
The Physiological Roles of the Exon Junction Complex in Development and Diseases

 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
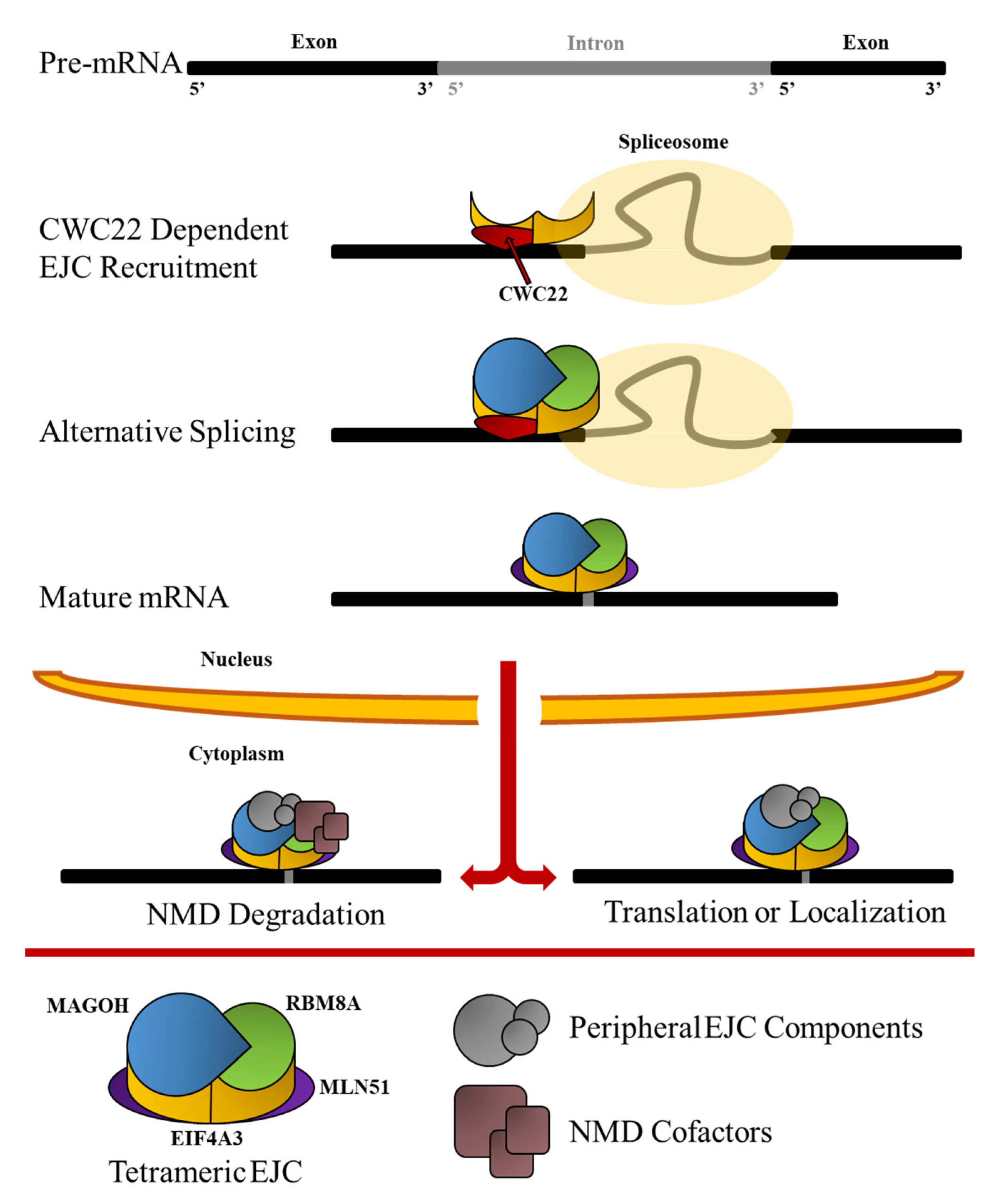
1.1. Formation of the EJC
1.2. The EJC at the Cellular Level
1.3. Overview of the EJC’s Position in Development
2. The Functions of EJC in mRNA Translation, Localization, and NMD
2.1. The mRNA Localization and Translation
2.2. Implications of MLN51 in P-Body Formation
2.3. The EJC Serves as a Crucial Link between Splicing and NMD
3. The Functions of EJC in Development
3.1. The EJC Regulates Neural Proliferation and Differentiation
3.2. Regulation of Splicing during Development
3.3. Control of mRNA Fate by the EJC during Neural Development
3.4. Crucial Roles of NMD in Processing mRNAs during Development
3.5. Crucial Roles of NMD in Diseases
4. EJC Components Are Implicated in Phenotypically Diverse Genetic Disorders
4.1. The Importance of RBM8A in the Developing Organism
4.2. EIF4A3, MAGOH, and Peripheral Components’ Differential Associations in Human Disease
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berk, A.J. Discovery of RNA splicing and genes in pieces. Proc. Natl. Acad. Sci. USA 2016, 113, 801–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartor, F.; Anderson, J.; McCaig, C.; Miedzybrodzka, Z.; Müller, B. Mutation of genes controlling mRNA metabolism and protein synthesis predisposes to neurodevelopmental disorders. Biochem. Soc. Trans. 2015, 43, 1259–1265. [Google Scholar] [CrossRef] [PubMed]
- Le Guienr, C.; Lejeune, F.; Galiana, D.; Kister, L.; Breathnach, R.; Stevenin, J.; Del Gatto-Konczak, F. TIA-1 and TIAR activate splicing of alternative exons with weak 5′ splice sites followed by a U-rich stretch on their own pre-mRNAs. J. Biol. Chem. 2001, 276, 40638–40646. [Google Scholar] [CrossRef] [Green Version]
- Girardot, M.; Bayet, E.; Maurin, J.; Fort, P.; Roux, P.; Raynaud, P. SOX9 has distinct regulatory roles in alternative splicing and transcription. Nucleic Acids Res. 2018, 46, 9106–9118. [Google Scholar] [CrossRef] [PubMed]
- Das, R.; Zhou, Z.L.; Reed, R. Functional association of U2 snRNP with the ATP-independent spliceosomal complex E. Mol. Cell 2000, 5, 779–787. [Google Scholar] [CrossRef]
- Chew, S.L.; Liu, H.X.; Mayeda, A.; Krainer, A.R. Evidence for the function of an exonic splicing enhancer after the first catalytic step of pre-mRNA splicing. Proc. Natl. Acad. Sci. USA 1999, 96, 10655–10660. [Google Scholar] [CrossRef] [Green Version]
- Mayeda, A.; Helfman, D.M.; Krainer, A.R. Modulation of exon skipping and inclusion by heterogeneous nuclear ribonucleoprotein A1 and pre-mRNA splicing factor SF2/ASF. Mol. Cell. Biol. 1993, 13, 2993–3001. [Google Scholar] [CrossRef] [Green Version]
- Kim, V.N.; Yong, J.; Kataoka, N.; Abel, L.; Diem, M.D.; Dreyfuss, G. The Y14 protein communicates to the cytoplasm the position of exon-exon junctions. EMBO J. 2001, 20, 2062–2068. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, N.; Diem, M.D.; Kim, V.N.; Yong, J.; Dreyfuss, G. Magoh, a human homolog of Drosophila mago nashi protein, is a component of the splicing-dependent exon-exon junction complex. EMBO J. 2001, 20, 6424–6433. [Google Scholar] [CrossRef]
- Gehring, N.H.; Kunz, J.B.; Neu-Yilik, G.; Breit, S.; Viegas, M.H.; Hentze, M.W.; Kulozik, A.E. Exon-junction complex components specify distinct routes of nonsense-mediated mRNA decay with differential cofactor requirements. Mol. Cell 2005, 20, 65–75. [Google Scholar] [CrossRef]
- Ballut, L.; Marchadier, B.; Baguet, A.; Tomasetto, C.; Seraphin, B.; Le Hir, H. The exon junction core complex is locked onto RNA by inhibition of eIF4AIII ATPase activity. Nat. Struct. Mol. Biol. 2005, 12, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Ferraiuolo, M.A.; Lee, C.S.; Ler, L.W.; Hsu, J.L.; Costa-Mattioli, M.; Luo, M.J.; Reed, R.; Sonenberg, N. A nuclear translation-like factor elF4AIII is recruited to the mRNA during splicing and functions in nonsense-mediated decay. Proc. Natl. Acad. Sci. USA 2004, 101, 4118–4123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.C.; Dostie, J.; Diem, M.D.; Feng, W.Q.; Mann, M.; Rappsilber, J.; Dreyfuss, G. eIF4A3 is a novel component of the exon junction complex. RNA 2004, 10, 200–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehring, N.H.; Lamprinaki, S.; Hentze, M.W.; Kulozik, A.E. The Hierarchy of Exon-Junction Complex Assembly by the Spliceosome Explains Key Features of Mammalian Nonsense-Mediated mRNA Decay. PLoS Biol. 2009, 7, e1000120. [Google Scholar] [CrossRef]
- Gerbracht, J.V.; Boehm, V.; Britto-Borges, T.; Kallabis, S.; Wiederstein, J.L.; Ciriello, S.; Aschemeier, D.U.; Krüger, M.; Frese, C.K.; Altmüller, J.; et al. CASC3 promotes transcriptome-wide activation of nonsense-mediated decay by the exon junction complex. Nucleic Acids Res. 2020, 48, 8626–8644. [Google Scholar] [CrossRef]
- Gerbracht, J.V.; Gehring, N.H. The exon junction complex: Structural insights into a faithful companion of mammalian mRNPs. Biochem. Soc. Trans. 2018, 46, 153–161. [Google Scholar] [CrossRef]
- Schlautmann, L.P.; Gehring, N.H. A Day in the Life of the Exon Junction Complex. Biomolecules 2020, 10, 866. [Google Scholar] [CrossRef]
- Steckelberg, A.L.; Altmueller, J.; Dieterich, C.; Gehring, N.H. CWC22-dependent pre-mRNA splicing and eIF4A3 binding enables global deposition of exon junction complexes. Nucleic Acids Res. 2015, 43, 4687–4700. [Google Scholar] [CrossRef] [Green Version]
- Steckelberg, A.-L.; Boehm, V.; Gromadzka, A.M.; Gehring, N.H. CWC22 connects pre-mRNA splicing and exon junction complex assembly. Cell Rep. 2012, 2, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Michelle, L.; Cloutier, A.; Toutant, J.; Shkreta, L.; Thibault, P.; Durand, M.; Garneau, D.; Gendron, D.; Lapointe, E.; Couture, S.; et al. Proteins Associated with the Exon Junction Complex Also Control the Alternative Splicing of Apoptotic Regulators. Mol. Cell. Biol. 2012, 32, 954–967. [Google Scholar] [CrossRef] [Green Version]
- Hauer, C.; Sieber, J.; Schwarzl, T.; Hollerer, I.; Curk, T.; Alleaume, A.-M.; Hentze, M.W.; Kulozik, A.E. Exon Junction Complexes Show a Distributional Bias toward Alternatively Spliced mRNAs and against mRNAs Coding for Ribosomal Proteins. Cell Rep. 2016, 16, 1588–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehm, V.; Britto-Borges, T.; Steckelberg, A.-L.; Singh, K.K.; Gerbracht, J.V.; Gueney, E.; Blazquez, L.; Altmüller, J.; Dieterich, C.; Gehring, N.H. Exon Junction Complexes Suppress Spurious Splice Sites to Safeguard Transcriptome Integrity. Mol. Cell 2018, 72, 482–495.e487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blazquez, L.; Emmett, W.; Faraway, R.; Pineda, J.M.B.; Bajew, S.; Gohr, A.; Haberman, N.; Sibley, C.R.; Bradley, R.K.; Irimia, M.; et al. Exon Junction Complex Shapes the Transcriptome by Repressing Recursive Splicing. Mol. Cell 2018, 72, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Mabin, J.W.; Woodward, L.A.; Patton, R.D.; Yi, Z.X.; Jia, M.X.; Wysocki, V.H.; Bundschuh, R.; Singh, G. The Exon Junction Complex Undergoes a Compositional Switch that Alters mRNP Structure and Nonsense-Mediated mRNA Decay Activity. Cell Rep. 2018, 25, 2431–2446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murachelli, A.G.; Ebert, J.; Basquin, C.; Le Hir, H.; Conti, E. The structure of the ASAP core complex reveals the existence of a Pinin-containing PSAP complex. Nat. Struct. Mol. Biol. 2012, 19, 378–386. [Google Scholar] [CrossRef] [PubMed]
- TANGE, T.Ø.; SHIBUYA, T.; JURICA, M.S.; MOORE, M.J. Biochemical analysis of the EJC reveals two new factors and a stable tetrameric protein core. RNA 2005, 11, 1869–1883. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.K.; Diem, M.D.; Dreyfuss, G.; Van Duyne, G.D. Structure of the Y14-Magoh core of the exon junction complex. Curr. Biol. CB 2003, 13, 933–941. [Google Scholar] [CrossRef] [Green Version]
- Kim, V.N.; Kataoka, N.; Dreyfuss, G. Role of the nonsense-mediated decay factor hUpf3 in the splicing-dependent exon-exon junction complex. Science 2001, 293, 1832–1836. [Google Scholar] [CrossRef] [Green Version]
- Le Hir, H.; Gatfield, D.; Izaurralde, E.; Moore, M.J. The exon–exon junction complex provides a binding platform for factors involved in mRNA export and nonsense-mediated mRNA decay. EMBO J. 2001, 20, 4987–4997. [Google Scholar] [CrossRef] [Green Version]
- Reichert, V.L.; Le Hir, H.; Jurica, M.S.; Moore, M.J. 5′ exon interactions within the human spliceosome establish a framework for exon junction complex structure and assembly. Genes Dev. 2002, 16, 2778–2791. [Google Scholar] [CrossRef] [Green Version]
- Gehring, N.H.; Lamprinaki, S.; Kulozik, A.E.; Hentze, M.W. Disassembly of exon junction complexes by PYM. Cell 2009, 137, 536–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diem, M.D.; Chan, C.C.; Younis, I.; Dreyfuss, G. PYM binds the cytoplasmic exon-junction complex and ribosomes to enhance translation of spliced mRNAs. Nat. Struct. Mol. Biol. 2007, 14, 1173–1179. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.S.; Johnson, T.L. The exon junction complex: A multitasking guardian of the transcriptome. Mol. Cell 2018, 72, 799–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehm, V.; Gehring, N.H. Exon junction complexes: Supervising the gene expression assembly line. Trends Genet. 2016, 32, 724–735. [Google Scholar] [CrossRef]
- Le Hir, H.; Andersen, G.R. Structural insights into the exon junction complex. Curr. Opin. Struct. Biol. 2008, 18, 112–119. [Google Scholar] [CrossRef]
- Le Hir, H.; Seraphin, B. EJCs at the heart of translational control. Cell 2008, 133, 213–216. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, N.; Yong, J.; Kim, V.N.; Velazquez, F.; Perkinson, R.A.; Wang, F.; Dreyfuss, G. Pre-mRNA splicing imprints mRNA in the nucleus with a novel RNA-binding protein that persists in the cytoplasm. Mol. Cell 2000, 6, 673–682. [Google Scholar] [CrossRef]
- Mohr, S.E.; Dillon, S.T.; Boswell, R.E. The RNA-binding protein Tsunagi interacts with Mago Nashi to establish polarity and localize oskar mRNA during Drosophila oogenesis. Genes Dev. 2001, 15, 2886–2899. [Google Scholar] [CrossRef]
- Salicioni, A.M.; Xi, M.; Vanderveer, L.A.; Balsara, B.; Testa, J.R.; Dunbrack Jr, R.L.; Godwin, A.K. Identification and structural analysis of human RBM8A and RBM8B: Two highly conserved RNA-binding motif proteins that interact with OVCA1, a candidate tumor suppressor. Genomics 2000, 69, 54–62. [Google Scholar] [CrossRef]
- Boswell, R.E.; Prout, M.E.; Steichen, J.C. Mutations in a newly identified Drosophila melanogaster gene, mago nashi, disrupt germ cell formation and result in the formation of mirror-image symmetrical double abdomen embryos. Development 1991, 113, 373–384. [Google Scholar] [CrossRef]
- Holzmann, K.; Gerner, C.; Pöltl, A.; Schäfer, R.; Obrist, P.; Ensinger, C.; Grimm, R.; Sauermann, G. A human common nuclear matrix protein homologous to eukaryotic translation initiation factor 4A. Biochem. Biophys. Res. Commun. 2000, 267, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Kong, X.-T.; Zhang, G.-A.; Hou, X.-J.; Qiang, H.; Zhong, R.-Q. Proteomics-based identification of DEAD-box protein 48 as a novel autoantigen, a prospective serum marker for pancreatic cancer. Biochem. Biophys. Res. Commun. 2005, 330, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Baguet, A.; Degot, S.; Cougot, N.; Bertrand, E.; Chenard, M.P.; Wendling, C.; Kessler, P.; Le Hir, H.; Rio, M.C.; Tomasetto, C. The exon-junction-complex-component metastatic lymph node 51 functions in stress-granule assembly. J. Cell Sci. 2007, 120, 2774–2784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dostie, J.; Dreyfuss, G. Translation is required to remove Y14 from mRNAs in the cytoplasm. Curr. Biol. 2002, 12, 1060–1067. [Google Scholar] [CrossRef] [Green Version]
- Buchwald, G.; Schussler, S.; Basquin, C.; Le Hir, H.; Conti, E. Crystal structure of the human eIF4AIII-CWC22 complex shows how a DEAD-box protein is inhibited by a MIF4G domain. Proc. Natl. Acad. Sci. USA 2013, 110, E4611–E4618. [Google Scholar] [CrossRef] [Green Version]
- Noble, C.G.; Song, H.W. MLN51 Stimulates the RNA-Helicase Activity of eIF4AIII. PLoS ONE 2007, 2, e303. [Google Scholar] [CrossRef] [Green Version]
- Le Hir, H.; Izaurralde, E.; Maquat, L.E.; Moore, M.J. The spliceosome deposits multiple proteins 20–24 nucleotides upstream of mRNA exon–exon junctions. EMBO J. 2000, 19, 6860–6869. [Google Scholar] [CrossRef] [Green Version]
- Andersen, C.B.F.; Ballut, L.; Johansen, J.S.; Chamieh, H.; Nielsen, K.H.; Oliveira, C.L.P.; Pedersen, J.S.; Seraphin, B.; Le Hir, H.; Andersen, G.R. Structure of the exon junction core complex with a trapped DEAD-box ATPase bound to RNA. Science 2006, 313, 1968–1972. [Google Scholar] [CrossRef]
- Wahl, M.C.; Will, C.L.; Luhrmann, R. The Spliceosome: Design Principles of a Dynamic RNP Machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [Green Version]
- Alexandrov, A.; Colognori, D.; Shu, M.D.; Steitz, J.A. Human spliceosomal protein CWC22 plays a role in coupling splicing to exon junction complex deposition and nonsense-mediated decay. Proc. Natl. Acad. Sci. USA 2012, 109, 21313–21318. [Google Scholar] [CrossRef] [Green Version]
- Shiimori, M.; Inoue, K.; Sakamoto, H. A specific set of exon junction complex subunits is required for the nuclear retention of unspliced RNAs in Caenorhabditis elegans. Mol. Cell. Biol. 2013, 33, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, R.; Handler, D.; Ish-Horowicz, D.; Brennecke, J. The exon junction complex is required for definition and excision of neighboring introns in Drosophila. Genes Dev. 2014, 28, 1772–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Murigneux, V.; Le Hir, H. Transcriptome-wide modulation of splicing by the exon junction complex. Genome Biol. 2014, 15, 18. [Google Scholar] [CrossRef] [PubMed]
- Irimia, M.; Weatheritt, R.J.; Ellis, J.D.; Parikshak, N.N.; Gonatopoulos-Pournatzis, T.; Babor, M.; Quesnel-Vallieres, M.; Tapial, J.; Raj, B.; O’Hanlon, D.; et al. A Highly Conserved Program of Neuronal Microexons Is Misregulated in Autistic Brains. Cell 2014, 159, 1511–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malone, C.D.; Mestdagh, C.; Akhtar, J.; Kreim, N.; Deinhard, P.; Sachidanandam, R.; Treisman, J.; Roignant, J.Y. The exon junction complex controls transposable element activity by ensuring faithful splicing of the piwi transcript. Genes Dev. 2014, 28, 1786–1799. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, U.; Im, K.-B.; Benzing, C.; Janjetovic, S.; Rippe, K.; Lichter, P.; Wachsmuth, M. Assembly and mobility of exon–exon junction complexes in living cells. RNA 2009, 15, 862–876. [Google Scholar] [CrossRef] [Green Version]
- Daguenet, E.; Baguet, A.; Degot, S.; Schmidt, U.; Alpy, F.; Wendling, C.; Spiegelhalter, C.; Kessler, P.; Rio, M.C.; Le Hir, H.; et al. Perispeckles are major assembly sites for the exon junction core complex. Mol. Biol. Cell 2012, 23, 1765–1782. [Google Scholar] [CrossRef]
- Wiegand, H.L.; Lu, S.H.; Cullen, B.R. Exon junction complexes mediate the enhancing effect of splicing on mRNA expression. Proc. Natl. Acad. Sci. USA 2003, 100, 11327–11332. [Google Scholar] [CrossRef] [Green Version]
- Nott, A.; Le Hir, H.; Moore, M.J. Splicing enhances translation in mammalian cells: An additional function of the exon junction complex. Genes Dev. 2004, 18, 210–222. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.C.; Choe, J.; Chi, S.G.; Kim, Y.K. Exon junction complex enhances translation of spliced mRNAs at multiple steps. Biochem. Biophys. Res. Commun. 2009, 384, 334–340. [Google Scholar] [CrossRef]
- Ghosh, S.; Obrdlik, A.; Marchand, V.; Ephrussi, A. The EJC Binding and Dissociating Activity of PYM Is Regulated in Drosophila. PLoS Genet. 2014, 10, e1004455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacios, I.M.; Gatfield, D.; St Johnston, D.; Izaurralde, E. An eIF4AIII-containing complex required for mRNA localization and nonsense-mediated mRNA decay. Nature 2004, 427, 753–757. [Google Scholar] [CrossRef] [PubMed]
- Cougot, N.; Daguenet, E.; Baguet, A.; Cavalier, A.; Thomas, D.; Bellaud, P.; Fautrel, A.; Godey, F.; Bertrand, E.; Tomasetto, C.; et al. Overexpression of MLN51 triggers P-body disassembly and formation of a new type of RNA granules. J. Cell Sci. 2014, 127, 4692–4701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hug, N.; Longman, D.; Cáceres, J.F. Mechanism and regulation of the nonsense-mediated decay pathway. Nucleic Acids Res. 2016, 44, 1483–1495. [Google Scholar] [CrossRef] [Green Version]
- Kurosaki, T.; Popp, M.W.; Maquat, L.E. Quality and quantity control of gene expression by nonsense-mediated mRNA decay. Nat. Rev. Mol. Cell Biol. 2019, 20, 406–420. [Google Scholar] [CrossRef]
- Yi, Z.; Sanjeev, M.; Singh, G. The Branched Nature of the Nonsense-Mediated mRNA Decay Pathway. Trends Genet. 2021, 37, 143–159. [Google Scholar] [CrossRef]
- Holbrook, J.A.; Neu-Yilik, G.; Hentze, M.W.; Kulozik, A.E. Nonsense-mediated decay approaches the clinic. Nat. Genet. 2004, 36, 801–808. [Google Scholar] [CrossRef]
- Khajavi, M.; Inoue, K.; Lupski, J.R. Nonsense-mediated mRNA decay modulates clinical outcome of genetic disease. Eur. J. Hum. Genet. 2006, 14, 1074–1081. [Google Scholar] [CrossRef] [Green Version]
- Bhuvanagiri, M.; Schlitter, A.M.; Hentze, M.W.; Kulozik, A.E. NMD: RNA biology meets human genetic medicine. Biochem. J. 2010, 430, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Gardner, L.B. Nonsense-mediated RNA decay regulation by cellular stress: Implications for tumorigenesis. Mol. Cancer Res. 2010, 8, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Wittkopp, N.; Huntzinger, E.; Weiler, C.; Sauliere, J.; Schmidt, S.; Sonawane, M.; Izaurralde, E. Nonsense-Mediated mRNA Decay Effectors Are Essential for Zebrafish Embryonic Development and Survival. Mol. Cell. Biol. 2009, 29, 3517–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, D.L.; Watkins-Chow, D.E.; Schreck, K.C.; Pierfelice, T.J.; Larson, D.M.; Burnetti, A.J.; Liaw, H.J.; Myung, K.; Walsh, C.A.; Gaiano, N.; et al. The exon junction complex component Magoh controls brain size by regulating neural stem cell division. Nature Neurosci. 2010, 13, 551–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McSweeney, C.; Dong, F.P.; Chen, M.; Vitale, J.; Xu, L.; Crowley, N.; Luscher, B.; Zou, D.H.; Mao, Y.W. Full function of exon junction complex factor, Rbm8a, is critical for interneuron development. Transl. Psychiatry 2020, 10, 17. [Google Scholar] [CrossRef]
- Zou, D.H.; McSweeney, C.; Sebastian, A.; Reynolds, D.J.; Dong, F.P.; Zhou, Y.J.; Deng, D.Z.; Wang, Y.G.; Liu, L.; Zhu, J.; et al. A critical role of RBM8a in proliferation and differentiation of embryonic neural progenitors. Neural Dev. 2015, 10, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgi, C.; Yeo, G.W.; Stone, M.E.; Katz, D.B.; Burge, C.; Turrigiano, G.; Moore, M.J. The EJC factor eIF4AIII modulates synaptic strength and neuronal protein expression. Cell 2007, 130, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Alachkar, A.; Jiang, D.; Harrison, M.; Zhou, Y.; Chen, G.; Mao, Y. An EJC factor RBM8a Regulates Anxiety Behaviors. Curr. Mol. Med. 2013, 13, 887–899. [Google Scholar] [CrossRef]
- Ohbayashi, N.; Taira, N.; Kawakami, S.; Togi, S.; Sato, N.; Ikeda, O.; Kamitani, S.; Muromoto, R.; Sekine, Y.; Matsuda, T. An RNA biding protein, Y14 interacts with and modulates STAT3 activation. Biochem. Biophys. Res. Commun. 2008, 372, 475–479. [Google Scholar] [CrossRef] [Green Version]
- Muromoto, R.; Taira, N.; Ikeda, O.; Shiga, K.; Kamitani, S.; Togi, S.; Kawakami, S.; Sekine, Y.; Nanbo, A.; Oritani, K.; et al. The exon-junction complex proteins, Y14 and MAGOH regulate STAT3 activation. Biochem. Biophys. Res. Commun. 2009, 382, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Ishigaki, Y.; Nakamura, Y.; Tatsuno, T.; Hashimoto, M.; Shimasaki, T.; Iwabuchi, K.; Tomosugi, N. Depletion of RNA-binding protein RBM8A (Y14) causes cell cycle deficiency and apoptosis in human cells. Exp. Biol. Med. 2013, 238, 889–897. [Google Scholar] [CrossRef]
- Albers, C.A.; Paul, D.S.; Schulze, H.; Freson, K.; Stephens, J.C.; Smethurst, P.A.; Jolley, J.D.; Cvejic, A.; Kostadima, M.; Bertone, P.; et al. Compound inheritance of a low-frequency regulatory SNP and a rare null mutation in exon-junction complex subunit RBM8A causes TAR syndrome. Nature Genet. 2012, 44, 435-U248. [Google Scholar] [CrossRef] [Green Version]
- Klopocki, E.; Schulze, H.; Strauß, G.; Ott, C.-E.; Hall, J.; Trotier, F.; Fleischhauer, S.; Greenhalgh, L.; Newbury-Ecob, R.A.; Neumann, L.M. Complex inheritance pattern resembling autosomal recessive inheritance involving a microdeletion in thrombocytopenia–absent radius syndrome. Am. J. Hum. Genet. 2007, 80, 232–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favaro, F.P.; Alvizi, L.; Zechi-Ceide, R.M.; Bertola, D.; Felix, T.M.; de Souza, J.; Raskin, S.; Twigg, S.R.F.; Weiner, A.M.J.; Armas, P.; et al. A Noncoding Expansion in EIF4A3 Causes Richieri-Costa-Pereira Syndrome, a Craniofacial Disorder Associated with Limb Defects. Am. J. Hum. Genet. 2014, 94, 120–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarpey, P.S.; Raymond, F.L.; Nguyen, L.S.; Rodriguez, J.; Hackett, A.; Vandeleur, L.; Smith, R.; Shoubridge, C.; Edkins, S.; Stevens, C. Mutations in UPF3B, a member of the nonsense-mediated mRNA decay complex, cause syndromic and nonsyndromic mental retardation. Nat. Genet. 2007, 39, 1127–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safieddine, A.; Coleno, E.; Salloum, S.; Imbert, A.; Traboulsi, A.M.; Kwon, O.S.; Lionneton, F.; Georget, V.; Robert, M.C.; Gostan, T.; et al. A choreography of centrosomal mRNAs reveals a conserved localization mechanism involving active polysome transport. Nat. Commun. 2021, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Chouaib, R.; Safieddine, A.; Pichon, X.; Imbert, A.; Kwon, O.S.; Samacoits, A.; Traboulsi, A.M.; Robert, M.C.; Tsanov, N.; Coleno, E.; et al. A Dual Protein-mRNA Localization Screen Reveals Compartmentalized Translation and Widespread Co-translational RNA Targeting. Dev. Cell 2020, 54, 773–791. [Google Scholar] [CrossRef]
- Hubstenberger, A.; Courel, M.; Bénard, M.; Souquere, S.; Ernoult-Lange, M.; Chouaib, R.; Yi, Z.; Morlot, J.-B.; Munier, A.; Fradet, M.; et al. P-Body Purification Reveals the Condensation of Repressed mRNA Regulons. Mol. Cell 2017, 68, 144–157.e145. [Google Scholar] [CrossRef] [Green Version]
- Chin, A.; Lécuyer, E. RNA localization: Making its way to the center stage. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 2956–2970. [Google Scholar] [CrossRef]
- Holt, C.E.; Bullock, S.L. Subcellular mRNA Localization in Animal Cells and Why It Matters. Science 2009, 326, 1212–1216. [Google Scholar] [CrossRef] [Green Version]
- Bullock, S.L. Translocation of mRNAs by molecular motors: Think complex? Semin Cell Dev. Biol. 2007, 18, 194–201. [Google Scholar] [CrossRef]
- Bertrand, E.; Chartrand, P.; Schaefer, M.; Shenoy, S.M.; Singer, R.H.; Long, R.M. Localization of ASH1 mRNA particles in living yeast. Mol. Cell 1998, 2, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Buxbaum, A.R.; Haimovich, G.; Singer, R.H. In the right place at the right time: Visualizing and understanding mRNA localization. Nat. Rev. Mol. Cell Biol. 2015, 16, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Trcek, T.; Singer, R.H. The cytoplasmic fate of an mRNP is determined cotranscriptionally: Exception or rule? Genes Dev. 2010, 24, 1827–1831. [Google Scholar] [CrossRef] [Green Version]
- Kannaiah, S.; Amster-Choder, O. Protein targeting via mRNA in bacteria. Biochim. Biophys. Acta 2014, 1843, 1457–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.H.; Lee, S.J.; Gardiner, A.S.; Perrone-Bizzozero, N.I.; Yoo, S. Different motif requirements for the localization zipcode element of β-actin mRNA binding by HuD and ZBP1. Nucleic Acids Res. 2015, 43, 7432–7446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, P.; Blobel, G. Purification of a membrane-associated protein complex required for protein translocation across the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 1980, 77, 7112–7116. [Google Scholar] [CrossRef] [Green Version]
- Walter, P.; Blobel, G. Signal recognition particle contains a 7S RNA essential for protein translocation across the endoplasmic reticulum. Nature 1982, 299, 691–698. [Google Scholar] [CrossRef]
- Gilmore, R.; Blobel, G.; Walter, P. Protein translocation across the endoplasmic reticulum. I. Detection in the microsomal membrane of a receptor for the signal recognition particle. J. Cell Biol. 1982, 95, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Hachet, O.; Ephrussi, A. Drosophila Y14 shuttles to the posterior of the oocyte and is required for oskar mRNA transport. Curr. Biol. CB 2001, 11, 1666–1674. [Google Scholar] [CrossRef] [Green Version]
- Hachet, O.; Ephrussi, A. Splicing of oskar RNA in the nucleus is coupled to its cytoplasmic localization. Nature 2004, 428, 959–963. [Google Scholar] [CrossRef]
- Kwon, O.S.; Mishra, R.; Safieddine, A.; Coleno, E.; Alasseur, Q.; Faucourt, M.; Barbosa, I.; Bertrand, E.; Spassky, N.; Le Hir, H. Exon junction complex dependent mRNA localization is linked to centrosome organization during ciliogenesis. Nat. Commun. 2021, 12, s41467. [Google Scholar] [CrossRef]
- Lambert, J.D.; Nagy, L.M. Asymmetric inheritance of centrosomally localized mRNAs during embryonic cleavages. Nature 2002, 420, 682–686. [Google Scholar] [CrossRef] [PubMed]
- Lécuyer, E.; Yoshida, H.; Parthasarathy, N.; Alm, C.; Babak, T.; Cerovina, T.; Hughes, T.R.; Tomancak, P.; Krause, H.M. Global analysis of mRNA localization reveals a prominent role in organizing cellular architecture and function. Cell 2007, 131, 174–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sepulveda, G.; Antkowiak, M.; Brust-Mascher, I.; Mahe, K.; Ou, T.; Castro, N.M.; Christensen, L.N.; Cheung, L.; Jiang, X.; Yoon, D.; et al. Co-translational protein targeting facilitates centrosomal recruitment of PCNT during centrosome maturation in vertebrates. eLife 2018, 7, e34959. [Google Scholar] [CrossRef] [PubMed]
- Aznarez, I.; Nomakuchi, T.T.; Tetenbaum-Novatt, J.; Rahman, M.A.; Fregoso, O.; Rees, H.; Krainer, A.R. Mechanism of Nonsense-Mediated mRNA Decay Stimulation by Splicing Factor SRSF1. Cell Rep. 2018, 23, 2186–2198. [Google Scholar] [CrossRef] [Green Version]
- Fukumura, K.; Inoue, K.; Mayeda, A. Splicing activator RNPS1 suppresses errors in pre-mRNA splicing: A key factor for mRNA quality control. Biochem. Biophys. Res. Commun. 2018, 496, 921–926. [Google Scholar] [CrossRef]
- Kimball, C.; Powers, K.; Dustin, J.; Poirier, V.; Pellettieri, J. The exon junction complex is required for stem and progenitor cell maintenance in planarians. Dev. Biol. 2020, 457, 119–127. [Google Scholar] [CrossRef]
- Mao, H.Q.; McMahon, J.J.; Tsai, Y.H.; Wang, Z.F.; Silver, D.L. Haploinsufficiency for Core Exon Junction Complex Components Disrupts Embryonic Neurogenesis and Causes p53-Mediated Microcephaly. PLoS Genet. 2016, 12, e1006282. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, C.J.; McMahon, J.J.; Serdar, L.D.; Silver, D.L. Dosage-dependent requirements of Magoh for cortical interneuron generation and survival. Development 2020, 147, dev182295. [Google Scholar] [CrossRef]
- Pilaz, L.J.; McMahon, J.J.; Miller, E.E.; Lennox, A.L.; Suzuki, A.; Salmon, E.; Silver, D.L. Prolonged Mitosis of Neural Progenitors Alters Cell Fate in the Developing Brain. Neuron 2016, 89, 83–99. [Google Scholar] [CrossRef] [Green Version]
- Silver, D.L.; Leeds, K.E.; Hwang, H.W.; Miller, E.E.; Pavan, W.J. The EJC component Magoh regulates proliferation and expansion of neural crest-derived melanocytes. Dev. Biol. 2013, 375, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Volodarsky, M.; Lichtig, H.; Leibson, T.; Sadaka, Y.; Kadir, R.; Perez, Y.; Liani-Leibson, K.; Gradstein, L.; Shaco-Levy, R.; Shorer, Z.; et al. CDC174, a novel component of the exon junction complex whose mutation underlies a syndrome of hypotonia and psychomotor developmental delay. Hum. Mol. Genet. 2015, 24, 6485–6491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.Q.; Pilaz, L.J.; McMahon, J.J.; Golzio, C.; Wu, D.W.; Shi, L.; Katsanis, N.; Silver, D.L. Rbm8a Haploinsufficiency Disrupts Embryonic Cortical Development Resulting in Microcephaly. J. Neurosci. 2015, 35, 7003–7018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, T.-W.; Lu, C.-C.; Su, C.-H.; Wu, P.-Y.; Easwvaran, S.; Lee, C.-C.; Kuo, H.-C.; Hung, K.-Y.; Lee, K.-M.; Tsai, C.-Y. The RNA Processing Factor Y14 Participates in DNA Damage Response and Repair. iScience 2019, 13, 402–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Shen, B.Q.; Zhang, D.D.; Wang, Y.Y.; Tang, Z.M.; Ni, N.; Jin, X.L.; Luo, M.; Sun, H.; Gu, P. miR-29a regulates the proliferation and differentiation of retinal progenitors by targeting Rbm8a. Oncotarget 2017, 8, 31993–32008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, C.H.; Paolantoni, C.; Bawankar, P.; Tang, Z.; Brown, S.; Roignant, J.Y.; Treisman, J.E. An exon junction complex-independent function of Barentsz in neuromuscular synapse growth. EMBO Rep. 2022, 23, e53231. [Google Scholar] [CrossRef]
- Zou, D.H.; Li, R.J.; Huang, X.H.; Chen, G.Y.; Liu, Y.; Meng, Y.S.; Wang, Y.M.; Wu, Y.; Mao, Y.W. Identification of molecular correlations of RBM8A with autophagy in Alzheimer’s disease. Aging-Us 2019, 11, 11673–11685. [Google Scholar] [CrossRef]
- Kawano, T.; Kataoka, N.; Dreyfuss, G.; Sakamoto, H. Ce-Y14 and MAG-1, components of the exon–exon junction complex, are required for embryogenesis and germline sexual switching in Caenorhabditis elegans. Mech. Dev. 2004, 121, 27–35. [Google Scholar] [CrossRef]
- Roignant, J.Y.; Treisman, J.E. Exon junction complex subunits are required to splice Drosophila MAP kinase, a large heterochromatic gene. Cell 2010, 143, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Li, Y.; Liu, A.; Li, R.; Su, Y.; Du, J.; Li, C.; Zhu, A.J. The exon junction complex regulates the splicing of cell polarity gene dlg1 to control Wingless signaling in development. eLife 2016, 5, e17200. [Google Scholar] [CrossRef] [Green Version]
- Pilaz, L.J.; Lennox, A.L.; Rouanet, J.P.; Silver, D.L. Dynamic mRNA Transport and Local Translation in Radial Glial Progenitors of the Developing Brain. Curr. Biol. 2016, 26, 3383–3392. [Google Scholar] [CrossRef] [Green Version]
- Farris, S.; Lewandowski, G.; Cox, C.D.; Steward, O. Selective Localization of Arc mRNA in Dendrites Involves Activity- and Translation-Dependent mRNA Degradation. J. Neurosci. 2014, 34, 4481–4493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker-Haliski, M.L.; Pastuzyn, E.D.; Keefe, K.A. Expression of the core exon-junction complex factor eukaryotic initiation factor 4A3 is increased during spatial exploration and striatally-mediated learning. Neuroscience 2012, 226, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notaras, M.; Allen, M.; Longo, F.; Volk, N.; Toth, M.; Jeon, N.L.; Klann, E.; Colak, D. UPF2 leads to degradation of dendritically targeted mRNAs to regulate synaptic plasticity and cognitive function. Mol. Psychiatry 2020, 25, 3360–3379. [Google Scholar] [CrossRef] [PubMed]
- Son, H.G.; Lee, S.-J.V. Longevity regulation by NMD-mediated mRNA quality control. BMB Rep. 2017, 50, 160. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.-K.; Huang, L.; Gudikote, J.P.; Chang, Y.-F.; Imam, J.S.; MacLean II, J.A.; Wilkinson, M.F. An alternative branch of the nonsense-mediated decay pathway. EMBO J. 2007, 26, 1820–1830. [Google Scholar] [CrossRef] [Green Version]
- Metze, S.; Herzog, V.A.; Ruepp, M.-D.; Mühlemann, O. Comparison of EJC-enhanced and EJC-independent NMD in human cells reveals two partially redundant degradation pathways. RNA 2013, 19, 1432–1448. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.K.; Bhalla, A.D.; Le Hir, H.; Nguyen, L.S.; Huang, L.; Gecz, J.; Wilkinson, M.F. A UPF3-mediated regulatory switch that maintains RNA surveillance. Nat. Struct Mol. Biol. 2009, 16, 747–753. [Google Scholar] [CrossRef]
- Domingo, D.; Nawaz, U.; Corbett, M.; Espinoza, J.L.; Tatton-Brown, K.; Coman, D.; Wilkinson, M.F.; Gecz, J.; Jolly, L.A. A synonymous UPF3B variant causing a speech disorder implicates NMD as a regulator of neurodevelopmental disorder gene networks. Hum. Mol. Genet. 2020, 29, 2568–2578. [Google Scholar] [CrossRef]
- Long, A.A.; Mahapatra, C.T.; Woodruff, E.A.; Rohrbough, J.; Leung, H.-T.; Shino, S.; An, L.; Doerge, R.W.; Metzstein, M.M.; Pak, W.L. The nonsense-mediated decay pathway maintains synapse architecture and synaptic vesicle cycle efficacy. J. Cell Sci. 2010, 123, 3303–3315. [Google Scholar] [CrossRef] [Green Version]
- Guerra, G.M.; May, D.; Kroll, T.; Koch, P.; Groth, M.; Wang, Z.-Q.; Li, T.-L.; Grigaravičius, P. Cell Type-Specific Role of RNA Nuclease SMG6 in Neurogenesis. Cells 2021, 10, 3365. [Google Scholar] [CrossRef]
- Lou, C.-H.; Dumdie, J.; Goetz, A.; Shum, E.Y.; Brafman, D.; Liao, X.; Mora-Castilla, S.; Ramaiah, M.; Cook-Andersen, H.; Laurent, L.; et al. Nonsense-Mediated RNA Decay Influences Human Embryonic Stem Cell Fate. Stem Cell Rep. 2016, 6, 844–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huth, M.; Santini, L.; Galimberti, E.; Ramesmayer, J.; Titz-Teixeira, F.; Sehlke, R.; Oberhuemer, M.; Stummer, S.; Herzog, V.; Garmhausen, M.; et al. NMD is required for timely cell fate transitions by fine-tuning gene expression and regulating translation. Genes Dev. 2022. [Google Scholar] [CrossRef] [PubMed]
- McIlwain, D.R.; Pan, Q.; Reilly, P.T.; Elia, A.J.; McCracken, S.; Wakeham, A.C.; Itie-Youten, A.; Blencowe, B.J.; Mak, T.W. Smg1 is required for embryogenesis and regulates diverse genes via alternative splicing coupled to nonsense-mediated mRNA decay. Proc. Natl. Acad. Sci. USA 2010, 107, 12186–12191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, J.; Tang, C.; Yuan, S.; Porse, B.T.; Yan, W. UPF2, a nonsense-mediated mRNA decay factor, is required for prepubertal Sertoli cell development and male fertility by ensuring fidelity of the transcriptome. Development 2015, 142, 352–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoren, L.A.; Norgaard, G.A.; Weischenfeldt, J.; Waage, J.; Jakobsen, J.S.; Damgaard, I.; Bergstrom, F.C.; Blom, A.M.; Borup, R.; Bisgaard, H.C.; et al. UPF2 is a critical regulator of liver development, function and regeneration. PLoS ONE 2010, 5, e11650. [Google Scholar] [CrossRef]
- Jolly, L.A.; Homan, C.C.; Jacob, R.; Barry, S.; Gecz, J. The UPF3B gene, implicated in intellectual disability, autism, ADHD and childhood onset schizophrenia regulates neural progenitor cell behaviour and neuronal outgrowth. Hum. Mol. Genet. 2013, 22, 4673–4687. [Google Scholar] [CrossRef] [Green Version]
- Addington, A.M.; Gauthier, J.; Piton, A.; Hamdan, F.F.; Raymond, A.; Gogtay, N.; Miller, R.; Tossell, J.; Bakalar, J.; Germain, G.; et al. A novel frameshift mutation in UPF3B identified in brothers affected with childhood onset schizophrenia and autism spectrum disorders. Mol. Psychiatry 2011, 16, 238–239. [Google Scholar] [CrossRef]
- Alrahbeni, T.; Sartor, F.; Anderson, J.; Miedzybrodzka, Z.; McCaig, C.; Muller, B. Full UPF3B function is critical for neuronal differentiation of neural stem cells. Mol. Brain 2015, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Shum, E.Y.; Jones, S.H.; Lou, C.H.; Dumdie, J.; Kim, H.; Roberts, A.J.; Jolly, L.A.; Espinoza, J.L.; Skarbrevik, D.M.; et al. A Upf3b-mutant mouse model with behavioral and neurogenesis defects. Mol. Psychiatry 2018, 23, 1773–1786. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.; Jones, S.H.; Lake, B.B.; Dumdie, J.N.; Shum, E.Y.; Zhang, L.; Chen, S.; Sohni, A.; Pandya, S.; Gallo, R.L.; et al. The role of the NMD factor UPF3B in olfactory sensory neurons. eLife 2020, 9, e57525. [Google Scholar] [CrossRef]
- Johnson, J.L.; Stoica, L.; Liu, Y.W.; Zhu, P.J.; Bhattacharya, A.; Buffington, S.A.; Huq, R.; Eissa, N.T.; Larsson, O.; Porse, B.T.; et al. Inhibition of Upf2-Dependent Nonsense-Mediated Decay Leads to Behavioral and Neurophysiological Abnormalities by Activating the Immune Response. Neuron 2019, 104, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Kamelgarn, M.; Chen, J.; Kuang, L.; Jin, H.; Kasarskis, E.J.; Zhu, H. ALS mutations of FUS suppress protein translation and disrupt the regulation of nonsense-mediated decay. Proc. Natl. Acad. Sci. USA 2018, 115, E11904–E11913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontaine, K.A.; Leon, K.E.; Khalid, M.M.; Tomar, S.; Jimenez-Morales, D.; Dunlap, M.; Kaye, J.A.; Shah, P.S.; Finkbeiner, S.; Krogan, N.J. The cellular NMD pathway restricts Zika virus infection and is targeted by the viral capsid protein. MBio 2018, 9, e02126-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruno, I.G.; Karam, R.; Huang, L.L.; Bhardwaj, A.; Lou, C.H.; Shum, E.Y.; Song, H.W.; Corbett, M.A.; Gifford, W.D.; Gecz, J.; et al. Identification of a MicroRNA that Activates Gene Expression by Repressing Nonsense-Mediated RNA Decay. Mol. Cell 2011, 42, 500–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, T.; Zhang, C.; Wang, H.; Lay, K.; Fak, J.; Noebels, J.L.; Darnell, R.B. NOVA-dependent regulation of cryptic NMD exons controls synaptic protein levels after seizure. eLife 2013, 2, e00178. [Google Scholar] [CrossRef]
- Gonatopoulos-Pournatzis, T.; Niibori, R.; Salter, E.W.; Weatheritt, R.J.; Tsang, B.; Farhangmehr, S.; Liang, X.Y.; Braunschweig, U.; Roth, J.; Zhang, S.; et al. Autism-Misregulated eIF4G Microexons Control Synaptic Translation and Higher Order Cognitive Functions. Mol. Cell 2020, 77, 1176–1192. [Google Scholar] [CrossRef]
- Nguyen, L.S.; Kim, H.G.; Rosenfeld, J.A.; Shen, Y.P.; Gusella, J.F.; Lacassie, Y.; Layman, L.C.; Shaffer, L.G.; Gecz, J. Contribution of copy number variants involving nonsense-mediated mRNA decay pathway genes to neuro-developmental disorders. Hum. Mol. Genet. 2013, 22, 1816–1825. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.N.; Chan, C.-H.; Pearce, D.A. The role of nonsense-mediated decay in neuronal ceroid lipofuscinosis. Hum. Mol. Genet. 2013, 22, 2723–2734. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.-Q.; Shen, W.; Macdonald, R.L. Two molecular pathways (NMD and ERAD) contribute to a genetic epilepsy associated with the GABAA receptor GABRA1 PTC mutation, 975delC, S326fs328X. J. Neurosci. 2009, 29, 2833–2844. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Low, A.; Damle, S.S.; Keenan, M.M.; Kuntz, S.; Murray, S.F.; Monia, B.P.; Guo, S. Antisense suppression of the nonsense mediated decay factor Upf3b as a potential treatment for diseases caused by nonsense mutations. Genome Biol. 2018, 19, 4. [Google Scholar] [CrossRef] [Green Version]
- Tewes, A.-C.; Rall, K.K.; Römer, T.; Hucke, J.; Kapczuk, K.; Brucker, S.; Wieacker, P.; Ledig, S. Variations in RBM8A and TBX6 are associated with disorders of the müllerian ducts. Fertil. Steril. 2015, 103, 1313–1318. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, J.; Hu, B.; Qin, G.; Liang, R.; Lin, Y.; Wei, J.; Qian, Z.; Zou, D. RBM8A promotes growth and invasion through the Notch/STAT3 pathway in glioblastoma. Front Oncol. 2021, 11, 736941. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Liang, R.; Qiu, Y.; Lv, Y.; Zhang, J.; Qin, G.; Yuan, C.; Liu, Z.; Li, Y.; Zou, D.; et al. Expression and gene regulation network of RBM8A in hepatocellular carcinoma based on data mining. Aging 2019, 11, 423–447. [Google Scholar] [CrossRef] [PubMed]
- Liang, R.; Lin, Y.; Ye, J.-Z.; Yan, X.-X.; Liu, Z.-H.; Li, Y.-Q.; Luo, X.-L.; Ye, H.-H. High expression of RBM8A predicts poor patient prognosis and promotes tumor progression in hepatocellular carcinoma. Oncol. Rep. 2017, 37, 2167–2176. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-J.; Choi, J.-J.; Kim, W.Y.; Choi, C.H.; Lee, J.-W.; Bae, D.-S.; Son, D.-S.; Kim, J.; Park, B.K.; Ahn, G.; et al. Gene expression profiling for the prediction of lymph node metastasis in patients with cervical cancer. Cancer Sci. 2008, 99, 31–38. [Google Scholar] [CrossRef]
- Meznad, K.; Paget-Bailly, P.; Jacquin, E.; Peigney, A.; Aubin, F.; Guittaut, M.; Mougin, C.; Prétet, J.-L.; Baguet, A. The exon junction complex core factor eIF4A3 is a key regulator of HPV16 gene expression. Biosci. Rep. 2021, 41, BSR20203488. [Google Scholar] [CrossRef]
- Lin, Y.; Liang, R.; Mao, Y.W.; Ye, J.Z.; Mai, R.Y.; Gao, X.; Liu, Z.Y.; Wainwright, T.; Li, Q.; Luo, M.; et al. Comprehensive analysis of biological networks and the eukaryotic initiation factor 4A-3 gene as pivotal in hepatocellular carcinoma. J. Cell. Biochem. 2020, 121, 4094–4107. [Google Scholar] [CrossRef]
- Bernier, R.; Steinman, K.J.; Reilly, B.; Wallace, A.S.; Sherr, E.H.; Pojman, N.; Mefford, H.C.; Gerdts, J.; Earl, R.; Hanson, E.; et al. Clinical phenotype of the recurrent 1q21.1 copy-number variant. Genet. Med. 2016, 18, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Mefford, H.C.; Sharp, A.J.; Baker, C.; Itsara, A.; Jiang, Z.; Buysse, K.; Huang, S.; Maloney, V.K.; Crolla, J.A.; Baralle, D. Recurrent rearrangements of chromosome 1q21. 1 and variable pediatric phenotypes. N. Engl. J. Med. 2008, 359, 1685–1699. [Google Scholar] [CrossRef] [Green Version]
- Brunetti-Pierri, N.; Berg, J.S.; Scaglia, F.; Belmont, J.; Bacino, C.A.; Sahoo, T.; Lalani, S.R.; Graham, B.; Lee, B.; Shinawi, M. Recurrent reciprocal 1q21. 1 deletions and duplications associated with microcephaly or macrocephaly and developmental and behavioral abnormalities. Nat. Genet. 2008, 40, 1466–1471. [Google Scholar] [CrossRef]
- Carvill, G.L.; Helbig, K.L.; Myers, C.T.; Scala, M.; Huether, R.; Lewis, S.; Kruer, T.N.; Guida, B.S.; Bakhtiari, S.; Sebe, J.; et al. Damaging de novo missense variants in EEF1A2 lead to a developmental and degenerative epileptic-dyskinetic encephalopathy. Hum. Mutat. 2020, 41, 1263–1279. [Google Scholar] [CrossRef] [PubMed]
- Boussion, S.; Escande, F.; Jourdain, A.S.; Smol, T.; Brunelle, P.; Duhamel, C.; Alembik, Y.; Attié-Bitach, T.; Baujat, G.; Bazin, A. TAR syndrome: Clinical and molecular characterization of a cohort of 26 patients and description of novel noncoding variants of RBM8A. Hum. Mutat. 2020, 41, 1220–1225. [Google Scholar] [CrossRef] [PubMed]
- Kumar, C.; Sharma, D.; Pandita, A.; Bhalerao, S. Thrombocytopenia absent radius syndrome with Tetralogy of Fallot: A rare association. Int. Med. Case Rep. J. 2015, 8, 81. [Google Scholar] [PubMed] [Green Version]
- Gamba, B.F.; Zechi-Ceide, R.M.; Kokitsu-Nakata, N.M.; Vendrarnini-Pittoli, S.; Rosenberg, C.; Santos, A.C.K.; Ribeiro-Bicudo, L.; Richieri-Costa, A. Interstitial 1q21.1 Microdeletion Is Associated with Severe Skeletal Anomalies, Dysmorphic Face and Moderate Intellectual Disability. Mol. Syndromol. 2016, 7, 344–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.-H.; Liao, W.-J.; Ke, W.-C.; Yang, R.-B.; Tarn, W.-Y. The Y14-p53 regulatory circuit in megakaryocyte differentiation and thrombocytopenia. iScience 2021, 24, 103368. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, J.P.; Sheehan, K.B.; Bennett Jr, P.E.; Boswell, R.E. Mago Nashi, Tsunagi/Y14, and Ranshi form a complex that influences oocyte differentiation in Drosophila melanogaster. Dev. Biol. 2010, 339, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.-P.; Zhang, P.; Zhang, W.-N.; Huang, D.-K.; Jia, X.-M.; Gui, L.; Liu, M.; Shen, J.-J. Schistosoma japonicum: Tsunagi/Y14 protein plays a critical role in the development of the reproductive organs and eggs. Exp. Parasitol. 2013, 135, 430–436. [Google Scholar] [CrossRef]
- Miller, E.E.; Kobayashi, G.S.; Musso, C.M.; Allen, M.; Ishiy, F.A.A.; de Caires, L.C.; Goulart, E.; Griesi-Oliveira, K.; Zechi-Ceide, R.M.; Richieri-Costa, A.; et al. EIF4A3 deficient human iPSCs and mouse models demonstrate neural crest defects that underlie Richieri-Costa-Pereira syndrome. Hum. Mol. Genet. 2017, 26, 2177–2191. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Snider, L.; Jagannathan, S.; Tawil, R.; van der Maarel, S.M.; Tapscott, S.J.; Bradley, R.K. A feedback loop between nonsense-mediated decay and the retrogene DUX4 in facioscapulohumeral muscular dystrophy. eLife 2015, 4, e04996. [Google Scholar] [CrossRef]


{kind=link}
| EJC Component | Common Aliases | Model Organism | Reference |
|---|---|---|---|
| RBM8A | Y14 | Homo sapiens | [37] |
| Tsunagi | Drosophila melanogaster | [38] | |
| BOV 1 | Saccharomyces cerevisiae | [39] | |
| MAGOH | Mago-Nashi | Drosophila melanogaster | [40] |
| EIF4A3 | DEAD Box Protein | Mus muculus | [13] |
| NMP 265 | Homo sapiens | [41] | |
| DDX48 | Homo sapiens | [42] | |
| MLN51 | CASC3 | Homo sapiens | [43] |
| BTZ | Homo sapiens |
| EJC Component | Implicated Human Disorders | Tools Employed in Discovery | References |
|---|---|---|---|
| RBM8A | Thrombocytopenia-Absent Radius Syndrome (TAR) | Genetics analysis | [80,81] |
| Autism Spectrum Disorder | Mouse RNAi/Over-expression | [76] | |
| Mayer–Rokitansky–Küster–Hauser syndrome | Genetic variation analyses | [151] | |
| West Nile Virus | RNAi | [143] | |
| Glioblastoma | RNAseq Analysis | [152] | |
| Hepatocellular Carcinoma | Gene Expression Profiling Interactive Analysis | [153,154] | |
| Cervical Cancer | Microarray Gene Analysis | [155] | |
| Alzheimer’s Disease | GSEA and Identification of D.E.G.’s | [116] | |
| MAGOH 1 | |||
| EIF4A3 | Richieri-Costa-Pereira syndrome (RCP) | Genetics analysis | [82] |
| Anogenital Cancers | Gene expression analysis and RT-PCR | [156] | |
| Hepatocellular Carcinoma | Gene Expression Profiling Interactive Analysis | [157] | |
| MLN51 | HER2+ Breast Cancer | Immunofluorescence analysis | [63] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Asthana, S.; Martin, H.; Rupkey, J.; Patel, S.; Yoon, J.; Keegan, A.; Mao, Y. The Physiological Roles of the Exon Junction Complex in Development and Diseases. Cells 2022, 11, 1192. https://doi.org/10.3390/cells11071192
Asthana S, Martin H, Rupkey J, Patel S, Yoon J, Keegan A, Mao Y. The Physiological Roles of the Exon Junction Complex in Development and Diseases. Cells. 2022; 11(7):1192. https://doi.org/10.3390/cells11071192
Chicago/Turabian StyleAsthana, Shravan, Hannah Martin, Julian Rupkey, Shray Patel, Joy Yoon, Abiageal Keegan, and Yingwei Mao. 2022. "The Physiological Roles of the Exon Junction Complex in Development and Diseases" Cells 11, no. 7: 1192. https://doi.org/10.3390/cells11071192
APA StyleAsthana, S., Martin, H., Rupkey, J., Patel, S., Yoon, J., Keegan, A., & Mao, Y. (2022). The Physiological Roles of the Exon Junction Complex in Development and Diseases. Cells, 11(7), 1192. https://doi.org/10.3390/cells11071192