
The Cell Biology of Heterochromatin

{kind=link}
{kind=link}
Abstract
:1. Introduction
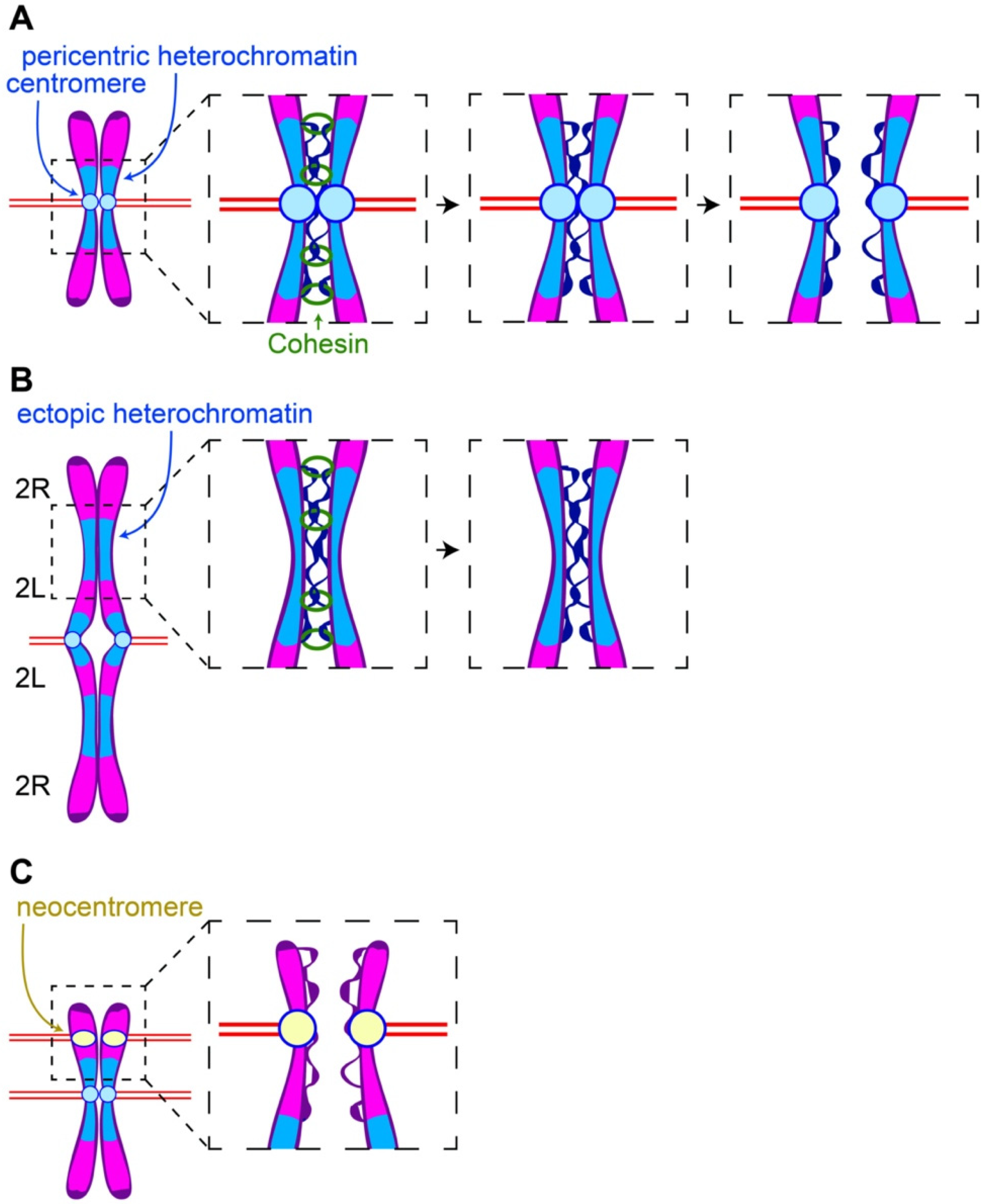
2. Pericentric Heterochromatin and Sister Chromatid Separation
2.1. Delayed Cohesin Removal at Ectopic Heterochromatic Results in Delayed Sister Separation at These Sites
2.2. Pericentric Heterochromatin and Centromeres Balance Binding and Pulling Forces on the Sister Chromatids
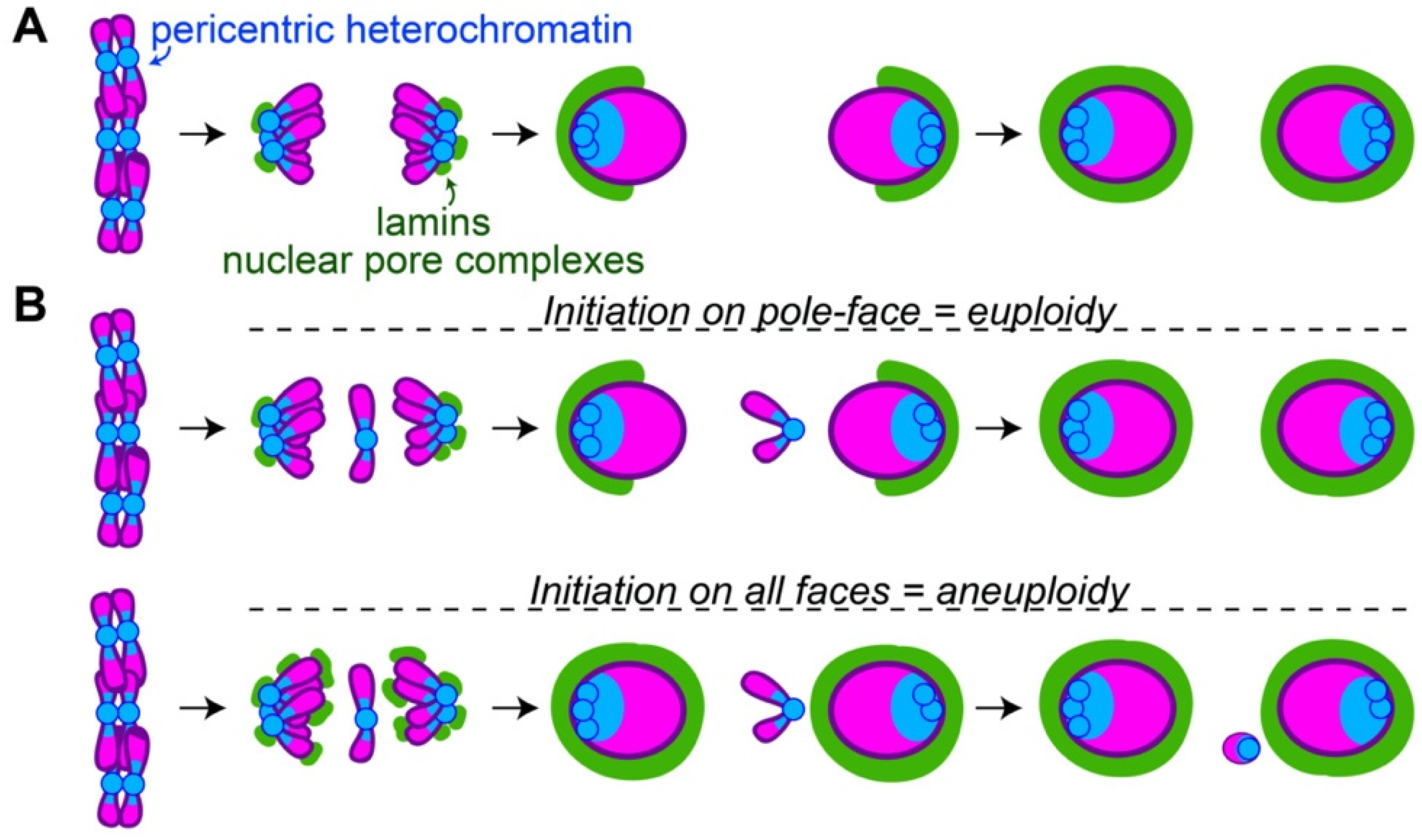
3. Heterochromatin and Nuclear Envelope Assembly
3.1. Initiation of Nuclear Envelope Assembly Often Occurs in Areas of High Heterochromatin Abundance
3.2. Pole-to-Midzone Nuclear Envelope Assembly Provides Additional Time for Segregating Chromosomes to Form a Single Nucleus
3.3. Variations of Mitosis Provide Additional Insights into the Functions of Heterochromatin–Nuclear Envelope Interactions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heitz, E. Das heterochromatin der moose. Jahrb. Wiss. Bot. 1928, 69, 762–818. [Google Scholar]
- Janssen, A.; Colmenares, S.U.; Karpen, G.H. Heterochromatin: Guardian of the Genome. Annu. Rev. Cell Dev. Biol. 2018, 34, 265–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allshire, R.C.; Madhani, H.D. Ten principles of heterochromatin formation and function. Nat. Rev. Mol. Cell Biol. 2018, 19, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Miga, K.H.; Koren, S.; Rhie, A.; Vollger, M.R.; Gershman, A.; Bzikadze, A.; Brooks, S.; Howe, E.; Porubsky, D.; Logsdon, G.A.; et al. Telomere-to-telomere assembly of a complete human X chromosome. Nature 2020, 585, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Yunis, J.J.; Yasmineh, W.G. Heterochromatin, satellite DNA, and cell function. Structural DNA of eucaryotes may support and protect genes and aid in speciation. Science 1971, 174, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.Y.; Crest, J.; Schubiger, G. Genetic interactions between Cdk1-CyclinB and the Separase complex in Drosophila. Development 2005, 132, 1875–1884. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Liu, H. Functioning mechanisms of Shugoshin-1 in centromeric cohesion during mitosis. Essays Biochem. 2020, 64, 289–297. [Google Scholar] [CrossRef]
- Kawashima, S.A.; Yamagishi, Y.; Honda, T.; Ishiguro, K.; Watanabe, Y. Phosphorylation of H2A by Bub1 prevents chromosomal instability through localizing shugoshin. Science 2010, 327, 172–177. [Google Scholar] [CrossRef]
- Luo, S.; Tong, L. Structure and Function of the Separase-Securin Complex. Subcell Biochem. 2021, 96, 217–232. [Google Scholar] [CrossRef]
- Novitski, E.; Grace, D.; Strommen, C. The entire compound autosomes of Drosophila melanogaster. Genetics 1981, 98, 257–273. [Google Scholar] [CrossRef]
- Martins, T.; Kotadia, S.; Malmanche, N.; Sunkel, C.E.; Sullivan, W. Strategies for outcrossing and genetic manipulation of Drosophila compound autosome stocks. G3 2013, 3, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, R.A.; Kotadia, S.; Tavares, A.; Mirkovic, M.; Bowlin, K.; Eichinger, C.S.; Nasmyth, K.; Sullivan, W. Centromere-independent accumulation of cohesin at ectopic heterochromatin sites induces chromosome stretching during anaphase. PLoS Biol. 2014, 12, e1001962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haarhuis, J.H.; Elbatsh, A.M.; Rowland, B.D. Cohesin and its regulation: On the logic of X-shaped chromosomes. Dev Cell 2014, 31, 7–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finardi, A.; Massari, L.F.; Visintin, R. Anaphase Bridges: Not All Natural Fibers Are Healthy. Genes 2020, 11, 902. [Google Scholar] [CrossRef]
- Farcas, A.M.; Uluocak, P.; Helmhart, W.; Nasmyth, K. Cohesin’s concatenation of sister DNAs maintains their intertwining. Mol. Cell 2011, 44, 97–107. [Google Scholar] [CrossRef]
- Mariezcurrena, A.; Uhlmann, F. Observation of DNA intertwining along authentic budding yeast chromosomes. Genes Dev 2017, 31, 2151–2161. [Google Scholar] [CrossRef] [Green Version]
- Rhoades, M.M.; McClintock, B. The cytogenetics of maize. Bot. Rev. 1935, 1, 292–325. [Google Scholar] [CrossRef]
- Fluminhan, A.; Kameya, T. Behaviour of chromosomes in anaphase cells in embryogenic callus cultures of maize (Zea mays L.). Theor. Appl. Genet. 1996, 92, 982–990. [Google Scholar] [CrossRef]
- Rhoades, M.M.; Vilkomerson, H. On the Anaphase Movement of Chromosomes. Proc. Natl. Acad. Sci. USA 1942, 28, 433–436. [Google Scholar] [CrossRef] [Green Version]
- Dawe, R.K.; Lowry, E.G.; Gent, J.I.; Stitzer, M.C.; Swentowsky, K.W.; Higgins, D.M.; Ross-Ibarra, J.; Wallace, J.G.; Kanizay, L.B.; Alabady, M.; et al. A Kinesin-14 Motor Activates Neocentromeres to Promote Meiotic Drive in Maize. Cell 2018, 173, 839–850.e18. [Google Scholar] [CrossRef] [Green Version]
- Swentowsky, K.W.; Gent, J.I.; Lowry, E.G.; Schubert, V.; Ran, X.; Tseng, K.F.; Harkess, A.E.; Qiu, W.; Dawe, R.K. Distinct kinesin motors drive two types of maize neocentromeres. Genes Dev. 2020, 34, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, M.M. Preferential Segregation in Maize. Genetics. 1942, 27, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Pineda, M.; Valente, L.P.; Dumont, M.; Mata, J.F.; Fachinetti, D.; Jansen, L.E.T. Induction of spontaneous human neocentromere formation and long-term maturation. J. Cell Biol. 2021, 220, e202007210. [Google Scholar] [CrossRef]
- Heun, P.; Erhardt, S.; Blower, M.D.; Weiss, S.; Skora, A.D.; Karpen, G.H. Mislocalization of the Drosophila centromere-specific histone CID promotes formation of functional ectopic kinetochores. Dev. Cell 2006, 10, 303–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeBose-Scarlett, E.M.; Sullivan, B.A. Genomic and Epigenetic Foundations of Neocentromere Formation. Annu. Rev. Genet. 2021, 55, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Burrack, L.S.; Berman, J. Neocentromeres and epigenetically inherited features of centromeres. Chromosome Res. 2012, 20, 607–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Steensel, B.; Belmont, A.S. Lamina-Associated Domains: Links with Chromosome Architecture, Heterochromatin, and Gene Repression. Cell 2017, 169, 780–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, P.R.; Nietlispach, D.; Mott, H.R.; Callaghan, J.; Bannister, A.; Kouzarides, T.; Murzin, A.G.; Murzina, N.V.; Laue, E.D. Structure of the HP1 chromodomain bound to histone H3 methylated at lysine 9. Nature 2002, 416, 103–107. [Google Scholar] [CrossRef]
- Ye, Q.; Worman, H.J. Interaction between an integral protein of the nuclear envelope inner membrane and human chromodomain proteins homologous to Drosophila HP1. J. Biol. Chem. 1996, 271, 14653–14656. [Google Scholar] [CrossRef] [Green Version]
- Ye, Q.; Callebaut, I.; Pezhman, A.; Courvalin, J.C.; Worman, H.J. Domain-specific interactions of human HP1-type chromodomain proteins and inner nuclear membrane protein LBR. J. Biol. Chem. 1997, 272, 14983–14989. [Google Scholar] [CrossRef] [Green Version]
- Hirano, Y.; Asakawa, H.; Sakuno, T.; Haraguchi, T.; Hiraoka, Y. Nuclear Envelope Proteins Modulating the Heterochromatin Formation and Functions in Fission Yeast. Cells 2020, 9, 1908. [Google Scholar] [CrossRef] [PubMed]
- Probst, A.V.; Almouzni, G. Heterochromatin establishment in the context of genome-wide epigenetic reprogramming. Trends Genet. 2011, 27, 177–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, K.; Tasaka, H.; Dotsu, M. Molecular behavior in living mitotic cells of human centromere heterochromatin protein HPLalpha ectopically expressed as a fusion to red fluorescent protein. Cell Struct. Funct. 2001, 26, 705–718. [Google Scholar] [CrossRef] [Green Version]
- Warecki, B.; Sullivan, W. Micronuclei Formation Is Prevented by Aurora B-Mediated Exclusion of HP1a from Late-Segregating Chromatin in Drosophila. Genetics 2018, 210, 171–187. [Google Scholar] [CrossRef] [Green Version]
- Dechat, T.; Gajewski, A.; Korbei, B.; Gerlich, D.; Daigle, N.; Haraguchi, T.; Furukawa, K.; Ellenberg, J.; Foisner, R. LAP2alpha and BAF transiently localize to telomeres and specific regions on chromatin during nuclear assembly. J. Cell Sci. 2004, 117, 6117–6128. [Google Scholar] [CrossRef] [Green Version]
- Katsani, K.R.; Karess, R.E.; Dostatni, N.; Doye, V. In vivo dynamics of Drosophila nuclear envelope components. Mol. Biol. Cell 2008, 19, 3652–3666. [Google Scholar] [CrossRef] [Green Version]
- Zullo, J.M.; Demarco, I.A.; Piqué-Regi, R.; Gaffney, D.J.; Epstein, C.B.; Spooner, C.J.; Luperchio, T.R.; Bernstein, B.E.; Pritchard, J.K.; Reddy, K.L.; et al. DNA sequence-dependent compartmentalization and silencing of chromatin at the nuclear lamina. Cell 2012, 149, 1474–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poleshko, A.; Mansfield, K.M.; Burlingame, C.C.; Andrake, M.D.; Shah, N.R.; Katz, R.A. The human protein PRR14 tethers heterochromatin to the nuclear lamina during interphase and mitotic exit. Cell Rep. 2013, 5, 292–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poleshko, A.; Smith, C.L.; Nguyen, S.C.; Sivaramakrishnan, P.; Wong, K.G.; Murray, J.I.; Lakadamyali, M.; Joyce, E.F.; Jain, R.; Epstein, J.A. H3K9me2 orchestrates inheritance of spatial positioning of peripheral heterochromatin through mitosis. eLife 2019, 8, e49278. [Google Scholar] [CrossRef]
- Kourmouli, N.; Theodoropoulos, P.A.; Dialynas, G.; Bakou, A.; Politou, A.S.; Cowell, I.G.; Singh, P.B.; Georgatos, S.D. Dynamic associations of heterochromatin protein 1 with the nuclear envelope. EMBO J. 2000, 19, 6558–6568. [Google Scholar] [CrossRef] [Green Version]
- Rasala, B.A.; Ramos, C.; Harel, A.; Forbes, D.J. Capture of AT-rich chromatin by ELYS recruits POM121 and NDC1 to initiate nuclear pore assembly. Mol. Biol. Cell 2008, 19, 3982–3996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Saldivar, G.; Fernandez, A.; Hirano, Y.; Mauro, M.; Lai, A.; Ayuso, C.; Haraguchi, T.; Hiraoka, Y.; Piano, F.; Askjaer, P. Identification of Conserved MEL-28/ELYS Domains with Essential Roles in Nuclear Assembly and Chromosome Segregation. PLoS Genet. 2016, 12, e1006131. [Google Scholar] [CrossRef] [PubMed]
- Talbert, P.B.; Henikoff, S. What makes a centromere? Exp. Cell. Res. 2020, 389, 111895. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.G.; Piano, F. MEL-28 is downstream of the Ran cycle and is required for nuclear-envelope function and chromatin maintenance. Curr. Biol. 2006, 16, 1757–1763. [Google Scholar] [CrossRef] [Green Version]
- Rasala, B.A.; Orjalo, A.V.; Shen, Z.; Briggs, S.; Forbes, D.J. ELYS is a dual nucleoporin/kinetochore protein required for nuclear pore assembly and proper cell division. Proc. Natl. Acad. Sci. USA 2006, 103, 17801–17806. [Google Scholar] [CrossRef] [Green Version]
- Franz, C.; Walczak, R.; Yavuz, S.; Santarella, R.; Gentzel, M.; Askjaer, P.; Galy, V.; Hetzer, M.; Mattaj, I.W.; Antonin, W. MEL-28/ELYS is required for the recruitment of nucleoporins to chromatin and postmitotic nuclear pore complex assembly. EMBO Rep. 2007, 8, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Ulbert, S.; Platani, M.; Boue, S.; Mattaj, I.W. Direct membrane protein-DNA interactions required early in nuclear envelope assembly. J. Cell Biol. 2006, 173, 469–476. [Google Scholar] [CrossRef] [Green Version]
- Royou, A.; Gagou, M.E.; Karess, R.; Sullivan, W. BubR1- and Polo-coated DNA tethers facilitate poleward segregation of acentric chromatids. Cell 2010, 140, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Afonso, O.; Matos, I.; Pereira, A.J.; Aguiar, P.; Lampson, M.A.; Maiato, H. Feedback control of chromosome separation by a midzone Aurora B gradient. Science 2014, 345, 332–336. [Google Scholar] [CrossRef] [Green Version]
- Bretscher, H.S.; Fox, D.T. Proliferation of Double-Strand Break-Resistant Polyploid Cells Requires Drosophila FANCD2. Dev. Cell 2016, 37, 444–457. [Google Scholar] [CrossRef] [Green Version]
- Karg, T.; Warecki, B.; Sullivan, W. Aurora B-mediated localized delays in nuclear envelope formation facilitate inclusion of late-segregating chromosome fragments. Mol. Biol. Cell 2015, 26, 2227–2241. [Google Scholar] [CrossRef] [PubMed]
- Afonso, O.; Castellani, C.M.; Cheeseman, L.P.; Ferreira, J.G.; Orr, B.; Ferreira, L.T.; Chambers, J.J.; Morais-de-Sa, E.; Maresca, T.J.; Maiato, H. Spatiotemporal control of mitotic exit during anaphase by an aurora B-Cdk1 crosstalk. eLife 2019, 8, e47646. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Kwon, M.; Mannino, M.; Yang, N.; Renda, F.; Khodjakov, A.; Pellman, D. Nuclear envelope assembly defects link mitotic errors to chromothripsis. Nature 2018, 561, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, Y.; Maekawa, H.; Asakawa, H.; Chikashige, Y.; Kojidani, T.; Osakada, H.; Matsuda, A.; Haraguchi, T. Inner nuclear membrane protein Ima1 is dispensable for intranuclear positioning of centromeres. Genes Cells 2011, 16, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Álvarez, A.; Cooper, J.P. The functionally elusive RabI chromosome configuration directly regulates nuclear membrane remodeling at mitotic onset. Cell Cycle 2017, 16, 1392–1396. [Google Scholar] [CrossRef]
- Maddox, P.S.; Oegema, K.; Desai, A.; Cheeseman, I.M. “Holo”er than thou: Chromosome segregation and kinetochore function in C. elegans. Chromosome Res. 2004, 12, 641–653. [Google Scholar] [CrossRef]
- Garrigues, J.M.; Sidoli, S.; Garcia, B.A.; Strome, S. Defining heterochromatin in C. elegans through genome-wide analysis of the heterochromatin protein 1 homolog HPL-2. Genome Res. 2015, 25, 76–88. [Google Scholar] [CrossRef] [Green Version]
- Gorjánácz, M.; Jaedicke, A.; Mattaj, I.W. What can Caenorhabditis elegans tell us about the nuclear envelope? FEBS Lett. 2007, 581, 2794–2801. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Gao, S.; Mostovoy, Y.; Kang, Y.; Zagoskin, M.; Sun, Y.; Zhang, B.; White, L.K.; Easton, A.; Nutman, T.B.; et al. Comparative genome analysis of programmed DNA elimination in nematodes. Genome Res. 2017, 27, 2001–2014. [Google Scholar] [CrossRef]
- Wang, J.; Veronezi, G.M.B.; Kang, Y.; Zagoskin, M.; O’Toole, E.T.; Davis, R.E. Comprehensive Chromosome End Remodeling during Programmed DNA Elimination. Curr. Biol. 2020, 30, 3397–3413.e3394. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Warecki, B.; Sullivan, W. The Cell Biology of Heterochromatin. Cells 2022, 11, 1247. https://doi.org/10.3390/cells11071247
Warecki B, Sullivan W. The Cell Biology of Heterochromatin. Cells. 2022; 11(7):1247. https://doi.org/10.3390/cells11071247
Chicago/Turabian StyleWarecki, Brandt, and William Sullivan. 2022. "The Cell Biology of Heterochromatin" Cells 11, no. 7: 1247. https://doi.org/10.3390/cells11071247
APA StyleWarecki, B., & Sullivan, W. (2022). The Cell Biology of Heterochromatin. Cells, 11(7), 1247. https://doi.org/10.3390/cells11071247