Induction of Cellular Senescence in Ovarian Cancer Cells. Cancer Med. 2015, 4, 745–758. [Google Scholar] [CrossRef]
- Pink, R.C.; Samuel, P.; Massa, D.; Caley, D.P.; Brooks, S.A.; Carter, D.R.F. The Passenger Strand, MiR-21-3p, Plays a Role in Mediating Cisplatin Resistance in Ovarian Cancer Cells. Gynecol. Oncol. 2015, 137, 143–151. [Google Scholar] [CrossRef]
- Kanlikilicer, P.; Bayraktar, R.; Denizli, M.; Rashed, M.H.; Ivan, C.; Aslan, B.; Mitra, R.; Karagoz, K.; Bayraktar, E.; Zhang, X.; et al. Exosomal MiRNA Confers Chemo Resistance via Targeting Cav1/p-Gp/M2-Type Macrophage Axis in Ovarian Cancer. EBioMedicine 2018, 38, 100–112. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Bao, W.; Liu, Y.; Wang, S.; Xu, S.; Li, X.; Li, Y.; Wu, S. MiR-98-5p Contributes to Cisplatin Resistance in Epithelial Ovarian Cancer by Suppressing MiR-152 Biogenesis via Targeting Dicer1. Cell Death Dis. 2018, 9, 447. [Google Scholar] [CrossRef]
- Zhu, X.; Shen, H.; Yin, X.; Yang, M.; Wei, H.; Chen, Q.; Feng, F.; Liu, Y.; Xu, W.; Li, Y. Macrophages Derived Exosomes Deliver MiR-223 to Epithelial Ovarian Cancer Cells to Elicit a Chemoresistant Phenotype. J. Exp. Clin. Cancer Res. 2019, 38, 81. [Google Scholar] [CrossRef]
- Alharbi, M.; Sharma, S.; Guanzon, D.; Lai, A.; Zuñiga, F.; Shiddiky, M.J.A.; Yamauchi, Y.; Salas-Burgos, A.; He, Y.; Pejovic, T.; et al. MiRNa Signature in Small Extracellular Vesicles and Their Association with Platinum Resistance and Cancer Recurrence in Ovarian Cancer. Nanomedicine 2020, 28, 102207. [Google Scholar] [CrossRef]
- Zhuang, L.; Zhang, B.; Liu, X.; Lin, L.; Wang, L.; Hong, Z.; Chen, J. Exosomal MiR-21-5p Derived from Cisplatin-Resistant SKOV3 Ovarian Cancer Cells Promotes Glycolysis and Inhibits Chemosensitivity of Its Progenitor SKOV3 Cells by Targeting PDHA1. Cell Biol. Int. 2021, 45, 2140–2149. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Lin, L.; Liu, Q.; Gao, W.; Chen, L.; Sha, C.; Chen, Q.; Xu, W.; Li, Y.; Zhu, X. Exosomal Transfer of MiR-429 Confers Chemoresistance in Epithelial Ovarian Cancer. Am. J. Cancer Res. 2021, 11, 2124–2141. [Google Scholar] [PubMed]
- Dorayappan, K.D.P.; Wanner, R.; Wallbillich, J.J.; Saini, U.; Zingarelli, R.; Suarez, A.A.; Cohn, D.E.; Selvendiran, K. Hypoxia-Induced Exosomes Contribute to a More Aggressive and Chemoresistant Ovarian Cancer Phenotype: A Novel Mechanism Linking STAT3/Rab Proteins. Oncogene 2018, 37, 3806–3821. [Google Scholar] [CrossRef] [PubMed]
- El-Arabey, A.A.; Denizli, M.; Kanlikilicer, P.; Bayraktar, R.; Ivan, C.; Rashed, M.; Kabil, N.; Ozpolat, B.; Calin, G.A.; Salama, S.A.; et al. GATA3 as a Master Regulator for Interactions of Tumor-Associated Macrophages with High-Grade Serous Ovarian Carcinoma. Cell Signal. 2020, 68, 109539. [Google Scholar] [CrossRef] [PubMed]
- Piktel, E.; Levental, I.; Durnaś, B.; Janmey, P.A.; Bucki, R. Plasma Gelsolin: Indicator of Inflammation and Its Potential as a Diagnostic Tool and Therapeutic Target. Int. J. Mol. Sci. 2018, 19, 2516. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.L.; Kwiatkowski, D.J.; Mole, J.E.; Cole, F.S. Structure and Biosynthesis of Cytoplasmic and Secreted Variants of Gelsolin. J. Biol. Chem. 1984, 259, 5271–5276. [Google Scholar] [CrossRef]
- Feldt, J.; Schicht, M.; Garreis, F.; Welss, J.; Schneider, U.W.; Paulsen, F. Structure, Regulation and Related Diseases of the Actin-Binding Protein Gelsolin. Expert Rev. Mol. Med. 2019, 20, e7. [Google Scholar] [CrossRef]
- Nag, S.; Larsson, M.; Robinson, R.C.; Burtnick, L.D. Gelsolin: The Tail of a Molecular Gymnast. Cytoskeleton 2013, 70, 360–384. [Google Scholar] [CrossRef]
- Garg, R.; Peddada, N.; Sagar, A.; Nihalani, D. Ashish Visual Insight into How Low PH Alone Can Induce Actin-Severing Ability in Gelsolin under Calcium-Free Conditions. J. Biol. Chem. 2011, 286, 20387–20397. [Google Scholar] [CrossRef] [Green Version]
- Janmey, P.A.; Stossel, T.P. Modulation of Gelsolin Function by Phosphatidylinositol 4,5-Bisphosphate. Nature 1987, 325, 362–364. [Google Scholar] [CrossRef]
- Vouyiouklis, D.A.; Brophy, P.J. A Novel Gelsolin Isoform Expressed by Oligodendrocytes in the Central Nervous System. J. Neurochem. 1997, 69, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Silacci, P.; Mazzolai, L.; Gauci, C.; Stergiopulos, N.; Yin, H.L.; Hayoz, D. Gelsolin Superfamily Proteins: Key Regulators of Cellular Functions. Cell Mol. Life Sci. 2004, 61, 2614–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiatkowski, D.J.; Stossel, T.P.; Orkin, S.H.; Mole, J.E.; Colten, H.R.; Yin, H.L. Plasma and Cytoplasmic Gelsolins Are Encoded by a Single Gene and Contain a Duplicated Actin-Binding Domain. Nature 1986, 323, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Allen, P.G. Functional Consequences of Disulfide Bond Formation in Gelsolin. FEBS Lett. 1997, 401, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Wen, D.; Corina, K.; Chow, E.P.; Miller, S.; Janmey, P.A.; Pepinsky, R.B. The Plasma and Cytoplasmic Forms of Human Gelsolin Differ in Disulfide Structure. Biochemistry 1996, 35, 9700–9709. [Google Scholar] [CrossRef]
- Smith, D.B.; Janmey, P.A.; Herbert, T.J.; Lind, S.E. Quantitative Measurement of Plasma Gelsolin and Its Incorporation into Fibrin Clots. J. Lab. Clin. Med. 1987, 110, 189–195. [Google Scholar]
- Kwiatkowski, D.J.; Mehl, R.; Izumo, S.; Nadal-Ginard, B.; Yin, H.L. Muscle Is the Major Source of Plasma Gelsolin. J. Biol. Chem. 1988, 263, 8239–8243. [Google Scholar] [CrossRef]
- Chen, C.-C.; Chiou, S.-H.; Yang, C.-L.; Chow, K.-C.; Lin, T.-Y.; Chang, H.-W.; You, W.-C.; Huang, H.-W.; Chen, C.-M.; Chen, N.-C.; et al. Secreted Gelsolin Desensitizes and Induces Apoptosis of Infiltrated Lymphocytes in Prostate Cancer. Oncotarget 2017, 8, 77152–77167. [Google Scholar] [CrossRef] [Green Version]
- Asare-Werehene, M.; Nakka, K.; Reunov, A.; Chiu, C.-T.; Lee, W.-T.; Abedini, M.R.; Wang, P.-W.; Shieh, D.-B.; Dilworth, F.J.; Carmona, E.; et al. The Exosome-Mediated Autocrine and Paracrine Actions of Plasma Gelsolin in Ovarian Cancer Chemoresistance. Oncogene 2020, 39, 1600–1616. [Google Scholar] [CrossRef] [Green Version]
- Asare-Werehene, M.; Tsuyoshi, H.; Zhang, H.; Salehi, R.; Chang, C.-Y.; Carmona, E.; Librach, C.L.; Mes-Masson, A.-M.; Chang, C.-C.; Burger, D.; et al. Plasma Gelsolin Confers Chemoresistance in Ovarian Cancer by Resetting the Relative Abundance and Function of Macrophage Subtypes. Cancers 2022, 14, 1039. [Google Scholar] [CrossRef]
- Giampazolias, E.; Schulz, O.; Lim, K.H.J.; Rogers, N.C.; Chakravarty, P.; Srinivasan, N.; Gordon, O.; Cardoso, A.; Buck, M.D.; Poirier, E.Z.; et al. Secreted Gelsolin Inhibits DNGR-1-Dependent Cross-Presentation and Cancer Immunity. Cell 2021, 184, 4016–4031. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.K.; Sawada, K.; Tiwari, P.; Mui, K.; Gwin, K.; Lengyel, E. Ligand-Independent Activation of c-Met by Fibronectin and α(5)β(1)-Integrin Regulates Ovarian Cancer Invasion and Metastasis. Oncogene 2011, 30, 1566–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.; Cao, X.; Luo, Y.; Zhang, G.; Zhang, D. A Positive Feedback Loop of LncRNA DSCR8/MiR-98-5p/STAT3/HIF-1α Plays a Role in the Progression of Ovarian Cancer. Front. Oncol. 2020, 10, 1713. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Deng, X.; Ma, X.; Zang, N.; Li, H.; Li, G.; Li, D.; Li, C.; Huang, W.; He, M. Cellular Transcriptomics: Gelsolin Negatively Regulates the Expression of Apoptosis-Associated Genes and Inhibits Apoptosis in Hepatocarcinoma Cells. Int. J. Clin. Exp. Pathol. 2015, 8, 13871–13885. [Google Scholar]
- Koya, R.C.; Fujita, H.; Shimizu, S.; Ohtsu, M.; Takimoto, M.; Tsujimoto, Y.; Kuzumaki, N. Gelsolin Inhibits Apoptosis by Blocking Mitochondrial Membrane Potential Loss and Cytochrome c Release. J. Biol. Chem. 2000, 275, 15343–15349. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.J.; Azuma, T.; Tang, J.X.; Hartwig, J.H.; Muszynski, M.; Wu, Q.; Libby, P.; Kwiatkowski, D.J. Caspase-3-Induced Gelsolin Fragmentation Contributes to Actin Cytoskeletal Collapse, Nucleolysis, and Apoptosis of Vascular Smooth Muscle Cells Exposed to Proinflammatory Cytokines. Eur. J. Cell Biol. 1998, 77, 294–302. [Google Scholar] [CrossRef]
- Deng, R.; Hao, J.; Han, W.; Ni, Y.; Huang, X.; Hu, Q. Gelsolin Regulates Proliferation, Apoptosis, Migration and Invasion in Human Oral Carcinoma Cells. Oncol. Lett. 2015, 9, 2129–2134. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Ye, Z.; Wen, J.; Ma, L.; He, Y.; Lian, G.; Wang, Z.; Wei, L.; Wu, D.; Jiang, B. Gelsolin, but Not Its Cleavage, Is Required for TNF-Induced ROS Generation and Apoptosis in MCF-7 Cells. Biochem. Biophys. Res. Commun. 2009, 385, 284–289. [Google Scholar] [CrossRef]
- Kothakota, S.; Azuma, T.; Reinhard, C.; Klippel, A.; Tang, J.; Chu, K.; McGarry, T.J.; Kirschner, M.W.; Koths, K.; Kwiatkowski, D.J.; et al. Caspase-3-Generated Fragment of Gelsolin: Effector of Morphological Change in Apoptosis. Science 1997, 278, 294–298. [Google Scholar] [CrossRef]
- Sakurai, N.; Utsumi, T. Posttranslational N-Myristoylation Is Required for the Anti-Apoptotic Activity of Human TGelsolin, the C-Terminal Caspase Cleavage Product of Human Gelsolin. J. Biol. Chem. 2006, 281, 14288–14295. [Google Scholar] [CrossRef] [Green Version]
- Chhabra, D.; Nosworthy, N.J.; dos Remedios, C.G. The N-Terminal Fragment of Gelsolin Inhibits the Interaction of DNase I with Isolated Actin, but Not with the Cofilin-Actin Complex. Proteomics 2005, 5, 3131–3136. [Google Scholar] [CrossRef] [PubMed]
- Binju, M.; Amaya-Padilla, M.A.; Wan, G.; Gunosewoyo, H.; Suryo Rahmanto, Y.; Yu, Y. Therapeutic Inducers of Apoptosis in Ovarian Cancer. Cancers 2019, 11, 1786. [Google Scholar] [CrossRef] [PubMed]
- Abedini, M.R.; Muller, E.J.; Bergeron, R.; Gray, D.A.; Tsang, B.K. Akt Promotes Chemoresistance in Human Ovarian Cancer Cells by Modulating Cisplatin-Induced, P53-Dependent Ubiquitination of FLICE-like Inhibitory Protein. Oncogene 2010, 29, 11–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abedini, M.R.; Muller, E.J.; Brun, J.; Bergeron, R.; Gray, D.A.; Tsang, B.K. Cisplatin Induces P53-Dependent FLICE-like Inhibitory Protein Ubiquitination in Ovarian Cancer Cells. Cancer Res. 2008, 68, 4511–4517. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, L.; Espona-Fiedler, M.; Longley, D.B. FLIP as a Therapeutic Target in Cancer. FEBS J. 2018, 285, 4104–4123. [Google Scholar] [CrossRef] [Green Version]
- Abedini, M.R.; Qiu, Q.; Yan, X.; Tsang, B.K. Possible Role of FLICE-like Inhibitory Protein (FLIP) in Chemoresistant Ovarian Cancer Cells In Vitro. Oncogene 2004, 23, 6997–7004. [Google Scholar] [CrossRef] [Green Version]
- Abedini, M.R.; Wang, P.-W.; Huang, Y.-F.; Cao, M.; Chou, C.-Y.; Shieh, D.-B.; Tsang, B.K. Cell Fate Regulation by Gelsolin in Human Gynecologic Cancers. Proc. Natl. Acad. Sci. USA 2014, 111, 14442–14447. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhang, T.; Ye, J.; Li, H.; Huang, J.; Li, X.; Wu, B.; Huang, X.; Hou, J. Tumor-Infiltrating Lymphocytes Predict Response to Chemotherapy in Patients with Advance Non-Small Cell Lung Cancer. Cancer Immunol. Immunother. 2012, 61, 1849–1856. [Google Scholar] [CrossRef]
- Zhang, L.; Conejo-Garcia, J.R.; Katsaros, D.; Gimotty, P.A.; Massobrio, M.; Regnani, G.; Makrigiannakis, A.; Gray, H.; Schlienger, K.; Liebman, M.N.; et al. Intratumoral T Cells, Recurrence, and Survival in Epithelial Ovarian Cancer. N. Engl. J. Med. 2003, 348, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Rapoport, B.L.; Nayler, S.; Mlecnik, B.; Smit, T.; Heyman, L.; Bouquet, I.; Martel, M.; Galon, J.; Benn, C.-A.; Anderson, R. Tumor-Infiltrating Lymphocytes (TILs) in Early Breast Cancer Patients: High CD3+, CD8+, and Immunoscore Are Associated with a Pathological Complete Response. Cancers 2022, 14, 2525. [Google Scholar] [CrossRef]
- Zhou, J.; Li, X.; Wu, X.; Zhang, T.; Zhu, Q.; Wang, X.; Wang, H.; Wang, K.; Lin, Y.; Wang, X. Exosomes Released from Tumor-Associated Macrophages Transfer MiRNAs That Induce a Treg/Th17 Cell Imbalance in Epithelial Ovarian Cancer. Cancer Immunol. Res. 2018, 6, 1578–1592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenoy, G.N.; Loyall, J.; Berenson, C.S.; Kelleher, R.J., Jr.; Iyer, V.; Balu-Iyer, S.V.; Odunsi, K.; Bankert, R.B. Sialic Acid-Dependent Inhibition of T Cells by Exosomal Ganglioside GD3 in Ovarian Tumor Microenvironments. J. Immunol. 2018, 201, 3750–3758. [Google Scholar] [CrossRef] [PubMed]
- Czystowska-Kuzmicz, M.; Sosnowska, A.; Nowis, D.; Ramji, K.; Szajnik, M.; Chlebowska-Tuz, J.; Wolinska, E.; Gaj, P.; Grazul, M.; Pilch, Z.; et al. Small Extracellular Vesicles Containing Arginase-1 Suppress T-Cell Responses and Promote Tumor Growth in Ovarian Carcinoma. Nat. Commun. 2019, 10, 3000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Kadamberi, I.P.; Mittal, S.; Tsaih, S.-W.; George, J.; Kumar, S.; Vijayan, D.K.; Geethadevi, A.; Parashar, D.; Topchyan, P.; et al. Tumor Derived Extracellular Vesicles Drive T Cell Exhaustion in Tumor Microenvironment through Sphingosine Mediated Signaling and Impacting Immunotherapy Outcomes in Ovarian Cancer. Adv. Sci. 2022, 9, e2104452. [Google Scholar] [CrossRef]
- Kelleher, R.J., Jr.; Balu-Iyer, S.; Loyall, J.; Sacca, A.J.; Shenoy, G.N.; Peng, P.; Iyer, V.; Fathallah, A.M.; Berenson, C.S.; Wallace, P.K.; et al. Extracellular Vesicles Present in Human Ovarian Tumor Microenvironments Induce a Phosphatidylserine-Dependent Arrest in the T-Cell Signaling Cascade. Cancer Immunol. Res. 2015, 3, 1269–1278. [Google Scholar] [CrossRef] [Green Version]
- Giuntoli, R.L., 2nd; Webb, T.J.; Zoso, A.; Rogers, O.; Diaz-Montes, T.P.; Bristow, R.E.; Oelke, M. Ovarian Cancer-Associated Ascites Demonstrates Altered Immune Environment: Implications for Antitumor Immunity. Anticancer Res. 2009, 29, 2875–2884. [Google Scholar]
- Cândido, E.B.; Silva, L.M.; Carvalho, A.T.; Lamaita, R.M.; Filho, R.M.P.; Cota, B.D.C.V.; da Silva-Filho, A.L. Immune Response Evaluation through Determination of Type 1, Type 2, and Type 17 Patterns in Patients with Epithelial Ovarian Cancer. Reprod. Sci. 2013, 20, 828–837. [Google Scholar] [CrossRef]
- Lu, S.C. Glutathione Synthesis. Biochim. Biophys. Acta 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Nunes, S.C.; Serpa, J. Glutathione in Ovarian Cancer: A Double-Edged Sword. Int. J. Mol. Sci. 2018, 19, 1882. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Cui, F.; Cheng, Y.; Han, L.; Wang, J.; Sun, D.; Liu, Y.-L.; Zhou, P.-K.; Min, R. Gelsolin: Role of a Functional Protein in Mitigating Radiation Injury. Cell Biochem. Biophys. 2015, 71, 389–396. [Google Scholar] [CrossRef]
- Wang, W.; Kryczek, I.; Dostál, L.; Lin, H.; Tan, L.; Zhao, L.; Lu, F.; Wei, S.; Maj, T.; Peng, D.; et al. Effector T Cells Abrogate Stroma-Mediated Chemoresistance in Ovarian Cancer. Cell 2016, 165, 1092–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najafi, M.; Hashemi Goradel, N.; Farhood, B.; Salehi, E.; Nashtaei, M.S.; Khanlarkhani, N.; Khezri, Z.; Majidpoor, J.; Abouzaripour, M.; Habibi, M.; et al. Macrophage Polarity in Cancer: A Review. J. Cell Biochem. 2019, 120, 2756–2765. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Yu, Y.; Wang, X.; Zhang, T. Tumor-Associated Macrophages in Tumor Immunity. Front. Immunol. 2020, 11, 583084. [Google Scholar] [CrossRef]
- Diskin, C.; Pålsson-McDermott, E.M. Metabolic Modulation in Macrophage Effector Function. Front. Immunol. 2018, 9, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, X.; Wu, Q.; Wu, X.; Zhu, Q.; Wang, X.; Jiang, L.; Chen, X.; Wang, X. Epithelial Ovarian Cancer-Secreted Exosomal MiR-222-3p Induces Polarization of Tumor-Associated Macrophages. Oncotarget 2016, 7, 43076–43087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, J.; He, X.; Xu, Y.; Zhang, W.; Fu, F. MiR-200b Is Upregulated in Plasma-Derived Exosomes and Functions as an Oncogene by Promoting Macrophage M2 Polarization in Ovarian Cancer. J. Ovarian Res. 2021, 14, 74. [Google Scholar] [CrossRef]
- Chen, X.; Ying, X.; Wang, X.; Wu, X.; Zhu, Q.; Wang, X. Exosomes Derived from Hypoxic Epithelial Ovarian Cancer Deliver MicroRNA-940 to Induce Macrophage M2 Polarization. Oncol. Rep. 2017, 38, 522–528. [Google Scholar] [CrossRef] [Green Version]
- Peng, P.; Yan, Y.; Keng, S. Exosomes in the Ascites of Ovarian Cancer Patients: Origin and Effects on Anti-Tumor Immunity. Oncol. Rep. 2011, 25, 749–762. [Google Scholar]
- Labani-Motlagh, A.; Israelsson, P.; Ottander, U.; Lundin, E.; Nagaev, I.; Nagaeva, O.; Dehlin, E.; Baranov, V.; Mincheva-Nilsson, L. Differential Expression of Ligands for NKG2D and DNAM-1 Receptors by Epithelial Ovarian Cancer-Derived Exosomes and Its Influence on NK Cell Cytotoxicity. Tumour Biol. 2016, 37, 5455–5466. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, H.; Xing, X.; Wang, L.; Zhang, J.; Yan, L.; Zheng, X.; Zhang, M. Gelsolin Regulates Proliferation, Apoptosis and Invasion in Natural Killer/T-Cell Lymphoma Cells. Biol. Open 2018, 7, bio027557. [Google Scholar] [CrossRef] [Green Version]
- Nossov, V.; Amneus, M.; Su, F.; Lang, J.; Janco, J.M.T.; Reddy, S.T.; Farias-Eisner, R. The Early Detection of Ovarian Cancer: From Traditional Methods to Proteomics. Can We Really Do Better than Serum CA-125? Am. J. Obstet. Gynecol. 2008, 199, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Asare-Werehene, M.; Communal, L.; Carmona, E.; Le, T.; Provencher, D.; Mes-Masson, A.-M.; Tsang, B.K. Pre-Operative Circulating Plasma Gelsolin Predicts Residual Disease and Detects Early Stage Ovarian Cancer. Sci. Rep. 2019, 9, 13924. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.-T.; Wang, P.-W.; Asare-Werehene, M.; Tsang, B.K.; Shieh, D.-B. Circulating Plasma Gelsolin: A Predictor of Favorable Clinical Outcomes in Head and Neck Cancer and Sensitive Biomarker for Early Disease Diagnosis Combined with Soluble Fas Ligand. Cancers 2020, 12, 1569. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, K.; Yin, X.; Li, H.; Li, Y.; Zhang, Q.; Wang, H.; Qiu, Y. Lower Expression of Gelsolin in Colon Cancer and Its Diagnostic Value in Colon Cancer Patients. J. Cancer 2019, 10, 1288–1296. [Google Scholar] [CrossRef] [PubMed]
- Ueland, F.R.; Desimone, C.P.; Seamon, L.G.; Miller, R.A.; Goodrich, S.; Podzielinski, I.; Sokoll, L.; Smith, A.; van Nagell, J.R., Jr.; Zhang, Z. Effectiveness of a Multivariate Index Assay in the Preoperative Assessment of Ovarian Tumors. Obstet. Gynecol. 2011, 117, 1289–1297. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Janas, Ł.; Stachowiak, G.; Stetkiewicz, T.; Wilczyński, J.R. Current Clinical Application of Serum Biomarkers to Detect Ovarian Cancer. Prz Menopauzalny 2015, 14, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Narod, S. Can Advanced-Stage Ovarian Cancer Be Cured? Nat. Rev. Clin. Oncol. 2016, 13, 255–261. [Google Scholar] [CrossRef]
- Tomao, F.; D’Incalci, M.; Biagioli, E.; Peccatori, F.A.; Colombo, N. Restoring Platinum Sensitivity in Recurrent Ovarian Cancer by Extending the Platinum-Free Interval: Myth or Reality? Cancer 2017, 123, 3450–3459. [Google Scholar] [CrossRef] [Green Version]
- Bristow, R.E.; Duska, L.R.; Lambrou, N.C.; Fishman, E.K.; O’Neill, M.J.; Trimble, E.L.; Montz, F.J. A Model for Predicting Surgical Outcome in Patients with Advanced Ovarian Carcinoma Using Computed Tomography. Cancer 2000, 89, 1532–1540. [Google Scholar] [CrossRef]
- Forstner, R.; Hricak, H.; Occhipinti, K.A.; Powell, C.B.; Frankel, S.D.; Stern, J.L. Ovarian Cancer: Staging with CT and MR Imaging. Radiology 1995, 197, 619–626. [Google Scholar] [CrossRef]
- Suidan, R.S.; Ramirez, P.T.; Sarasohn, D.M.; Teitcher, J.B.; Iyer, R.B.; Zhou, Q.; Iasonos, A.; Denesopolis, J.; Zivanovic, O.; Long Roche, K.C.; et al. A Multicenter Assessment of the Ability of Preoperative Computed Tomography Scan and CA-125 to Predict Gross Residual Disease at Primary Debulking for Advanced Epithelial Ovarian Cancer. Gynecol. Oncol. 2017, 145, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pergialiotis, V.; Karampetsou, N.; Bellos, I.; Thomakos, N.; Daskalakis, G. The Diagnostic Accuracy of Human Epididymis Factor 4 for the Prediction of Optimal Debulking in Patients with Ovarian Cancer: A Meta-Analysis of Observational Studies. Int. J. Gynecol. Cancer 2018, 28, 1471–1477. [Google Scholar] [CrossRef] [PubMed]
- Şahin, A.; Enver, N.; Erçetin, S.Y.; Cinel, Z.L.; Batman, A.Ç. The Promising Role of Gelsolin Expression to Predict Survival in Patients with Squamous Cell Carcinoma of the Larynx. Braz. J. Otorhinolaryngol. 2021, S1808-8694(21)00094-X. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Luo, X.; Lin, J.; Fu, S.; Feng, P.; Su, H.; He, X.; Liang, X.; Liu, K.; Deng, W. Gelsolin Promotes Cancer Progression by Regulating Epithelial-Mesenchymal Transition in Hepatocellular Carcinoma and Correlates with a Poor Prognosis. J. Oncol. 2020, 2020, 1980368. [Google Scholar] [CrossRef]
- Ma, X.; Sun, W.; Shen, J.; Hua, Y.; Yin, F.; Sun, M.; Cai, Z. Gelsolin Promotes Cell Growth and Invasion through the Upregulation of P-AKT and p-P38 Pathway in Osteosarcoma. Tumour Biol. 2016, 37, 7165–7174. [Google Scholar] [CrossRef] [PubMed]
- Maiorano, B.A.; Maiorano, M.F.P.; Lorusso, D.; Maiello, E. Ovarian Cancer in the Era of Immune Checkpoint Inhibitors: State of the Art and Future Perspectives. Cancers 2021, 13, 4438. [Google Scholar] [CrossRef]
- Yu, T.; Tang, B.; Sun, X. Development of Inhibitors Targeting Hypoxia-Inducible Factor 1 and 2 for Cancer Therapy. Yonsei Med. J. 2017, 58, 489–496. [Google Scholar] [CrossRef]
- Muñoz-Galván, S.; Carnero, A. Leveraging Genomics, Transcriptomics, and Epigenomics to Understand the Biology and Chemoresistance of Ovarian Cancer. Cancers 2021, 13, 4029. [Google Scholar] [CrossRef]
- Seto, E.; Yoshida, M. Erasers of Histone Acetylation: The Histone Deacetylase Enzymes. Cold Spring Harb. Perspect. Biol. 2014, 6, a018713. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Sharma, P.; Capalash, N. DNA Methyltransferase-1 Inhibitors as Epigenetic Therapy for Cancer. Curr. Cancer Drug Targets 2013, 13, 379–399. [Google Scholar] [CrossRef]
- Mielnicki, L.M.; Ying, A.M.; Head, K.L.; Asch, H.L.; Asch, B.B. Epigenetic Regulation of Gelsolin Expression in Human Breast Cancer Cells. Exp. Cell Res. 1999, 249, 161–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-C.; Chen, C.-W.; Yang, C.-L.; Tsai, I.-M.; Hou, Y.-C.; Chen, C.-J.; Shan, Y.-S. Tumor-Associated Macrophages Promote Epigenetic Silencing of Gelsolin through DNA Methyltransferase 1 in Gastric Cancer Cells. Cancer Immunol. Res. 2017, 5, 885–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asch, H.L.; Head, K.; Dong, Y.; Natoli, F.; Winston, J.S.; Connolly, J.L.; Asch, B.B. Widespread Loss of Gelsolin in Breast Cancers of Humans, Mice, and Rats. Cancer Res. 1996, 56, 4841–4845. [Google Scholar] [PubMed]
- Dosaka-Akita, H.; Hommura, F.; Fujita, H.; Kinoshita, I.; Nishi, M.; Morikawa, T.; Katoh, H.; Kawakami, Y.; Kuzumaki, N. Frequent Loss of Gelsolin Expression in Non-Small Cell Lung Cancers of Heavy Smokers. Cancer Res. 1998, 58, 322–327. [Google Scholar]
- Lee, H.K.; Driscoll, D.; Asch, H.; Asch, B.; Zhang, P.J. Downregulated Gelsolin Expression in Hyperplastic and Neoplastic Lesions of the Prostate. Prostate 1999, 40, 14–19. [Google Scholar] [CrossRef]
- Tanaka, M.; Müllauer, L.; Ogiso, Y.; Fujita, H.; Moriya, S.; Furuuchi, K.; Harabayashi, T.; Shinohara, N.; Koyanagi, T.; Kuzumaki, N. Gelsolin: A Candidate for Suppressor of Human Bladder Cancer. Cancer Res. 1995, 55, 3228–3232. [Google Scholar]
- Noske, A.; Denkert, C.; Schober, H.; Sers, C.; Zhumabayeva, B.; Weichert, W.; Dietel, M.; Wiechen, K. Loss of Gelsolin Expression in Human Ovarian Carcinomas. Eur. J. Cancer 2005, 41, 461–469. [Google Scholar] [CrossRef]
- Perets, R.; Drapkin, R. It’s Totally Tubular…Riding the New Wave of Ovarian Cancer Research. Cancer Res. 2016, 76, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Porter, R.M.; Holme, T.C.; Newman, E.L.; Hopwood, D.; Wilkinson, J.M.; Cuschieri, A. Monoclonal Antibodies to Cytoskeletal Proteins: An Immunohistochemical Investigation of Human Colon Cancer. J. Pathol. 1993, 170, 435–440. [Google Scholar] [CrossRef]
- Rutten, I.J.G.; van Dijk, D.P.J.; Kruitwagen, R.F.P.M.; Beets-Tan, R.G.H.; Olde Damink, S.W.M.; van Gorp, T. Loss of Skeletal Muscle during Neoadjuvant Chemotherapy Is Related to Decreased Survival in Ovarian Cancer Patients. J. Cachexia Sarcopenia Muscle 2016, 7, 458–466. [Google Scholar] [CrossRef] [Green Version]
- Tisdale, M.J. Cachexia in Cancer Patients. Nat. Rev. Cancer 2002, 2, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Pin, F.; Barreto, R.; Kitase, Y.; Mitra, S.; Erne, C.E.; Novinger, L.J.; Zimmers, T.A.; Couch, M.E.; Bonewald, L.F.; Bonetto, A. Growth of Ovarian Cancer Xenografts Causes Loss of Muscle and Bone Mass: A New Model for the Study of Cancer Cachexia. J. Cachexia Sarcopenia Muscle 2018, 9, 685–700. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onuma, T.; Asare-Werehene, M.; Yoshida, Y.; Tsang, B.K. Exosomal Plasma Gelsolin Is an Immunosuppressive Mediator in the Ovarian Tumor Microenvironment and a Determinant of Chemoresistance. Cells 2022, 11, 3305. https://doi.org/10.3390/cells11203305
Onuma T, Asare-Werehene M, Yoshida Y, Tsang BK. Exosomal Plasma Gelsolin Is an Immunosuppressive Mediator in the Ovarian Tumor Microenvironment and a Determinant of Chemoresistance. Cells. 2022; 11(20):3305. https://doi.org/10.3390/cells11203305
Chicago/Turabian StyleOnuma, Toshimichi, Meshach Asare-Werehene, Yoshio Yoshida, and Benjamin K. Tsang. 2022. "Exosomal Plasma Gelsolin Is an Immunosuppressive Mediator in the Ovarian Tumor Microenvironment and a Determinant of Chemoresistance" Cells 11, no. 20: 3305. https://doi.org/10.3390/cells11203305
APA StyleOnuma, T., Asare-Werehene, M., Yoshida, Y., & Tsang, B. K. (2022). Exosomal Plasma Gelsolin Is an Immunosuppressive Mediator in the Ovarian Tumor Microenvironment and a Determinant of Chemoresistance. Cells, 11(20), 3305. https://doi.org/10.3390/cells11203305