
miR-126-3p and miR-21-5p as Hallmarks of Bio-Positive Ageing; Correlation Analysis and Machine Learning Prediction in Young to Ultra-Centenarian Sicilian Population

 ,
,  , , ,
, , ,  ,
,  , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design, Participants, and Anamnestic Data
2.2. Plasma Sample Acquisition and RNA Isolation
2.3. TaqMan RT-qPCR miRNA Assays
2.4. Cell Cultures
2.5. Collection of Cell-Conditioned Media and miRNA Isolation
2.6. Senescence-Associated β-Galactosidase Staining
2.7. MTS Assay
2.8. miRNA Targeted Gene Prediction and KEGG Pathway Analyses by miRWalk
2.9. Statistical Analyses of miRNA Levels
2.10. ML Techniques
3. Results
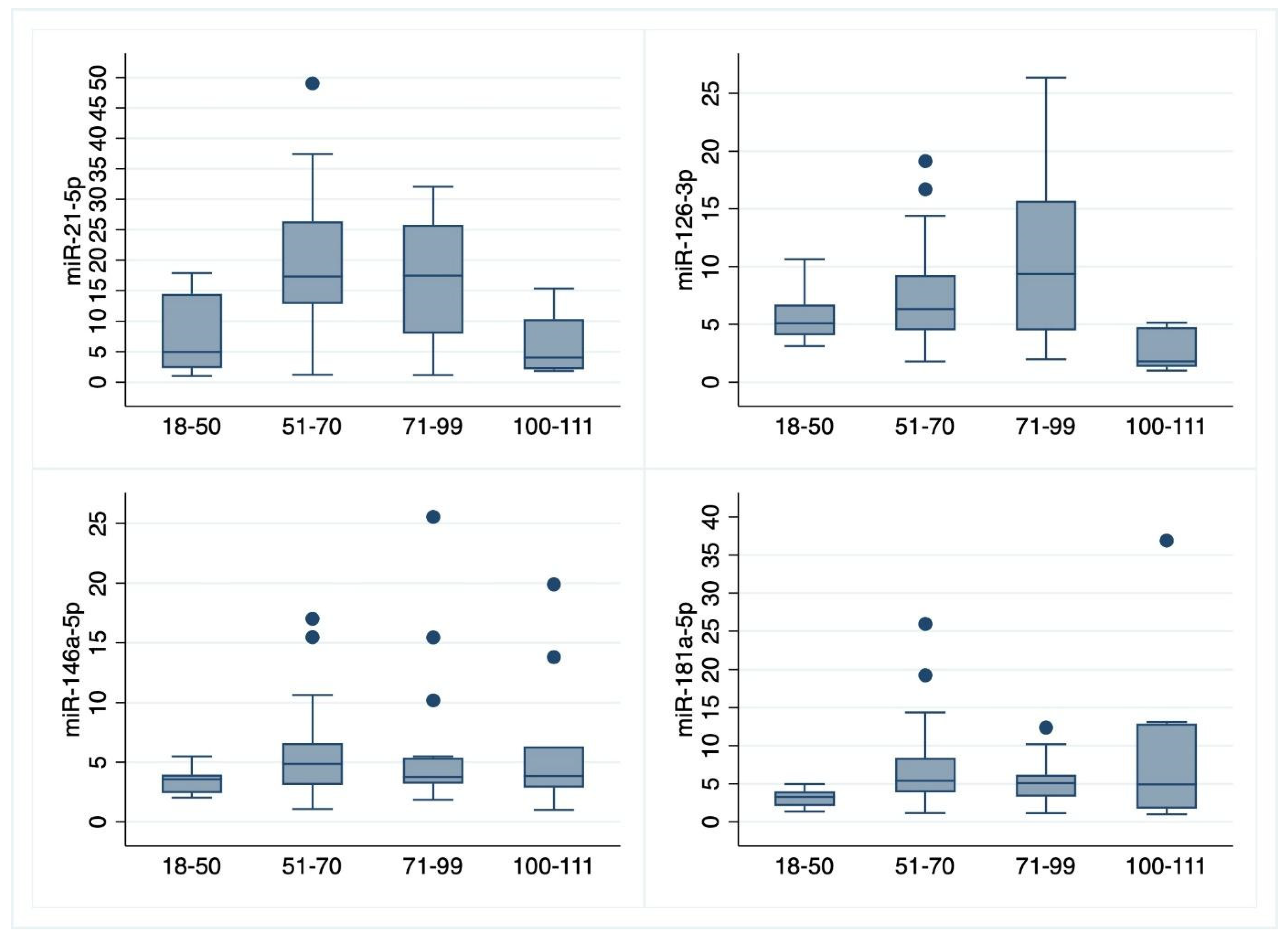
3.1. Plasma Values of miRNAs
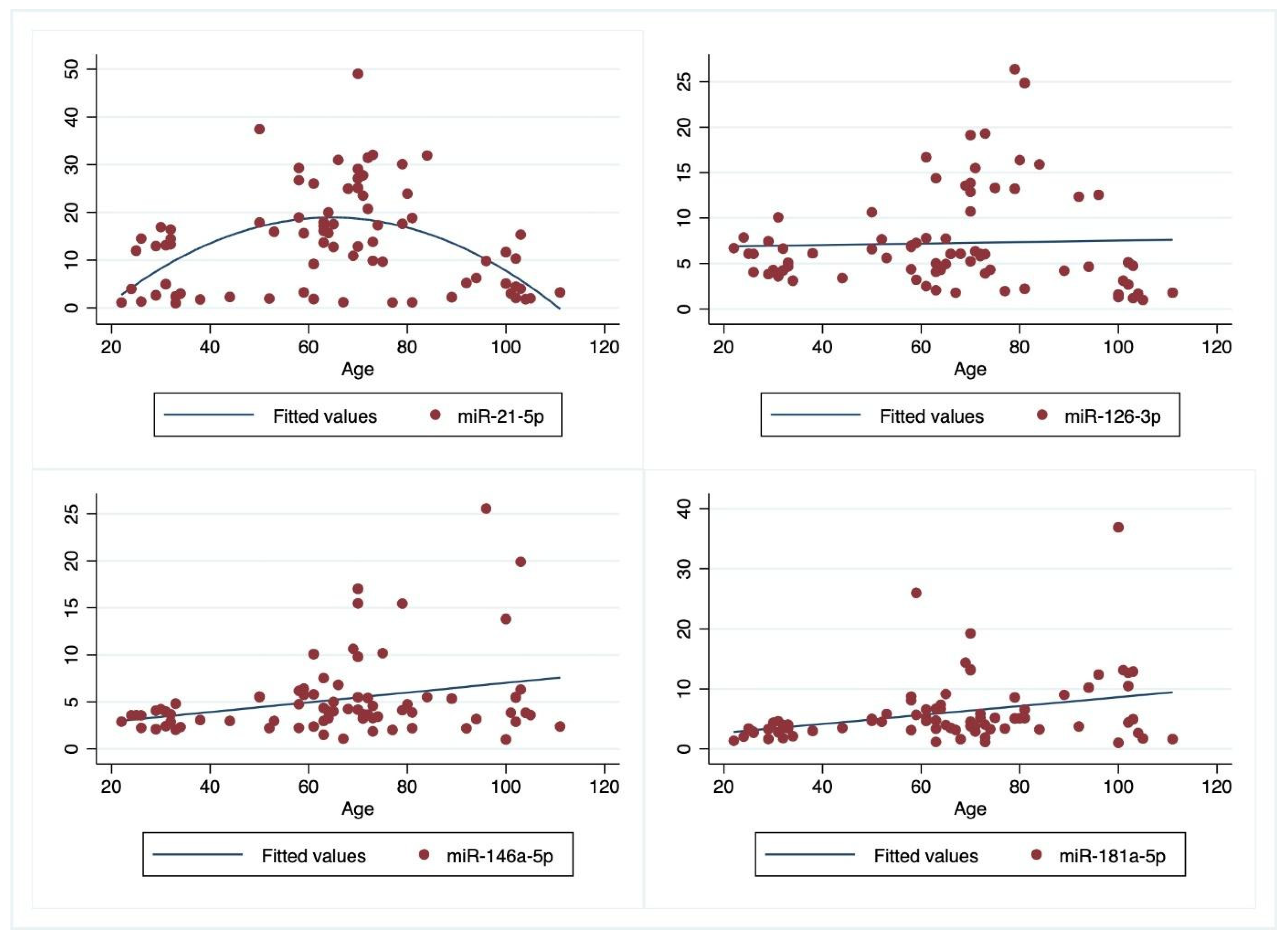
3.2. Correlation of Plasma Values of miRNAs with Some Parameters
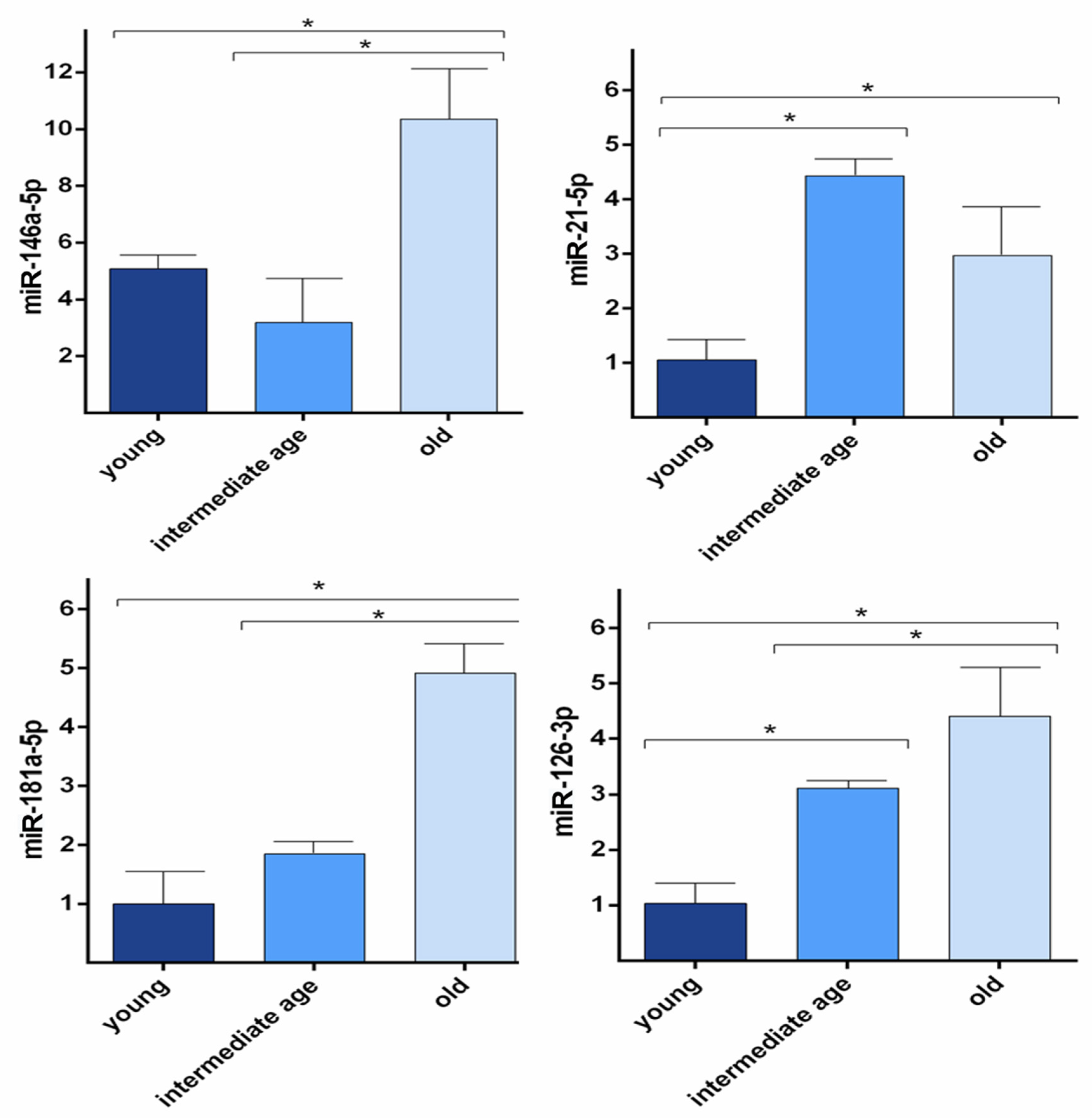
3.3. miRNAs Levels in HUVECs Undergoing Replicative Senescence
3.4. Enriched KEGG Pathway Clustered by Validated Targets of miR-21-5p, miR-126-3p, miR-146a-5p, and miR-181a-5p and Corresponding Target Genes
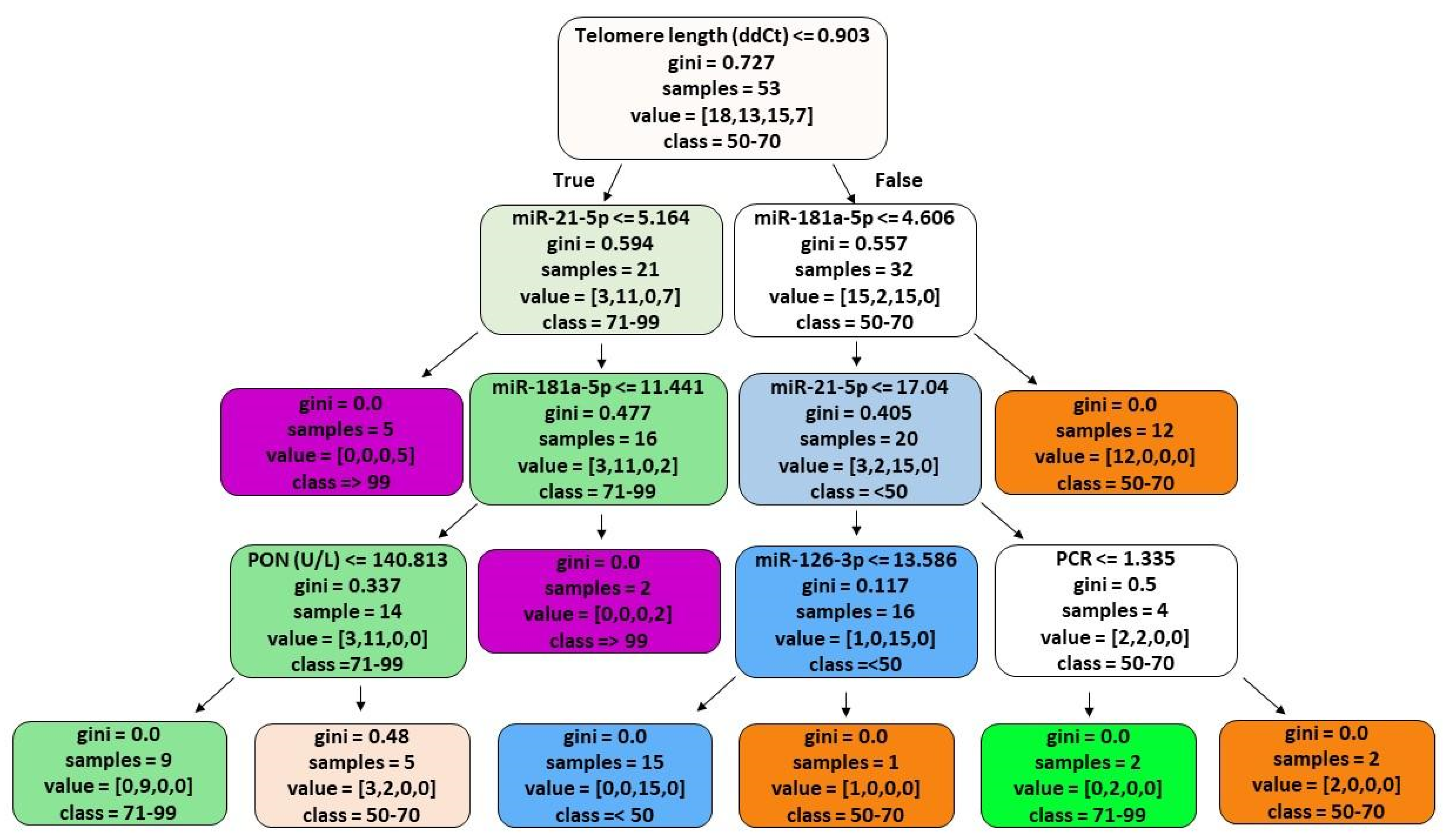
3.5. ML Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jung, H.J.; Suh, Y. Circulating miRNAs in ageing and ageing-related diseases. J. Genet. Genom. 2014, 41, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Curto, A.; Taverna, S.; Costa, M.A.; Passantino, R.; Augello, G.; Adamo, G.; Aiello, A.; Colomba, P.; Zizzo, C.; Zora, M.; et al. Can Be miR-126-3p a Biomarker of Premature Aging? An Ex Vivo and In Vitro Study in Fabry Disease. Cells 2021, 10, 356. [Google Scholar] [CrossRef] [PubMed]
- Pucci, M.; Reclusa Asiáin, P.; Duréndez Sáez, E.; Jantus-Lewintre, E.; Malarani, M.; Khan, S.; Fontana, S.; Naing, A.; Passiglia, F.; Raez, L.E.; et al. Extracellular Vesicles As miRNA Nano-Shuttles: Dual Role in Tumor Progression. Target Oncol. 2018, 13, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Kinser, H.E.; Pincus, Z. MicroRNAs as modulators of longevity and the aging process. Hum. Genet. 2020, 139, 291–308. [Google Scholar] [CrossRef] [PubMed]
- miRbase: The microRNA database Home Page. Available online: https://www.miRbase.org/index.shtml (accessed on 20 February 2022).
- Kato, M.; Chen, X.; Inukai, S.; Zhao, H.; Slack, F.J. Age-associated changes in expression of small, noncoding RNAs, including microRNAs, in C. elegans. RNA 2011, 17, 1804–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith-Vikos, T.; Slack, F.J. MicroRNAs and their roles in aging. J. Cell Sci. 2012, 125, 7–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Vijayan, M.; Bhatti, J.S.; Reddy, P.H. MicroRNAs as Peripheral Biomarkers in Aging and Age-Related Diseases. Prog. Mol. Biol. Transl. Sci. 2017, 146, 47–94. [Google Scholar] [CrossRef]
- Ogonowski, N.; Salcidua, S.; Leon, T.; Chamorro-Veloso, N.; Valls, C.; Avalos, C.; Bisquertt, A.; Rentería, M.E.; Orellana, P.; Duran-Aniotz, C. Systematic Review: microRNAs as Potential Biomarkers in Mild Cognitive Impairment Diagnosis. Front. Aging Neurosci. 2022, 13, 807764. [Google Scholar] [CrossRef]
- Olivieri, F.; Capri, M.; Bonafè, M.; Morsiani, C.; Jung, H.J.; Spazzafumo, L.; Viña, J.; Suh, Y. Circulating miRNAs and miRNA shuttles as biomarkers: Perspective trajectories of healthy and unhealthy aging. Mech. Ageing Dev. 2017, 165, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Olivieri, F.; Prattichizzo, F.; Giuliani, A.; Matacchione, G.; Rippo, M.R.; Sabbatinelli, J.; Bonafè, M. miR-21 and miR-146a: The microRNAs of inflammaging and age-related diseases. Ageing Res. Rev. 2021, 70, 101374. [Google Scholar] [CrossRef]
- Olivieri, F.; Rippo, M.R.; Procopio, A.D.; Fazioli, F. Circulating inflamma-miRs in aging and age-related diseases. Front. Genet. 2013, 4, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balzano, F.; Deiana, M.; Dei Giudici, S.; Oggiano, A.; Pasella, S.; Pinna, S.; Mannu, A.; Deiana, N.; Porcu, B.; Masala, A.G.E.; et al. MicroRNA Expression Analysis of Centenarians and Rheumatoid Arthritis Patients Reveals a Common Expression Pattern. Int. J. Med. Sci. 2017, 14, 622–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serna, E.; Gambini, J.; Borras, C.; Abdelaziz, K.M.; Belenguer, A.; Sanchis, P.; Avellana, J.A.; Rodriguez-Mañas, L.; Viña, J. Centenarians, but not octogenarians, up-regulate the expression of microRNAs. Sci. Rep. 2012, 2, 961, Erratum in: Sci. Rep. 2013, 3, 1575. Mohammed, Kheira [corrected to Abdelaziz, Kheira M]. [Google Scholar] [CrossRef]
- Smith-Vikos, T.; Liu, Z.; Parsons, C.; Gorospe, M.; Ferrucci, L.; Gill, T.M.; Slack, F.J. A serum miRNA profile of human longevity: Findings from the Baltimore Longitudinal Study of Aging (BLSA). Aging 2016, 8, 2971–2987. [Google Scholar] [CrossRef] [Green Version]
- Cammarata, G.; Duro, G.; Chiara, T.D.; Curto, A.L.; Taverna, S.; Candore, G. Circulating miRNAs in Successful and Unsuccessful Aging. A Mini-review. Curr. Pharm. Des. 2019, 25, 4150–4153. [Google Scholar] [CrossRef]
- Yamakuchi, M.; Hashiguchi, T. Endothelial Cell Aging: How miRNAs Contribute? J. Clin. Med. 2018, 7, 170. [Google Scholar] [CrossRef] [Green Version]
- Venkat, P.; Cui, C.; Chopp, M.; Zacharek, A.; Wang, F.; Landschoot-Ward, J.; Shen, Y.; Chen, J. MiR-126 Mediates Brain Endothelial Cell Exosome Treatment-Induced Neurorestorative Effects After Stroke in Type 2 Diabetes Mellitus Mice. Stroke 2019, 50, 2865–2874. [Google Scholar] [CrossRef]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Jebari-Benslaiman, S.; Galicia-García, U.; Larrea-Sebal, A.; Olaetxea, J.R.; Alloza, I.; Vandenbroeck, K.; Benito-Vicente, A.; Martín, C. Pathophysiology of Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 3346. [Google Scholar] [CrossRef]
- Borja-Gonzalez, M.; Casas-Martinez, J.C.; McDonagh, B.; Goljanek-Whysall, K. Inflamma-miR-21 Negatively Regulates Myogenesis during Ageing. Antioxidants 2020, 9, 345. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.; Lin, P.; Wang, J.; Yu, H.; Lv, T.; Sun, L.; Du, G. Circular RNAs: Promising Molecular Biomarkers of Human Aging-Related Diseases via Functioning as an miRNA Sponge. Mol. Ther. Methods Clin. Dev. 2020, 18, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Lazzarini, R.; Babini, L.; Prattichizzo, F.; Rippo, M.R.; Tiano, L.; Di Nuzzo, S.; Graciotti, L.; Festa, R.; Brugè, F.; et al. Anti-inflammatory effect of ubiquinol-10 on young and senescent endothelial cells via miR-146a modulation. Free Radic. Biol. Med. 2013, 63, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Rippo, M.R.; Olivieri, F.; Monsurrò, V.; Prattichizzo, F.; Albertini, M.C.; Procopio, A.D. MitomiRs in human inflamm-aging: A hypothesis involving miR-181a, miR-34a and miR-146a. Exp. Gerontol. 2014, 56, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Bi, Q.; Goodman, K.E.; Kaminsky, J.; Lessler, J. What is Machine Learning? A Primer for the Epidemiologist. Am. J. Epidemiol. 2019, 188, 2222–2239. [Google Scholar] [CrossRef]
- Khatri, S.; Arora, A.; Prakash, A. Supervised Machine Learning Algorithms for Credit Card Fraud Detection: A Comparison. In Proceedings of the 2020 10th International Conference on Cloud Computing, Data Science & Engineering (Confluence), Noida, India, 29–31 January 2020; pp. 680–683. [Google Scholar] [CrossRef]
- Aiello, A.; Accardi, G.; Aprile, S.; Caldarella, R.; Carru, C.; Ciaccio, M.; De Vivo, I.; Gambino, C.M.; Ligotti, M.E.; Vasto, S.; et al. Age and Gender-related Variations of Molecular and Phenotypic Parameters in A Cohort of Sicilian Population: From Young to Centenarians. Aging Dis. 2021, 12, 1773–1793. [Google Scholar] [CrossRef]
- Boisen, L.; Drasbek, K.R.; Pedersen, A.S.; Kristensen, P. Evaluation of endothelial cell culture as a model system of vascular ageing. Exp. Gerontol. 2010, 45, 779–787. [Google Scholar] [CrossRef] [Green Version]
- miRWalk. Home—miRWalk. Available online: uni-heidelberg.de (accessed on 20 February 2022).
- KEGG Pathway. KEGG Overview. Available online: genome.jp (accessed on 20 February 2022).
- Kanehisa, M.; Sato, Y.; Furumichi, M.; Morishima, K.; Tanabe, M. New approach for understanding genome variations in KEGG. Nucleic Acids Res. 2019, 47, D590–D595. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, P.; Amador, S.; Lucas-Ochoa, A.M.; Cuenca-Bermejo, L.; Fernández-Villalba, E.; Raparelli, V.; Norris, C.; Kautzky-Willer, A.; Kublickiene, K.; Pilote, L.; et al. Sex, rurality and socioeconomical status in Spanish centennial population (2017). Aging 2021, 13, 22059–22077. [Google Scholar] [CrossRef]
- Quinlan, J.R. Induction of Decision Trees. Mach. Learn. 1986, 1, 81–106. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Zhang, J.; Zhou, Y.; Guo, X.; Ma, Y. A feature selection algorithm of decision tree based on feature weight. Expert Syst. Appl. 2021, 164, 113842. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Sahin, U.; de Thé, H.; Lallemand-Breitenbach, V. Sumoylation in Physiology, Pathology and Therapy. Cells 2022, 11, 814. [Google Scholar] [CrossRef] [PubMed]
- Caruso, C.; Accardi, G.; Ligotti, M.E.; Vasto, S.; Candore, G. Pathobiology of aging: An introduction to age-related diseases. In Human Aging; Caruso, C., Candore, G., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 35–74. ISBN 9780128225691. [Google Scholar]
- Accardi, G.; Ligotti, M.E.; Candore, G. Phenotypic Aspects of Longevity. In Centenarians; Caruso, C., Ed.; Springer: Cham, Switzerland, 2019; ISBN 978-3-030-20762-5. [Google Scholar]
- Caruso, C.; Passarino, G.; Puca, A.; Scapagnini, G. “Positive biology”: The centenarian lesson. Immun. Ageing 2012, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameling, S.; Kacprowski, T.; Chilukoti, R.K.; Malsch, C.; Liebscher, V.; Suhre, K.; Pietzner, M.; Friedrich, N.; Homuth, G.; Hammer, E.; et al. Associations of circulating plasma microRNAs with age, body mass index and sex in a population-based study. BMC Med. Genom. 2015, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; Luettich, K. MicroRNAs as potential biomarkers of smoking-related diseases. Biomark. Med. 2012, 6, 671–684. [Google Scholar] [CrossRef] [Green Version]
- Aiello, A.; Accardi, G.; Aprile, S.; Caldarella, R.; Cammarata, G.; Carru, C.; Caruso, C.; Ciaccio, M.; De Vivo, I.; Gambino, C.M.; et al. Pro-inflammatory status is not a limit for longevity: Case report of a Sicilian centenarian. Aging Clin. Exp. Res. 2021, 33, 1403–1407. [Google Scholar] [CrossRef]
- Ong, J.; Woldhuis, R.R.; Boudewijn, I.M.; van den Berg, A.; Kluiver, J.; Kok, K.; Terpstra, M.M.; Guryev, V.; de Vries, M.; Vermeulen, C.J.; et al. Age-related gene and miRNA expression changes in airways of healthy individuals. Sci. Rep. 2019, 9, 3765. [Google Scholar] [CrossRef] [Green Version]
- Lescai, F.; Marchegiani, F.; Franceschi, C. PON1 is a longevity gene: Results of a meta-analysis. Ageing Res. Rev. 2009, 8, 277–284. [Google Scholar] [CrossRef]
- Arbeev, K.G.; Verhulst, S.; Steenstrup, T.; Kark, J.D.; Bagley, O.; Kooperberg, C.; Reiner, A.P.; Hwang, S.J.; Levy, D.; Fitzpatrick, A.L.; et al. Association of Leukocyte Telomere Length With Mortality Among Adult Participants in 3 Longitudinal Studies. JAMA Netw. Open 2020, 3, 200023. [Google Scholar] [CrossRef] [Green Version]
- Boldin, M.P.; Baltimore, D. MicroRNAs, new effectors and regulators of NF-κB. Immunol. Rev. 2012, 246, 205–220. [Google Scholar] [CrossRef]
- Olivieri, F.; Lazzarini, R.; Recchioni, R.; Marcheselli, F.; Rippo, M.R.; Di Nuzzo, S.; Albertini, M.C.; Graciotti, L.; Babini, L.; Mariotti, S.; et al. MiR-146a as marker of senescence-associated pro-inflammatory status in cells involved in vascular remodelling. Age 2013, 35, 1157–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roganović, J. Downregulation of microRNA-146a in diabetes, obesity and hypertension may contribute to severe COVID-19. Med. Hypotheses 2021, 146, 110448. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.; Gaetani, S.; Sorgentoni, G.; Agarbati, S.; Laggetta, M.; Matacchione, G.; Gobbi, M.; Rossi, T.; Galeazzi, R.; Piccinini, G.; et al. Circulating Inflamma-miRs as Potential Biomarkers of Cognitive Impairment in Patients Affected by Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 647015. [Google Scholar] [CrossRef] [PubMed]
- Bonafè, M.; Olivieri, F. Circulating microRNAs in aging. Oncotarget 2015, 6, 6–1340. [Google Scholar] [CrossRef] [Green Version]
- Noren Hooten, N.; Fitzpatrick, M.; Wood, W.H., III; De, S.; Ejiogu, N.; Zhang, Y.; Mattison, J.A.; Becker, K.G.; Zonderman, A.B.; Evans, M.K. Age-related changes in microRNA levels in serum. Aging 2013, 5, 725–740. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Li, Z.; Li, M.; Xu, N.; Zhang, Y. miR-181a and inflammation: miRNA homeostasis response to inflammatory stimuli in vivo. Biochem. Biophys. Res. Commun. 2013, 430, 647–652. [Google Scholar] [CrossRef]
- Su, Y.; Yuan, J.; Zhang, F.; Lei, Q.; Zhang, T.; Li, K.; Guo, J.; Hong, Y.; Bu, G.; Lv, X.; et al. MicroRNA-181a-5p and microRNA-181a-3p cooperatively restrict vascular inflammation and atherosclerosis. Cell Death Dis. 2019, 10, 365. [Google Scholar] [CrossRef] [Green Version]
- Flachsbart, F.; Dose, J.; Gentschew, L.; Geismann, C.; Caliebe, A.; Knecht, C.; Nygaard, M.; Badarinarayan, N.; ElSharawy, A.; May, S.; et al. Identification and characterization of two functional variants in the human longevity gene FOXO3. Nat. Commun. 2017, 8, 2063. [Google Scholar] [CrossRef] [Green Version]
- Princz, A.; Tavernarakis, N. The role of SUMOylation in ageing and senescent decline. Mech. Ageing Dev. 2017, 162, 85–90. [Google Scholar] [CrossRef]
- Gong, L.; Sun, Q.; Li, D.W. Sumoylation in Cellular Senescence and Aging. Curr. Mol. Med. 2017, 16, 871–876. [Google Scholar] [CrossRef]
- Olivieri, F.; Bonafè, M.; Spazzafumo, L.; Gobbi, M.; Prattichizzo, F.; Recchioni, R.; Marcheselli, F.; La Sala, L.; Galeazzi, R.; Rippo, M.R.; et al. Age- and glycemia-related miR-126-3p levels in plasma and endothelial cells. Aging 2014, 6, 771–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | (a) Young Adults (22–50 y.o.) N = 19, M = 7, W = 12 | (b) Adults (51–70 y.o.) N = 28, M = 14, W = 14 | (c) Older Adults (71–99 y.o.) N = 20, M = 13, W = 7 | (d) Ultracentenarians (100–111 y.o.) N = 11, M = 2, W = 9 | p-Value of Kruskal–Wallis Test | p-Value Bonferroni Test | |||||||||
| mean | median | SD | mean | median | SD | mean | median | SD | mean | median | SD | p = 0.0003 | (a vs. b), p = 0.0028; (a vs. c), p = 0.0284; (b vs. d), p = 0.0024; (c vs. d), p = 0.0171. | ||
| miR-21-5p | N | 8.22 | 4.96 | 6.45 | 18.87 | 17.34 | 11.06 | 16.73 | 17.47 | 10.74 | 5.73 | 4.01 | 4.58 | ||
| M | 7.71 | 4.96 | 6.35 | 19.56 | 18.07 | 13.24 | 15.40 | 17.32 | 8.75 | 3.05 | 3.05 | 1.36 | |||
| W | 8.51 | 7.50 | 6.78 | 18.18 | 17.34 | 8.83 | 19.19 | 27.77 | 14.19 | 6.33 | 4.43 | 4.88 | |||
| miR-126-3p | N | 5.70 | 5.10 | 2.16 | 7.58 | 6.33 | 4.54 | 10.77 | 9.36 | 7.33 | 2.67 | 1.80 | 1.62 | p = 0.0002 | (a vs. d), p = 0.0238; (b vs. d), p = 0.0005; (c vs. d), p = 0.0001. |
| M | 6.27 | 6.06 | 2.19 | 6.98 | 5.84 | 4.80 | 11.85 | 12.36 | 8.21 | 3.72 | 3.72 | 1.46 | |||
| W | 5.37 | 4.49 | 2.17 | 8.18 | 6.71 | 4.35 | 8.76 | 6.35 | 5.30 | 2.44 | 1.68 | 1.64 | |||
| miR-146a-5p | N | 3.34 | 3.57 | 0.94 | 3.34 | 3.57 | 0.94 | 5.67 | 3.79 | 5.63 | 6.23 | 3.85 | 5.63 | p = 0.0491 | (a vs. b), p = 0.0177. |
| M | 3.11 | 3.57 | 0.78 | 5.41 | 3.71 | 4.92 | 5.05 | 3.43 | 6.23 | 4.60 | 4.60 | 2.43 | |||
| W | 3.48 | 3.32 | 1.03 | 6.13 | 5.66 | 2.47 | 6.83 | 5.41 | 4.52 | 6.60 | 3.85 | 6.17 | |||
| miR-181a-5p | N | 3.11 | 3.28 | 1.03 | 7.22 | 5.41 | 5.52 | 5.27 | 5.07 | 2.85 | 9.30 | 4.93 | 10.36 | p = 0.0024 | (a vs. b), p = 0.0006; (a vs. c), p = 0.0358. |
| M | 2.87 | 2.82 | 0.78 | 6.30 | 4.83 | 4.73 | 4.99 | 4.03 | 3.12 | 4.65 | 4.65 | 0.40 | |||
| W | 3.25 | 3.43 | 1.16 | 8.15 | 6.62 | 6.24 | 5.80 | 5.19 | 2.39 | 10.34 | 10.47 | 11.30 | |||
| R2 = 0.346 | Coefficient | p |
|---|---|---|
| Age | 1.46 | <0.0005 |
| Age2 | −0.01 | <0.0005 |
| BMI | −0.20 | =0.54 |
| Smoke | ||
| Smoker (reference) | ||
| Ex-smokers | 1.94 | =0.57 |
| Never smoked | 5.24 | =0.06 |
| Gender | ||
| M (reference) | ||
| F | −2.02 | =0.40 |
| b0 (constant) | −23.32 | =0.01 |
| miR-126-3p | miR-146a-5p | miR-181a-5p | ||||
|---|---|---|---|---|---|---|
| Coefficient | p | Coefficient | p | Coefficient | p | |
| Age Class | ||||||
| 22–50 (Reference) | ||||||
| 51–70 | 2.08 | =0.14 | 2.84 | <0.0005 | 4.23 | 0.00 |
| 71–99 | 4.37 | =0.05 | 2.39 | =0.16 | 1.61 | 0.30 |
| 100–111 | −3.63 | <0.0005 | 1.15 | =0.34 | 5.28 | 0.13 |
| BMI | 0.07 | =0.61 | −0.05 | =0.74 | −0.01 | 0.95 |
| Smoke | ||||||
| Smoker (Reference) | ||||||
| Ex-smoker | 2.05 | =0.25 | 2.14 | =0.10 | 4.13 | 0.00 |
| Never smoked | 2.91 | =0.04 | 1.36 | =0.06 | 1.93 | 0.01 |
| Gender | ||||||
| M (Reference) | ||||||
| F | −0.85 | =0.49 | 0.72 | =0.46 | 1.63 | 0.16 |
| b0 | 2.50 | =0.49 | 2.95 | =0.44 | 0.68 | 0.87 |
| R2 = 0.2779 | R2 = 0.116 | R2 = 0.2169 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Accardi, G.; Bono, F.; Cammarata, G.; Aiello, A.; Herrero, M.T.; Alessandro, R.; Augello, G.; Carru, C.; Colomba, P.; Costa, M.A.; et al. miR-126-3p and miR-21-5p as Hallmarks of Bio-Positive Ageing; Correlation Analysis and Machine Learning Prediction in Young to Ultra-Centenarian Sicilian Population. Cells 2022, 11, 1505. https://doi.org/10.3390/cells11091505
Accardi G, Bono F, Cammarata G, Aiello A, Herrero MT, Alessandro R, Augello G, Carru C, Colomba P, Costa MA, et al. miR-126-3p and miR-21-5p as Hallmarks of Bio-Positive Ageing; Correlation Analysis and Machine Learning Prediction in Young to Ultra-Centenarian Sicilian Population. Cells. 2022; 11(9):1505. https://doi.org/10.3390/cells11091505
Chicago/Turabian StyleAccardi, Giulia, Filippa Bono, Giuseppe Cammarata, Anna Aiello, Maria Trinidad Herrero, Riccardo Alessandro, Giuseppa Augello, Ciriaco Carru, Paolo Colomba, Maria Assunta Costa, and et al. 2022. "miR-126-3p and miR-21-5p as Hallmarks of Bio-Positive Ageing; Correlation Analysis and Machine Learning Prediction in Young to Ultra-Centenarian Sicilian Population" Cells 11, no. 9: 1505. https://doi.org/10.3390/cells11091505
APA StyleAccardi, G., Bono, F., Cammarata, G., Aiello, A., Herrero, M. T., Alessandro, R., Augello, G., Carru, C., Colomba, P., Costa, M. A., De Vivo, I., Ligotti, M. E., Lo Curto, A., Passantino, R., Taverna, S., Zizzo, C., Duro, G., Caruso, C., & Candore, G. (2022). miR-126-3p and miR-21-5p as Hallmarks of Bio-Positive Ageing; Correlation Analysis and Machine Learning Prediction in Young to Ultra-Centenarian Sicilian Population. Cells, 11(9), 1505. https://doi.org/10.3390/cells11091505