
Tany-Seq: Integrated Analysis of the Mouse Tanycyte Transcriptome


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hypothalamic Single Cell RNA Sequencing Dataset Transformation, Filtering, and Clustering
2.2. Cell Population Identification and Integration of Tanycyte Cell Clusters
2.3. Gene Ontology Analysis
2.4. Hindbrain and Neuronal Progenitor Transformation, Filtering, and Clustering
2.5. Mapping
2.6. Comparison of Transcript Expression between Tanycyte and Tanycyte-Like-Cells
2.7. Specific Sample Usage and Package Versions
3. Results
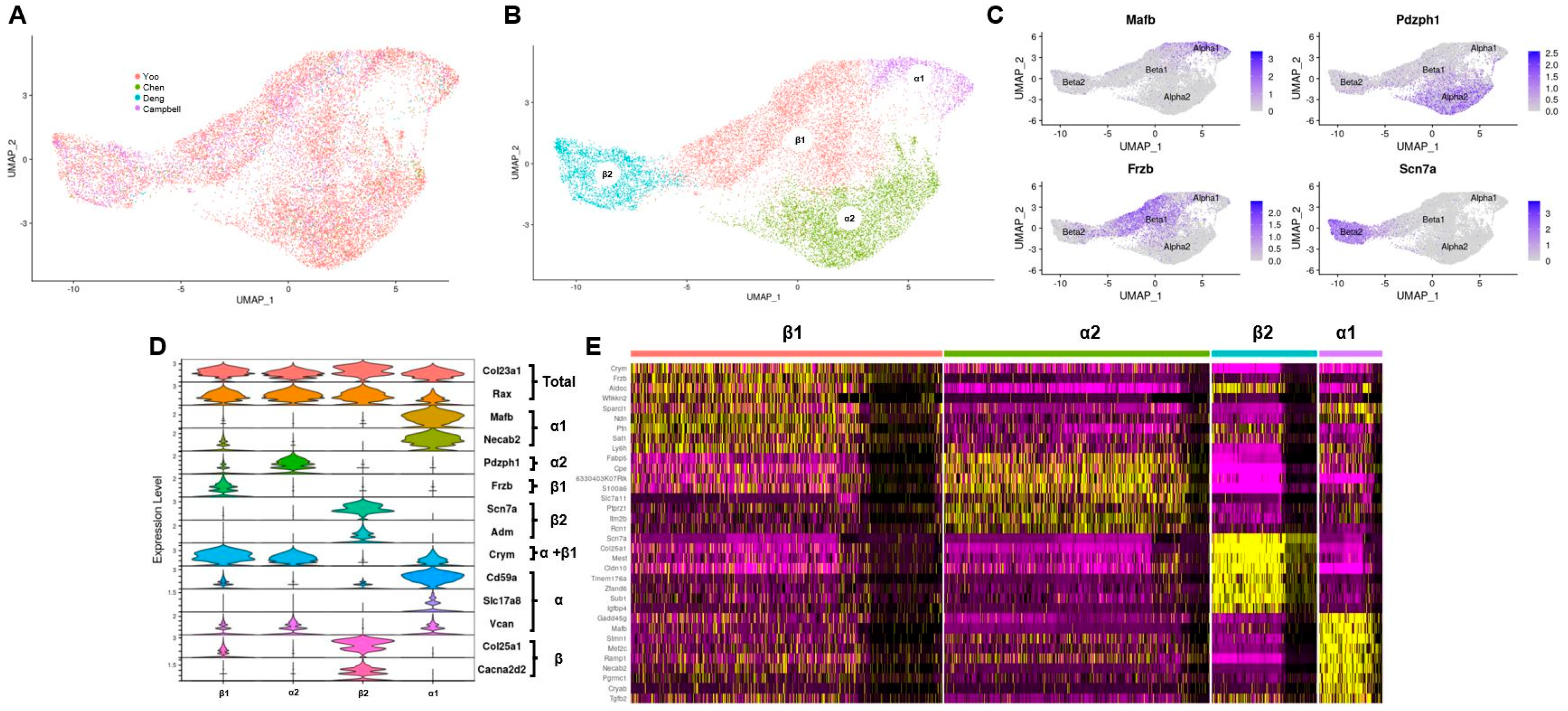
3.1. Integration of Tanycytes and Verification of Identity
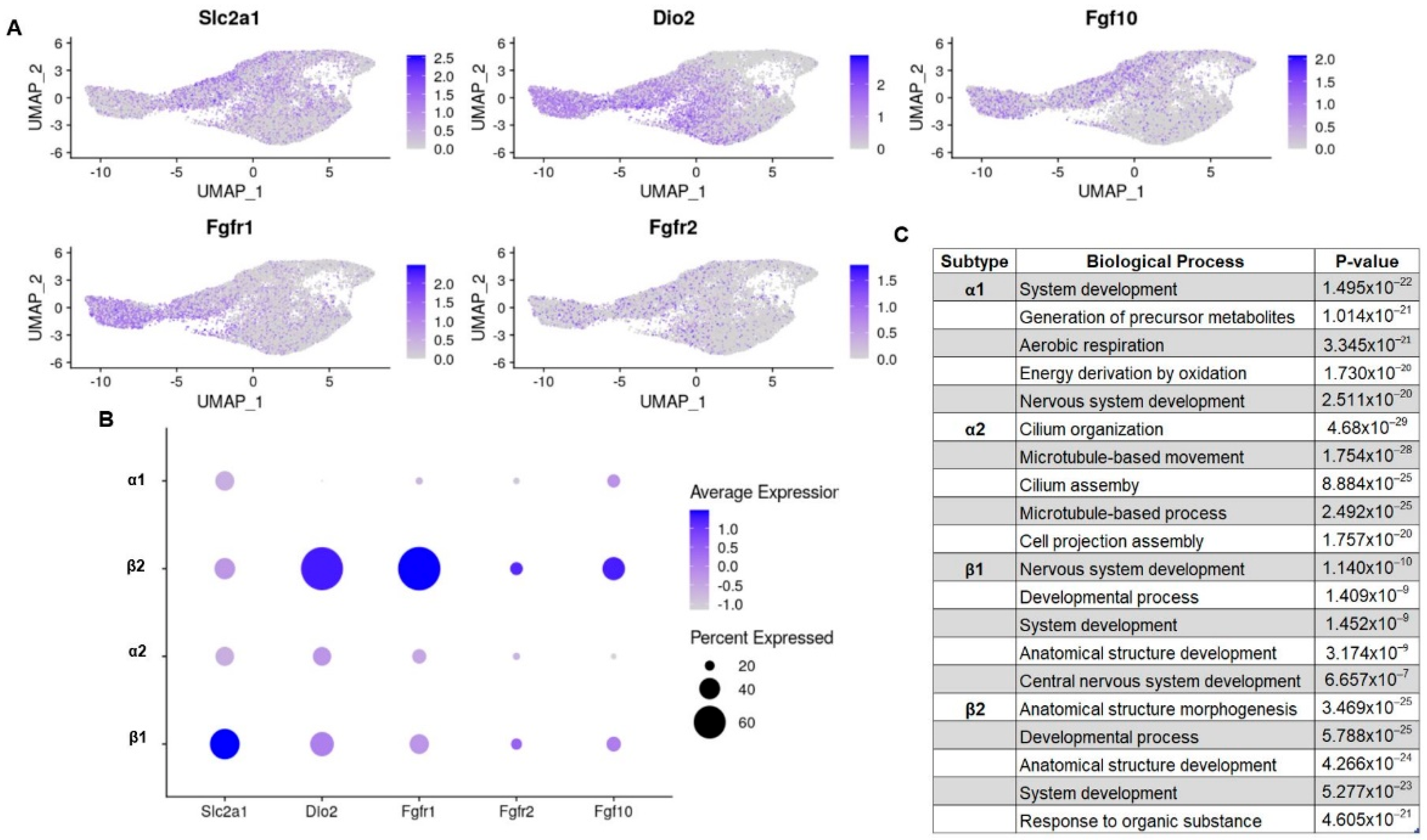
3.2. Transcripts Relevant to Tanycyte Phenotypes Have Subtype Specific Expression
3.2.1. Glucose-Sensing and Energy Balance
3.2.2. Thyroid Hormone Regulation and Import
3.2.3. Gene Ontology
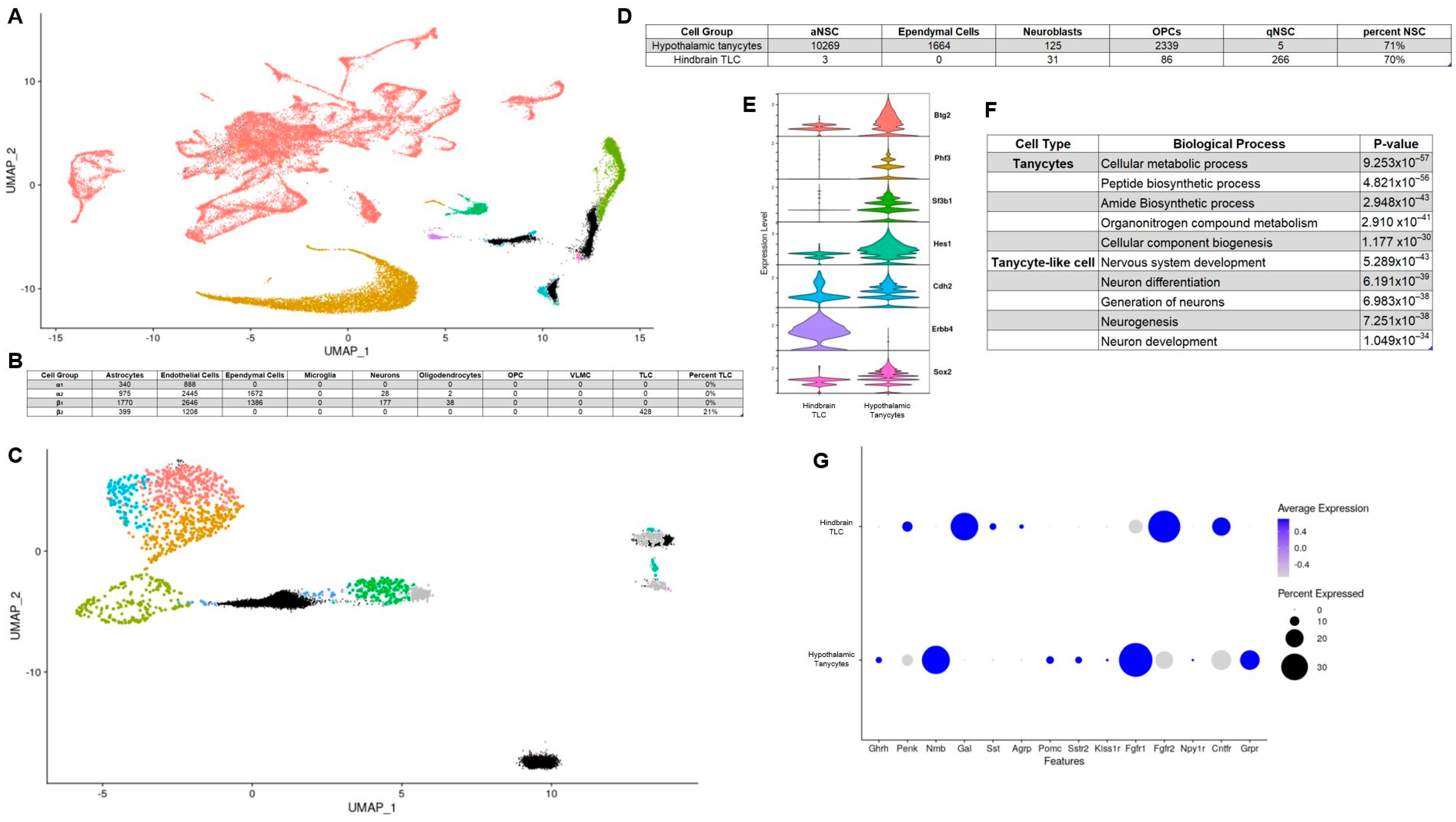
3.3. Hypothalamic Tanycytes and Purported NTS Tanycyte-Like Cells Lack Similar Transcriptomic Profiles and Are Primed to Fulfil Distinct Biological Functions
4. Discussion
5. Limitations
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luquet, S.; Magnan, C. The central nervous system at the core of the regulation of energy homeostasis. Front. Biosci. (Sch. Ed.) 2009, 1, 448–465. [Google Scholar] [CrossRef] [PubMed]
- Langlet, F. Tanycyte Gene Expression Dynamics in the Regulation of Energy Homeostasis. Front. Endocrinol. 2019, 10, 286. [Google Scholar] [CrossRef] [Green Version]
- Bolborea, M.; Langlet, F. What is the physiological role of hypothalamic tanycytes in metabolism? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2021, 320, R994–R1003. [Google Scholar] [CrossRef] [PubMed]
- Timper, K.; Brüning, J.C. Hypothalamic circuits regulating appetite and energy homeostasis: Pathways to obesity. Dis. Model. Mech. 2017, 10, 679–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roh, E.; Song, D.K.; Kim, M.-S. Emerging role of the brain in the homeostatic regulation of energy and glucose metabolism. Exp. Mol. Med. 2016, 48, e216. [Google Scholar] [CrossRef] [Green Version]
- Rui, L. Brain regulation of energy balance and body weight. Rev. Endocr. Metab. Disord. 2013, 14, 387–407. [Google Scholar] [CrossRef] [Green Version]
- Mullier, A.; Bouret, S.G.; Prevot, V.; Dehouck, B. Differential distribution of tight junction proteins suggests a role for tanycytes in blood-hypothalamus barrier regulation in the adult mouse brain. J. Comp. Neurol. 2010, 518, 943–962. [Google Scholar] [CrossRef]
- Pasquettaz, R.; Kolotuev, I.; Rohrbach, A.; Gouelle, C.; Pellerin, L.; Langlet, F. Peculiar protrusions along tanycyte processes face diverse neural and nonneural cell types in the hypothalamic parenchyma. J. Comp. Neurol. 2020, 529, 553–575. [Google Scholar] [CrossRef]
- Yoo, S.; Kim, J.; Lyu, P.; Hoang, T.V.; Ma, A.; Trinh, V.; Dai, W.; Jiang, L.; Leavey, P.; Duncan, L.; et al. Control of neurogenic competence in mammalian hypothalamic tanycytes. Sci. Adv. 2021, 7, eabg3777. [Google Scholar] [CrossRef]
- Campbell, J.N.; Macosko, E.Z.; Fenselau, H.; Pers, T.H.; Lyubetskaya, A.; Tenen, D.; Goldman, M.; Verstegen, A.M.; Resch, J.M.; McCarroll, S.A.; et al. A molecular census of arcuate hypothalamus and median eminence cell types. Nat. Neurosci. 2017, 20, 484–496. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Wu, X.; Jiang, L.; Zhang, Y. Single-Cell RNA-Seq Reveals Hypothalamic Cell Diversity. Cell Rep. 2017, 18, 3227–3241. [Google Scholar] [CrossRef] [PubMed]
- Deng, G.; Morselli, L.L.; Wagner, V.A.; Balapattabi, K.; Sapouckey, S.A.; Knudtson, K.L.; Rahmouni, K.; Cui, H.; Sigmund, C.D.; Kwitek, A.E.; et al. Single-Nucleus RNA Sequencing of the Hypothalamic Arcuate Nucleus of C57BL/6J Mice after Prolonged Diet-Induced Obesity. Hypertension 2020, 76, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Barahona, M.J.; Llanos, P.; Recabal, A.; Escobar-Acuña, K.; Elizondo-Vega, R.; Salgado, M.; Ordenes, P.; Uribe, E.; Sepúlveda, F.J.; Araneda, R.; et al. Glial hypothalamic inhibition of GLUT2 expression alters satiety, impacting eating behavior. Glia 2017, 66, 592–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uranga, R.M.; Millán, C.; Barahona, M.J.; Recabal, A.; Salgado, M.; Martinez, F.; Ordenes, P.; Elizondo-Vega, R.; Sepúlveda, F.; Uribe, E.; et al. Adenovirus-mediated suppression of hypothalamic glucokinase affects feeding behavior. Sci. Rep. 2017, 7, 3697. [Google Scholar] [CrossRef] [Green Version]
- Peruzzo, B.; Pastor, F.E.; Blázquez, J.L.; Schöbitz, K.; Peláez, B.; Amat, P.; Rodríguez, E.M. A second look at the barriers of the medial basal hypothalamus. Exp. Brain Res. 2000, 132, 10–26. [Google Scholar] [CrossRef]
- Garcia, M.D.L.A.; Carrasco, M.; Godoy, A.; Reinicke, K.; Montecinos, V.P.; Aguayo, L.G.; Tapia, J.C.; Vera, J.C.; Nualart, F. Elevated expression of glucose transporter-1 in hypothalamic ependymal cells not involved in the formation of the brain-cerebrospinal fluid barrier. J. Cell. Biochem. 2001, 80, 491–503. [Google Scholar] [CrossRef]
- Bentsen, M.A.; Rausch, D.M.; Mirzadeh, Z.; Muta, K.; Scarlett, J.M.; Brown, J.M.; Herranz-Pérez, V.; Baquero, A.F.; Thompson, J.; Alonge, K.M.; et al. Transcriptomic analysis links diverse hypothalamic cell types to fibroblast growth factor 1-induced sustained diabetes remission. Nat. Commun. 2020, 11, 4458. [Google Scholar] [CrossRef]
- Kaminskas, B.; Goodman, T.; Hagan, A.; Bellusci, S.; Ornitz, D.M.; Hajihosseini, M.K. Characterisation of endogenous players in fibroblast growth factor-regulated functions of hypothalamic tanycytes and energy-balance nuclei. J. Neuroendocr. 2019, 31, e12750. [Google Scholar] [CrossRef]
- Haan, N.; Goodman, T.; Najdi-Samiei, A.; Stratford, C.M.; Rice, R.; El Agha, E.; Bellusci, S.; Hajihosseini, M.K. Fgf10-Expressing Tanycytes Add New Neurons to the Appetite/Energy-Balance Regulating Centers of the Postnatal and Adult Hypothalamus. J. Neurosci. 2013, 33, 6170–6180. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.E. Thyroid Hormone Control of Thermogenesis and Energy Balance. Thyroid 1995, 5, 481–492. [Google Scholar] [CrossRef]
- Coppola, A.; Liu, Z.-W.; Andrews, Z.B.; Paradis, E.; Roy, M.-C.; Friedman, J.M.; Ricquier, D.; Richard, D.; Horvath, T.L.; Gao, X.-B.; et al. A Central Thermogenic-like Mechanism in Feeding Regulation: An Interplay between Arcuate Nucleus T3 and UCP2. Cell Metab. 2007, 5, 21–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.; Ebling, F.J.P. The Role of Hypothalamic Tri-Iodothyronine Availability in Seasonal Regulation of Energy Balance and Body Weight. J. Thyroid Res. 2011, 2011, 387562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, H.M.; Kim, S.W.; Salvatore, D.; Bartha, T.; Legradi, G.; Larsen, P.R.; Lechan, R.M. Regional distribution of type 2 thyroxine deiodinase messenger ribonucleic acid in rat hypothalamus and pituitary and its regulation by thyroid hormone. Endocrinology 1997, 138, 3359–3368. [Google Scholar] [CrossRef]
- Müller-Fielitz, H.; Stahr, M.; Bernau, M.; Richter, M.; Abele, S.; Krajka, V.; Benzin, A.; Wenzel, J.; Kalies, K.; Mittag, J.; et al. Tanycytes control the hormonal output of the hypothalamic-pituitary-thyroid axis. Nat. Commun. 2017, 8, 484. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.Q.; Cheng, W.; Gordian, D.; Lee, J.; Paulsen, S.J.; Hansen, S.N.; Egerod, K.L.; Barkholt, P.; Rhodes, C.J.; Secher, A.; et al. A genetic map of the mouse dorsal vagal complex and its role in obesity. Nat. Metab. 2021, 3, 530–545. [Google Scholar] [CrossRef] [PubMed]
- Dowsett, G.K.C.; Lam, B.Y.H.; Tadross, J.A.; Cimino, I.; Rimmington, D.; Coll, A.P.; Polex-Wolf, J.; Knudsen, L.B.; Pyke, C.; Yeo, G.S.H. A survey of the mouse hindbrain in the fed and fasted states using single-nucleus RNA sequencing. Mol. Metab. 2021, 53, 101240. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.T.; Stratton, J.; Stykel, M.; Abbasi, S.; Sharma, S.; Mayr, K.; Koblinger, K.; Whelan, P.; Biernaskie, J. Single-Cell Transcriptomics and Fate Mapping of Ependymal Cells Reveals an Absence of Neural Stem Cell Function. Cell 2018, 173, 1045–1057.e9. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sullivan, A.I.; Potthoff, M.J.; Flippo, K.H. Tany-Seq: Integrated Analysis of the Mouse Tanycyte Transcriptome. Cells 2022, 11, 1565. https://doi.org/10.3390/cells11091565
Sullivan AI, Potthoff MJ, Flippo KH. Tany-Seq: Integrated Analysis of the Mouse Tanycyte Transcriptome. Cells. 2022; 11(9):1565. https://doi.org/10.3390/cells11091565
Chicago/Turabian StyleSullivan, Andrew I., Matthew J. Potthoff, and Kyle H. Flippo. 2022. "Tany-Seq: Integrated Analysis of the Mouse Tanycyte Transcriptome" Cells 11, no. 9: 1565. https://doi.org/10.3390/cells11091565
APA StyleSullivan, A. I., Potthoff, M. J., & Flippo, K. H. (2022). Tany-Seq: Integrated Analysis of the Mouse Tanycyte Transcriptome. Cells, 11(9), 1565. https://doi.org/10.3390/cells11091565