
The Autocrine Role of Placental Extracellular Vesicles from Missed Miscarriage in Causing Senescence: Possible Pathogenesis of Missed Miscarriage

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Tissue Collection and Preparation
2.2. Placental EVs Collection and Characterisation
2.3. RNA Extraction
2.4. RNA Sequencing
2.5. RNA Sequencing Analysis
2.6. Measurement of miRNAs in the Placentae by RT-PCR
2.7. Transfection of Trophoblast Cells with a Mimic or an Inhibitor of miRNA
2.8. Determine of the Senescent Changes in Placental Explants and Trophoblast Cells
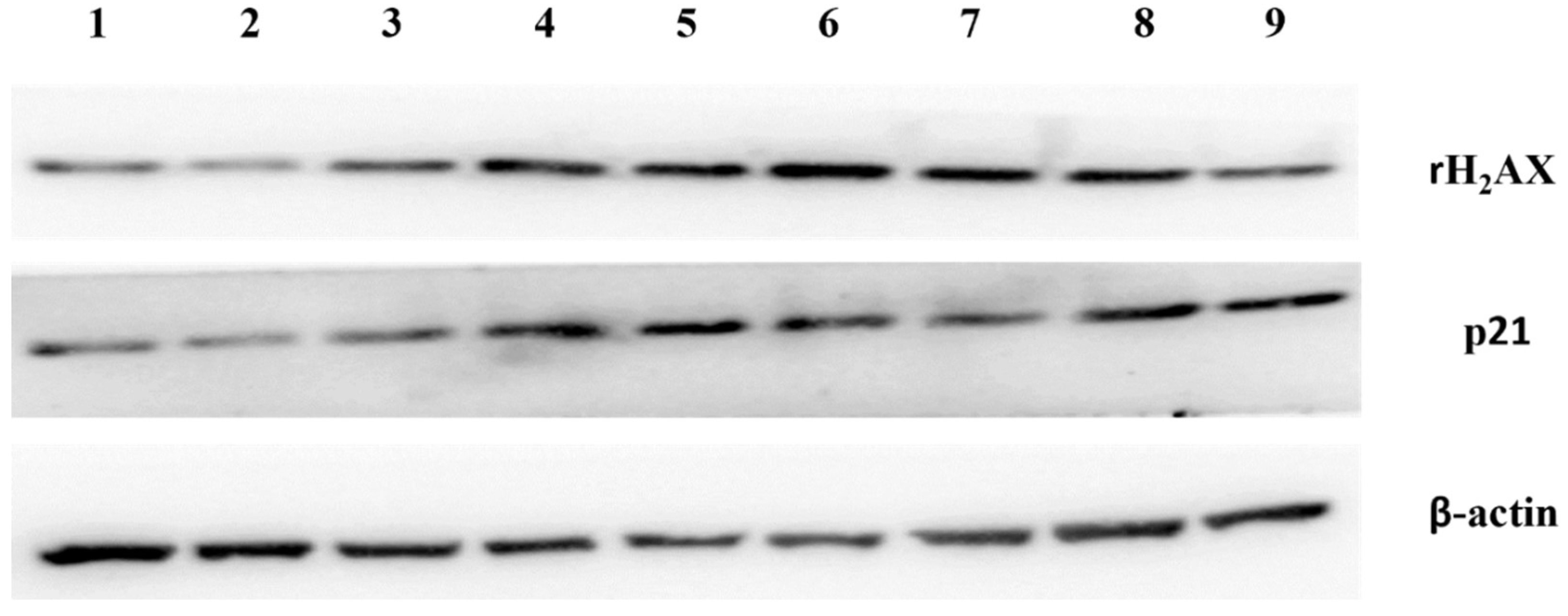
2.9. Western Blotting
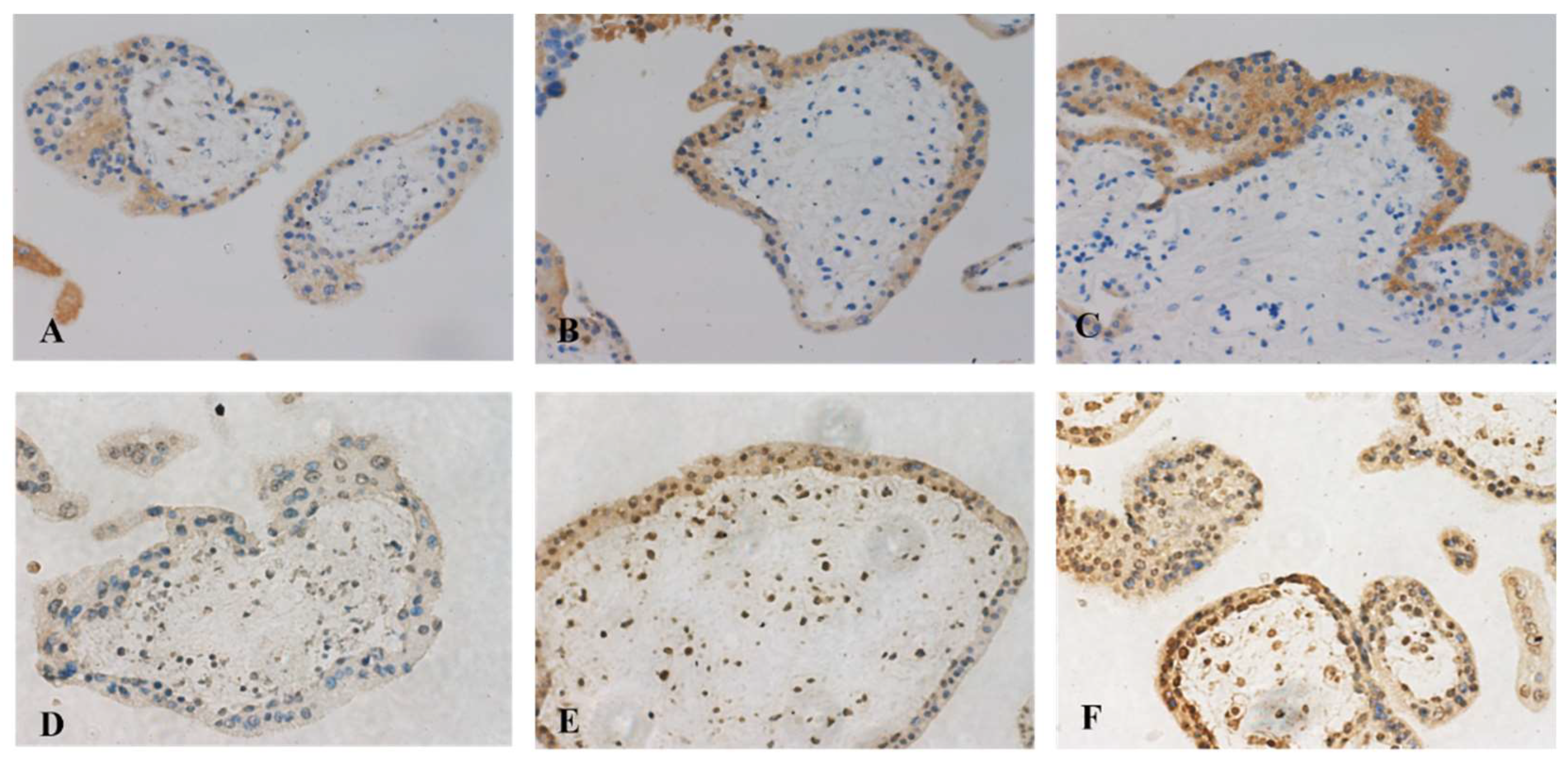
2.10. Immunohistochemistry
2.11. Statistical Analysis
3. Results
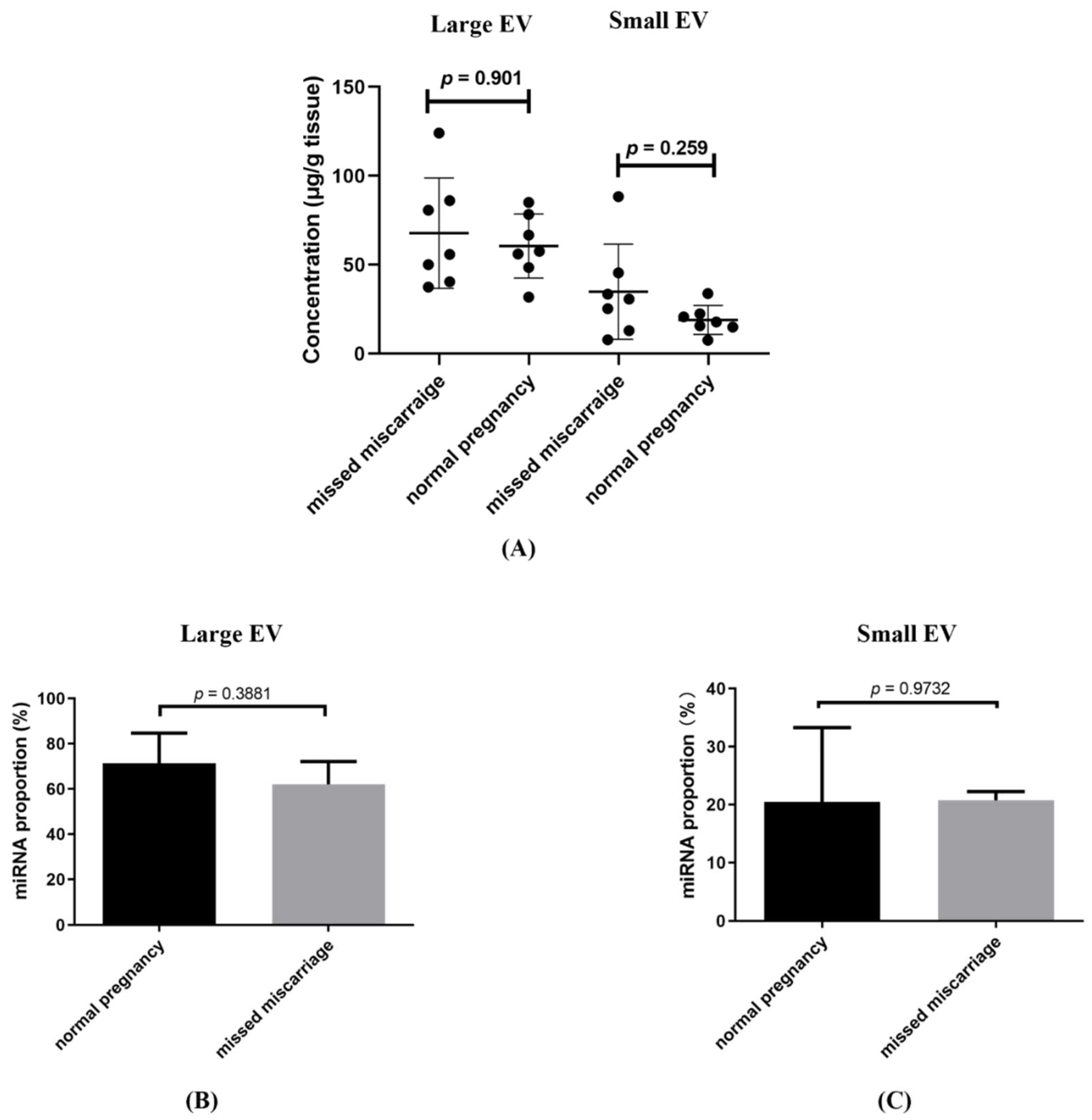
3.1. The Total Concentration of RNA Associated with Placental Large and Small EVs
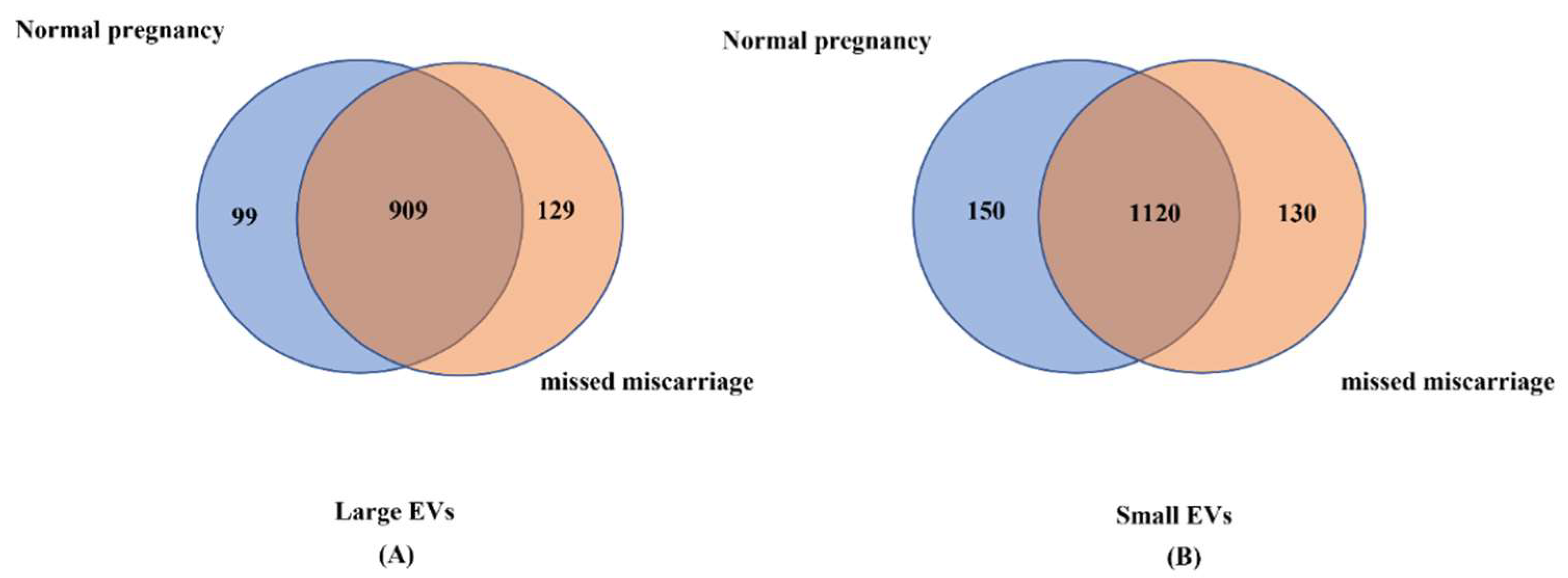
3.2. Analysis of miRNA Profiles in Placental EVs
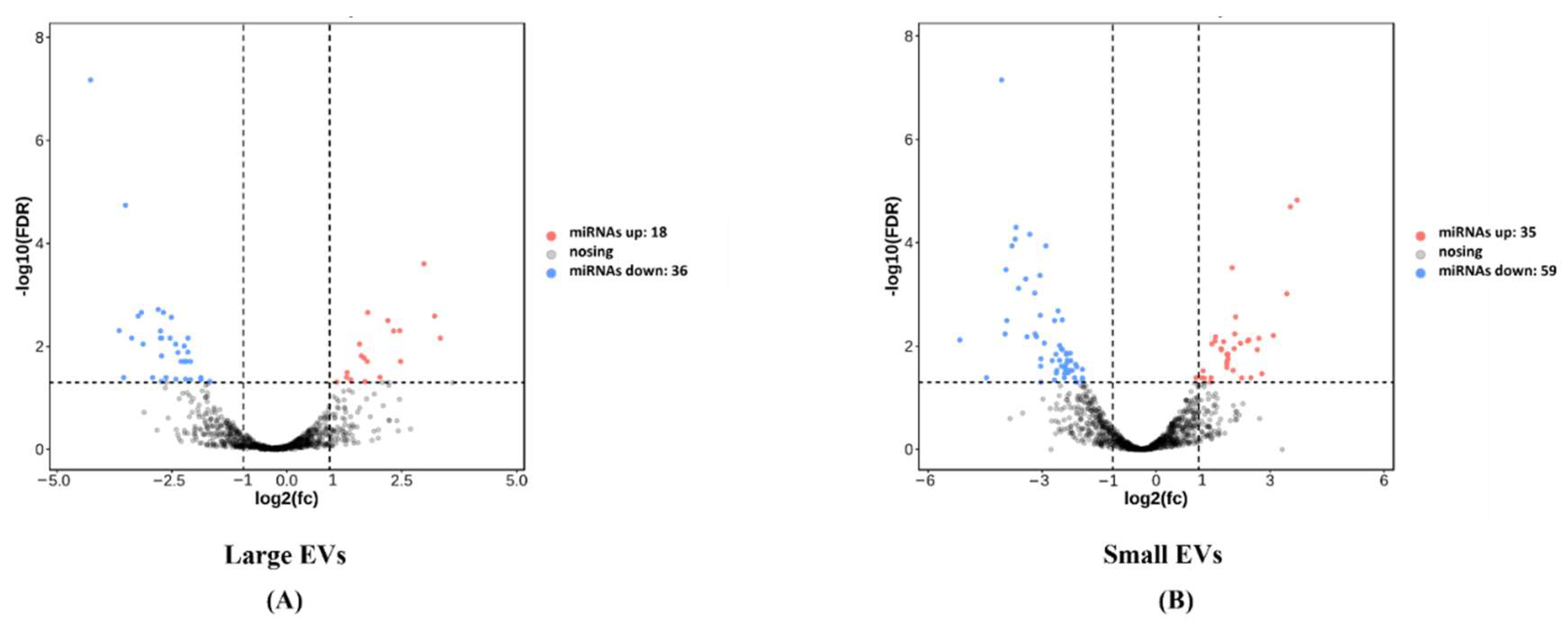
3.3. Comparison of the Expression Levels of miRNAs between EVs from Healthy and Missed Miscarriage Placentae
3.4. Analysis of the Functions or Signalling Pathways That the Differentially Expressed miRNAs Are Involved in
3.5. Differences in the miRNAs Encoded on Chromosome 19 (C19MC)
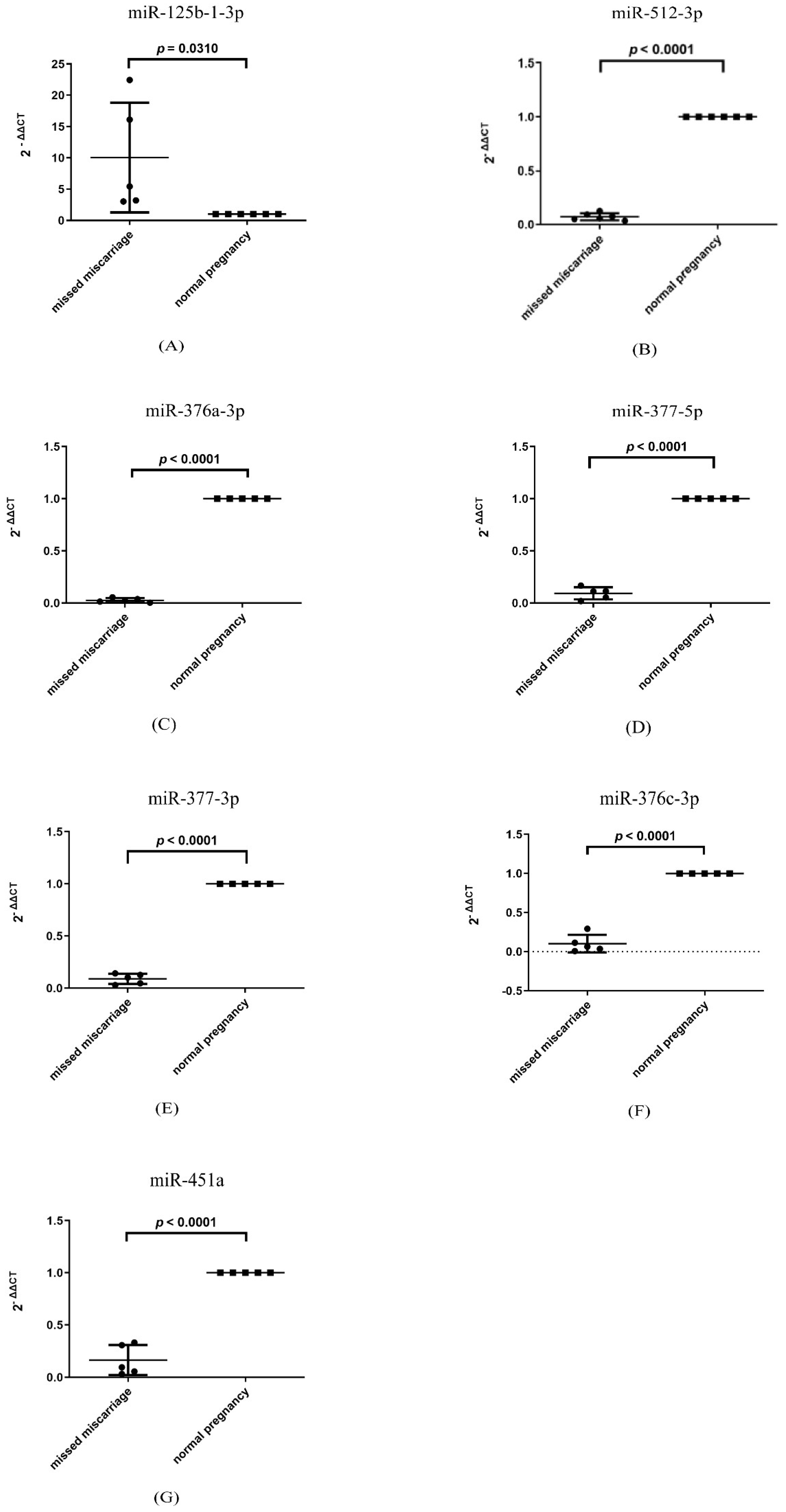
3.6. Changes in the Level of miRNAs That Are Involved in Senescence in Missed Miscarriage Placentae
3.7. Treatment with a Single miRNA Did Not Affect the Growth of Trophoblasts and Did Not Induce the Senescent Changes
3.8. Placental EVs from Missed Miscarriage Induced Senescent Changes in Normal Placentae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leveno, K.J.; Corton, M.M.; Bloom, S.L. Manual of Pregnancy Complications, 23th ed.; McGraw-Hill Medical: New York, NY, USA, 2013. [Google Scholar]
- Vaiman, D. Genetic regulation of recurrent spontaneous abortion in humans. Biomed. J. 2015, 38, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Oliver, A.; Overton, C. Diagnosis and management of miscarriage. Practitioner 2014, 258, 25–28. [Google Scholar] [PubMed]
- Burton, G.J.; Jauniaux, E. Placental oxidative stress: From miscarriage to preeclampsia. J. Soc. Gynecol. Investig. 2004, 11, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Jauniaux, E.; Burton, G.J. Pathophysiology of histological changes in early pregnancy loss. Placenta 2005, 26, 114–123. [Google Scholar] [CrossRef]
- Jauniaux, E.; Hempstock, J.; Greenwold, N.; Burton, G.J. Trophoblastic oxidative stress in relation to temporal and regional differences in maternal placental blood flow in normal and abnormal early pregnancies. Am. J. Pathol. 2003, 162, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Hustin, J.; Jauniaux, E.; Schaaps, J.P. Histological study of the materno-embryonic interface in spontaneous abortion. Placenta 1990, 11, 477–486. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, X.; Zhang, Y.; Feng, H.; Gao, J.; Liu, H.; Guo, F.; Chen, Q. Senescent Changes and Endoplasmic Reticulum Stress May Be Involved in the Pathogenesis of Missed Miscarriage. Front. Cell Dev. Biol. 2021, 9, 656549. [Google Scholar] [CrossRef]
- Liu, H.; Kang, M.; Wang, J.; Blenkiron, C.; Lee, A.; Wise, M.; Chamley, L.; Chen, Q. Estimation of the burden of human placental micro- and nano-vesicles extruded into the maternal blood from 8 to 12 weeks of gestation. Placenta 2018, 72, 41–47. [Google Scholar] [CrossRef]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular organelles important in intercellular communication. J. Proteom. 2010, 73, 1907–1920. [Google Scholar] [CrossRef]
- Nair, S.; Ormazabal, V.; Lappas, M.; McIntyre, H.D.; Salomon, C. Extracellular vesicles and their potential role inducing changes in maternal insulin sensitivity during gestational diabetes mellitus. Am. J. Reprod. Immunol. 2021, 85, e13361. [Google Scholar] [CrossRef]
- Tannetta, D.; Masliukaite, I.; Vatish, M.; Redman, C.; Sargent, I. Update of syncytiotrophoblast derived extracellular vesicles in normal pregnancy and preeclampsia. J. Reprod. Immunol. 2017, 119, 98–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiley, C.D.; Campisi, J. From Ancient Pathways to Aging Cells-Connecting Metabolism and Cellular Senescence. Cell Metab. 2016, 23, 1013–1021. [Google Scholar]
- Bartel, D.P. Metazoan MicroRNAs. Cell 2018, 173, 20–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Prieto, D.M.; Ospina-Prieto, S.; Chaiwangyen, W.; Schoenleben, M.; Markert, U.R. Pregnancy-associated miRNA-clusters. J. Reprod. Immunol. 2013, 97, 51–61. [Google Scholar] [CrossRef]
- Inno, R.; Kikas, T.; Lillepea, K.; Laan, M. Coordinated Expressional Landscape of the Human Placental miRNome and Transcriptome. Front. Cell Dev. Biol. 2021, 9, 697947. [Google Scholar] [CrossRef]
- Kotlabova, K.; Doucha, J.; Hromadnikova, I. Placental-specific microRNA in maternal circulation--identification of appropriate pregnancy-associated microRNAs with diagnostic potential. J. Reprod. Immunol. 2011, 89, 185–191. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Boriachek, K.; Islam, M.N.; Möller, A.; Salomon, C.; Nguyen, N.T.; Hossain, M.S.A.; Yamauchi, Y.; Shiddiky, M.J.A. Biological Functions and Current Advances in Isolation and Detection Strategies for Exosome Nanovesicles. Small 2018, 14, 1702153. [Google Scholar] [CrossRef]
- Xiao, X.; Xiao, F.; Zhao, M.; Tong, M.; Wise, M.R.; Stone, P.R.; Chamley, L.W.; Chen, Q. Treating normal early gestation placentae with preeclamptic sera produces extracellular micro and nano vesicles that activate endothelial cells. J. Reprod. Immunol. 2017, 120, 34–41. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, Y.; Sun, X.; Kang, M.; Zhao, M.; Wan, J.; Chen, Q. Exporting Proteins Associated with Senescence Repair via Extracellular Vesicles May Be Associated with Early Pregnancy Loss. Cells 2022, 11, 2772. [Google Scholar] [CrossRef]
- Shahnawaz, S.; Nawaz, U.S.; Zaugg, J.; Hussain, G.; Malik, N.; Dogar, M.Z.; Malik, S.A.; Albrecht, C. Dysregulated Autophagy Leads to Oxidative Stress and Aberrant Expression of ABC Transporters in Women with Early Miscarriage. Antioxidants 2021, 10, 1742. [Google Scholar] [CrossRef] [PubMed]
- Horibe, S.; Tanahashi, T.; Kawauchi, S.; Murakami, Y.; Rikitake, Y. Mechanism of recipient cell-dependent differences in exosome uptake. BMC Cancer 2018, 18, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sancho-Albero, M.; Navascués, N.; Mendoza, G.; Sebastián, V.; Arruebo, M.; Martín-Duque, P.; Santamaría, J. Exosome origin determines cell targeting and the transfer of therapeutic nanoparticles towards target cells. J. Nanobiotechnol. 2019, 17, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barteneva, N.S.; Fasler-Kan, E.; Bernimoulin, M.; Stern, J.N.; Ponomarev, E.D.; Duckett, L.; Vorobjev, I.A. Circulating microparticles: Square the circle. BMC Cell Biol. 2013, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular vesicles for drug delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Saber, S.H.; Ali, H.E.A.; Gaballa, R.; Gaballah, M.; Ali, H.I.; Zerfaoui, M.; Elmageed, Z.Y.A. Exosomes are the Driving Force in Preparing the Soil for the Metastatic Seeds: Lessons from the Prostate Cancer. Cells 2020, 9, 564. [Google Scholar] [CrossRef] [Green Version]
- Nyholm, A.M.; Lerche, C.M.; Manfé, V.; Biskup, E.; Johansen, P.; Morling, N.; Thomsen, B.M.; Glud, M.; Gniadecki, R. miR-125b induces cellular senescence in malignant melanoma. BMC Dermatol. 2014, 14, 8. [Google Scholar] [CrossRef] [Green Version]
- Maes, O.C.; Sarojini, H.; Wang, E. Stepwise up-regulation of microRNA expression levels from replicating to reversible and irreversible growth arrest states in WI-38 human fibroblasts. J. Cell. Physiol. 2009, 221, 109–119. [Google Scholar] [CrossRef]
- Shao, Y.; Dong, L.J.; Takahashi, Y.; Chen, J.; Liu, X.; Chen, Q.; Ma, J.X.; Li, X.R. miRNA-451a regulates RPE function through promoting mitochondrial function in proliferative diabetic retinopathy, American journal of physiology. Endocrinol. Metab. 2019, 316, E443–E452. [Google Scholar]
- Faraonio, R.; Salerno, P.; Passaro, F.; Sedia, C.; Iaccio, A.; Bellelli, R.; Nappi, T.C.; Comegna, M.; Romano, S.; Salvatore, G.; et al. A set of miRNAs participates in the cellular senescence program in human diploid fibroblasts. Cell Death Differ. 2012, 19, 713–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, Y.J.; Kwon, E.S.; Lee, S.M.; Kim, S.K.; Min, K.W.; Lim, J.Y.; Lee, B.; Kang, J.S.; Kwak, J.Y.; Son, Y.H.; et al. A subset of microRNAs in the Dlk1-Dio3 cluster regulates age-associated muscle atrophy by targeting Atrogin-1. J. Cachexia Sarcopenia Muscle 2020, 11, 1336–1350. [Google Scholar] [CrossRef] [PubMed]
- Akbari, A.; Majd, H.M.; Rahnama, R.; Heshmati, J.; Morvaridzadeh, M.; Agah, S.; Amini, S.M.; Masoodi, M. Cross-talk between oxidative stress signaling and microRNA regulatory systems in carcinogenesis: Focused on gastrointestinal cancers. Biomed. Pharmacother. 2020, 131, 110729. [Google Scholar] [CrossRef] [PubMed]
- Mouillet, J.F.; Goff, J.; Sadovsky, E.; Sun, H.; Parks, T.; Chu, T.; Sadovsky, Y. Transgenic expression of human C19MC miRNAs impacts placental morphogenesis. Placenta 2020, 101, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Ye, G.; Nadeem, L.; Ji, L.; Manchanda, T.; Wang, Y.; Zhao, Y.; Qiao, J.; Wang, Y.L.; Lye, S.; et al. MicroRNA-376c impairs transforming growth factor-β and nodal signaling to promote trophoblast cell proliferation and invasion. Hypertension 2013, 61, 864–872. [Google Scholar] [CrossRef] [Green Version]
- Sabapatha, A.; Gercel-Taylor, C.; Taylor, D.D. Specific isolation of placenta-derived exosomes from the circulation of pregnant women and their immunoregulatory consequences. Am. J. Reprod. Immunol. 2006, 56, 345–355. [Google Scholar] [CrossRef]
- Miura, K.; Higashijima, A.; Murakami, Y.; Tsukamoto, O.; Hasegawa, Y.; Abe, S.; Fuchi, N.; Miura, S.; Kaneuchi, M.; Masuzaki, H. Circulating chromosome 19 miRNA cluster microRNAs in pregnant women with severe pre-eclampsia. J. Obstet. Gynaecol. Res. 2015, 41, 1526–1532. [Google Scholar] [CrossRef]
- Montenegro, D.; Romero, R.; Kim, S.S.; Tarca, A.L.; Draghici, S.; Kusanovic, J.P.; Kim, J.S.; Lee, D.C.; Erez, O.; Gotsch, F.; et al. Expression patterns of microRNAs in the chorioamniotic membranes: A role for microRNAs in human pregnancy and parturition. J. Pathol. 2009, 217, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Rao, Z.; He, Z.; He, Y.; Guo, Z.; Kong, D.; Liu, J. MicroRNA-512-3p is upregulated, and promotes proliferation and cell cycle progression, in prostate cancer cells. Mol. Med. Rep. 2018, 17, 586–593. [Google Scholar] [CrossRef] [Green Version]
- Borgdorff, V.; Lleonart, M.E.; Bishop, C.L.; Fessart, D.; Bergin, A.H.; Overhoff, M.G.; Beach, D.H. Multiple microRNAs rescue from Ras-induced senescence by inhibiting p21(Waf1/Cip1). Oncogene 2010, 29, 2262–2271. [Google Scholar] [CrossRef] [Green Version]
- Karasu, E.; Eisenhardt, S.U.; Harant, J.; Huber-Lang, M. Extracellular Vesicles: Packages Sent With Complement. Front. Immunol. 2018, 9, 721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambe, S.; Yoshitake, H.; Yuge, K.; Ishida, Y.; Ali, M.M.; Takizawa, T.; Kuwata, T.; Ohkuchi, A.; Matsubara, S.; Suzuki, M.; et al. Human exosomal placenta-associated miR-517a-3p modulates the expression of PRKG1 mRNA in Jurkat cells. Biol. Reprod. 2014, 91, 129. [Google Scholar] [CrossRef] [PubMed]
- Collett, G.P.; Redman, C.W.; Sargent, I.L.; Vatish, M. Endoplasmic reticulum stress stimulates the release of extracellular vesicles carrying danger-associated molecular pattern (DAMP) molecules. Oncotarget 2018, 9, 6707–6717. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Tang, Y.; Liu, Y.; Wang, J.; Shen, Y.; Sun, X.; Kang, M.; Zhao, M.; Chen, Q. The Autocrine Role of Placental Extracellular Vesicles from Missed Miscarriage in Causing Senescence: Possible Pathogenesis of Missed Miscarriage. Cells 2022, 11, 3873. https://doi.org/10.3390/cells11233873
Zhang Y, Tang Y, Liu Y, Wang J, Shen Y, Sun X, Kang M, Zhao M, Chen Q. The Autocrine Role of Placental Extracellular Vesicles from Missed Miscarriage in Causing Senescence: Possible Pathogenesis of Missed Miscarriage. Cells. 2022; 11(23):3873. https://doi.org/10.3390/cells11233873
Chicago/Turabian StyleZhang, Yi, Yunhui Tang, Yang Liu, Jiayi Wang, Ye Shen, Xinyi Sun, Matthew Kang, Min Zhao, and Qi Chen. 2022. "The Autocrine Role of Placental Extracellular Vesicles from Missed Miscarriage in Causing Senescence: Possible Pathogenesis of Missed Miscarriage" Cells 11, no. 23: 3873. https://doi.org/10.3390/cells11233873
APA StyleZhang, Y., Tang, Y., Liu, Y., Wang, J., Shen, Y., Sun, X., Kang, M., Zhao, M., & Chen, Q. (2022). The Autocrine Role of Placental Extracellular Vesicles from Missed Miscarriage in Causing Senescence: Possible Pathogenesis of Missed Miscarriage. Cells, 11(23), 3873. https://doi.org/10.3390/cells11233873