

Dynamic Balance between PTH1R-Dependent Signal Cascades Determines Its Pro- or Anti-Osteogenic Effects on MSC

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
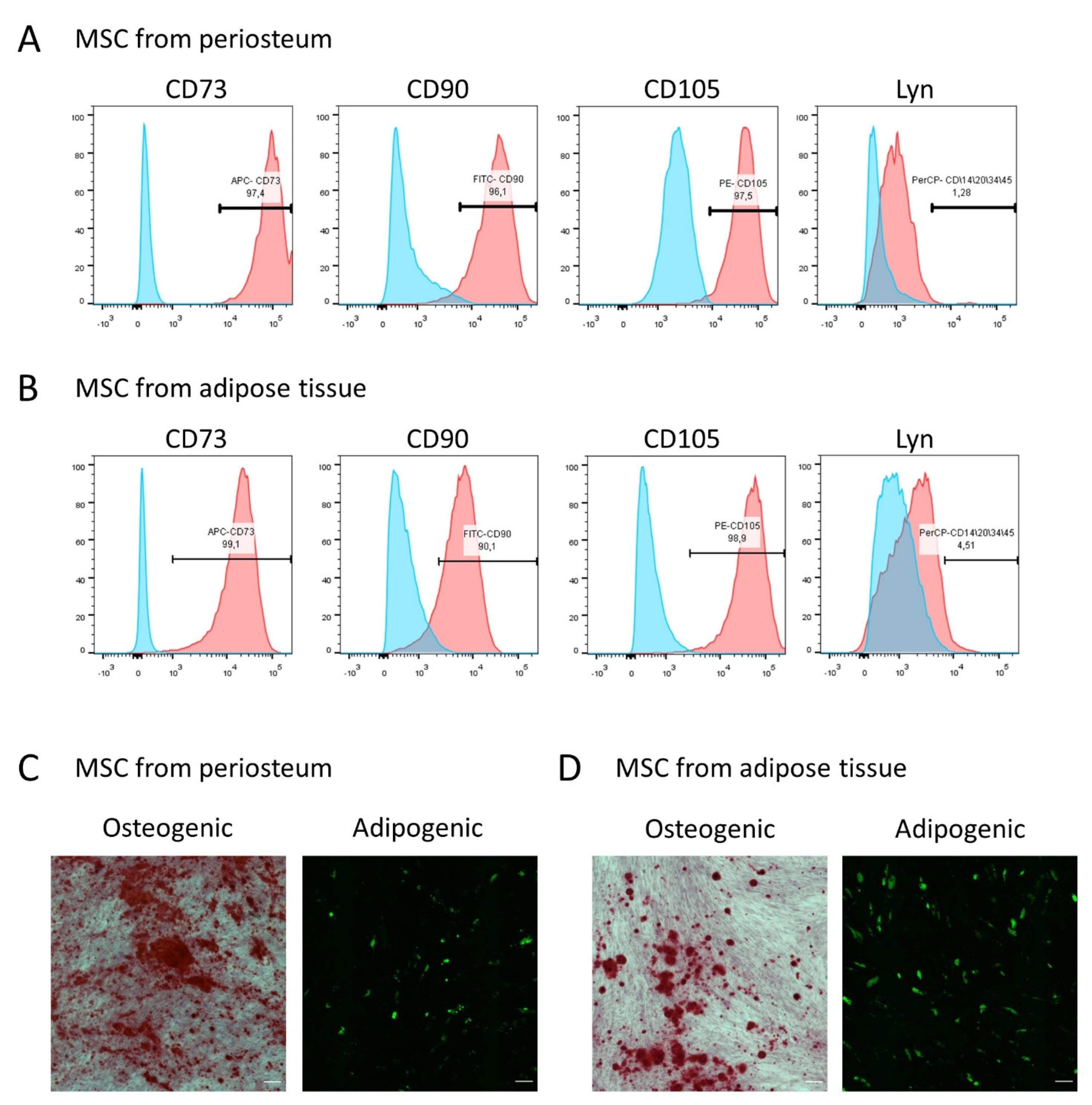
2.1. MSC Isolation and Culturing
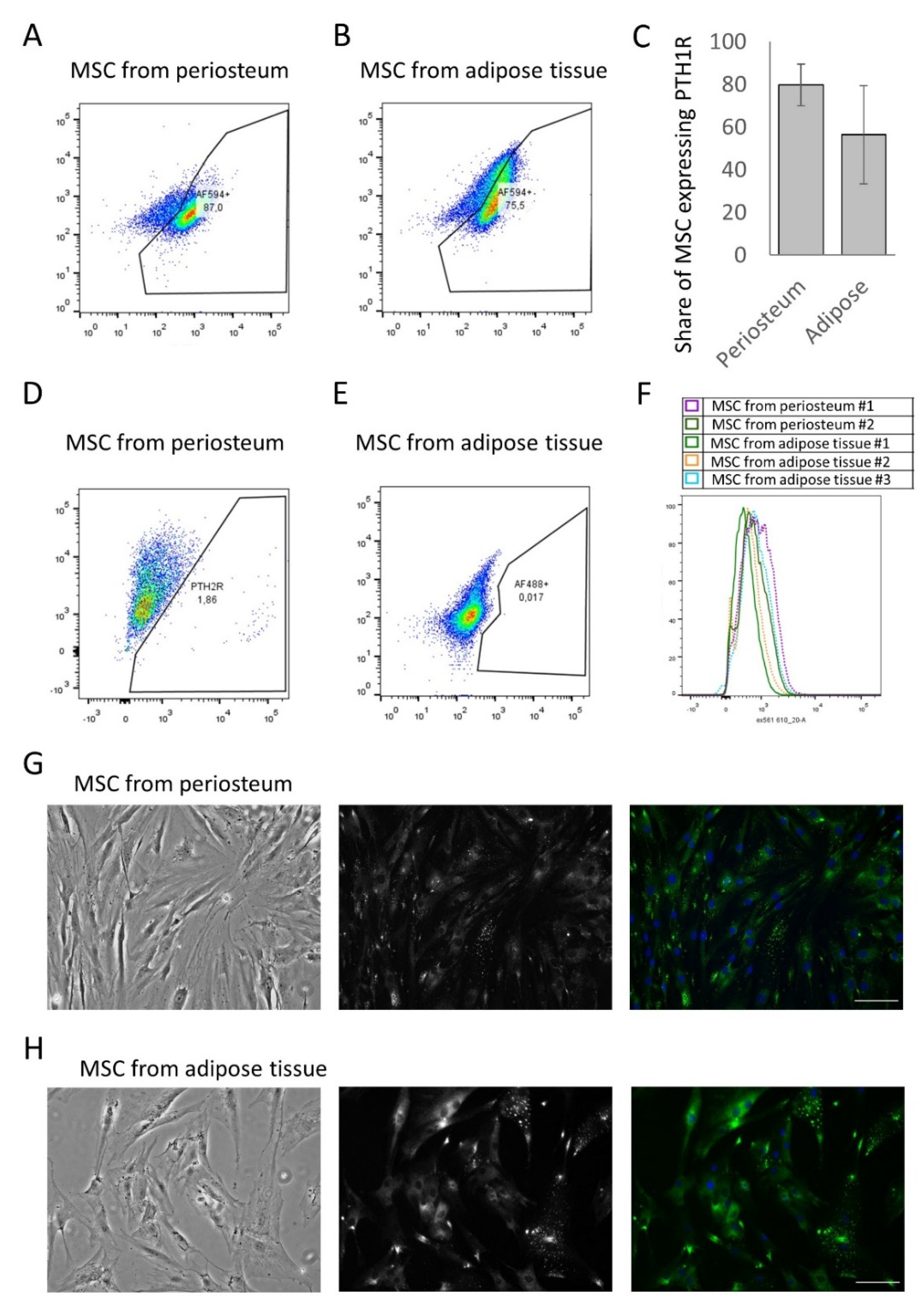
2.2. Flow Cytometry
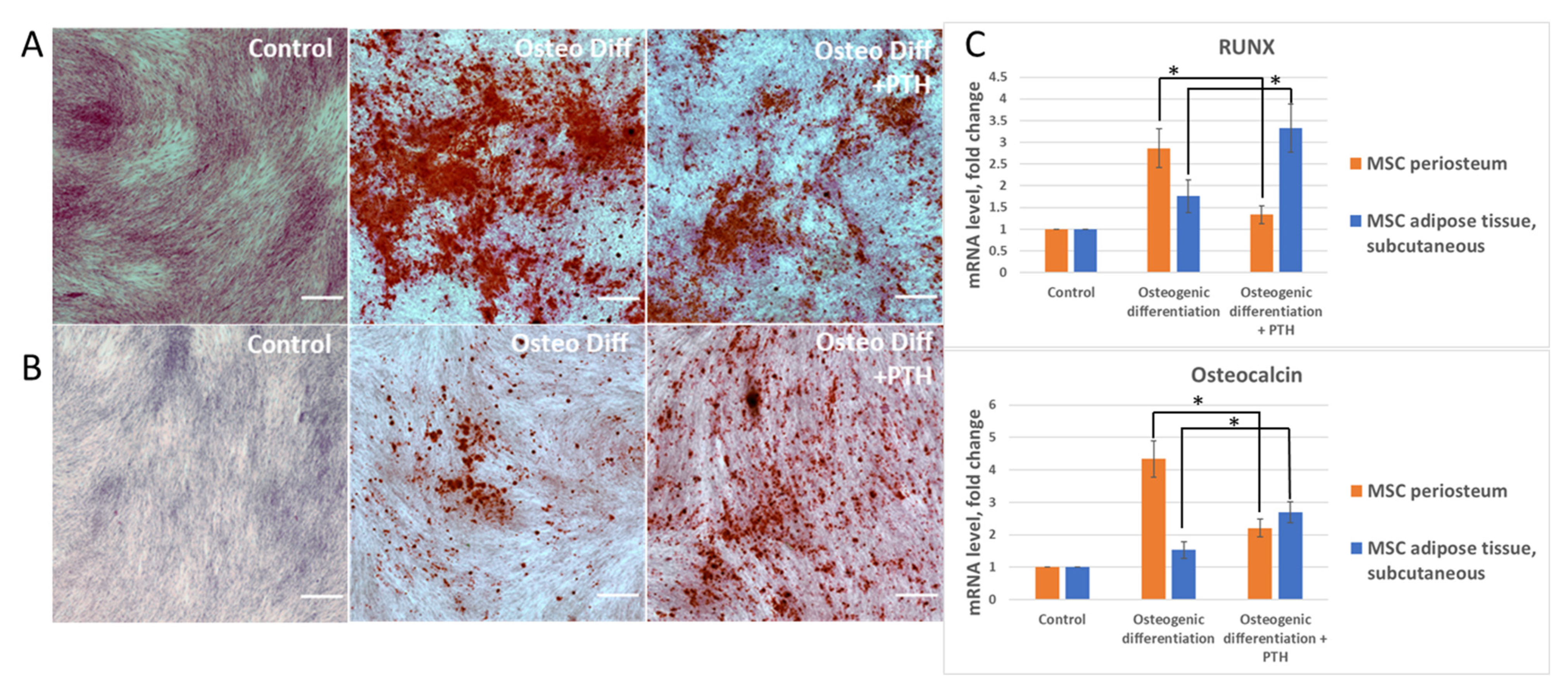
2.3. MSCs Differentiation Assays
2.4. Histologic Staining
2.5. RT-PCR
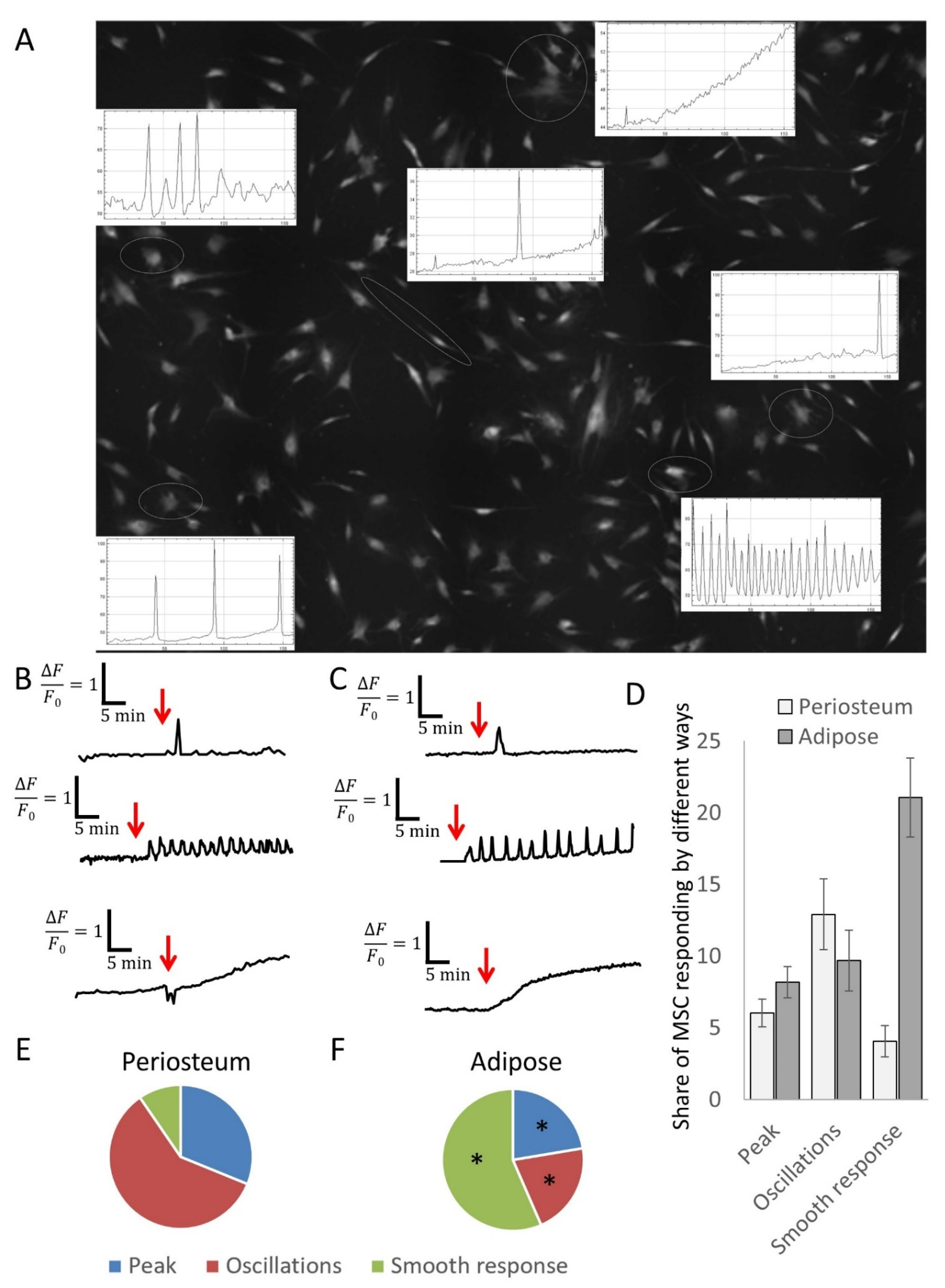
2.6. Calcium Imaging
3. Results
3.1. MSCs from Periosteum and Adipose Tissue Contain a Comparable Number of PTH1R-Positive Cells
3.2. Parathyroid Hormone (PTH) Has a Different Effect on the Differentiation of MSCs Derived from Periosteum and Adipose Tissue
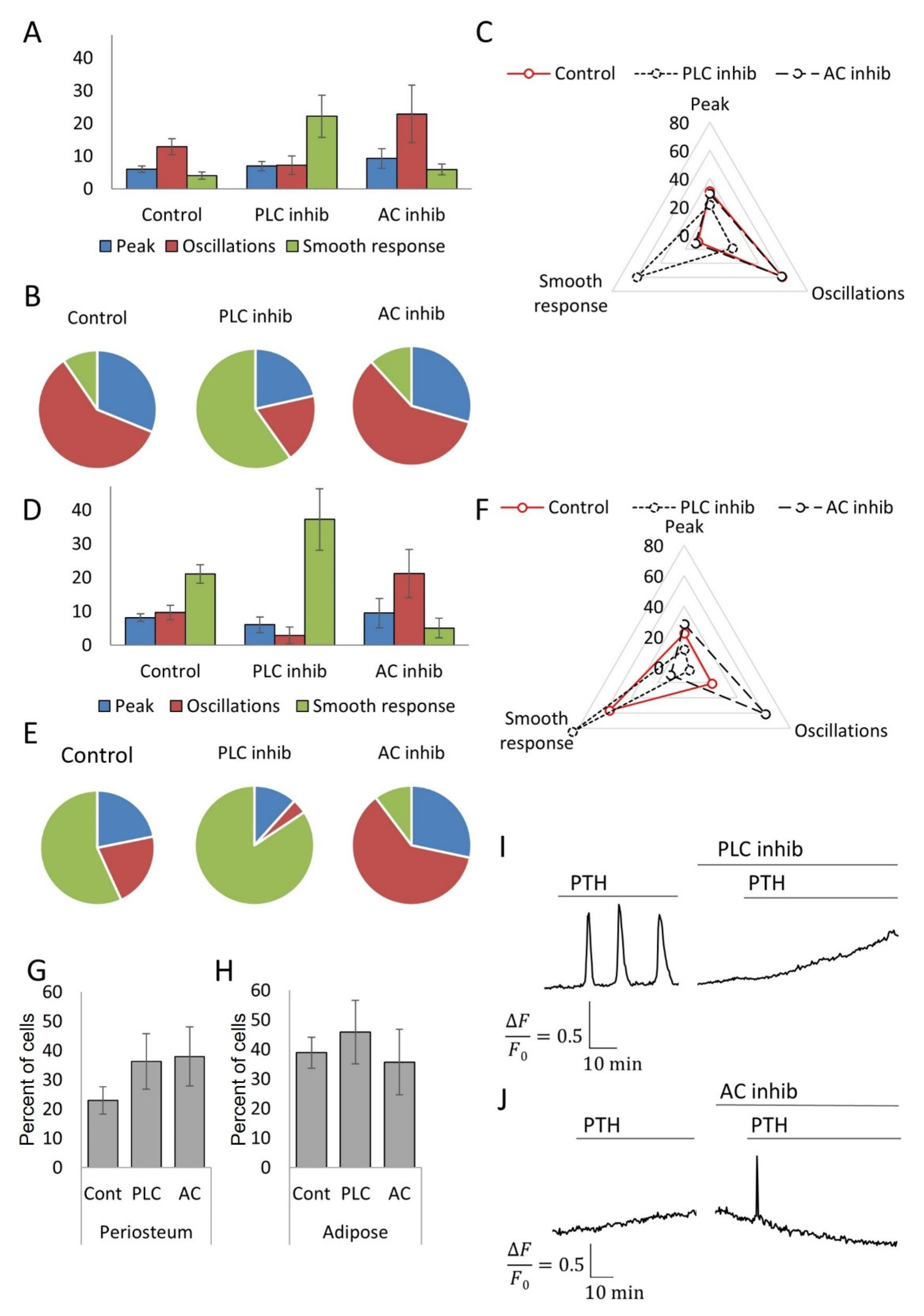
3.3. MSCs Derived from Different Tissue Depots Show a Different Profile of Calcium Signals in Response to PTH
3.4. Inhibitory Analysis Demonstrates the Possibility of Switching the Types of Ca2+ Responses to PTH in MSC Populations
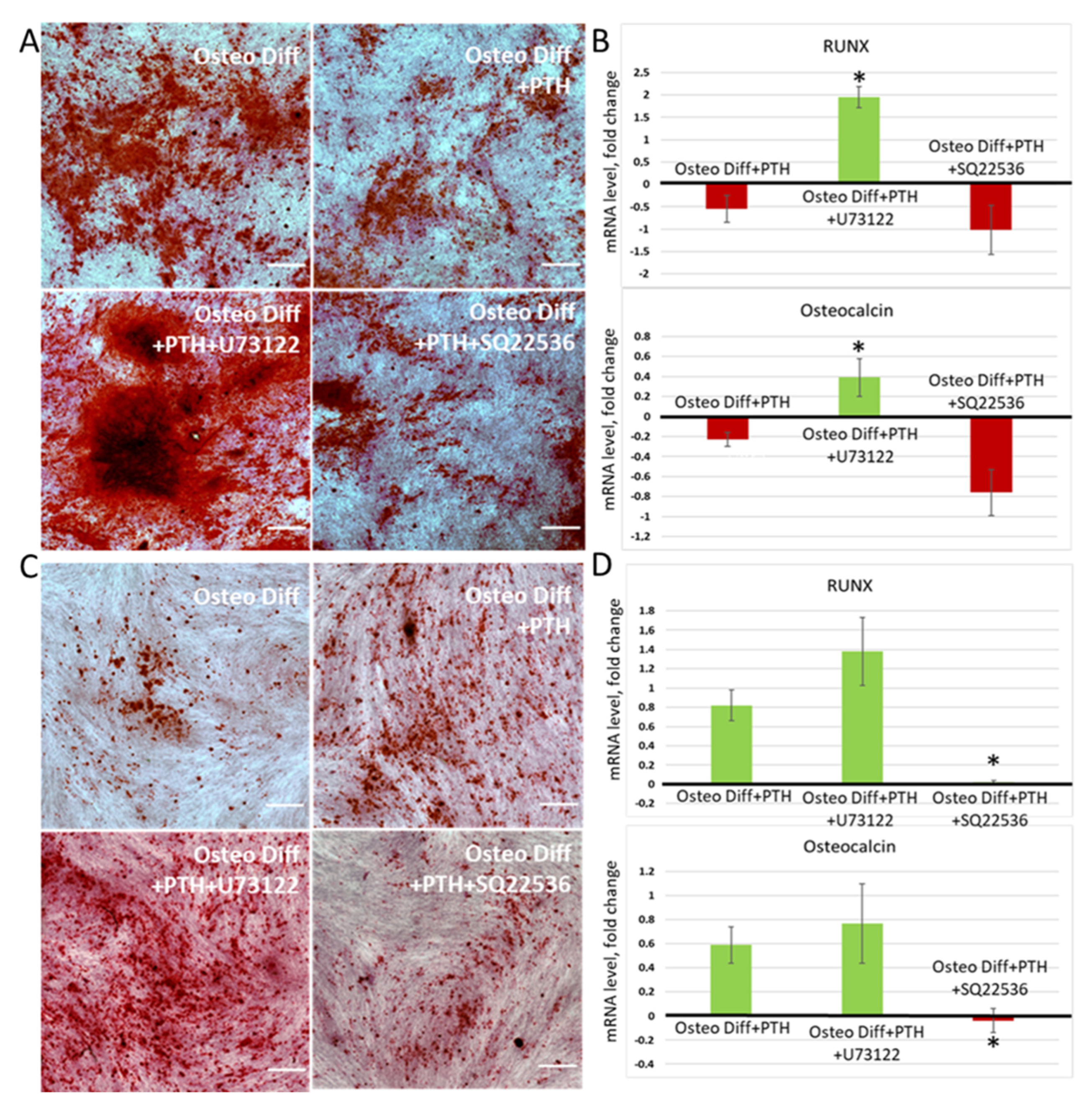
3.5. Switching of Intracellular Ca2+ Signaling Makes It Possible to Change the Vector of PTH Influence on the Osteogenic Differentiation of MSCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Knight, M.N.; Hankenson, K.D. Mesenchymal Stem Cells in Bone Regeneration. Adv. Wound Care 2013, 2, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Wein, M.N.; Kronenberg, H.M. Regulation of Bone Remodeling by Parathyroid Hormone. Cold Spring Harb. Perspect. Med. 2018, 8, a031237. [Google Scholar] [CrossRef] [Green Version]
- Wojda, S.J.; Donahue, S.W. Parathyroid hormone for bone regeneration. J. Orthop. Res. 2018, 36, 2586–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorontsova, M.V.; Kulebyakin, K.Y.; Makazan, N.V.; Sozaeva, L.S.; Tyurin-Kuzmin, P.A. Parathyroid Hormone in the Regulation of Bone Growth and Resorption in Health and Disease. Ann. Russ. Acad. Med. Sci. 2021, 76, 506–517. [Google Scholar] [CrossRef]
- Suzuki, A.; Ozono, K.; Kubota, T.; Kondou, H.; Tachikawa, K.; Michigami, T. PTH/cAMP/PKA signaling facilitates canonical Wnt signaling via inactivation of glycogen synthase kinase-3β in osteoblastic Saos-2 cells. J. Cell. Biochem. 2008, 104, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Zhao, X.; Yang, C.; Crane, J.; Xian, L.; Lu, W.; Wan, M.; Cao, X. Parathyroid hormone induces differentiation of mesenchymal stromal/stem cells by enhancing bone morphogenetic protein signaling. J. Bone Miner. Res. 2012, 27, 2001–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leder, B.Z. Parathyroid Hormone and Parathyroid Hormone-Related Protein Analogs in Osteoporosis Therapy. Curr. Osteoporos. Rep. 2017, 15, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Neer, R.M.; Arnaud, C.D.; Zanchetta, J.R.; Prince, R.; Gaich, G.A.; Reginster, J.Y.; Hodsman, A.B.; Eriksen, E.F.; Ish-Shalom, S.; Genant, H.K.; et al. Effect of parathyroid hormone (1-34) on fractures and bone mineral density in postmenopausal women with osteoporosis. N. Engl. J. Med. 2001, 344, 1434–1441. [Google Scholar] [CrossRef]
- Tashjian, A.H.; Chabner, B.A. Commentary on Clinical Safety of Recombinant Human Parathyroid Hormone 1-34 in the Treatment of Osteoporosis in Men and Postmenopausal Women. J. Bone Miner. Res. 2002, 17, 1151–1161. [Google Scholar] [CrossRef]
- Udagawa, N.; Takahashi, N.; Jimi, E.; Matsuzaki, K.; Tsurukai, T.; Itoh, K.; Nakagawa, N.; Yasuda, H.; Goto, M.; Tsuda, E.; et al. Osteoblasts/stromal cells stimulate osteoclast activation through expression of osteoclast differentiation factor/RANKL but not macrophage colony-stimulating factor: Receptor activator of NF-kappa B ligand. Bone 1999, 25, 517–523. [Google Scholar] [CrossRef]
- Tanaka, Y.; Nakayamada, S.; Okada, Y. Osteoblasts and osteoclasts in bone remodeling and inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Lotinun, S.; Sibonga, J.D.; Turner, R.T. Differential effects of intermittent and continuous administration of parathyroid hormone on bone histomorphometry and gene expression. Endocrine 2002, 17, 29–36. [Google Scholar] [CrossRef]
- Mantovani, G.; Bastepe, M.; Monk, D.; De Sanctis, L.; Thiele, S.; Usardi, A.; Ahmed, S.F.; Bufo, R.; Choplin, T.; De Filippo, G.; et al. Diagnosis and management of pseudohypoparathyroidism and related disorders: First international Consensus Statement. Nat. Rev. Endocrinol. 2018, 14, 476–500. [Google Scholar] [CrossRef] [PubMed]
- Pignolo, R.J.; Xu, M.; Russell, E.; Richardson, A.; Kaplan, J.; Billings, P.C.; Kaplan, F.S.; Shore, E.M. Heterozygous inactivation of Gnas in adipose-derived mesenchymal progenitor cells enhances osteoblast differentiation and promotes heterotopic ossification. J. Bone Miner. Res. 2011, 26, 2647–2655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.-H.; Ma, S.; Sutkeviciute, I.; Shen, D.-D.; Zhou, X.E.; de Waal, P.W.; Li, C.-Y.; Kang, Y.; Clark, L.J.; Jean-Alphonse, F.G.; et al. Structure and dynamics of the active human parathyroid hormone receptor-1. Science 2019, 364, 148–153. [Google Scholar] [CrossRef]
- Usdin, T.B.; Gruber, C.; Bonner, T.I. Identification and functional expression of a receptor selectively recognizing parathyroid hormone, the PTH2 receptor. J. Biol. Chem. 1995, 270, 15455–15458. [Google Scholar] [CrossRef] [Green Version]
- Bastepe, M.; Turan, S.; He, Q. Heterotrimeric G proteins in the control of parathyroid hormone actions. J. Mol. Endocrinol. 2017, 58, R203–R224. [Google Scholar] [CrossRef] [Green Version]
- Kulebyakin, K.; Tyurin-Kuzmin, P.; Efimenko, A.; Voloshin, N.; Kartoshkin, A.; Karagyaur, M.; Grigorieva, O.; Novoseletskaya, E.; Sysoeva, V.; Makarevich, P.; et al. Decreased Insulin Sensitivity in Telomerase-Immortalized Mesenchymal Stem Cells Affects Efficacy and Outcome of Adipogenic Differentiation in vitro. Front. Cell Dev. Biol. 2021, 9, 1961. [Google Scholar] [CrossRef]
- Tyurin-Kuzmin, P.A.; Fadeeva, J.I.; Kanareikina, M.A.; Kalinina, N.I.; Sysoeva, V.; Dyikanov, D.T.; Stambolsky, D.V.; Tkachuk, V.A. Activation of β-adrenergic receptors is required for elevated α1A-adrenoreceptors expression and signaling in mesenchymal stromal cells. Sci. Rep. 2016, 6, 32835. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Guo, J.; Iida-Klein, A.; Huang, X.; Abou-Samra, A.B.; Segre, G.V.; Bringhurst, F.R. Parathyroid hormone (PTH)/PTH-related peptide receptor density modulates activation of phospholipase C and phosphate transport by PTH in LLC-PK1 cells. Endocrinology 1995, 136, 3884–3891. [Google Scholar] [CrossRef] [PubMed]
- Takasu, H.; Guo, J.; Bringhurst, F.R. Dual Signaling and Ligand Selectivity of the Human PTH/PTHrP Receptor. J. Bone Miner. Res. 1999, 14, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Kotova, P.D.; Sysoeva, V.Y.; Rogachevskaja, O.A.; Bystrova, M.F.; Kolesnikova, A.S.; Tyurin-Kuzmin, P.A.; Fadeeva, J.I.; Tkachuk, V.A.; Kolesnikov, S.S. Functional expression of adrenoreceptors in mesenchymal stromal cells derived from the human adipose tissue. Biochim. Biophys. Acta-Mol. Cell Res. 2014, 1843, 1899–1908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, K.L.; Premont, R.T.; Lefkowitz, R.J. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Noto, Y.; Saito, M.; Ichizen, H.; Kida, H. Spinal cord compression by heterotopic ossification associated with pseudohypoparathyroidism. J. Int. Med. Res. 1997, 25, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Sarathi, V.; Patil, A.; Wade, R.; Bandgar, T.R.; Shah, N.S. Intraarticular heterotopic ossification as the initial manifestation in a child with pseudohypoparathyroidism 1a. Indian J. Pediatr. 2011, 78, 745–748. [Google Scholar] [CrossRef]
- Bastepe, M. GNAS mutations and heterotopic ossification. Bone 2018, 109, 80–85. [Google Scholar] [CrossRef]
- Yamaguchi, D.T.; Hahn, T.J.; Iida-Klein, A.; Kleeman, C.R.; Muallem, S. Parathyroid hormone-activated calcium channels in an osteoblast-like clonal osteosarcoma cell line. cAMP-dependent and cAMP-independent calcium channels. J. Biol. Chem. 1987, 262, 7711–7718. [Google Scholar] [CrossRef]
- Tyurin-Kuzmin, P.A.; Kalinina, N.I.; Kulebyakin, K.Y.; Balatskiy, A.V.; Sysoeva, V.Y.; Tkachuk, V.A. Angiotensin receptor subtypes regulate adipose tissue renewal and remodelling. FEBS J. 2020, 287, 1076–1087. [Google Scholar] [CrossRef] [Green Version]
- Mahon, M.J.; Donowitz, M.; Yun, C.C.; Segre, G.V. Na(+)/H(+) exchanger regulatory factor 2 directs parathyroid hormone 1 receptor signalling. Nature 2002, 417, 858–861. [Google Scholar] [CrossRef]
- Nelson, E.J.; Hellevuo, K.; Yoshimura, M.; Tabakoff, B. Ethanol-induced phosphorylation and potentiation of the activity of type 7 adenylyl cyclase. Involvement of protein kinase C delta. J. Biol. Chem. 2003, 278, 4552–4560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elli, F.M.; Mantovani, G. Pseudohypoparathyroidism, acrodysostosis, progressive osseous heteroplasia: Different names for the same spectrum of diseases? Endocrine 2021, 72, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Regard, J.B.; Malhotra, D.; Gvozdenovic-Jeremic, J.; Josey, M.; Chen, M.; Weinstein, L.S.; Lu, J.; Shore, E.M.; Kaplan, F.S.; Yang, Y. Activation of Hedgehog signaling by loss of GNAS causes heterotopic ossification. Nat. Med. 2013, 19, 1505–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regard, J.B.; Cherman, N.; Palmer, D.; Kuznetsov, S.A.; Celi, F.S.; Guettier, J.-M.; Chen, M.; Bhattacharyya, N.; Wess, J.; Coughlin, S.R.; et al. Wnt/β-catenin signaling is differentially regulated by Gα proteins and contributes to fibrous dysplasia. Proc. Natl. Acad. Sci. USA 2011, 108, 20101–20106. [Google Scholar] [CrossRef] [Green Version]
- Selvamurugan, N.; Chou, W.Y.; Pearman, A.T.; Pulumati, M.R.; Partridge, N.C. Parathyroid hormone regulates the rat collagenase-3 promoter in osteoblastic cells through the cooperative interaction of the activator protein-1 site and the runt domain binding sequence. J. Biol. Chem. 1998, 273, 10647–10657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellicelli, M.; Miller, J.A.; Arabian, A.; Gauthier, C.; Akhouayri, O.; Wu, J.Y.; Kronenberg, H.M.; St-Arnaud, R. The PTH-Gαs-Protein Kinase A Cascade Controls αNAC Localization To Regulate Bone Mass. Mol. Cell. Biol. 2014, 34, 1622–1633. [Google Scholar] [CrossRef] [Green Version]
- Thouverey, C.; Caverzasio, J. Suppression of p38α MAPK Signaling in Osteoblast Lineage Cells Impairs Bone Anabolic Action of Parathyroid Hormone. J. Bone Miner. Res. 2016, 31, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Liu, M.; Yang, D.; Bouxsein, M.L.; Saito, H.; Galvin, R.S.; Kuhstoss, S.A.; Thomas, C.C.; Schipani, E.; Baron, R.; et al. Suppression of Wnt Signaling by Dkk1 Attenuates PTH-Mediated Stromal Cell Response and New Bone Formation. Cell Metab. 2010, 11, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Schupbach, D.; Comeau-Gauthier, M.; Harvey, E.; Merle, G. Wnt modulation in bone healing. Bone 2020, 138, 115491. [Google Scholar] [CrossRef]
- Shore, E.M.; Ahn, J.; de Beur, S.J.; Li, M.; Xu, M.; Gardner, R.M.; Zasloff, M.A.; Whyte, M.P.; Levine, M.A.; Kaplan, F.S. Paternally Inherited Inactivating Mutations of the GNAS1 Gene in Progressive Osseous Heteroplasia. N. Engl. J. Med. 2002, 346, 99–106. [Google Scholar] [CrossRef]
- Wu, J.Y.; Aarnisalo, P.; Bastepe, M.; Sinha, P.; Fulzele, K.; Selig, M.K.; Chen, M.; Poulton, I.J.; Purton, L.E.; Sims, N.A.; et al. Gsα enhances commitment of mesenchymal progenitors to the osteoblast lineage but restrains osteoblast differentiation in mice. J. Clin. Investig. 2011, 121, 3492–3504. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Deng, P.; Iglesias-Bartolome, R.; Amornphimoltham, P.; Steffen, D.J.; Jin, Y.; Molinolo, A.A.; de Castro, L.F.; Ovejero, D.; Yuan, Q.; et al. Expression of an active Gαs mutant in skeletal stem cells is sufficient and necessary for fibrous dysplasia initiation and maintenance. Proc. Natl. Acad. Sci. USA 2018, 115, E428–E437. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Gene | Primers | Amplicon Size (bp) |
|---|---|---|---|
| 60S Ribosomal protein P0 | RPLP0 | F: GCTGCTGCCCGTGCTGGTG R: TGGTGCCCCTGGAGATTTTAGTGG | 130 |
| Runt-related transcription factor 2 | RUNX2 | F: TCTTAGAACAAATTCTGCCCTTT R: TGCTTTGGTCTTGAAATCACA | 136 |
| Osteocalcin | OCN | F: AGCAAAGGTGCAGCCTTTGT R: GCGCCTGGGTCTCTTCACT | 63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulebyakin, K.; Tyurin-Kuzmin, P.; Sozaeva, L.; Voloshin, N.; Nikolaev, M.; Chechekhin, V.; Vigovskiy, M.; Sysoeva, V.; Korchagina, E.; Naida, D.; et al. Dynamic Balance between PTH1R-Dependent Signal Cascades Determines Its Pro- or Anti-Osteogenic Effects on MSC. Cells 2022, 11, 3519. https://doi.org/10.3390/cells11213519
Kulebyakin K, Tyurin-Kuzmin P, Sozaeva L, Voloshin N, Nikolaev M, Chechekhin V, Vigovskiy M, Sysoeva V, Korchagina E, Naida D, et al. Dynamic Balance between PTH1R-Dependent Signal Cascades Determines Its Pro- or Anti-Osteogenic Effects on MSC. Cells. 2022; 11(21):3519. https://doi.org/10.3390/cells11213519
Chicago/Turabian StyleKulebyakin, Konstantin, Pyotr Tyurin-Kuzmin, Leila Sozaeva, Nikita Voloshin, Mikhail Nikolaev, Vadim Chechekhin, Maxim Vigovskiy, Veronika Sysoeva, Elizaveta Korchagina, Daria Naida, and et al. 2022. "Dynamic Balance between PTH1R-Dependent Signal Cascades Determines Its Pro- or Anti-Osteogenic Effects on MSC" Cells 11, no. 21: 3519. https://doi.org/10.3390/cells11213519
APA StyleKulebyakin, K., Tyurin-Kuzmin, P., Sozaeva, L., Voloshin, N., Nikolaev, M., Chechekhin, V., Vigovskiy, M., Sysoeva, V., Korchagina, E., Naida, D., & Vorontsova, M. (2022). Dynamic Balance between PTH1R-Dependent Signal Cascades Determines Its Pro- or Anti-Osteogenic Effects on MSC. Cells, 11(21), 3519. https://doi.org/10.3390/cells11213519