
Depletion of the Origin Recognition Complex Subunits Delays Aging in Budding Yeast

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Determination of the Growth Rate
2.3. Determination of the Mean Doubling Time
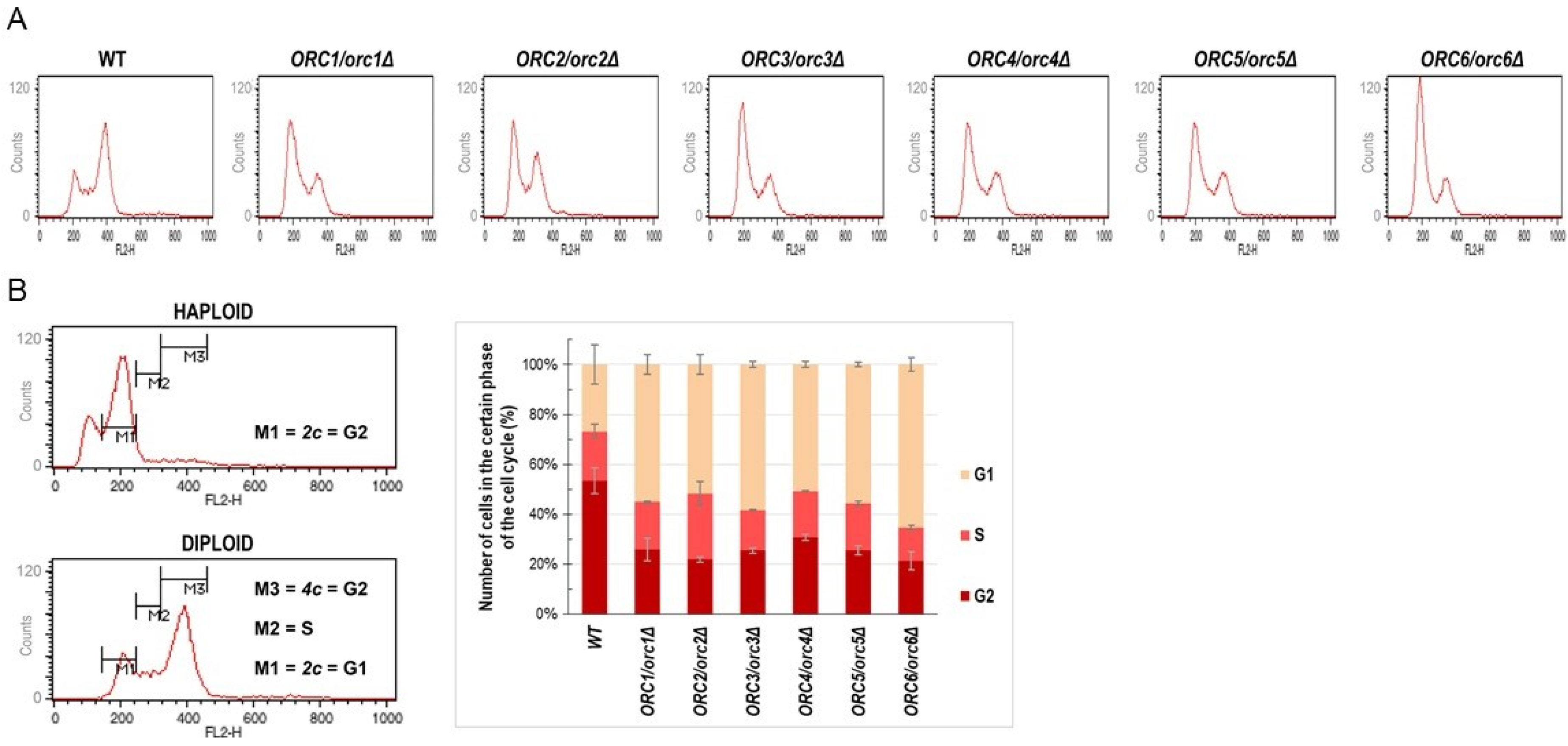
2.4. Cell Cycle Analysis
2.5. Measurement of Cell Metabolic Activity
2.6. Detection of Rad52-YFP Foci via Fluorescence Microscopy
2.7. Sporulation Efficiency Assay
2.8. Tetrad Dissection
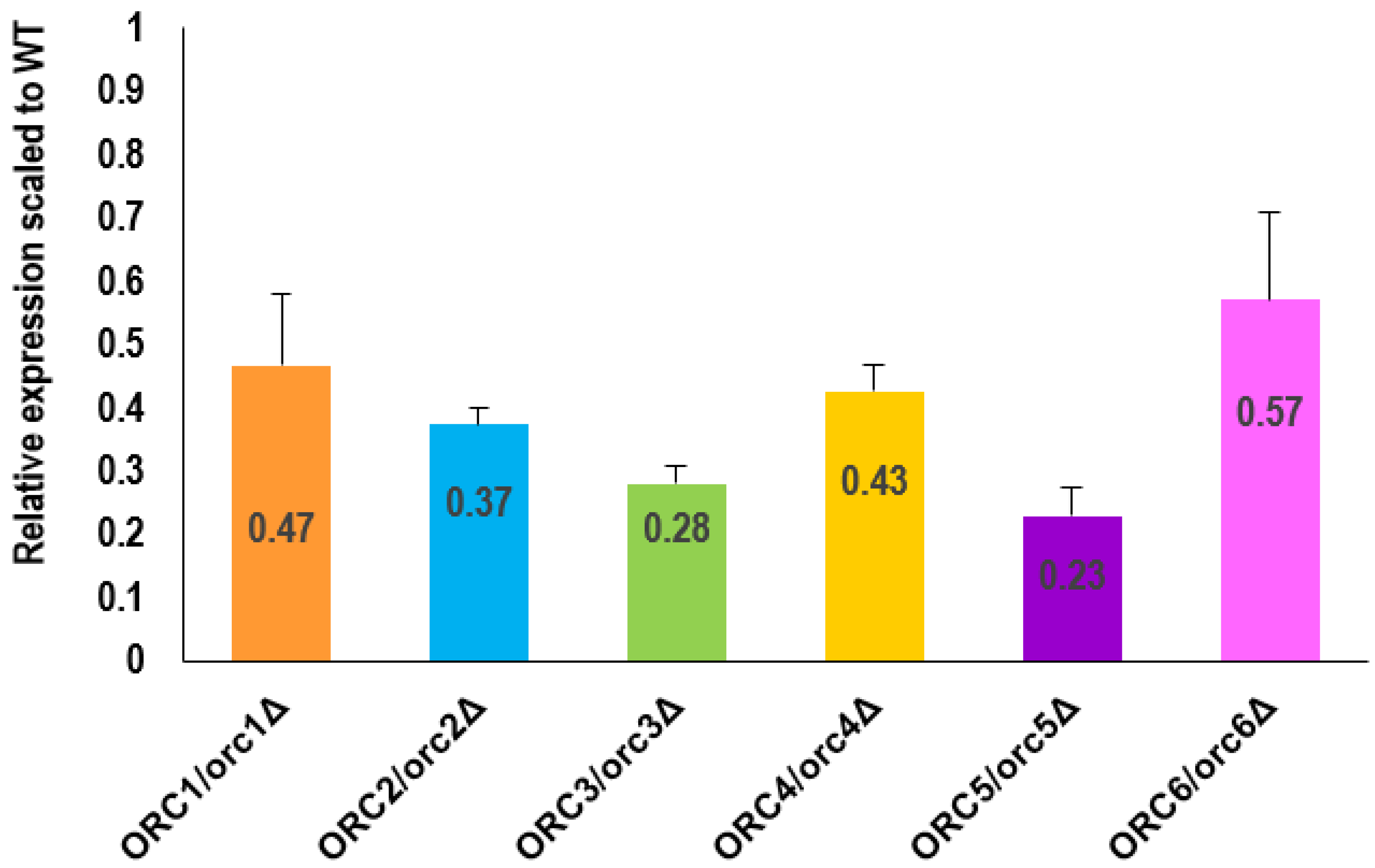
2.9. RNA Isolation and RT-qPCR
2.10. Determination of Budding Lifespan
2.11. Determination of the Total Lifespan
2.12. Chronological Lifespan (CLS) Assays
2.13. Cell Viability
2.14. Raman Spectroscopy
2.15. Statistical Analysis
3. Results
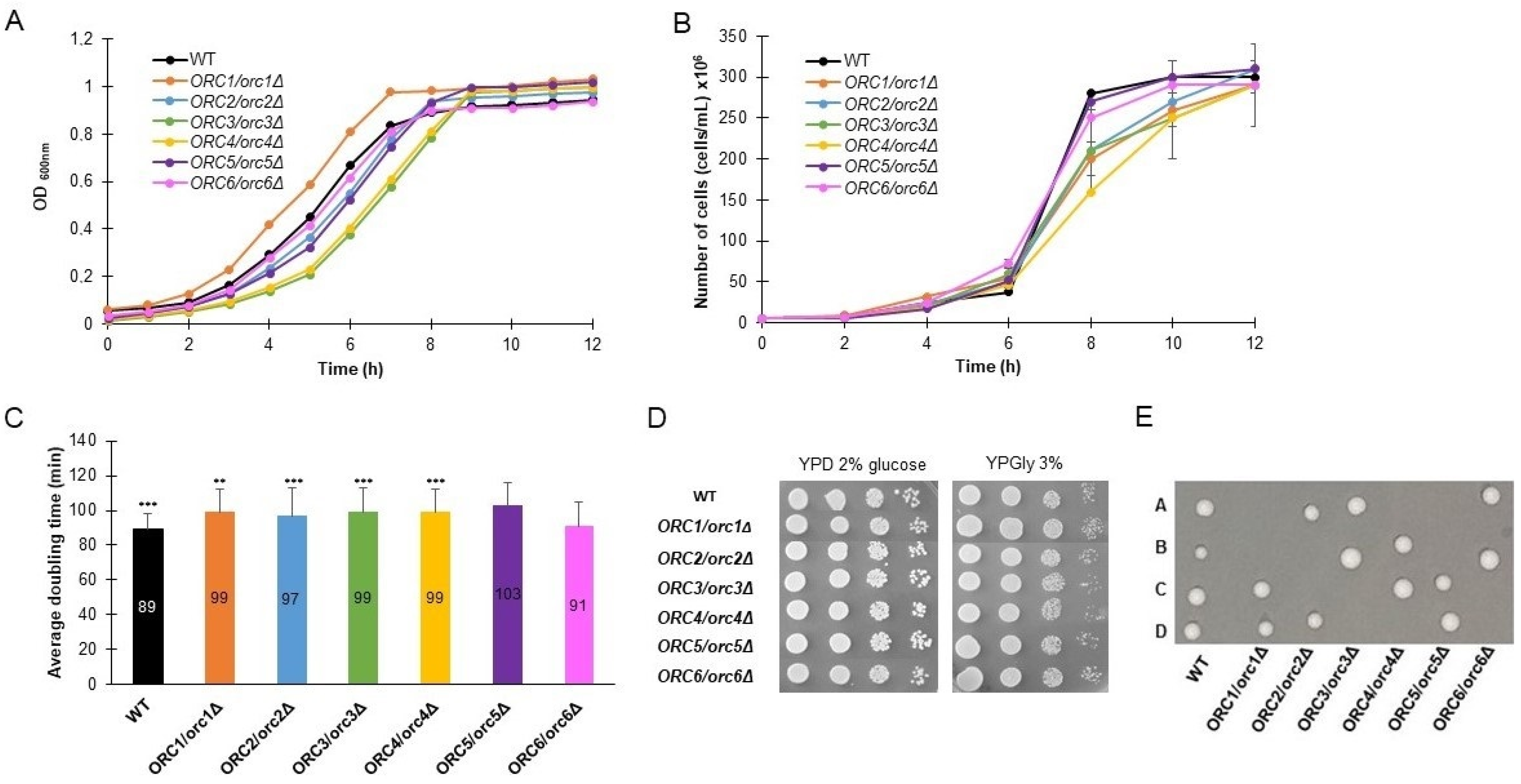
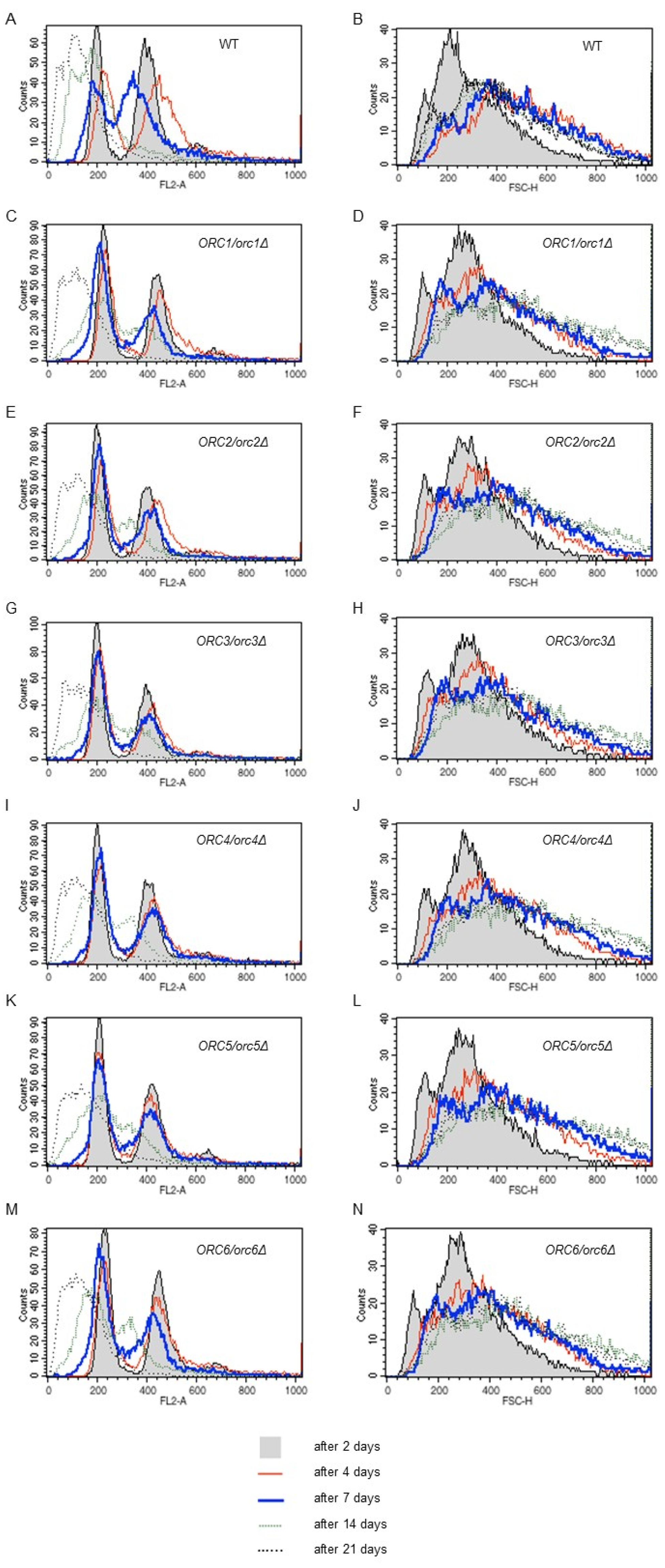
3.1. ORC/orcΔ Heterozygous Strains Grow Slowly Due to a Delay in G1/S Transition in the Cell Cycle
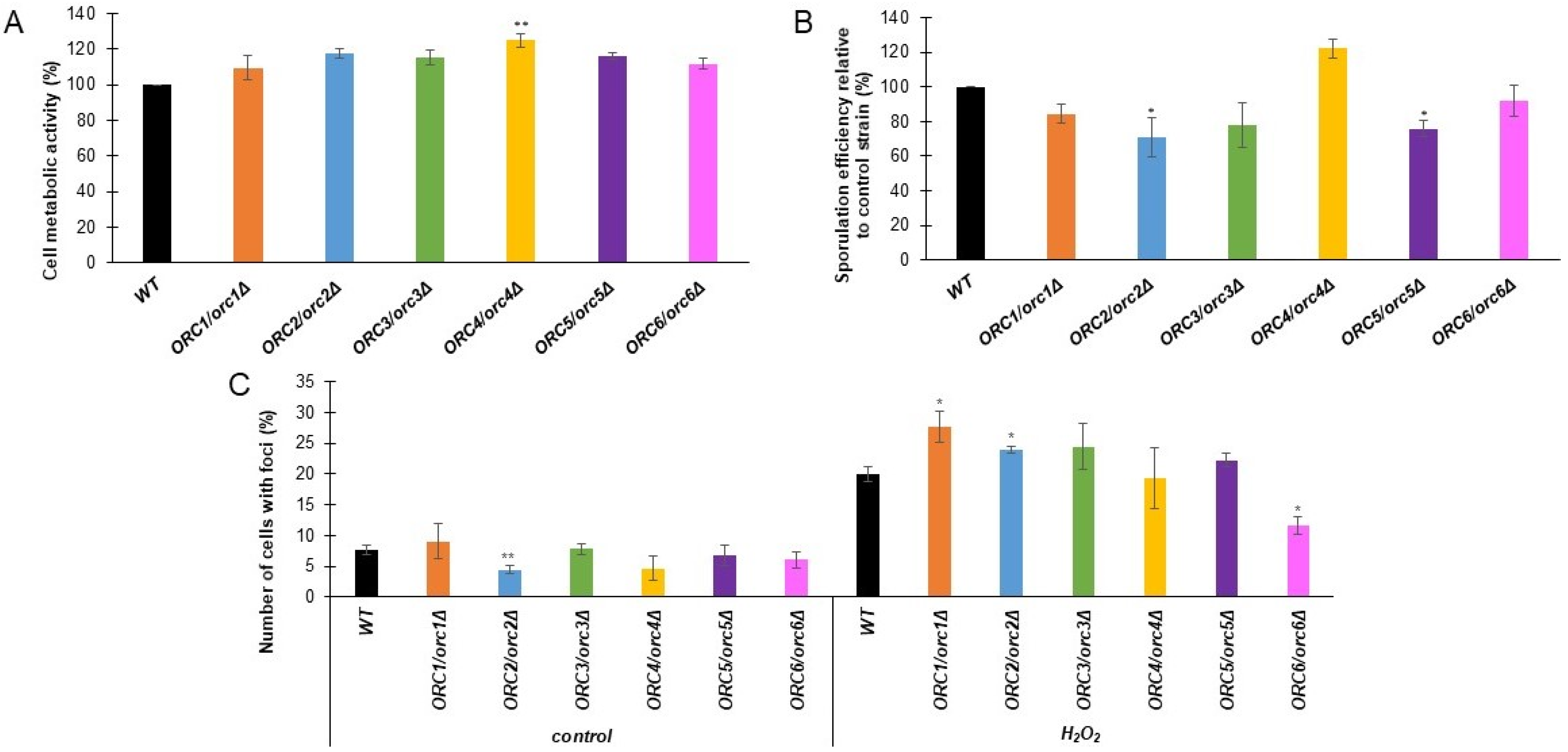
3.2. The Metabolic Activity, Sporulation Efficiency and Response to Oxidative Stress Are Changed in ORC/orcΔ Strains
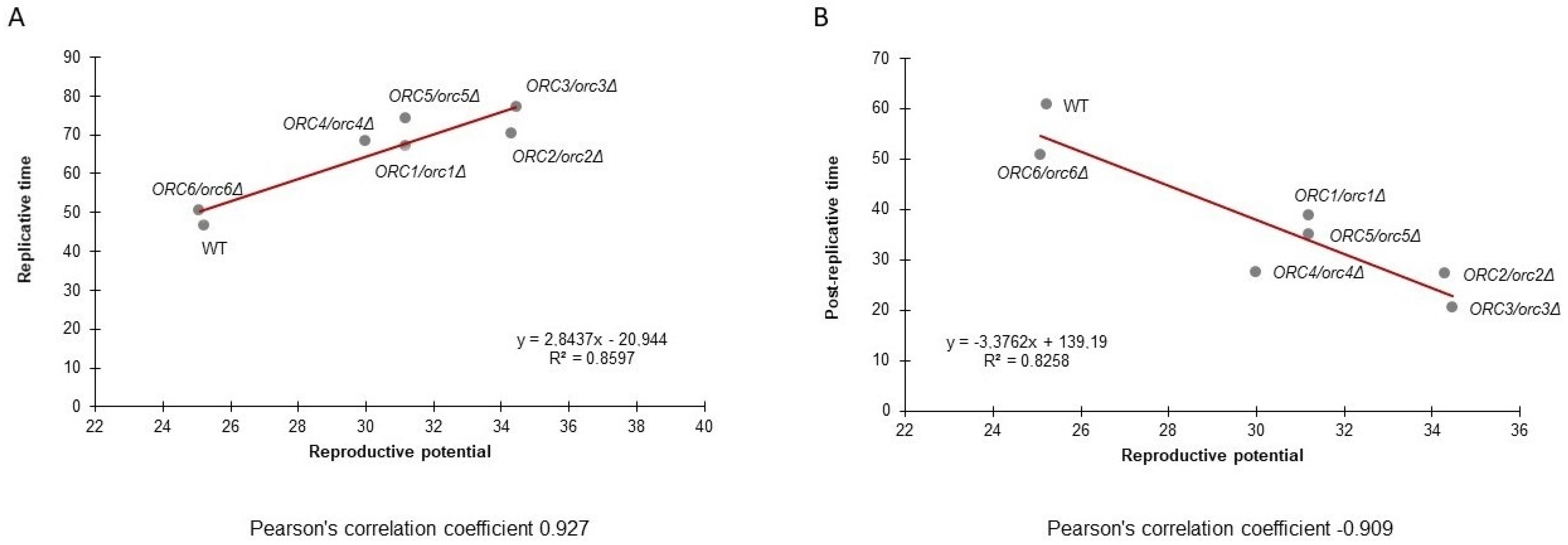
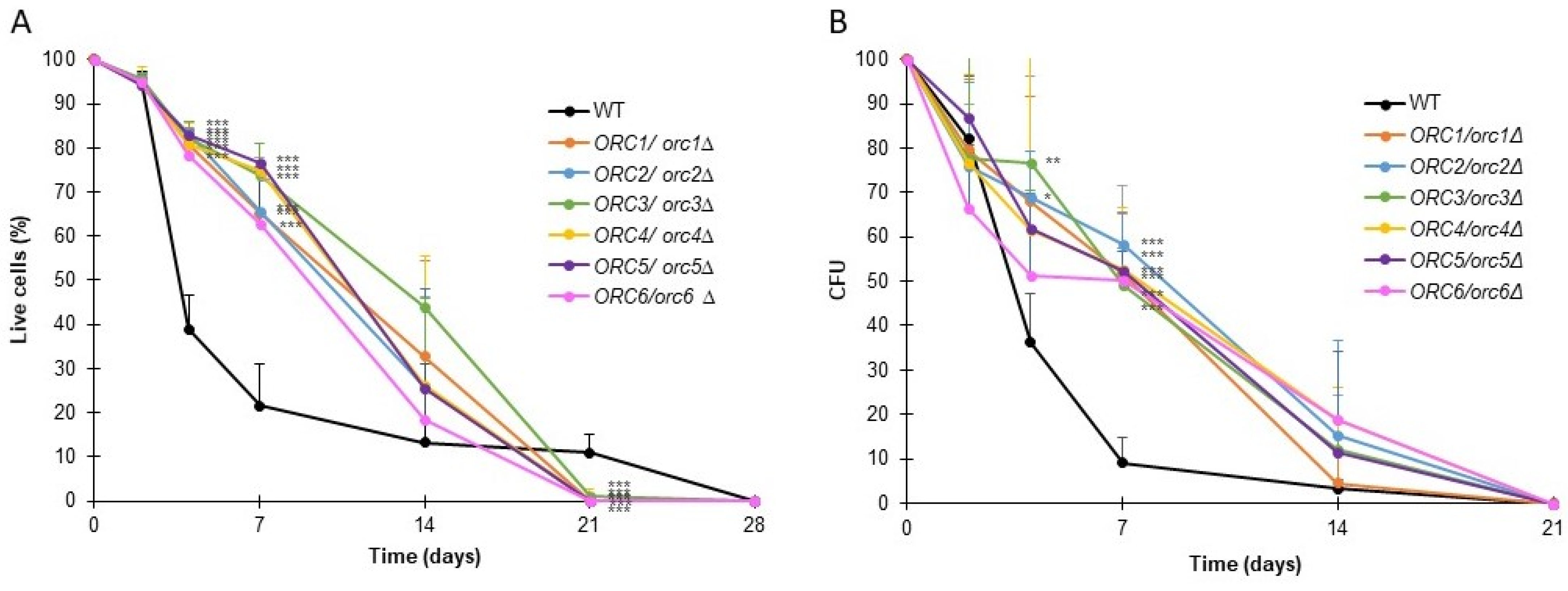
3.3. Strains with Reduced Expression of ORC Subunits Have Extended Budding and Chronological Lifespans
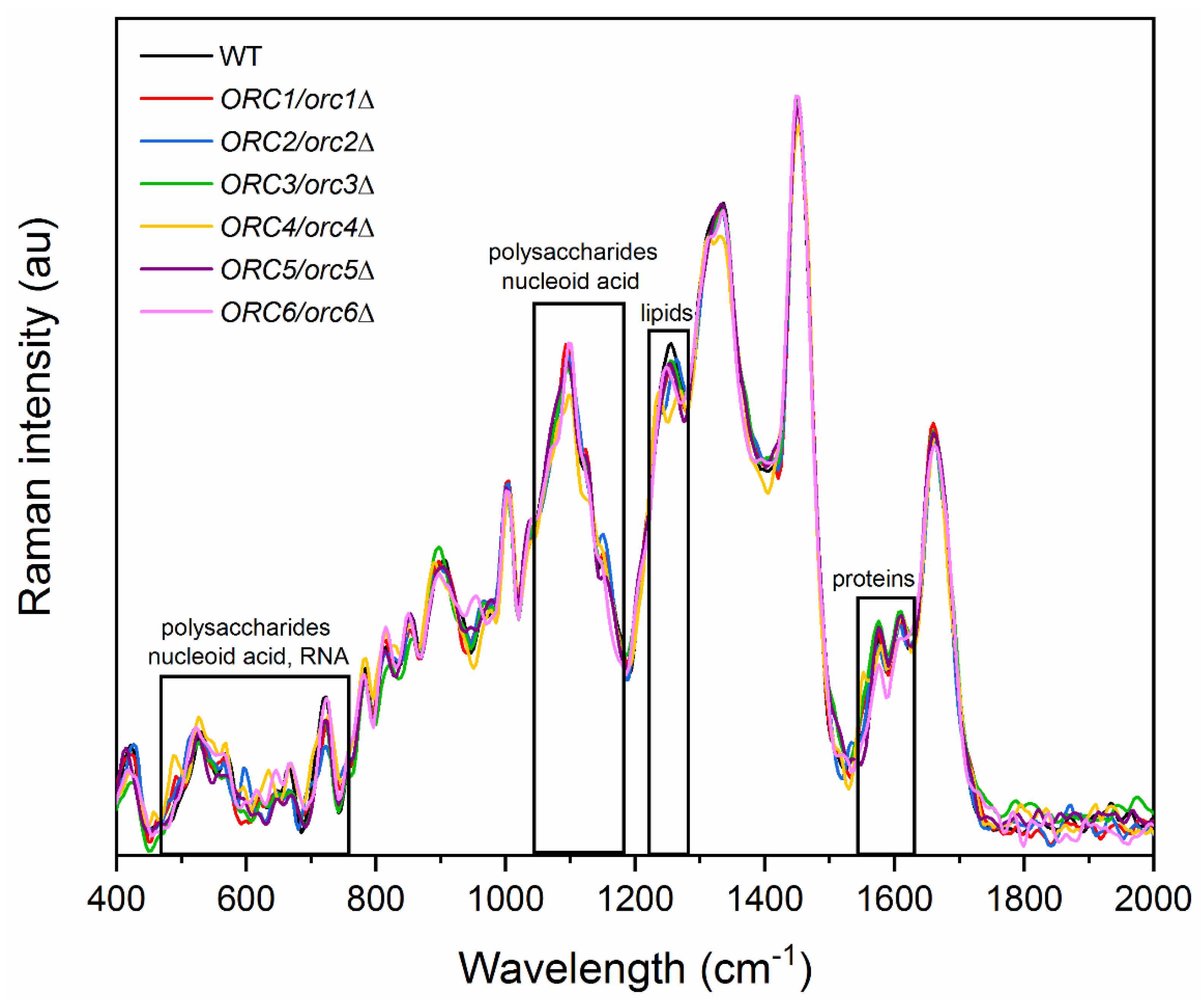
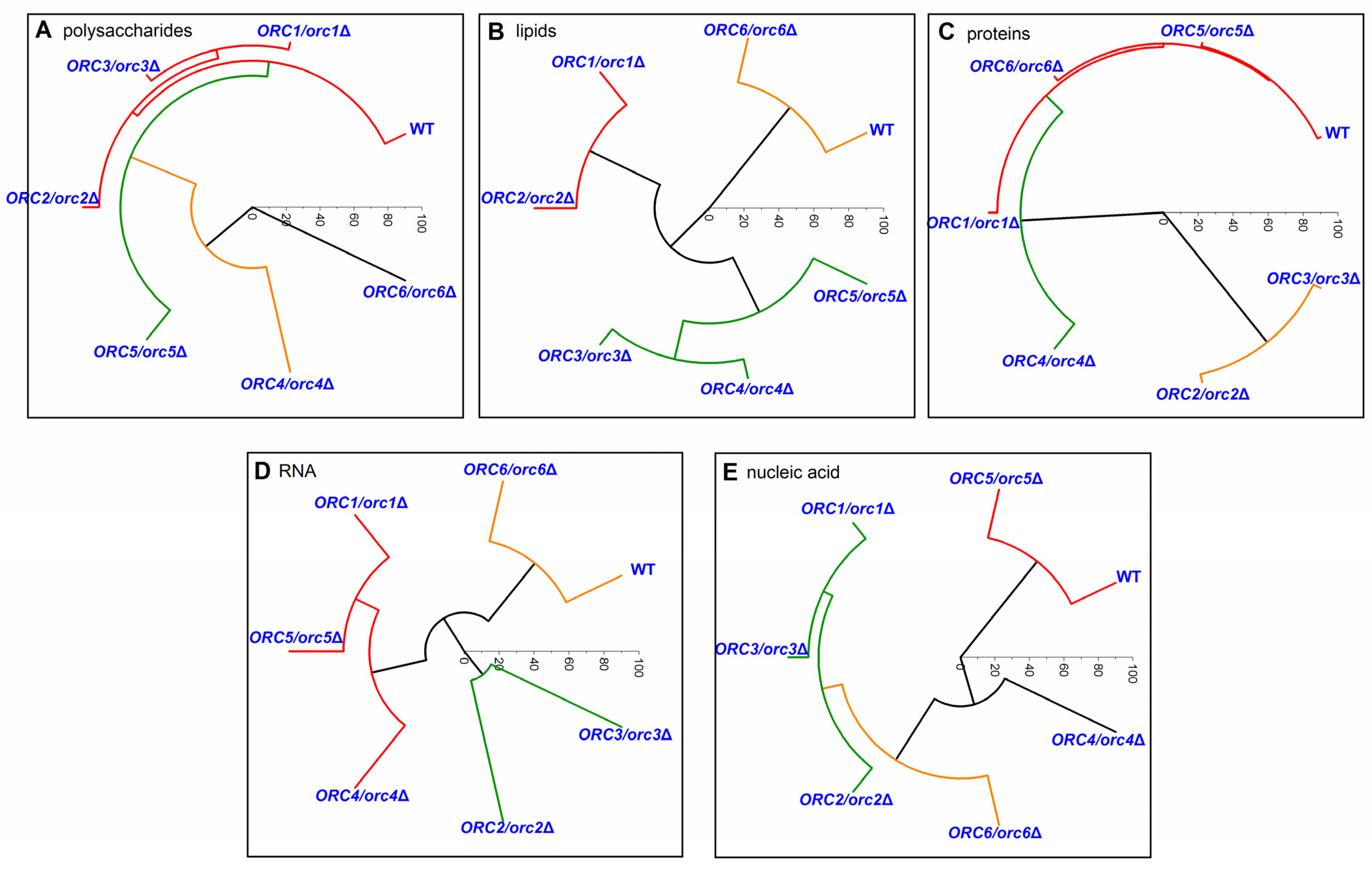
3.4. Reducing the Levels of Orc1-6 Proteins Influences the Chemical Composition of the Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diffley, J.F.X. Quality control in the initiation of eukaryotic DNA replication. Philos. Trans. R. Soc. B-Biol. Sci. 2011, 366, 3545–3553. [Google Scholar] [CrossRef] [Green Version]
- Bell, S.P.; Stillman, B. ATP-dependent recognition of eukaryotic origins of dna-replication by a multiprotein complex. Nature 1992, 357, 128–134. [Google Scholar] [CrossRef]
- Bell, S.P.; Mitchell, J.; Leber, J.; Kobayashi, R.; Stillman, B. The multidomain structure of orc1p reveals similarity to regulators of dna-replication and transcriptional silencing. Cell 1995, 83, 563–568. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, K.; Makise, M.; Sueyasu, Y.; Takehara, M.; Asano, T.; Mizushima, T. Yeast two-hybrid analysis of the origin recognition complex of Saccharomyces cerevisiae: Interaction between subunits and identification of binding proteins. Fems Yeast Res. 2007, 7, 1263–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnokov, I.N. Multiple functions of the origin recognition complex. Int. Rev. Cytol. 2007, 256, 69–109. [Google Scholar] [CrossRef] [PubMed]
- Gossen, M.; Pak, D.T.S.; Hansen, S.K.; Acharya, J.K.; Botchan, M.R. A Drosophila homolog of the yeast origin recognition complex. Science 1995, 270, 1674–1677. [Google Scholar] [CrossRef] [PubMed]
- Dhar, S.K.; Dutta, A. Identification and characterization of the human ORC6 homolog. J. Biol. Chem. 2000, 275, 34983–34988. [Google Scholar] [CrossRef] [Green Version]
- Bell, S.P. The origin recognition complex: From simple origins to complex functions. Genes Dev. 2002, 16, 659–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speck, C.; Chen, Z.Q.; Li, H.L.; Stillman, B. ATPase-dependent cooperative binding of ORC and Cdc6 to origin DNA. Nat. Struct. Mol. Biol. 2005, 12, 965–971. [Google Scholar] [CrossRef]
- Rao, H.; Stillman, B. The origin recognition complex interacts with a bipartite dna-binding site within yeast replicators. Proc. Natl. Acad. Sci. USA 1995, 92, 2224–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randell, J.C.W.; Bowers, J.L.; Rodriguez, H.K.; Bell, S.P. Sequential ATP hydrolysis by Cdc6 and ORC directs loading of the Mcm2-7 helicase. Mol. Cell 2006, 21, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Klemm, R.D.; Austin, R.J.; Bell, S.P. Coordinate binding of ATP and origin DNA regulates the ATPase activity of the origin recognition complex. Cell 1997, 88, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Semple, J.W.; Da-Silva, L.F.; Jervis, E.J.; Ah-Kee, J.; Al-Attar, H.; Kummer, L.; Heikkila, J.J.; Pasero, P.; Duncker, B.P. An essential role for Orc6 in DNA replication through maintenance of pre-replicative complexes. Embo J. 2006, 25, 5150–5158. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, J.C.; Kwan, E.X.; Pohl, T.J.; Amemiya, H.M.; Raghuraman, M.K.; Brewer, B.J. Defective replication initiation results in locus specific chromosome breakage and a ribosomal RNA deficiency in yeast. PLoS Genet. 2017, 13, e1007041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molon, M.; Zadrag-Tecza, R.; Bilinski, T. The longevity in the yeast Saccharomyces cerevisiae: A comparison of two approaches for assessment the lifespan. Biochem. Biophys. Res. Commun. 2015, 460, 651–656. [Google Scholar] [CrossRef]
- Fabrizio, P.; Longo, V.D. The chronological life span of Saccharomyces cerevisiae. Aging Cell 2003, 2, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, R.K.; Johnston, J.R. Life span of individual yeast cells. Nature 1959, 183, 1751–1752. [Google Scholar] [CrossRef] [Green Version]
- Minois, N.; Frajnt, M.; Wilson, C.; Vaupel, J.W. Advances in measuring lifespan in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2005, 102, 402–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaeberlein, M. Lessons on longevity from budding yeast. Nature 2010, 464, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Fabrizio, P.; Pletcher, S.D.; Minois, N.; Vaupel, J.W.; Longo, V.D. Chronological aging-independent replicative life span regulation by Msn2/Msn4 and Sod2 in Saccharomyces cerevisiae. FEBS Lett. 2004, 557, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Kaeberlein, M.; Hu, D.; Kerr, E.O.; Tsuchiya, M.; Westman, E.A.; Dang, N.; Fields, S.; Kennedy, B.K. Increased life span due to calorie restriction in respiratory-deficient yeast. PLoS Genet. 2005, 1, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Stepien, K.; Wojdyla, D.; Nowak, K.; Molon, M. Impact of curcumin on replicative and chronological aging in the Saccharomyces cerevisiae yeast. Biogerontology 2020, 21, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Czachor, J.; Milek, M.; Galiniak, S.; Stepien, K.; Dzugan, M.; Molon, M. Coffee Extends Yeast Chronological Lifespan through Antioxidant Properties. Int. J. Mol. Sci. 2020, 21, 9510. [Google Scholar] [CrossRef]
- Kwong, M.M.Y.; Lee, J.W.; Samian, M.R.; Wahab, H.A.; Watanabe, N.; Ong, E.B.B. Identification of Tropical Plant Extracts That Extend Yeast Chronological Life Span. Cells 2021, 10, 2718. [Google Scholar] [CrossRef] [PubMed]
- Steffen, K.K.; MacKay, V.L.; Kerr, E.O.; Tsuchiya, M.; Hu, D.; Fox, L.A.; Dang, N.; Johnston, E.D.; Oakes, J.A.; Tchao, B.N.; et al. Yeast life span extension by depletion of 60S ribosomal subunits is mediated by Gcn4. Cell 2008, 133, 292–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borkiewicz, L.; Molon, M.; Molestak, E.; Grela, P.; Horbowicz-Drozdzal, P.; Wawiorka, L.; Tchorzewski, M. Functional Analysis of the Ribosomal uL6 Protein of Saccharomyces cerevisiae. Cells 2019, 8, 718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molon, M.; Molestak, E.; Kula-Maximenko, M.; Grela, P.; Tchorzewski, M. Ribosomal Protein uL11 as a Regulator of Metabolic Circuits Related to Aging and Cell Cycle. Cells 2020, 9, 1745. [Google Scholar] [CrossRef]
- Delaney, J.R.; Murakami, C.; Chou, A.; Carr, D.; Schleit, J.; Sutphin, G.L.; An, E.H.; Castanza, A.S.; Fletcher, M.; Goswami, S.; et al. Dietary restriction and mitochondrial function link replicative and chronological aging in Saccharomyces cerevisiae. Exp. Gerontol. 2013, 48, 1006–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludovico, P.; Burhans, W.C. Reactive oxygen species, ageing and the hormesis police. Fems Yeast Res. 2014, 14, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, D.A.; Guarente, L. Extrachromosomal rDNA circles—A cause of aging in yeast. Cell 1997, 91, 1033–1042. [Google Scholar] [CrossRef] [Green Version]
- Molon, M.; Panek, A.; Molestak, E.; Skoneczny, M.; Tchorzewski, M.; Wnuk, M. Daughters of the budding yeast from old mothers have shorter replicative lifespans but not total lifespans. Are DNA damage and rDNA instability the factors that determine longevity? Cell Cycle 2018, 17, 1173–1187. [Google Scholar] [CrossRef] [Green Version]
- Molon, M.; Szajwaj, M.; Tchorzewski, M.; Skoczowski, A.; Niewiadomska, E.; Zadrag-Tecza, R. The rate of metabolism as a factor determining longevity of the Saccharomyces cerevisiae yeast. Age 2016, 38, 11. [Google Scholar] [CrossRef]
- Kwolek-Mirek, M.; Zadrag-Tecza, R. Comparison of methods used for assessing the viability and vitality of yeast cells. Fems Yeast Res. 2014, 14, 1068–1079. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rosell, J.; Sunjevaric, I.; De Piccoli, G.; Sacher, M.; Eckert-Boulet, N.; Reid, R.; Jentsch, S.; Rothstein, R.; Aragon, L.; Lisby, M. The Smc5-Smc6 complex and SUMO modification of Rad52 regulates recombinational repair at the ribosomal gene locus. Nat. Cell Biol. 2007, 9, U923–U972. [Google Scholar] [CrossRef]
- Krol, K.; Antoniuk-Majchrzak, J.; Skoneczny, M.; Sienko, M.; Jendrysek, J.; Rumienczyk, I.; Halas, A.; Kurlandzka, A.; Skoneczna, A. Lack of G1/S control destabilizes the yeast genome via replication stress-induced DSBs and illegitimate recombination. J. Cell Sci. 2018, 131, jcs226480. [Google Scholar] [CrossRef] [Green Version]
- Molon, M.; Zebrowski, J. Phylogenetic relationship and Fourier-transform infrared spectroscopy-derived lipid determinants of lifespan parameters in the Saccharomyces cerevisiae yeast. FEMS Yeast Res. 2017, 17, fox031. [Google Scholar] [CrossRef]
- Rosch, P.; Harz, M.; Schmitt, M.; Popp, J. Raman spectroscopic identification of single yeast cells. J. Raman Spectrosc. 2005, 36, 377–379. [Google Scholar] [CrossRef]
- Chan, J.W.; Winhold, H.; Corzett, M.H.; Ulloa, J.M.; Cosman, M.; Balhorn, R.; Huser, T. Monitoring dynamic protein expression in living E-coli. Bacterial Celts by laser tweezers raman spectroscopy. Cytom. Part A 2007, 71A, 468–474. [Google Scholar] [CrossRef]
- Lemma, T.; Wang, J.; Arstila, K.; Hytonen, V.P.; Toppari, J.J. Identifying yeasts using surface enhanced Raman spectroscopy. Spectrochim. Acta Part A-Mol. Biomol. Spectrosc. 2019, 218, 299–307. [Google Scholar] [CrossRef] [PubMed]
- De Gelder, J.; De Gussem, K.; Vandenabeele, P.; Moens, L. Reference database of Raman spectra of biological molecules. J. Raman Spectrosc. 2007, 38, 1133–1147. [Google Scholar] [CrossRef]
- Kawakami, H.; Katayama, T. DnaA, ORC, and Cdc6: Similarity beyond the domains of life and diversity. Biochem. Cell Biol. Biochim. Et Biol. Cell. 2010, 88, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.P.; Dutta, A. DNA replication in eukaryotic cells. Annu. Rev. Biochem. 2002, 71, 333–374. [Google Scholar] [CrossRef] [Green Version]
- Scholefield, G.; Veening, J.W.; Murray, H. DnaA and ORC: More than DNA replication initiators. Trends Cell Biol. 2011, 21, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, A.; Bell, S.P. Initiation of DNA replication in eukaryotic cells. Annu. Rev. Cell Dev. Biol. 1997, 13, 293–332. [Google Scholar] [CrossRef]
- Gibson, D.G.; Bell, S.P.; Aparicio, O.M. Cell cycle execution point analysis of ORC function and characterization of the checkpoint response to ORC inactivation in Saccharomyces cerevisiae. Genes Cells 2006, 11, 557–573. [Google Scholar] [CrossRef]
- Clarey, M.G.; Erzberger, J.P.; Grob, P.; Leschziner, A.E.; Berger, J.M.; Nogales, E.; Botchan, M. Nucleotide-dependent conformational changes in the DnaA-like core of the origin recognition complex. Nat. Struct. Mol. Biol. 2006, 13, 684–690. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Bell, S.P. Architecture of the yeast origin recognition complex bound to origins of DNA replication. Mol. Cell. Biol. 1997, 17, 7159–7168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, T.; Makise, M.; Takehara, M.; Mizushima, T. Interaction between ORC and Cdt1p of Saccharomyces cerevisiae. Fems Yeast Res. 2007, 7, 1256–1262. [Google Scholar] [CrossRef] [Green Version]
- Takara, T.J.; Bell, S.P. Multiple Cdt1 molecules act at each origin to load replication-competent Mcm2-7 helicases. Embo J. 2011, 30, 4885–4896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Prat, L.; Martinez-Vicente, M.; Perdiguero, E.; Ortet, L.; Rodriguez-Ubreva, J.; Rebollo, E.; Ruiz-Bonilla, V.; Gutarra, S.; Ballestar, E.; Serrano, A.L.; et al. Autophagy maintains stemness by preventing senescence. Nature 2016, 529, 37–42. [Google Scholar] [CrossRef]
- Bartkova, J.; Rezaei, N.; Liontos, M.; Karakaidos, P.; Kletsas, D.; Issaeva, N.; Vassiliou, L.V.F.; Kolettas, E.; Niforou, K.; Zoumpourlis, V.C.; et al. Oncogene-induced senescence is part of the tumorigenesis barrier imposed by DNA damage checkpoints. Nature 2006, 444, 633–637. [Google Scholar] [CrossRef]
- Gorgoulis, V.G.; Halazonetis, T.D. Oncogene-induced senescence: The bright and dark side of the response. Curr. Opin. Cell Biol. 2010, 22, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Bleichert, F. Mechanisms of replication origin licensing: A structural perspective. Curr. Opin. Struct. Biol. 2019, 59, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Fujita, M. Molecular Mechanism for Chromatin Regulation During MCM Loading in Mammalian Cells. DNA Replication: Old Princ. New Discov. 2017, 1042, 61–78. [Google Scholar] [CrossRef]
- Hills, S.A.; Diffley, J.F.X. DNA Replication and Oncogene-Induced Replicative Stress. Curr. Biol. 2014, 24, R435–R444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higa, M.; Matsuda, Y.; Fujii, J.; Sugimoto, N.; Yoshida, K.; Fujita, M. TRF2-mediated ORC recruitment underlies telomere stability upon DNA replication stress. Nucleic Acids Res. 2021, 49, 12234–12251. [Google Scholar] [CrossRef]
- Tatsumi, Y.; Ezura, K.; Yoshida, K.; Yugawa, T.; Narisawa-Saito, M.; Kiyono, T.; Ohta, S.; Obuse, C.; Fujita, M. Involvement of human ORC and TRF2 in pre-replication complex assembly at telomeres. Genes Cells 2008, 13, 1045–1059. [Google Scholar] [CrossRef]
- Yoshida, R.; Tamura, T.; Takaoka, C.; Harada, K.; Kobayashi, A.; Mukai, Y.; Fukusaki, E. Metabolomics-based systematic prediction of yeast lifespan and its application for semi-rational screening of ageing-related mutants. Aging Cell 2010, 9, 616–625. [Google Scholar] [CrossRef]
- Petschnigg, J.; Wolinski, H.; Kolb, D.; Zellnig, G.; Kurat, C.F.; Natter, K.; Kohlwein, S.D. Good Fat, Essential Cellular Requirements for Triacylglycerol Synthesis to Maintain Membrane Homeostasis in Yeast. J. Biol. Chem. 2009, 284, 30981–30993. [Google Scholar] [CrossRef] [Green Version]
- Handee, W.; Li, X.B.; Hall, K.W.; Deng, X.X.; Li, P.; Benning, C.; Williams, B.L.; Kuo, M.H. An Energy-Independent Pro-longevity Function of Triacylglycerol in Yeast. PLoS Genet. 2016, 12, e1005878. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.D.; Shadel, G.S.; Kaeberlein, M.; Kennedy, B. Replicative and Chronological Aging in Saccharomyces cerevisiae. Cell Metab. 2012, 16, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Longo, V.D.; Gralla, E.B.; Valentine, J.S. Superoxide dismutase activity is essential for stationary phase survival in Saccharomyces cerevisiae—Mitochondrial production of toxic oxygen species in vivo. J. Biol. Chem. 1996, 271, 12275–12280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrizio, P.; Pozza, F.; Pletcher, S.D.; Gendron, C.M.; Longo, V.D. Regulation of longevity and stress resistance by Sch9 in yeast. Science 2001, 292, 288–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrizio, P.; Liou, L.L.; Moy, V.N.; Diaspro, A.; SelverstoneValentine, J.; Gralla, E.B.; Longo, V.D. SOD2 functions downstream of Sch9 to extend longevity in yeast. Genetics 2003, 163, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, H.; Whiteway, M. Increased respiration in the sch9 Delta mutant is required for increasing chronological life span but not replicative life span. Eukaryot. Cell 2008, 7, 1127–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnuki, S.; Ohya, Y. High-dimensional single-cell phenotyping reveals extensive haploinsufficiency. PLoS Biol. 2018, 16, e2005130. [Google Scholar] [CrossRef] [Green Version]
- Legrand, M.; Lephart, P.; Forche, A.; Mueller, F.M.C.; Walsh, T.; Magee, P.T.; Magee, B.B. Homozygosity at the MTL locus in clinical strains of Candida albicans: Karyotypic rearrangements and tetraploid formation. Mol. Microbiol. 2004, 52, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Krol, K.; Brozda, I.; Skoneczny, M.; Bretne, M.; Skoneczna, A. A Genomic Screen Revealing the Importance of Vesicular Trafficking Pathways in Genome Maintenance and Protection against Genotoxic Stress in Diploid Saccharomyces cerevisiae Cells. PLoS ONE 2015, 10, e0120702. [Google Scholar] [CrossRef] [Green Version]
- Alabrudzinska, M.; Skoneczny, M.; Skoneczna, A. Diploid-Specific Genome Stability Genes of S. cerevisiae: Genomic Screen Reveals Haploidization as an Escape from Persisting DNA Rearrangement Stress. PLoS ONE 2011, 6, e21124. [Google Scholar] [CrossRef]
- Skoneczna, A.; Kaniak, A.; Skoneczny, M. Genetic instability in budding and fission yeast-sources and mechanisms. Fems Microbiol. Rev. 2015, 39, 917–967. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Petes, T.D. Haploidization in Saccharomyces cerevisiae Induced by a Deficiency in Homologous Recombination. Genetics 2012, 191, 279–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerstein, A.C.; McBride, R.M.; Otto, S.P. Ploidy reduction in Saccharomyces cerevisiae. Biol. Lett. 2008, 4, 91–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitale, I.; Galluzzi, L.; Castedo, M.; Kroemer, G. Mitotic catastrophe: A mechanism for avoiding genomic instability. Nat. Rev. Mol. Cell Biol. 2011, 12, 384–391. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| BY4743 a/α | Mat a/α; his3Δ1/his3Δ1; leu2Δ0/leu2Δ0; lys2Δ0/LYS2; MET15/met15Δ0; ura3Δ0/ura3Δ0 | Open Biosystems |
| ORC1/orc1Δ | Mat a/α; his3Δ1/his3Δ1; leu2Δ0/leu2Δ0; lys2Δ0/LYS2; MET15/met15Δ0; ura3Δ0/ura3Δ0; yml065w::kanMX4/YML065w | Open Biosystems |
| ORC2/orc2Δ | Mat a/α; his3Δ1/his3Δ1; leu2Δ0/leu2Δ0; lys2Δ0/LYS2; MET15/met15Δ0; ura3Δ0/ura3Δ0; ybr060c::kanMX4/YBR060c | Open Biosystems |
| ORC3/orc3Δ | Mat a/α; his3Δ1/his3Δ1; leu2Δ0/leu2Δ0; lys2Δ0/LYS2; MET15/met15Δ0; ura3Δ0/ura3Δ0; yll004w::kanMX4/YLL004w | Open Biosystems |
| ORC4/orc4Δ | Mat a/α; his3Δ1/his3Δ1; leu2Δ0/leu2Δ0; lys2Δ0/LYS2; MET15/met15Δ0; ura3Δ0/ura3Δ0; ypr162c::kanMX4/YPR162c | Open Biosystems |
| ORC5/orc5Δ | Mat a/α; his3Δ1/his3Δ1; leu2Δ0/leu2Δ0; lys2Δ0/LYS2; MET15/met15Δ0; ura3Δ0/ura3Δ0; ynl261w::kanMX4/YNL261w | Open Biosystems |
| ORC6/orc6Δ | Mat a/α; his3Δ1/his3Δ1; leu2Δ0/leu2Δ0; lys2Δ0/LYS2; MET15/met15Δ0; ura3Δ0/ura3Δ0; yhr118c::kanMX4/YHR118c | Open Biosystems |
| Gene Name | Forward Primer 5′->3′ | Reverse Primer 5′->3′ | Source |
|---|---|---|---|
| ACT1 | GTAAGGAATTATACGGTAACATC | TAGATGGACCACTTTCGTCG | in this study |
| ORC1 | TGGGTATACGCACGAAGAGC | TCCTCACGTCTTCAGGCAAC | in this study |
| ORC2 | ATTTACGCTCCGCTCCTCTG | CTTCAGCACCACTGCTGGTA | in this study |
| ORC3 | ACTGAGCAGATGTCCTACATTCA | GCCCGTTAATCGGGTTCTCT | in this study |
| ORC4 | AGCTCGTCTATCACCGCAAG | CCAGGGTCGCTGTCTTTACA | in this study |
| ORC5 | GGATTCCTCACGAAGTGCAGA | GGTAGAGCTGCTTATGGACG | in this study |
| ORC6 | AACCAGGAAACGACGGTTTG | TTGTTTCGTTCTCCCGCTTC | in this study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stępień, K.; Skoneczna, A.; Kula-Maximenko, M.; Jurczyk, Ł.; Mołoń, M. Depletion of the Origin Recognition Complex Subunits Delays Aging in Budding Yeast. Cells 2022, 11, 1252. https://doi.org/10.3390/cells11081252
Stępień K, Skoneczna A, Kula-Maximenko M, Jurczyk Ł, Mołoń M. Depletion of the Origin Recognition Complex Subunits Delays Aging in Budding Yeast. Cells. 2022; 11(8):1252. https://doi.org/10.3390/cells11081252
Chicago/Turabian StyleStępień, Karolina, Adrianna Skoneczna, Monika Kula-Maximenko, Łukasz Jurczyk, and Mateusz Mołoń. 2022. "Depletion of the Origin Recognition Complex Subunits Delays Aging in Budding Yeast" Cells 11, no. 8: 1252. https://doi.org/10.3390/cells11081252
APA StyleStępień, K., Skoneczna, A., Kula-Maximenko, M., Jurczyk, Ł., & Mołoń, M. (2022). Depletion of the Origin Recognition Complex Subunits Delays Aging in Budding Yeast. Cells, 11(8), 1252. https://doi.org/10.3390/cells11081252