
Growth Response and Differentiation of Bone Marrow-Derived Mesenchymal Stem/Stromal Cells in the Presence of Novel Multiple Myeloma Drug Melflufen

 ,
,  , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bone Marrow-Derived Mesenchymal Stem Cell Isolation and Culture
2.2. Metabolic Activity and Cytotoxicity
2.3. Proliferation Assay
2.4. Indirect Co-cultures of BMSCs and Myeloma Cell Line MM.1S
2.5. Adipogenic Differentiation
2.6. Osteogenic Differentiation
2.7. Angiogenesis Assay
2.8. Immunocytochemistry
2.9. Quantification of Endothelial Cell and Pericyte Coverage Using Image Analysis
2.10. Quantified Real Time-Polymerase Chain Reaction (qRT-PCR)
2.11. Statistical Analyses
3. Results
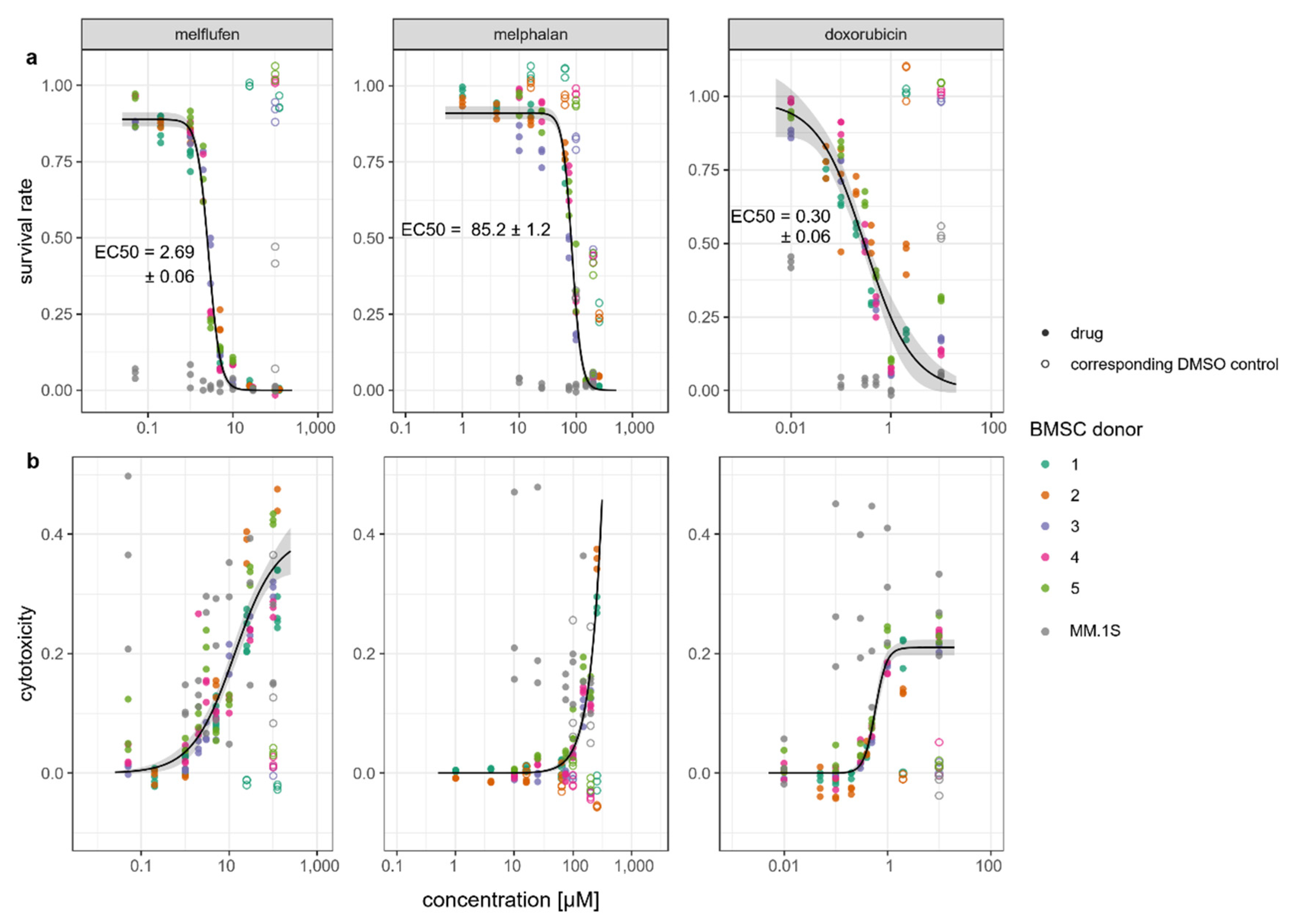
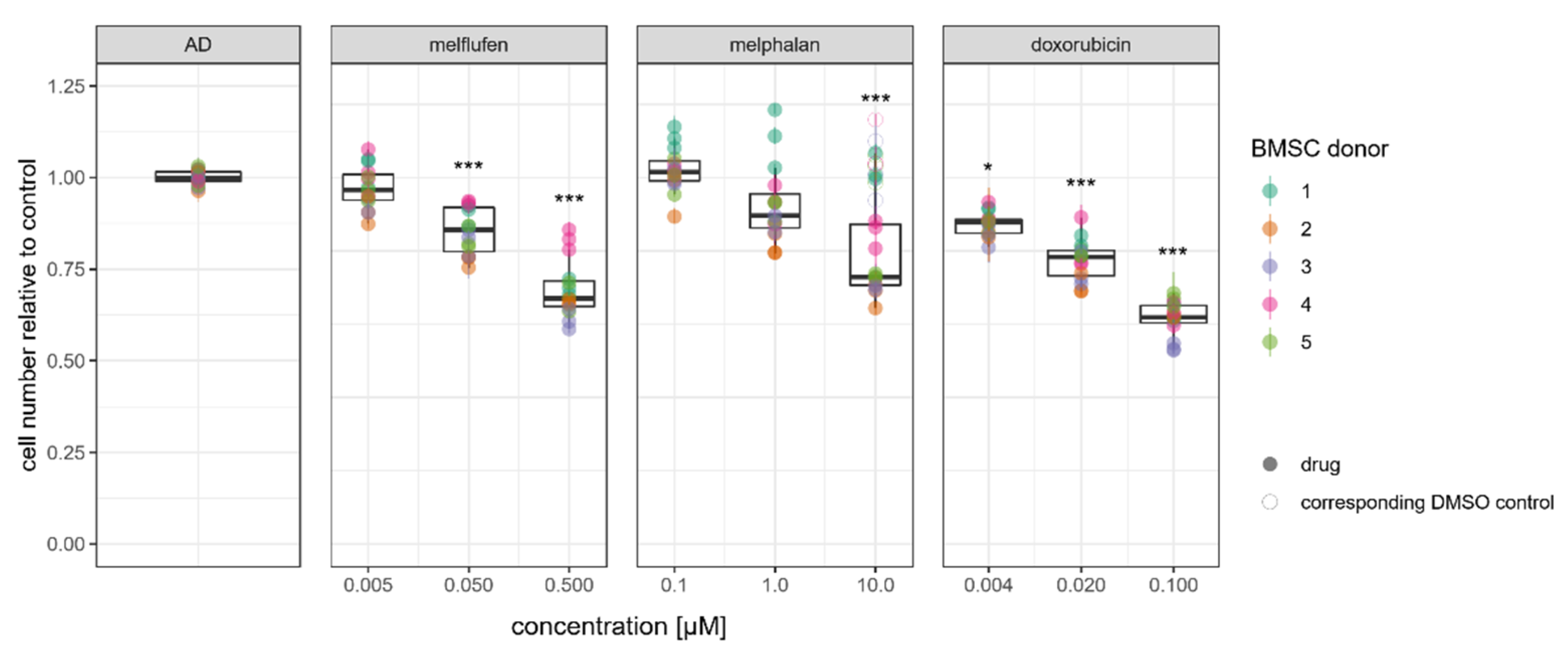
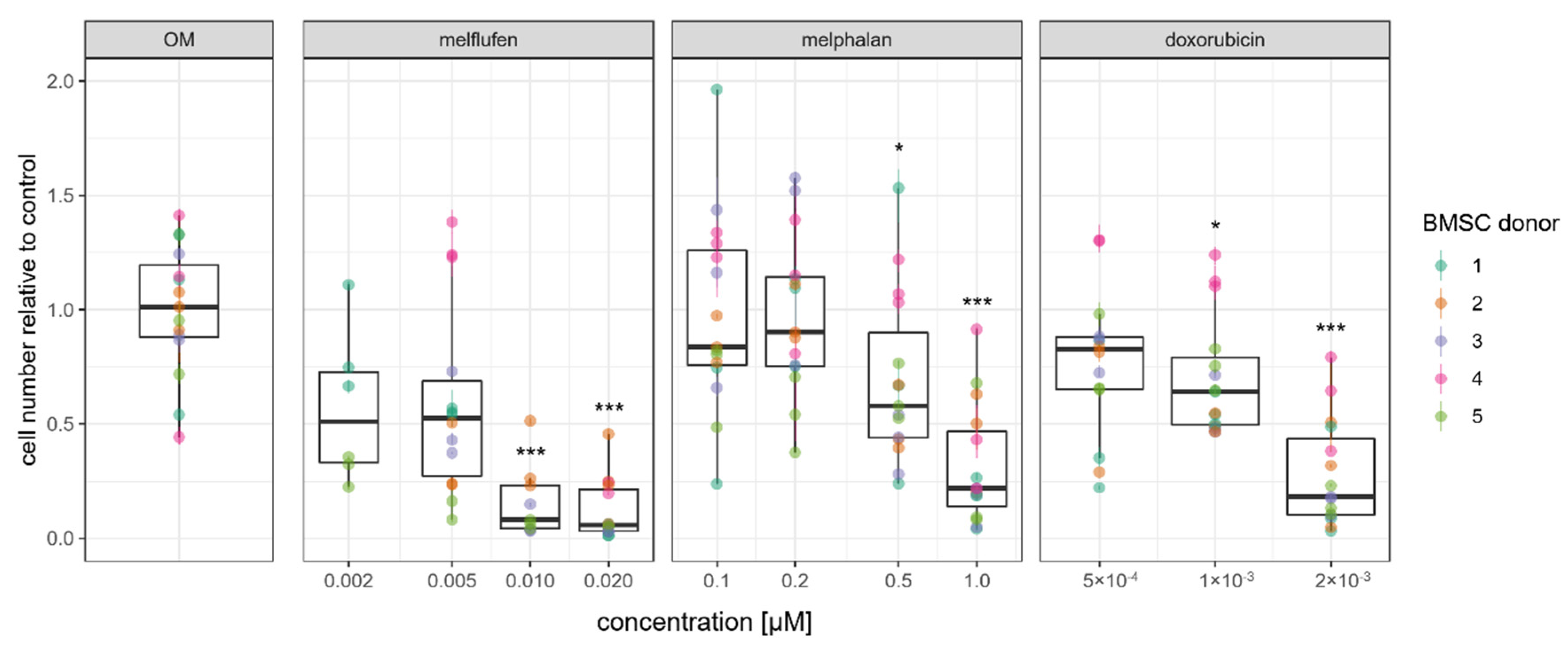
3.1. Viability, Cytotoxic Response, and Proliferation of BMSCs in the Presence of Standard-of-Care Drugs for Multiple Myeloma
3.2. Myeloma Cytotoxic Response in the Presence of BMSCs
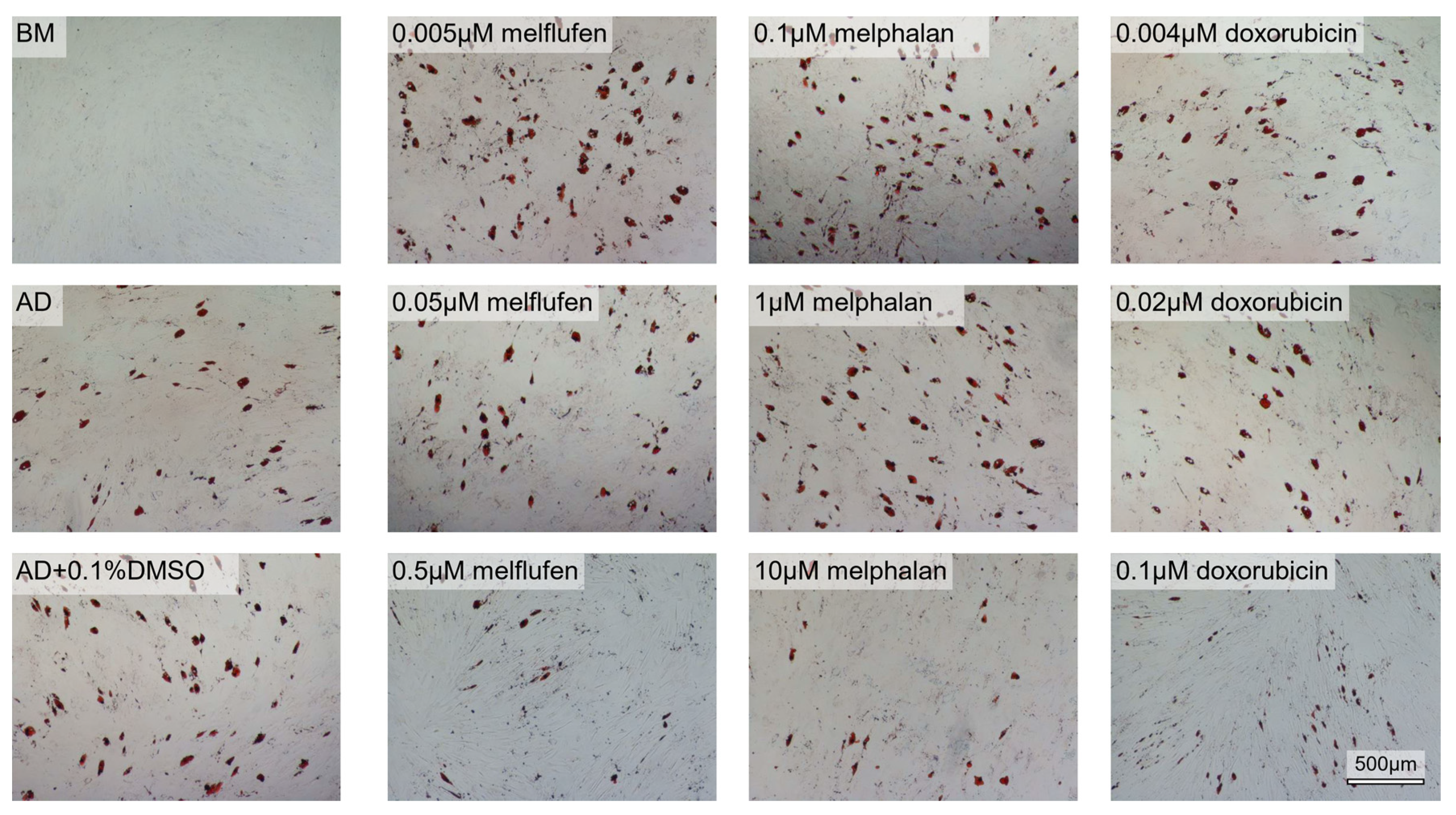
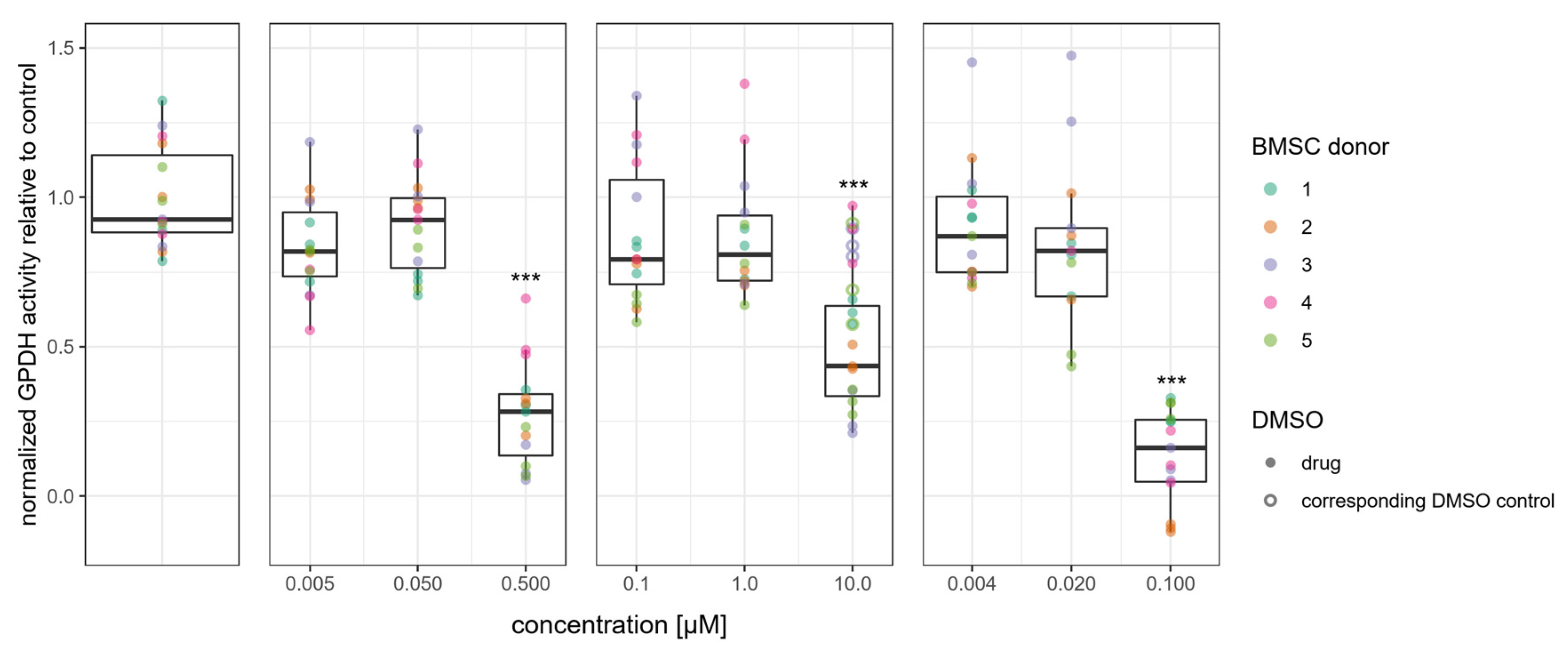
3.3. Adipogenic Differentiation
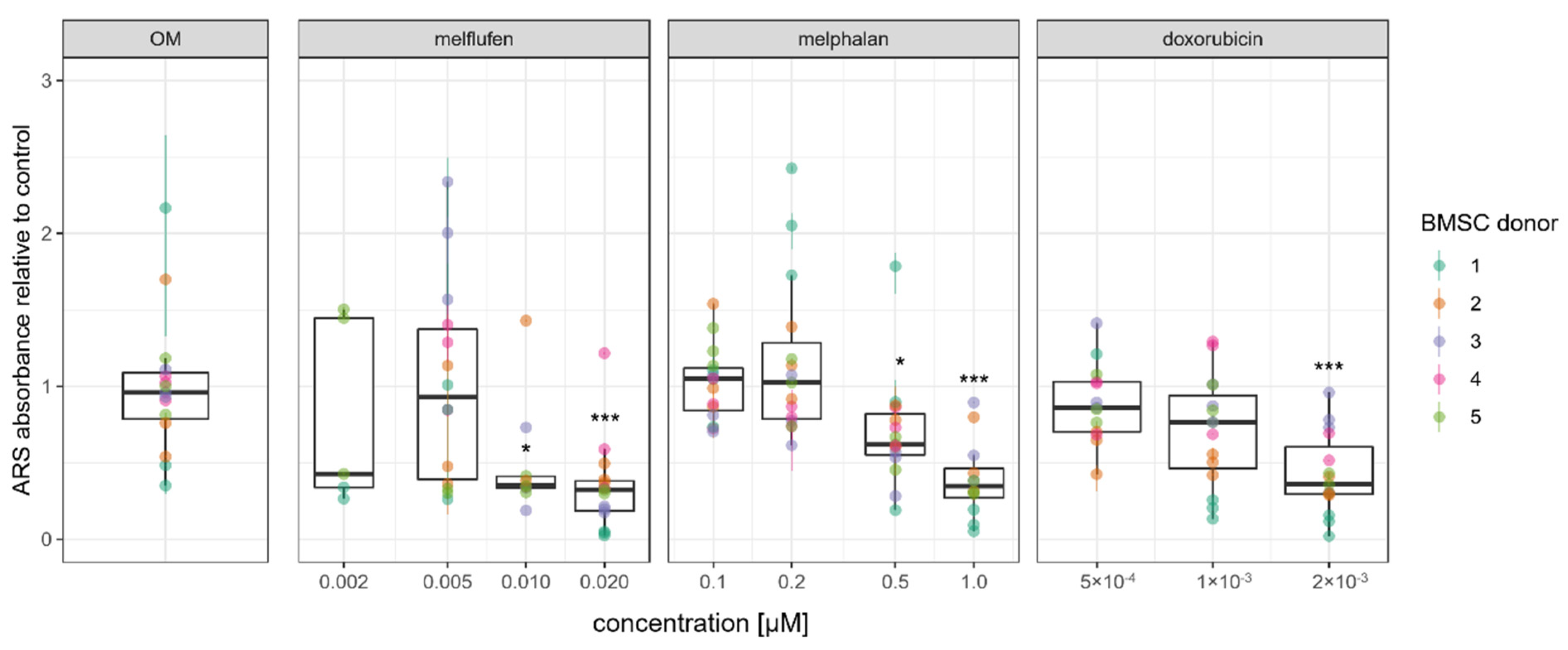
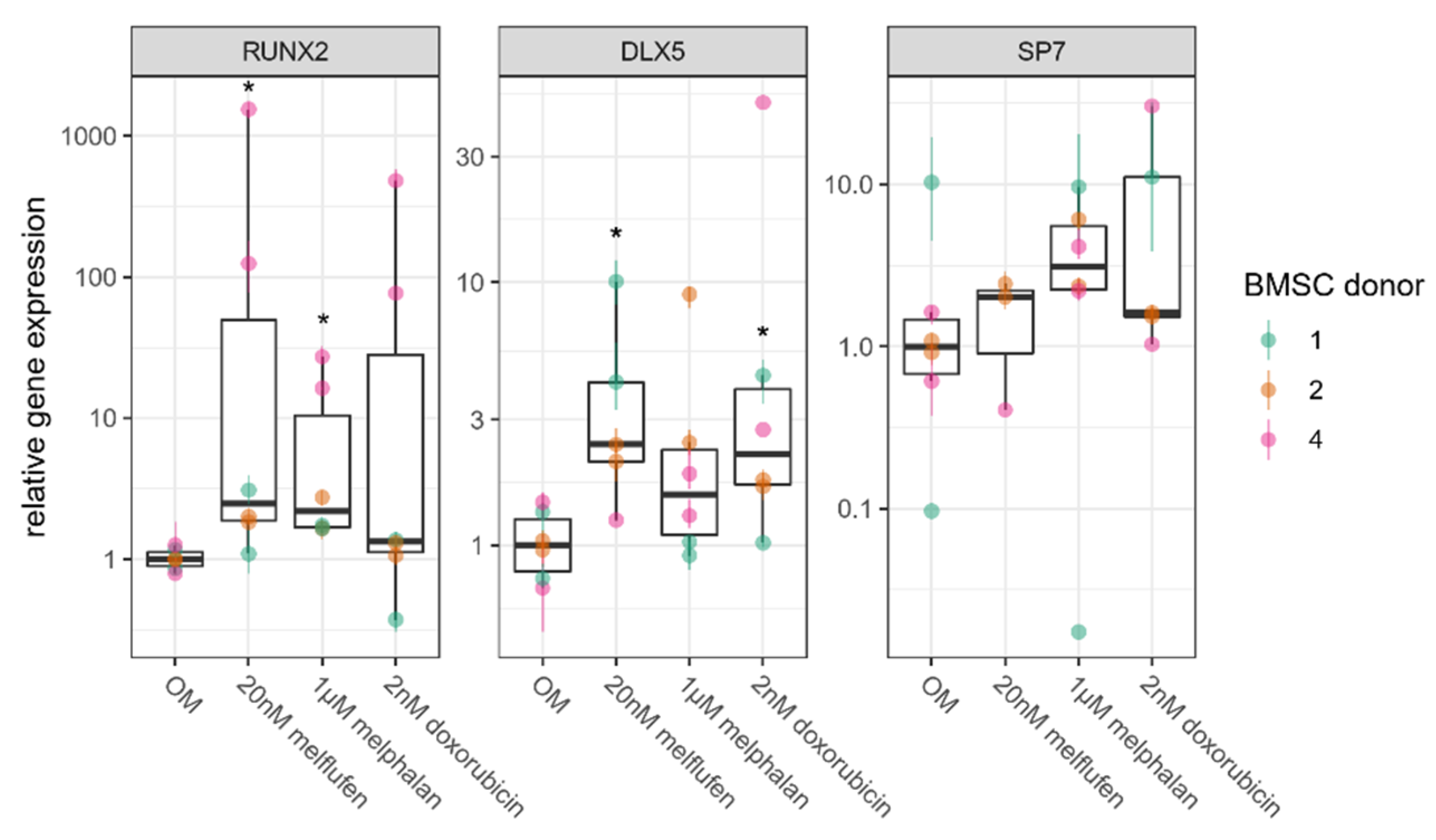
3.4. Osteogenic Differentiation
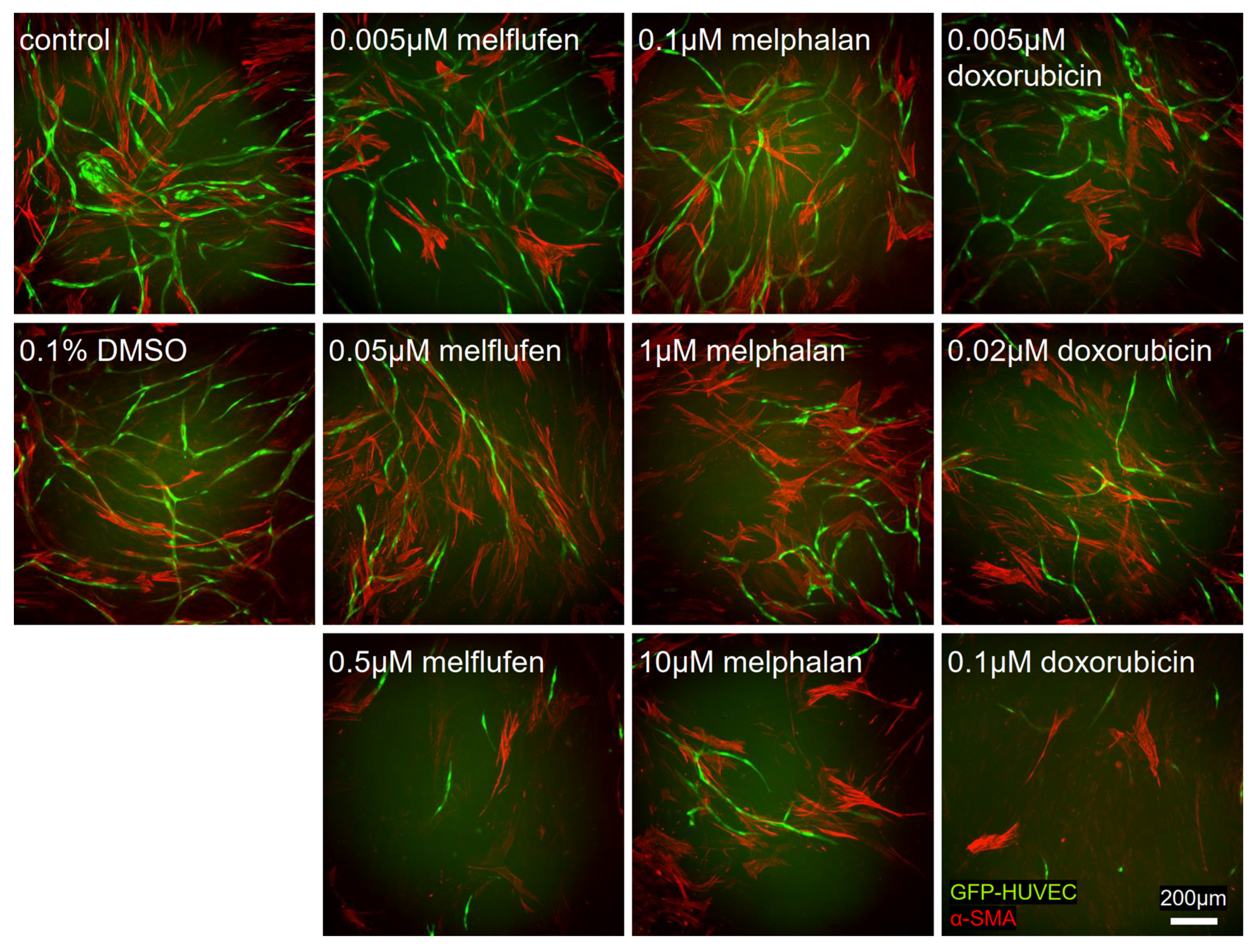
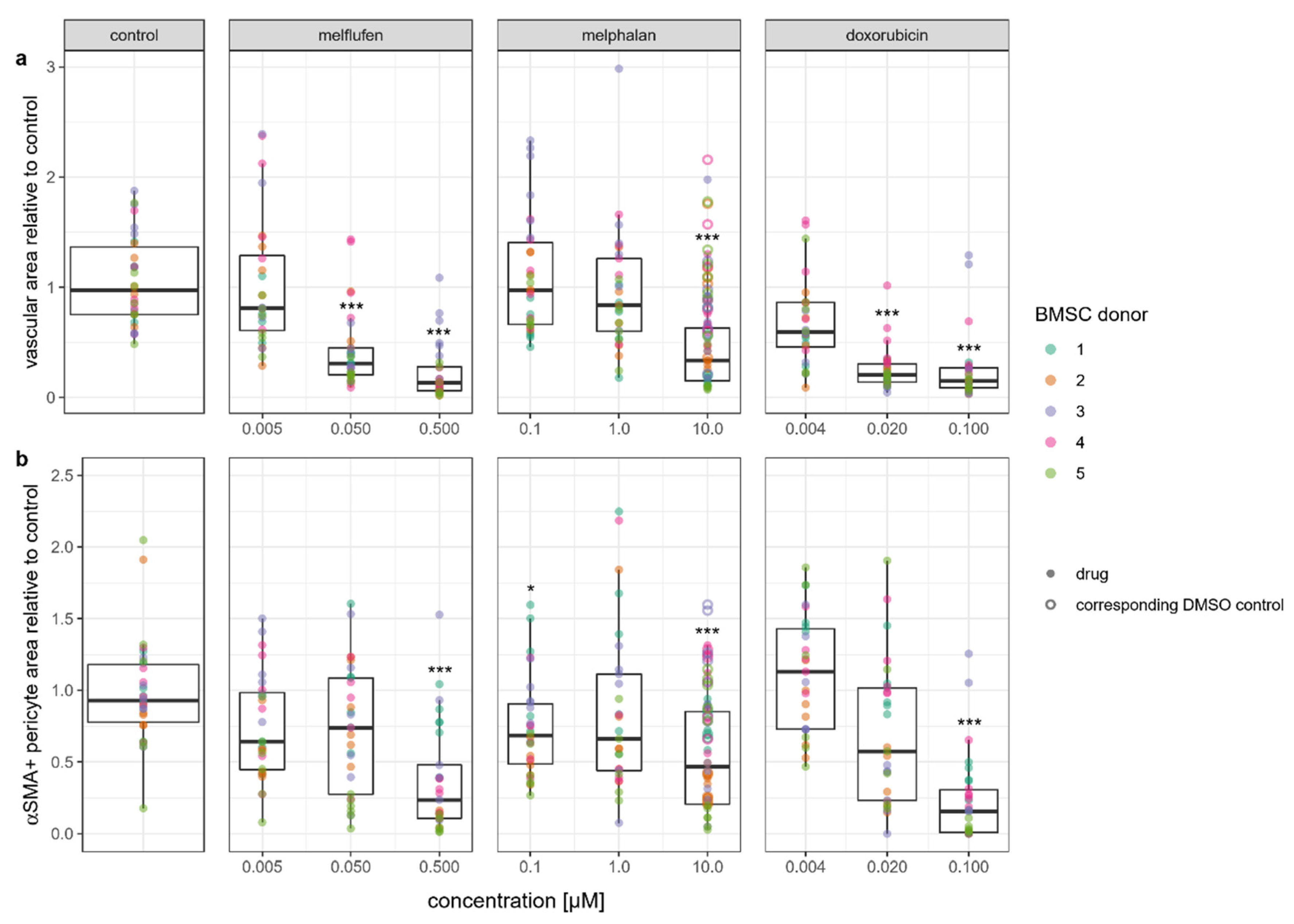
3.5. Angiogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, S.K.; Dimopoulos, M.A.; Kastritis, E.; Terpos, E.; Nahi, H.; Goldschmidt, H.; Hillengass, J.; Leleu, X.; Beksac, M.; Alsina, M.; et al. Natural history of relapsed myeloma, refractory to immunomodulatory drugs and proteasome inhibitors: A multicenter IMWG study. Leukemia 2017, 31, 2443–2448. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolock, S.L.; Krishnan, I.; Tenen, D.E.; Matkins, V.; Camacho, V.; Patel, S.; Agarwal, P.; Bhatia, R.; Tenen, D.G.; Klein, A.M.; et al. Mapping Distinct Bone Marrow Niche Populations and Their Differentiation Paths. Cell Rep. 2019, 28, 302–311.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, W.C.; Van Ness, B. The bone marrow stromal microenvironment influences myeloma therapeutic response in vitro. Leukemia 2001, 15, 264–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nefedova, Y.; Landowski, T.H.; Dalton, W.S. Bone marrow stromal-derived soluble factors and direct cell contact contribute to de novo drug resistance of myeloma cells by distinct mechanisms. Leukemia 2003, 17, 1175–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, Y.T.; Li, X.F.; Breitkreutz, I.; Song, W.; Neri, P.; Catley, L.; Podar, K.; Hideshima, T.; Chauhan, D.; Raje, N.; et al. Role of B-cell-activating factor in adhesion and growth of human multiple myeloma cells in the bone marrow microenvironment. Cancer Res. 2006, 66, 6675–6682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, M.; Foldes, G. It Takes Two: Endothelial-Perivascular Cell Cross-Talk in Vascular Development and Disease. Front. Cardiovasc. Med. 2018, 5, 154. [Google Scholar] [CrossRef] [PubMed]
- Reagan, M.R.; Liaw, L.; Rosen, C.J.; Ghobrial, I.M. Dynamic interplay between bone and multiple myeloma: Emerging roles of the osteoblast. Bone 2015, 75, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.B. Clinical pharmacology of melphalan and its implications for clinical resistance to anticancer agents. Cancer Treat. Res. 2002, 12, 305–322. [Google Scholar] [CrossRef]
- Gullbo, J.; Dhar, S.; Luthman, K.; Ehrsson, H.; Lewensohn, R.; Nygren, P.; Larsson, R. Antitumor activity of the alkylating oligopeptides J1 (L-melphalanyl-p-L-fluorophenylalanine ethyl ester) and P2 (L-prolyl-m-L-sarcolysyl-p-L-fluorophenylalanine ethyl ester): Comparison with melphalan. Anti-Cancer Drugs 2003, 14, 617–624. [Google Scholar] [CrossRef]
- Gullbo, J.; Lindhagen, E.; Bashir-Hassan, S.; Tullberg, M.; Ehrsson, H.; Lewensohn, R.; Nygren, P.; De La Torre, M.; Luthman, K.; Larsson, R. Antitumor efficacy and acute toxicity of the novel dipeptide melphalanyl-p-L-fluorophenylalanine ethyl ester (J1) in vivo. Investig. New Drugs 2004, 22, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Gullbo, J.; Tullberg, M.; Våbenø, J.; Ehrsson, H.; Lewensohn, R.; Nygren, P.; Larsson, R.; Luthman, K. Structure-activity relationship for alkylating dipeptide nitrogen mustard derivatives. Oncol. Res. 2003, 14, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Gullbo, J.; Wickström, M.; Tullberg, M.; Ehrsson, H.; Lewensohn, R.; Nygren, P.; Luthman, K.; Larsson, R. Activity of hydrolytic enzymes in tumour cells is a determinant for anti-tumour efficacy of the melphalan containing prodrug J1. J. Drug Target. 2003, 11, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Wickström, M.; Haglund, C.; Lindman, H.; Nygren, P.; Larsson, R.; Gullbo, J. The novel alkylating prodrug J1: Diagnosis directed activity profile ex vivo and combination analyses in vitro. Investig. New Drugs 2008, 26, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Wickström, M.; Johnsen, J.I.; Ponthan, F.; Segerström, L.; Sveinbjörnsson, B.; Lindskog, M.; Lövborg, H.; Viktorsson, K.; Lewensohn, R.; Kogner, P.; et al. The novel melphalan prodrug J1 inhibits neuroblastoma growth in vitro and in vivo. Mol. Cancer Ther. 2007, 6, 2409–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickström, M.; Viktorsson, K.; Lundholm, L.; Aesoy, R.; Nygren, H.; Sooman, L.; Fryknäs, M.; Vogel, L.K.; Lewensohn, R.; Larsson, R.; et al. The alkylating prodrug J1 can be activated by aminopeptidase N, leading to a possible target directed release of melphalan. Biochem Pharmacol. 2010, 79, 1281–1290. [Google Scholar] [CrossRef]
- Mateos, M.-V.; Bladé, J.; Bringhen, S.; Ocio, E.M.; Efebera, Y.; Pour, L.; Gay, F.; Sonneveld, P.; Gullbo, J.; Richardson, P.G. Melflufen: A Peptide–Drug Conjugate for the Treatment of Multiple Myeloma. J. Clin. Med. 2020, 9, 3120. [Google Scholar] [CrossRef]
- Richardson, P.G.; Oriol, A.; Larocca, A.; Bladé, J.; Cavo, M.; Rodriguez-Otero, P.; Leleu, X.; Nadeem, O.; Hiemenz, J.W.; Hassoun, H.; et al. HORIZON (OP-106) Investigators. Melflufen and Dexamethasone in Heavily Pretreated Relapsed and Refractory Multiple Myeloma. J. Clin. Oncol. 2021, 39, 757–767. [Google Scholar] [CrossRef]
- Li, Z.; Kupcsik, L.; Yao, S.J.; Alini, M.; Stoddart, M.J. Chondrogenesis of human bone marrow mesenchymal stem cells in fibrin-polyurethane composites. Tissue Eng. Part. A 2009, 15, 1729–1737. [Google Scholar] [CrossRef]
- Gebraad, A.; Kornilov, R.; Kaur, S.; Miettinen, S.; Haimi, S.; Peltoniemi, H.; Mannerström, B.; Seppänen-Kaijansinkko, R. Monocyte-derived extracellular vesicles stimulate cytokine secretion and gene expression of matrix metalloproteinases by mesenchymal stem/stromal cells. FEBS J. 2018, 285, 2337–2359. [Google Scholar] [CrossRef] [Green Version]
- Patrikoski, M.; Lee, M.H.C.; Mäkinen, L.; Ang, X.M.; Mannerström, B.; Raghunath, M.; Miettinen, S. Effects of Macromolecular Crowding on Human Adipose Stem Cell Culture in Fetal Bovine Serum, Human Serum, and Defined Xeno-Free/Serum-Free Conditions. Stem Cells Int. 2017, 2017, 6909163. [Google Scholar] [CrossRef] [PubMed]
- Patrikoski, M.; Juntunen, M.; Boucher, S.; Campbell, A.; Vemuri, M.C.; Mannerström, B.; Miettinen, S. Development of fully defined xeno-free culture system for the preparation and propagation of cell therapy-compliant human adipose stem cells. Stem Cell Res. Ther. 2013, 4, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojansivu, M.; Vanhatupa, S.; Björkvik, L.; Häkkänen, H.; Kellomäki, M.; Autio, R.; Ihalainen, J.A.; Hupa, L.; Miettinen, S. Bioactive glass ions as strong enhancers of osteogenic differentiation in human adipose stem cells. Acta Biomater. 2015, 21, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Zhang, L.; An, G.; Meng, H.; Han, Y.; Xie, Z.; Xu, Y.; Li, C.; Yu, Z.; Chang, H.; et al. Bone marrow stromal cells protect myeloma cells from bortezomib induced apoptosis by suppressing microRNA-15a expression. Leuk. Lymphoma. 2011, 52, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Lindroos, B.; Boucher, S.; Chase, L.; Kuokkanen, H.; Huhtala, H.; Haataja, R.; Vemuri, M.; Suuronen, R.; Miettinen, S. Serum-free, xeno-free culture media maintain the proliferation rate and multipotentiality of adipose stem cells in vitro. Cytotherapy 2009, 11, 958–972. [Google Scholar] [CrossRef]
- Kyllönen, L.; Haimi, S.; Mannerström, B.; Huhtala, H.; Rajala, K.M.; Skottman, H.; Sándor, G.K.; Miettinen, S. Effects of different serum conditions on osteogenic differentiation of human adipose stem cells in vitro. Stem Cell Res. Ther. 2013, 15, 17. [Google Scholar] [CrossRef] [Green Version]
- Juntunen, M.; Heinonen, S.; Huhtala, H.; Rissanen, A.; Kaprio, J.; Kuismanen, K.; Pietiläinen, K.H.; Miettinen, S.; Patrikoski, M. Evaluation of the effect of donor weight on adipose stromal/stem cell characteristics by using weight-discordant monozygotic twin pairs. Stem Cell Res. Ther. 2021, 12, 516. [Google Scholar] [CrossRef]
- Mykuliak, A.; Yrjänäinen, A.; Mäki, A.-J.; Gebraad, A.; Lampela, E.; Kääriäinen, M.; Pakarinen, T.-K.; Kallio, P.; Miettinen, S.; Vuorenpää, H. Vasculogenic Potency of Bone Marrow- and Adipose Tissue-Derived Mesenchymal Stem/Stromal Cells Results in Differing Vascular Network Phenotypes in a Microfluidic Chip. Front. Bioeng. Biotechnol. 2022, 10, 764237. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods. 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.H.; Alitalo, K. Molecular regulation of angiogenesis and lymphangiogenesis. Nat. Rev. Mol. Cell Biol. 2007, 8, 464–478. [Google Scholar] [CrossRef]
- Stratman, A.N.; Malotte, K.M.; Mahan, R.D.; Davis, M.J.; Davis, G.E. Pericyte recruitment during vasculogenic tube assembly stimulates endothelial basement membrane matrix formation. Blood 2009, 114, 5091–5101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haase, K.; Gillrie, M.R.; Hajal, C.; Kamm, R.D. Pericytes Contribute to Dysfunction in a Human 3D Model of Placental Microvasculature through VEGF-Ang-Tie2 Signaling. Adv. Sci. 2019, 6, 1900878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhatupa, S.; Ojansivu, M.; Autio, R.; Juntunen, M.; Miettinen, S. Bone Morphogenetic Protein-2 Induces Donor-Dependent Osteogenic and Adipogenic Differentiation in Human Adipose Stem Cells. Stem Cells Transl Med. 2015, 4, 1391–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-Response Analysis Using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorsten, P. PMCMRplus: Calculate Pairwise Multiple Comparisons of Mean Rank Sums Extended. Available online: https://CRAN.R-project.org/package=PMCMRplus (accessed on 24 March 2022).
- Xu, S.; De Veirman, K.; De Becker, A.; Vanderkerken, K.; Van Riet, I. Mesenchymal stem cells in multiple myeloma: A therapeutical tool or target? Leukemia 2018, 32, 1500–1514. [Google Scholar] [CrossRef]
- Lazarus, H.M.; Herzig, R.H.; Graham-Pole, J.; Wolff, S.N.; Phillips, G.L.; Strandjord, S.; Hurd, D.; Forman, W.; Gordon, E.M.; Coccia, P. Intensive melphalan chemotherapy and cryopreserved autologous bone marrow transplantation for the treatment of refractory cancer. J. Clin. Oncol. 1983, 1, 359–367. [Google Scholar] [CrossRef]
- Miettinen, J.J.; Kumari, R.; Traustadottir, G.A.; Huppunen, M.E.; Sergeev, P.; Majumder, M.M.; Schepsky, A.; Gudjonsson, T.; Lievonen, J.; Bazou, D.; et al. Aminopeptidase Expression in Multiple Myeloma Associates with Disease Progression and Sensitivity to Melflufen. Cancers 2021, 13, 1527. [Google Scholar] [CrossRef]
- Strese, S.; Wickström, M.; Fuchs, P.F.; Fryknäs, M.; Gerwins, P.; Dale, T.; Larsson, R.; Gullbo, J. The novel alkylating prodrug melflufen (J1) inhibits angiogenesis in vitro and in vivo. Biochem Pharmacol. 2013, 86, 888–895. [Google Scholar] [CrossRef]
- Moore, H.E.; Davenport, E.L.; Smith, E.M.; Muralikrishnan, S.; Dunlop, A.S.; Walker, B.A.; Krige, D.; Drummond, A.H.; Hooftman, L.; Morgan, G.J.; et al. Aminopeptidase inhibition as a targeted treatment strategy in myeloma. Mol. Cancer Ther. 2009, 8, 762–770. [Google Scholar] [CrossRef] [Green Version]
- Bellagamba, B.C.; de Abreu, B.R.R.; Grivicich, I.; Markarian, C.F.; Chem, E.; Camassola, M.; Nardi, N.B.; Dihl, R.R. Human mesenchymal stem cells are resistant to cytotoxic and genotoxic effects of cisplatin in vitro. Genet. Mol. Biol. 2016, 39, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Bosco, D.B.; Kenworthy, R.; Zorio, D.A.R.; Sang, Q.X.A. Human mesenchymal stem cells are resistant to paclitaxel by adopting a non-proliferative fibroblastic state. PLoS ONE. 2015, 10, e0128511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolay, N.H.; Perez, R.L.; Ruhle, A.; Trinh, T.; Sisombath, S.; Weber, K.J.; Ho, A.D.; Debus, J.; Saffrich, R.; Huber, P.E. Mesenchymal stem cells maintain their defining stem cell characteristics after treatment with cisplatin. Sci. Rep. 2016, 6, 20035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidmaier, R.; Baumann, P.; Emmerich, B.; Meinhardt, G. Evaluation of chemosensitivity of human bone marrow stromal cells--differences between common chemotherapeutic drugs. Anticancer Res. 2006, 26, 347–350. [Google Scholar] [PubMed]
- Oliveira, M.S.; Carvalho, J.L.; Campos, A.C.; Gomes, D.A.; de Goes, A.M.; Melo, M.M. Doxorubicin has in vivo toxicological effects on ex vivo cultured mesenchymal stem cells. Toxicol Lett. 2014, 224, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Cruet-Hennequart, S.; Prendergast, Á.M.; Shaw, G.; Barry, F.P.; Carty, M.P. Doxorubicin induces the DNA damage response in cultured human mesenchymal stem cells. Int. J. Hematol. 2012, 96, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Kemp, K.; Morse, R.; Sanders, K.; Hows, J.; Donaldson, C. Alkylating chemotherapeutic agents cyclophosphamide and melphalan cause functional injury to human bone marrow-derived mesenchymal stem cells. Ann. Hematol. 2011, 90, 777–789. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.A.; Keutmann, M.K.; Dowling, M.L.; Harris, E.; Chan, G.; Kao, G.D. Inactivation of the mitotic checkpoint as a determinant of the efficacy of microtubule-targeted drugs in killing human cancer cells. Mol. Cancer Ther. 2004, 3, 661–669. [Google Scholar]
- Hardin, J.; MacLeod, S.; Grigorieva, I.; Chang, R.; Barlogie, B.; Xiao, H.; Epstein, J. Interleukin-6 prevents dexamethasone-induced myeloma cell death. Blood 1994, 84, 3063–3070. [Google Scholar] [CrossRef] [Green Version]
- Somaiah, C.; Kumar, A.; Sharma, R.; Sharma, A.; Anand, T.; Bhattacharyya, J.; Das, D.; Deka Talukdar, S.; Jaganathan, B.G. Mesenchymal stem cells show functional defect and decreased anti-cancer effect after exposure to chemotherapeutic drugs. J. Biomed. Sci. 2018, 25, 5. [Google Scholar] [CrossRef] [Green Version]
- Nicolay, N.H.; Rühle, A.; Perez, R.L.; Trinh, T.; Sisombath, S.; Weber, K.J.; Ho, A.D.; Debus, J.; Saffrich, R.; Huber, P.E. Mesenchymal stem cells are sensitive to bleomycin treatment. Sci. Rep. 2016, 6, 26645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; De Veirman, K.; Evans, H.; Santini, G.C.; Vande Broek, I.; Leleu, X.; De Becker, A.; Van Camp, B.; Croucher, P.; Vanderkerken, K.; et al. Effect of the HDAC inhibitor vorinostat on the osteogenic differentiation of mesenchymal stem cells in vitro and bone formation in vivo. Acta Pharm. Sin. 2013, 34, 699–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Folkman, J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell 1996, 86, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Yoo, S.Y.; Kwon, S.M. Angiogenesis and its therapeutic opportunities. Mediators Inflamm. 2013, 2013, 127170. [Google Scholar] [CrossRef] [Green Version]
- Winter, U.; Mena, H.A.; Negrotto, S.; Arana, E.; Pascual-Pasto, G.; Laurent, V.; Suñol, M.; Chantada, G.L.; Carcaboso, A.M.; Schaiquevich, P. Schedule-Dependent Antiangiogenic and Cytotoxic Effects of Chemotherapy on Vascular Endothelial and Retinoblastoma Cells. PLoS ONE 2016, 11, e0160094. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xing, Y.; Liang, C.; Hu, L.; Xu, F.; Mei, Q. In search of underlying mechanisms and potential drugs of melphalan-induced vascular toxicity through retinal endothelial cells using bioinformatics approach. Tumour Biol. 2016, 37, 6709–6718. [Google Scholar] [CrossRef]
- Maragoudakis, M.E.; Peristeris, P.; Missirlis, E.; Aletras, A.; Andriopoulou, P.; Haralabopoulos, G. Inhibition of angiogenesis by anthracyclines and titanocene dichloride. Ann. N. Y. Acad Sci. 1994, 732, 280–293. [Google Scholar] [CrossRef]
- Uvez, A.; Aydinlik, S.; Esener, O.; Erkisa, M.; Karakus, D.; Armutak, E.I. Synergistic interactions between resveratrol and doxorubicin inhibit angiogenesis both in vitro and in vivo. Pol. J. Vet. Sci. 2020, 23, 571–580. [Google Scholar] [CrossRef]
- Gurel-Gurevin, E.; Kiyan, H.T.; Esener, O.; Aydinlik, S.; Uvez, A.; Ulukaya, E.; Dimas, K.; Armutak, E.I. Chloroquine Used in Combination with Chemotherapy Synergistically Suppresses Growth and Angiogenesis In Vitro and In Vivo. Anticancer Res. 2018, 38, 4011–4020. [Google Scholar] [CrossRef]
- Sato, M.; Arap, W.; Pasqualini, R. Molecular targets on blood vessels for cancer therapies in clinical trials. Oncology 2007, 21, 1346–1352. [Google Scholar]
- Fukasawa, K.; Fujii, H.; Saitoh, Y.; Koizumi, K.; Aozuka, Y.; Sekine, K.; Yamada, M.; Saiki, I.; Nishikawa, K. Aminopeptidase N (APN/CD13) is selectively expressed in vascular endothelial cells and plays multiple roles in angiogenesis. Cancer Lett. 2006, 243, 135–143. [Google Scholar] [CrossRef] [PubMed]















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebraad, A.; Ohlsbom, R.; Miettinen, J.J.; Emeh, P.; Pakarinen, T.-K.; Manninen, M.; Eskelinen, A.; Kuismanen, K.; Slipicevic, A.; Lehmann, F.; et al. Growth Response and Differentiation of Bone Marrow-Derived Mesenchymal Stem/Stromal Cells in the Presence of Novel Multiple Myeloma Drug Melflufen. Cells 2022, 11, 1574. https://doi.org/10.3390/cells11091574
Gebraad A, Ohlsbom R, Miettinen JJ, Emeh P, Pakarinen T-K, Manninen M, Eskelinen A, Kuismanen K, Slipicevic A, Lehmann F, et al. Growth Response and Differentiation of Bone Marrow-Derived Mesenchymal Stem/Stromal Cells in the Presence of Novel Multiple Myeloma Drug Melflufen. Cells. 2022; 11(9):1574. https://doi.org/10.3390/cells11091574
Chicago/Turabian StyleGebraad, Arjen, Roope Ohlsbom, Juho J. Miettinen, Promise Emeh, Toni-Karri Pakarinen, Mikko Manninen, Antti Eskelinen, Kirsi Kuismanen, Ana Slipicevic, Fredrik Lehmann, and et al. 2022. "Growth Response and Differentiation of Bone Marrow-Derived Mesenchymal Stem/Stromal Cells in the Presence of Novel Multiple Myeloma Drug Melflufen" Cells 11, no. 9: 1574. https://doi.org/10.3390/cells11091574
APA StyleGebraad, A., Ohlsbom, R., Miettinen, J. J., Emeh, P., Pakarinen, T. -K., Manninen, M., Eskelinen, A., Kuismanen, K., Slipicevic, A., Lehmann, F., Nupponen, N. N., Heckman, C. A., & Miettinen, S. (2022). Growth Response and Differentiation of Bone Marrow-Derived Mesenchymal Stem/Stromal Cells in the Presence of Novel Multiple Myeloma Drug Melflufen. Cells, 11(9), 1574. https://doi.org/10.3390/cells11091574