
The Role of Non-Coding RNAs in the Human Placenta


Abstract
:1. Introduction
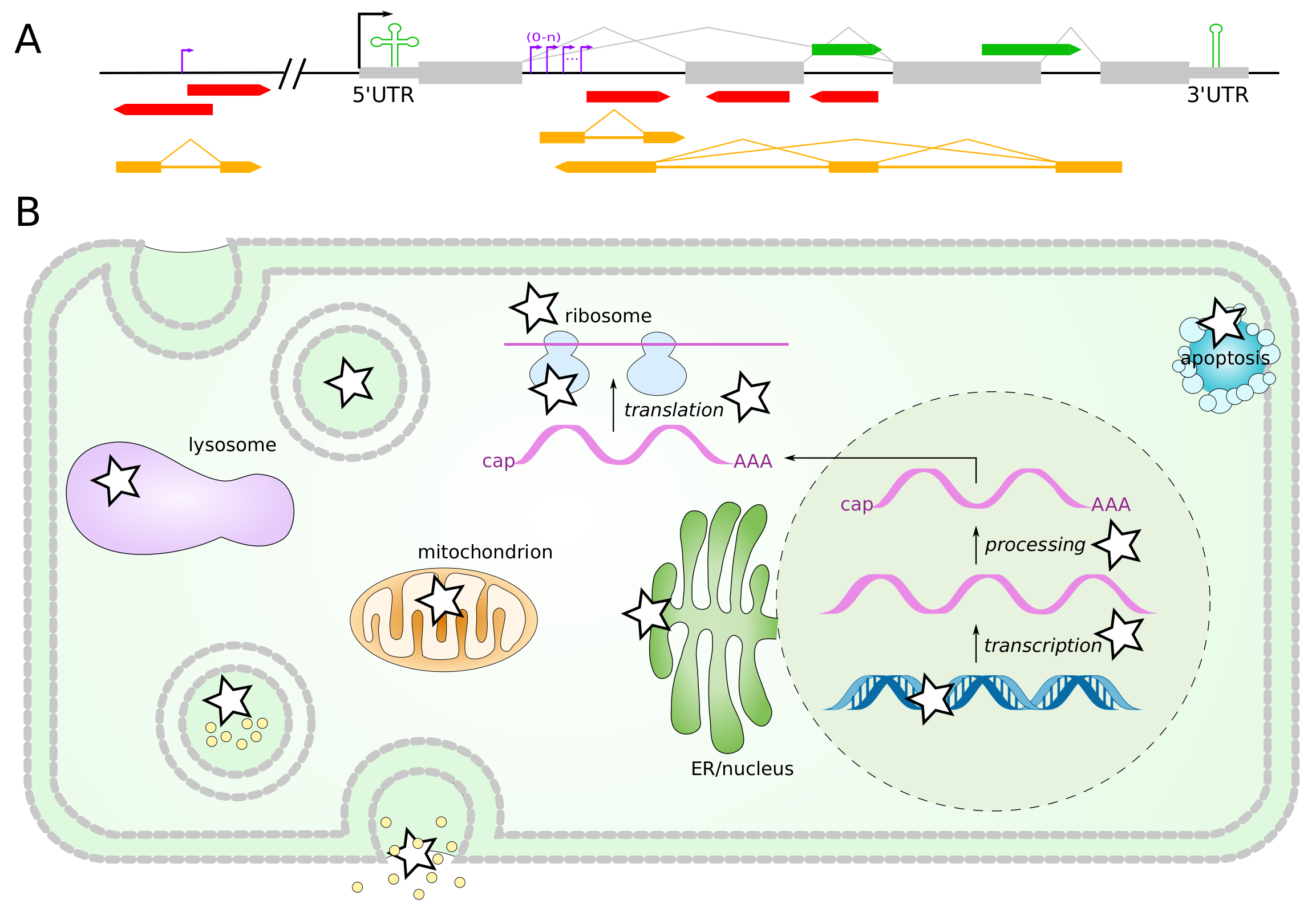
2. Prediction and Identification of ncRNAs
2.1. Identification of miRNAs as Marker Genes
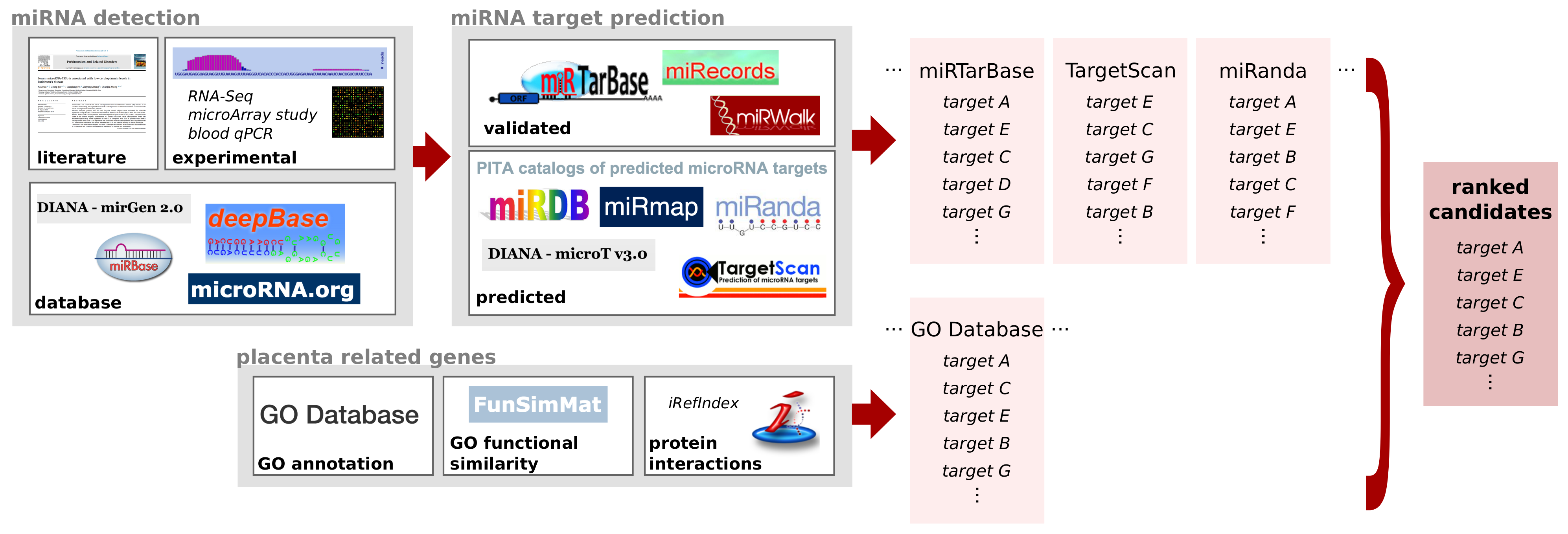
2.2. Non-Coding RNA Target Prediction
3. MicroRNAs and MicroRNA Clusters in Placenta
3.1. miRNA Clusters C14MC, C19MC and miR-371-3 in Trophoblast Cells
3.2. Placental MicroRNAs in Pregnancy Pathologies
3.2.1. miRNA and Preeclampsia
3.2.2. Gestational Diabetes Mellitus
3.2.3. Miscarriage
3.2.4. Other Pathologies
4. Long Non-Coding RNAs in Placenta
Long Non-CODING RNAs in Pathologies
5. Circular RNAs in Placenta
6. Circulating Non-Coding RNAs in Maternal Serum/Plasma
Free RNAs and Pathologies
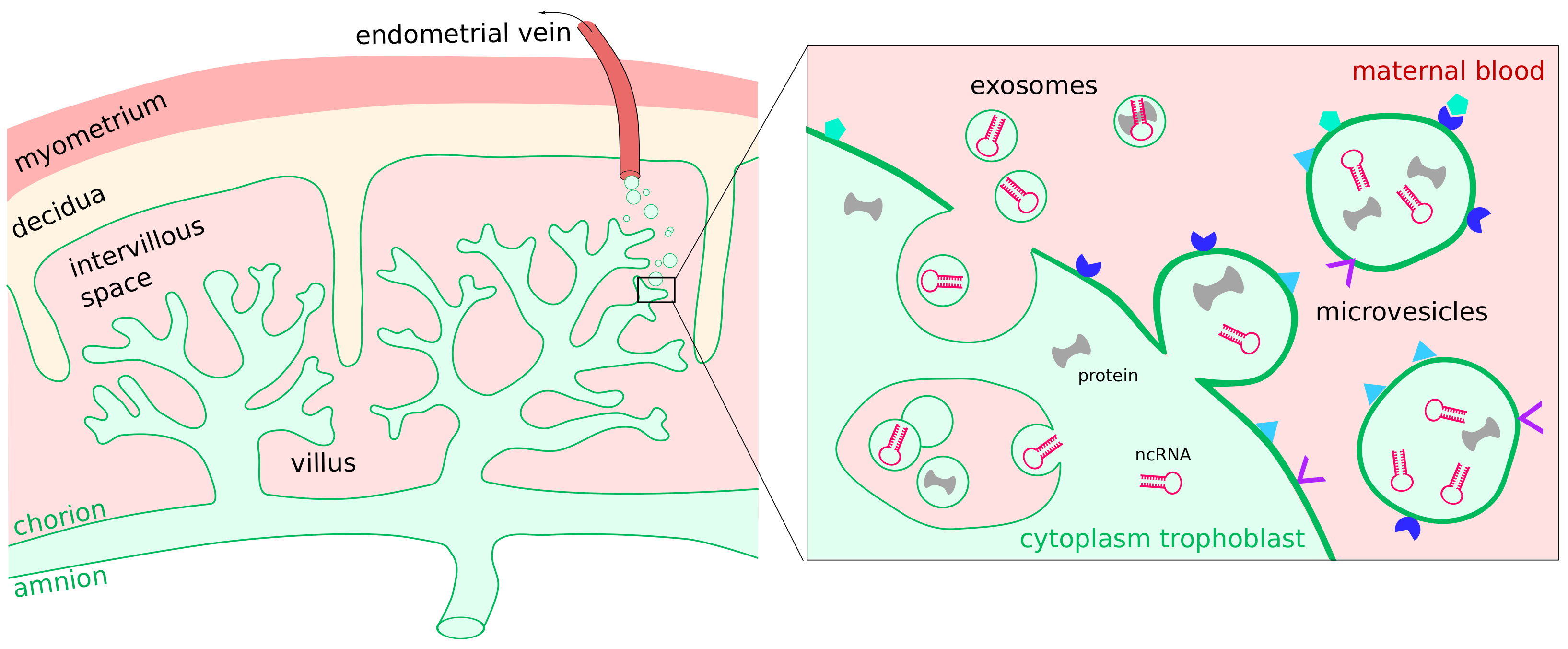
7. Extracellular Vesicles
EVs in Pregnancy Pathologies
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schmidt, A.; Morales-Prieto, D.M.; Pastuschek, J.; Froehlich, K.; Markert, U.R. Only humans have human placentas: Molecular differences between mice and humans. J. Reprod. Immunol. 2015, 108, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Göhner, C.; Weber, M.; Tannetta, D.S.; Groten, T.; Plösch, T.; Faas, M.M.; Scherjon, S.A.; Schleußner, E.; Markert, U.R.; Fitzgerald, J.S. A New Enzyme-linked Sorbent Assay (ELSA) to Quantify Syncytiotrophoblast Extracellular Vesicles in Biological Fluids. Am. J. Reprod. Immunol. 2015, 73, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Morales-Prieto, D.M.; Ospina-Prieto, S.; Schmidt, A.; Chaiwangyen, W.; Markert, U.R. Elsevier Trophoblast Research Award Lecture: Origin, evolution and future of placenta miRNAs. Placenta 2014, 35, S39–S45. [Google Scholar] [CrossRef] [PubMed]
- ENCODE Project Consortium; Birney, E.; Stamatoyannopoulos, J.A.; Dutta, A.; Guigó, R.; Gingeras, T.R.; Margulies, E.H.; Weng, Z.; Snyder, M.; Dermitzakis, E.T.; et al. Identification and analysis of functional elements in 1% of the human genome by the ENCODE pilot project. Nature 2007, 447, 799–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ENCODE Project Consortium. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Gregory, T.R. Synergy between sequence and size in large-scale genomics. Nat. Rev. Genet. 2005, 6, 699–708. [Google Scholar] [CrossRef]
- Kung, J.T.Y.; Colognori, D.; Lee, J.T. Long Noncoding RNAs: Past, Present, and Future. Genetics 2013, 193, 651–669. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.C.; Mendell, J.T. microRNAs in vertebrate physiology and human disease. Annu. Rev. Genom. Hum. Genet. 2007, 8, 215–239. [Google Scholar] [CrossRef] [Green Version]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef]
- Cox, B.; Leavey, K.; Nosi, U.; Wong, F.; Kingdom, J. Placental transcriptome in development and pathology: Expression, function, and methods of analysis. Am. J. Obstet. Gynecol. 2015, 213, S138–S151. [Google Scholar] [CrossRef]
- Kalvari, I.; Nawrocki, E.P.; Ontiveros-Palacios, N.; Argasinska, J.; Lamkiewicz, K.; Marz, M.; Griffiths-Jones, S.; Toffano-Nioche, C.; Gautheret, D.; Weinberg, Z.; et al. Rfam 14: Expanded coverage of metagenomic, viral and microRNA families. Nucleic Acids Res. 2021, 49, D192–D200. [Google Scholar] [CrossRef] [PubMed]
- Harrow, J.; Frankish, A.; Gonzalez, J.M.; Tapanari, E.; Diekhans, M.; Kokocinski, F.; Aken, B.L.; Barrell, D.; Zadissa, A.; Searle, S.; et al. GENCODE: The reference human genome annotation for The ENCODE Project. Genome Res. 2012, 22, 1760–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales Prieto, D.M.; Markert, U.R. MicroRNAs in pregnancy. J. Reprod. Immunol. 2011, 88, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Morales-Prieto, D.M.; Ospina-Prieto, S.; Chaiwangyen, W.; Schoenleben, M.; Markert, U.R. Pregnancy-associated miRNA-clusters. J. Reprod. Immunol. 2013, 97, 51–61. [Google Scholar] [CrossRef] [PubMed]
- He, X.; He, Y.; Xi, B.; Zheng, J.; Zeng, X.; Cai, Q.; OuYang, Y.; Wang, C.; Zhou, X.; Huang, H.; et al. LncRNAs expression in preeclampsia placenta reveals the potential role of lncRNAs contributing to preeclampsia pathogenesis. PLoS ONE 2013, 8, e81437. [Google Scholar]
- Zou, Y.; Jiang, Z.; Yu, X.; Sun, M.; Zhang, Y.; Zuo, Q.; Zhou, J.; Yang, N.; Han, P.; Ge, Z.; et al. Upregulation of long noncoding RNA SPRY4-IT1 modulates proliferation, migration, apoptosis, and network formation in trophoblast cells HTR-8SV/neo. PLoS ONE 2013, 8, e79598. [Google Scholar] [CrossRef] [Green Version]
- Wang, E.C.; Wang, A.Z. Nanoparticles and their applications in cell and molecular biology. Integr. Biol. 2014, 6, 9–26. [Google Scholar] [CrossRef] [Green Version]
- Ospina-Prieto, S.; Chaiwangyen, W.; Herrmann, J.; Groten, T.; Schleussner, E.; Markert, U.R.; Morales-Prieto, D.M. MicroRNA-141 is upregulated in preeclamptic placentae and regulates trophoblast invasion and intercellular communication. Transl. Res. 2016, 172, 61–72. [Google Scholar] [CrossRef]
- Moro, L.; Bardají, A.; Macete, E.; Barrios, D.; Morales-Prieto, D.M.; España, C.; Mandomando, I.; Sigaúque, B.; Dobaño, C.; Markert, U.R.; et al. Placental Microparticles and microRNAs in Pregnant Women with Plasmodium falciparum or HIV Infection. PLoS ONE 2016, 11, e0146361. [Google Scholar] [CrossRef]
- Gezer, U.; Özgür, E.; Cetinkaya, M.; Isin, M.; Dalay, N. Long non-coding RNAs with low expression levels in cells are enriched in secreted exosomes. Cell Biol. Int. 2014, 38, 1076–1079. [Google Scholar] [CrossRef]
- Takahashi, K.; Yan, I.K.; Wood, J.; Haga, H.; Patel, T. Involvement of extracellular vesicle long noncoding RNA (linc-VLDLR) in tumor cell responses to chemotherapy. Mol. Cancer Res. 2014, 12, 1377–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, A.R.; Findeiß, S.; Washietl, S.; Hofacker, I.L.; Stadler, P.F. RNAz 2.0: Improved noncoding RNA detection. In Biocomputing 2010; World Scientific: Singapore, 2010; pp. 69–79. [Google Scholar]
- Rivas, E.; Eddy, S.R. Noncoding RNA gene detection using comparative sequence analysis. BMC Bioinform. 2001, 2, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camargo, A.P.; Sourkov, V.; Pereira, G.A.G.; Carazzolle, M.F. RNAsamba: Neural network-based assessment of the protein-coding potential of RNA sequences. NAR Genom. Bioinform. 2020, 2, lqz024. [Google Scholar] [CrossRef] [Green Version]
- Wucher, V.; Legeai, F.; Hedan, B.; Rizk, G.; Lagoutte, L.; Leeb, T.; Jagannathan, V.; Cadieu, E.; David, A.; Lohi, H.; et al. FEELnc: A tool for long non-coding RNA annotation and its application to the dog transcriptome. Nucleic Acids Res. 2017, 45, e57. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Yin, H.; Li, B.; Yu, C.; Wang, F.; Xu, X.; Cao, J.; Bao, Y.; Wang, L.; Abbasi, A.A.; et al. Characterization and identification of long non-coding RNAs based on feature relationship. Bioinformatics 2019, 35, 2949–2956. [Google Scholar] [CrossRef]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef]
- Hu, L.; Xu, Z.; Hu, B.; Lu, Z.J. COME: A robust coding potential calculation tool for lncRNA identification and characterization based on multiple features. Nucleic Acids Res. 2017, 45, e2. [Google Scholar] [CrossRef]
- Li, A.; Zhang, J.; Zhou, Z. PLEK: A tool for predicting long non-coding RNAs and messenger RNAs based on an improved k-mer scheme. BMC Bioinform. 2014, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.F.; Jungreis, I.; Kellis, M. PhyloCSF: A comparative genomics method to distinguish protein coding and non-coding regions. Bioinformatics 2011, 27, i275–i282. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, H.; Zhang, L.; Meng, J. lncRScan-SVM: A tool for predicting long non-coding RNAs using support vector machine. PLoS ONE 2015, 10, e0139654. [Google Scholar] [CrossRef]
- Chen, J.; Shishkin, A.A.; Zhu, X.; Kadri, S.; Maza, I.; Guttman, M.; Hanna, J.H.; Regev, A.; Garber, M. Evolutionary analysis across mammals reveals distinct classes of long non-coding RNAs. Genome Biol. 2016, 17, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, C.M.; Weretilnyk, E.A.; Golding, G.B. Prediction of plant lncRNA by ensemble machine learning classifiers. BMC Genom. 2018, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.; Zhang, Q.; Deng, M.; Miao, J.; Guo, Y.; Gao, W.; Cui, Q. An analysis of human microRNA and disease associations. PLoS ONE 2008, 3, e3420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Shi, J.; Gao, Y.; Cui, C.; Zhang, S.; Li, J.; Zhou, Y.; Cui, Q. HMDD v3. 0: A database for experimentally supported human microRNA–disease associations. Nucleic Acids Res. 2019, 47, D1013–D1017. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.F. Non-coding RNAs: New players in eukaryotic biology. Gene 2005, 357, 83–94. [Google Scholar] [CrossRef]
- Chatzou, M.; Magis, C.; Chang, J.M.; Kemena, C.; Bussotti, G.; Erb, I.; Notredame, C. Multiple sequence alignment modeling: Methods and applications. Brief. Bioinform. 2016, 17, 1009–1023. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.; Feinbaum, R.; Ambros, V. The C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Hosseini, M.K.; Gunel, T.; Gumusoglu, E.; Benian, A.; Aydinli, K. MicroRNA expression profiling in placenta and maternal plasma in early pregnancy loss. Mol. Med. Rep. 2018, 17, 4941–4952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, M.; Kolluru, G.K.; Ahmed, A. Small Molecule, Big Prospects: MicroRNA in Pregnancy and Its Complications. J. Pregnancy 2017, 2017, 6972732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayder, H.; O’Brien, J.; Nadeem, U.; Peng, C. MicroRNAs: Crucial regulators of placental development. Reproduction 2018, 155, R259–R271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Ma, Y.; Wu, H.; Wang, Y.L. Placenta-Derived MicroRNAs in the Pathophysiology of Human Pregnancy. Front. Cell Dev. Biol. 2021, 9, 540. [Google Scholar] [CrossRef]
- Liu, W.; Niu, Z.; Li, Q.; Pang, R.T.K.; Chiu, P.C.N.; Yeung, W.S.B. MicroRNA and Embryo Implantation. Am. J. Reprod. Immunol. 2016, 75, 263–271. [Google Scholar] [CrossRef]
- Liang, Y.; Ridzon, D.; Wong, L.; Chen, C. Characterization of microRNA expression profiles in normal human tissues. BMC Genom. 2007, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.S.; Ishibashi, O.; Ishikawa, G.; Ishikawa, T.; Katayama, A.; Mishima, T.; Takizawa, T.; Shigihara, T.; Goto, T.; Izumi, A.; et al. Human villous trophoblasts express and secrete placenta-specific microRNAs into maternal circulation via exosomes. Biol. Reprod. 2009, 81, 717–729. [Google Scholar] [CrossRef] [Green Version]
- Donker, R.; Mouillet, J.; Chu, T.; Hubel, C.; Stolz, D.; Morelli, A.; Sadovsky, Y. The expression profile of C19MC microRNAs in primary human trophoblast cells and exosomes. Mol. Hum. Reprod. 2012, 18, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Morales-Prieto, D.M.; Chaiwangyen, W.; Ospina-Prieto, S.; Schneider, U.; Herrmann, J.; Gruhn, B.; Markert, U.R. MicroRNA expression profiles of trophoblastic cells. Placenta 2012, 33, 725–734. [Google Scholar] [CrossRef]
- Légaré, C.; Clément, A.A.; Desgagné, V.; Thibeault, K.; White, F.; Guay, S.P.; Arsenault, B.J.; Scott, M.S.; Jacques, P.É.; Perron, P.; et al. Human plasma pregnancy-associated miRNAs and their temporal variation within the first trimester of pregnancy. Reprod. Biol. Endocrinol. 2022, 20, 1–13. [Google Scholar] [CrossRef]
- Morales-Prieto, D.M.; Favaro, R.R.; Markert, U.R. Placental miRNAs in feto-maternal communication mediated by extracellular vesicles. Placenta 2020, 102, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Mouillet, J.F.; Goff, J.; Sadovsky, E.; Sun, H.; Parks, T.; Chu, T.; Sadovsky, Y. Transgenic expression of human C19MC miRNAs impacts placental morphogenesis. Placenta 2020, 101, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Paquette, A.G.; Chu, T.; Wu, X.; Wang, K.; Price, N.D.; Sadovsky, Y. Distinct communication patterns of trophoblastic miRNA among the maternal-placental-fetal compartments. Placenta 2018, 72, 28–35. [Google Scholar] [CrossRef]
- Benetatos, L.; Hatzimichael, E.; Londin, E.; Vartholomatos, G.; Loher, P.; Rigoutsos, I.; Briasoulis, E. The microRNAs within the DLK1-DIO3 genomic region: Involvement in disease pathogenesis. Cell. Mol. Life Sci. 2013, 70, 795–814. [Google Scholar] [CrossRef]
- Hu, H.Y.; He, L.; Fominykh, K.; Yan, Z.; Guo, S.; Zhang, X.; Taylor, M.S.; Tang, L.; Li, J.; Liu, J.; et al. Evolution of the human-specific microRNA miR-941. Nat. Commun. 2012, 3, 1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazov, E.A.; McWilliam, S.; Barris, W.C.; Dalrymple, B.P. Origin, evolution, and biological role of miRNA cluster in DLK-DIO3 genomic region in placental mammals. Mol. Biol. Evol. 2008, 25, 939–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, M.; Weise, A.; Vasheghani, F.; Göhner, C.; Fitzgerald, J.S.; Liehr, T.; Markert, U.R. Cytogenomics of six human trophoblastic cell lines. Placenta 2021, 103, 72–75. [Google Scholar] [CrossRef]
- Pastuschek, J.; Nonn, O.; Gutiérrez-Samudio, R.N.; Murrieta-Coxca, J.M.; Müller, J.; Sanft, J.; Huppertz, B.; Markert, U.R.; Groten, T.; Morales-Prieto, D.M. Molecular characteristics of established trophoblast-derived cell lines. Placenta 2021, 108, 122–133. [Google Scholar] [CrossRef]
- Noguer-Dance, M.; Abu-Amero, S.; Al-Khtib, M.; Lefèvre, A.; Coullin, P.; Moore, G.E.; Cavaillé, J. The primate-specific microRNA gene cluster (C19MC) is imprinted in the placenta. Hum. Mol. Genet. 2010, 19, 3566–3582. [Google Scholar] [CrossRef] [Green Version]
- Malnou, E.C.; Umlauf, D.; Mouysset, M.; Cavaillé, J. Imprinted microRNA gene clusters in the evolution, development, and functions of mammalian placenta. Front. Genet. 2019, 9, 706. [Google Scholar] [CrossRef]
- Flor, I.; Neumann, A.; Freter, C.; Helmke, B.M.; Langenbuch, M.; Rippe, V.; Bullerdiek, J. Abundant expression and hemimethylation of C19MC in cell cultures from placenta-derived stromal cells. Biochem. Biophys. Res. Commun. 2012, 422, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Mong, E.F.; Yang, Y.; Akat, K.M.; Canfield, J.; VanWye, J.; Lockhart, J.; Tsibris, J.C.; Schatz, F.; Lockwood, C.J.; Tuschl, T.; et al. Chromosome 19 microRNA cluster enhances cell reprogramming by inhibiting epithelial-to-mesenchymal transition. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumont, T.M.F.; Mouillet, J.F.; Bayer, A.; Gardner, C.L.; Klimstra, W.B.; Wolf, D.G.; Yagel, S.; Balmir, F.; Binstock, A.; Sanfilippo, J.S.; et al. The expression level of C19MC miRNAs in early pregnancy and in response to viral infection. Placenta 2017, 53, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hromadnikova, I.; Kotlabova, K.; Ivankova, K.; Krofta, L. Expression profile of C19MC microRNAs in placental tissue of patients with preterm prelabor rupture of membranes and spontaneous preterm birth. Mol. Med. Rep. 2017, 16, 3849–3862. [Google Scholar] [CrossRef]
- Bayer, A.; Lennemann, N.J.; Ouyang, Y.; Sadovsky, E.; Sheridan, M.A.; Roberts, R.M.; Coyne, C.B.; Sadovsky, Y. Chromosome 19 microRNAs exert antiviral activity independent from type III interferon signaling. Placenta 2018, 61, 33–38. [Google Scholar] [CrossRef]
- Flor, I.; Bullerdiek, J. The dark side of a success story: MicroRNAs of the C19MC cluster in human tumours. J. Pathol. 2012, 227, 270–274. [Google Scholar] [CrossRef]
- Wu, S.; Aksoy, M.; Shi, J.; Houbaviy, H.B. Evolution of the miR-290–295/miR-371–373 cluster family seed repertoire. PLoS ONE 2014, 9, e108519. [Google Scholar] [CrossRef]
- Shah, J.A.; Khattak, S.; Rauf, M.A.; Cai, Y.; Jin, J. Potential Biomarkers of miR-371–373 Gene Cluster in Tumorigenesis. Life 2021, 11, 984. [Google Scholar] [CrossRef]
- Escudero, C.A.; Herlitz, K.; Troncoso, F.; Acurio, J.; Aguayo, C.; Roberts, J.M.; Truong, G.; Duncombe, G.; Rice, G.; Salomon, C. Role of extracellular vesicles and microRNAs on dysfunctional angiogenesis during preeclamptic pregnancies. Front. Physiol. 2016, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Cai, Y. Down-regulation of microRNA let-7d inhibits the proliferation and invasion of trophoblast cells in preeclampsia. J. Cell Biochem. 2018, 119, 1141–1151. [Google Scholar] [CrossRef]
- Lu, T.M.; Lu, W.; Zhao, L.J. MicroRNA-137 Affects Proliferation and Migration of Placenta Trophoblast Cells in Preeclampsia by Targeting ERRα. Reprod. Sci. 2017, 24, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yan, J. MicroRNA-454 is involved in regulating trophoblast cell proliferation, apoptosis, and invasion in preeclampsia by modulating the expression of ephrin receptor B4. Biomed. Pharmacother. 2018, 107, 746–753. [Google Scholar] [CrossRef]
- Wu, L.; Song, W.Y.; Xie, Y.; Hu, L.L.; Hou, X.M.; Wang, R.; Gao, Y.; Zhang, J.N.; Zhang, L.; Li, W.W.; et al. miR-181a-5p suppresses invasion and migration of HTR-8/SVneo cells by directly targeting IGF2BP2. Cell Death Dis. 2018, 9, 16. [Google Scholar] [CrossRef]
- Gao, Y.; She, R.; Wang, Q.; Li, Y.; Zhang, H. Up-regulation of miR-299 suppressed the invasion and migration of HTR-8/SVneo trophoblast cells partly via targeting HDAC2 in pre-eclampsia. Biomed. Pharmacother. 2018, 97, 1222–1228. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Niu, H.; Cheng, L.; Li, X.; Zhang, Q.; Ning, Y. Invasion of trophoblast cell lines is inhibited by miR-93 via MMP-2. Placenta 2017, 53, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Long, A.; Tan, L.; Hong, M.; Wu, J.; Cai, L.; Li, Q. Elevated microRNA-520g in pre-eclampsia inhibits migration and invasion of trophoblasts. Placenta 2017, 51, 70–75. [Google Scholar] [CrossRef]
- Niu, Z.R.; Han, T.; Sun, X.L.; Luan, L.X.; Gou, W.L.; Zhu, X.M. MicroRNA-30a-3p is overexpressed in the placentas of patients with preeclampsia and affects trophoblast invasion and apoptosis by its effects on IGF-1. Am. J. Obstet. Gynecol. 2018, 218, 249.e1–249.e12. [Google Scholar] [CrossRef]
- Zou, A.X.; Chen, B.; Li, Q.X.; Liang, Y.C. MiR-134 inhibits infiltration of trophoblast cells in placenta of patients with preeclampsia by decreasing ITGB1 expression. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2199–2206. [Google Scholar] [CrossRef]
- Gao, T.; Deng, M.; Wang, Q. miRNA-320a inhibits trophoblast cell invasion by targeting estrogen-related receptor-gamma. J. Obstet. Gynaecol. Res. 2018, 44, 756–763. [Google Scholar] [CrossRef]
- Fang, M.; Du, H.; Han, B.; Xia, G.; Shi, X.; Zhang, F.; Fu, Q.; Zhang, T. Hypoxia-inducible microRNA-218 inhibits trophoblast invasion by targeting LASP1: Implications for preeclampsia development. Int. J. Biochem. Cell Biol. 2017, 87, 95–103. [Google Scholar] [CrossRef]
- Wu, H.; Wang, H.; Liu, M.; Bai, Y.; Li, Y.X.; Ji, L.; Peng, C.; Yu, Y.; Wang, Y.L. MiR-195 participates in the placental disorder of preeclampsia via targeting activin receptor type-2B in trophoblastic cells. J. Hypertens. 2016, 34, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Chen, Y.; Chen, L.; Wang, K.; Pan, T.; Liu, X.; Xu, W. miR-15b-AGO2 play a critical role in HTR8/SVneo invasion and in a model of angiogenesis defects related to inflammation. Placenta 2016, 41, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, P.; Mao, X.; Zhang, H. Regulatory mechanism of miR-525-5p in over-invasion of trophoblast. J. Obstet. Gynaecol. Res. 2021, 47, 679–688. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Yang, L.; Wang, H. miR-520 promotes DNA-damage-induced trophoblast cell apoptosis by targeting PARP1 in recurrent spontaneous abortion (RSA). Gynecol. Endocrinol. 2017, 33, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.C.; Chen, M.; Wang, Y.X.; Rui, C.; Xu, W.; Ding, H.J.; Shi, Z.H. MicroRNA-128a-induced apoptosis in HTR-8/SVneo trophoblast cells contributes to pre-eclampsia. Biomed. Pharmacother. 2016, 81, 63–70. [Google Scholar] [CrossRef]
- Guo, M.; Zhao, X.; Yuan, X.; Li, P. Elevated microRNA-34a contributes to trophoblast cell apoptosis in preeclampsia by targeting BCL-2. J. Hum. Hypertens. 2017, 31, 815–820. [Google Scholar] [CrossRef]
- Zhang, Y.; Pan, X.; Yu, X.; Li, L.; Qu, H.; Li, S. MicroRNA-590-3p inhibits trophoblast-dependent maternal spiral artery remodeling by repressing low-density lipoprotein receptor-related protein 6. Mol. Genet. Genom. Med. 2018, 6, 1124–1133. [Google Scholar] [CrossRef] [Green Version]
- Brkić, J.; Dunk, C.; O’Brien, J.; Fu, G.; Nadeem, L.; Wang, Y.l.; Rosman, D.; Salem, M.; Shynlova, O.; Yougbaré, I.; et al. MicroRNA-218-5p promotes endovascular trophoblast differentiation and spiral artery remodeling. Mol. Ther. 2018, 26, 2189–2205. [Google Scholar] [CrossRef] [Green Version]
- Hayder, H.; Fu, G.; Nadeem, L.; O’Brien, J.A.; Lye, S.J.; Peng, C. Overexpression of miR-210-3p Impairs Extravillous Trophoblast Functions Associated with Uterine Spiral Artery Remodeling. Int. J. Mol. Sci. 2021, 22, 3961. [Google Scholar] [CrossRef]
- Bounds, K.R.; Chiasson, V.L.; Pan, L.J.; Gupta, S.; Chatterjee, P. MicroRNAs: New Players in the Pathobiology of Preeclampsia. Front. Cardiovasc. Med. 2017, 4, 60. [Google Scholar] [CrossRef] [Green Version]
- Hemmatzadeh, M.; Shomali, N.; Yousefzadeh, Y.; Mohammadi, H.; Ghasemzadeh, A.; Yousefi, M. MicroRNAs: Small molecules with a large impact on pre-eclampsia. J. Cell. Physiol. 2020, 235, 3235–3248. [Google Scholar] [CrossRef] [PubMed]
- Jairajpuri, D.S.; Almawi, W.Y. MicroRNA expression pattern in pre-eclampsia (Review). Mol. Med. Rep. 2016, 13, 2351–2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Y.; Lu, C.; Ji, X.; Miao, Z.; Long, W.; Ding, H.; Lv, M. Roles of microRNAs in preeclampsia. J. Cell. Physiol. 2019, 234, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Skalis, G.; Katsi, V.; Miliou, A.; Georgiopoulos, G.; Papazachou, O.; Vamvakou, G.; Nihoyannopoulos, P.; Tousoulis, D.; Makris, T. MicroRNAs in preeclampsia. MicroRNA 2019, 8, 28–35. [Google Scholar] [CrossRef]
- Lagana, A.S.; Vitale, S.G.; Sapia, F.; Valenti, G.; Corrado, F.; Padula, F.; Rapisarda, A.M.C.; D’Anna, R. miRNA expression for early diagnosis of preeclampsia onset: Hope or hype? J. Matern. Fetal Neonatal Med. 2018, 31, 817–821. [Google Scholar] [CrossRef]
- Hornakova, A.; Kolkova, Z.; Holubekova, V.; Loderer, D.; Lasabova, Z.; Biringer, K.; Halasova, E. Diagnostic Potential of MicroRNAs as Biomarkers in the Detection of Preeclampsia. Genet. Test. Mol. Biomark. 2020, 24, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Feng, L.; Zhang, H.; Hachy, S.; Satohisa, S.; Laurent, L.C.; Parast, M.; Zheng, J.; Chen, D.b. Preeclampsia up-regulates angiogenesis-associated microRNA (ie., miR-17,-20a, and-20b) that target ephrin-B2 and EPHB4 in human placenta. J. Clin. Endocrinol. Metab. 2012, 97, E1051–E1059. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Kan, X.; Liu, P.; Yin, L.; Li, Q.; Xu, H. MiR-20b is implicated in preeclampsia progression via the regulation of myeloid cell leukemin-1. J. Biol. Regul. Homeost. Agents 2020, 34, 1709–1717. [Google Scholar] [PubMed]
- Shao, X.; Liu, Y.; Liu, M.; Wang, Y.; Yan, L.; Wang, H.; Ma, L.; Li, Y.X.; Zhao, Y.; Wang, Y.L. Testosterone Represses Estrogen Signaling by Upregulating miR-22: A Mechanism for Imbalanced Steroid Hormone Production in Preeclampsia. Hypertension 2017, 69, 721–730. [Google Scholar] [CrossRef]
- Rezaei, M.; Eskandari, F.; Mohammadpour-Gharehbagh, A.; Harati-Sadegh, M.; Teimoori, B.; Salimi, S. Hypomethylation of the miRNA-34a gene promoter is associated with Severe Preeclampsia. Clin. Exp. Hypertens. 2018, 41, 118–122. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, L.; Zhang, F.; Luan, T.; Yin, Z.; Rui, C.; Ding, H. Influence of miR-34a on preeclampsia through the Notch signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 923–931. [Google Scholar] [PubMed]
- Xue, F.; Yang, J.; Li, Q.; Zhou, H. Down-regulation of microRNA-34a-5p promotes trophoblast cell migration and invasion via targetting Smad4. Biosci. Rep. 2019, 39, BSR20181631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Wu, L.; Zhang, G.; Tang, R.; Zhou, X. Elevated MicroRNA-181a-5p contributes to trophoblast dysfunction and preeclampsia. Reprod. Sci. 2019, 26, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Shi, L.; Hong, L.; Chen, X.; Cen, H. MiR-135a-5p promotes the migration and invasion of trophoblast cells in preeclampsia by targeting β-TrCP. Placenta 2020, 99, 63–69. [Google Scholar] [CrossRef]
- Wu, D.; Chen, X.; Wang, L.; Chen, F.; Cen, H.; Shi, L. Hypoxia-induced microRNA-141 regulates trophoblast apoptosis, invasion, and vascularization by blocking CXCL12β/CXCR2/4 signal transduction. Biomed. Pharmacother. 2019, 116, 108836. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, K.; Zhou, W.; Liu, H.; Yang, T.; Hou, P.; Li, X. miR-141-5p regulate ATF2 via effecting MAPK1/ERK2 signaling to promote preeclampsia. Biomed. Pharmacother. 2019, 115, 108953. [Google Scholar] [CrossRef]
- Xiao, J.; Tao, T.; Yin, Y.; Zhao, L.; Yang, L.; Hu, L. miR-144 may regulate the proliferation, migration and invasion of trophoblastic cells through targeting PTEN in preeclampsia. Biomed. Pharmacother. 2017, 94, 341–353. [Google Scholar] [CrossRef]
- Hu, S.; Li, J.; Tong, M.; Li, Q.; Chen, Y.; Lu, H.; Wang, Y.; Min, L. MicroRNA-144-3p may participate in the pathogenesis of preeclampsia by targeting Cox-2. Mol. Med. Rep. 2019, 19, 4655–4662. [Google Scholar] [CrossRef] [Green Version]
- Gunel, T.; Kamali, N.; Hosseini, M.K.; Gumusoglu, E.; Benian, A.; Aydinli, K. Regulatory effect of miR-195 in the placental dysfunction of preeclampsia. J. Matern. Fetal Neonatal Med. 2020, 33, 901–908. [Google Scholar] [CrossRef]
- Hu, T.X.; Wang, G.; Guo, X.J.; Sun, Q.Q.; He, P.; Gu, H.; Huang, Y.; Gao, L.; Ni, X. MiR 20a,-20b and -200c are involved in hydrogen sulfide stimulation of VEGF production in human placental trophoblasts. Placenta 2016, 39, 101–110. [Google Scholar] [CrossRef]
- Liu, F.; Wu, K.; Wu, W.; Chen, Y.; Wu, H.; Wang, H.; Zhang, W. miR-203 contributes to pre-eclampsia via inhibition of VEGFA expression. Mol. Med. Rep. 2018, 17, 5627–5634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, N.; Jia, Z.; Li, L. miR-320a upregulation contributes to the development of preeclampsia by inhibiting the growth and invasion of trophoblast cells by targeting interleukin 4. Mol. Med. Rep. 2019, 20, 3256–3264. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; She, K.; Li, H.; Yuan, X.; Han, X.; Wang, Y. MicroRNA-454 contributes to sustaining the proliferation and invasion of trophoblast cells through inhibiting Nodal/ALK7 signaling in pre-eclampsia. Chem. Biol. Interact. 2019, 298, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhao, X.; Shan, H.; Gao, H.; Wang, P. microRNA-520c-3p suppresses NLRP3 inflammasome activation and inflammatory cascade in preeclampsia by downregulating NLRP3. Inflamm. Res. 2019, 68, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Li, H.; Wei, J.X. MiR-4421 regulates the progression of preeclampsia by regulating CYP11B2. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1533–1540. [Google Scholar] [CrossRef]
- Licini, C.; Avellini, C.; Picchiassi, E.; Mensà, E.; Fantone, S.; Ramini, D.; Tersigni, C.; Tossetta, G.; Castellucci, C.; Tarquini, F.; et al. Pre-eclampsia predictive ability of maternal miR-125b: A clinical and experimental study. Transl. Res. 2021, 228, 13–27. [Google Scholar] [CrossRef]
- Poirier, C.; Desgagné, V.; Guérin, R.; Bouchard, L. MicroRNAs in Pregnancy and Gestational Diabetes Mellitus: Emerging Role in Maternal Metabolic Regulation. Curr. Diab. Rep. 2017, 17, 35. [Google Scholar] [CrossRef]
- Iljas, J.D.; Guanzon, D.; Elfeky, O.; Rice, G.E.; Salomon, C. Review: Bio-compartmentalization of microRNAs in exosomes during gestational diabetes mellitus. Placenta 2017, 54, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Muralimanoharan, S.; Maloyan, A.; Myatt, L. Mitochondrial function and glucose metabolism in the placenta with gestational diabetes mellitus: Role of miR-143. Clin. Sci. 2016, 130, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.L.; Zhang, L.; Li, J.; Tian, S.; Lv, X.D.; Wang, X.Q.; Su, X.; Li, Y.; Hu, Y.; Ma, X.; et al. Up-regulation of miR-98 and unraveling regulatory mechanisms in gestational diabetes mellitus. Sci. Rep. 2016, 6, 32268. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.Y.; Li, M.Q.; Li, H.P. miR-137 restricts the viability and migration of HTR-8/SVneo cells by downregulating FNDC5 in gestational diabetes mellitus. Curr. Mol. Med. 2019, 19, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.G.; Tian, S.; Zhang, L.; Hu, Y.; Guan, C.Y.; Ma, X.; Xia, H.F. The miRNA-29b is downregulated in placenta during gestational diabetes mellitus and may alter placenta development by regulating trophoblast migration and invasion through a HIF3A-dependent mechanism. Front. Endocrinol. 2020, 11, 169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, K.; Tian, S.; Wang, X.Q.; Li, J.H.; Dong, Y.C.; Xia, H.F.; Ma, X. Down-regulation of microRNA-30d-5p is associated with gestational diabetes mellitus by targeting RAB8A. J. Diabetes Its Complicat. 2021, 35, 107959. [Google Scholar] [CrossRef]
- Wen, J.; Bai, X. miR-520h Inhibits cell survival by targeting mTOR in gestational diabetes mellitus. Acta Biochim. Pol. 2021, 68, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liu, Z.; Fang, J.; Qi, H. miR-96-5p: A potential diagnostic marker for gestational diabetes mellitus. Medicine 2021, 100, e25808. [Google Scholar] [CrossRef]
- Zhou, X.; Xiang, C.; Zheng, X. miR-132 serves as a diagnostic biomarker in gestational diabetes mellitus and its regulatory effect on trophoblast cell viability. Diagn. Pathol. 2019, 14, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Shen, W.W.; Cao, X.M.; Ding, W.Y.; Yan, L.P.; Gao, L.J.; Li, X.L.; Zhong, T.Y. Novel mechanism of miRNA-365-regulated trophoblast apoptosis in recurrent miscarriage. J. Cell. Mol. Med. 2017, 21, 2412–2425. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.N.; Tang, X.M.; Wang, X.Q.; Gao, J.; Li, N.; Wang, Y.Y.; Xia, H.F. MiR-93 inhibits trophoblast cell proliferation and promotes cell apoptosis by targeting BCL2L2 in recurrent spontaneous abortion. Reprod. Sci. 2020, 27, 152–162. [Google Scholar] [CrossRef]
- Ding, J.; Cheng, Y.; Zhang, Y.; Liao, S.; Yin, T.; Yang, J. The miR-27a-3p/USP25 axis participates in the pathogenesis of recurrent miscarriage by inhibiting trophoblast migration and invasion. J. Cell. Physiol. 2019, 234, 19951–19963. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Pan, L.; Xiong, Y.; Wang, L.; Cui, Y.; Liu, J.; Tang, L. Down-regulation of the Sp1 transcription factor by an increase of microRNA-4497 in human placenta is associated with early recurrent miscarriage. Reprod. Biol. Endocrinol. 2021, 19, 1–12. [Google Scholar] [CrossRef]
- Du, E.; Cao, Y.; Feng, C.; Lu, J.; Yang, H.; Zhang, Y. The Possible Involvement of miR-37la-5p Regulating XIAP in the Pathogenesis of Recurrent Pregnancy Loss. Reprod. Sci. 2019, 26, 1468–1475. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Yu, T.; Xu, H.; Hou, N.; Cheng, Q.; Lai, L.; Wang, Q.; Sheng, J.; Huang, H. Down-regulation of miR-378a-3p induces decidual cell apoptosis: A possible mechanism for early pregnancy loss. Hum. Reprod. 2018, 33, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.X.; Li, L.; Dong, Y.J.; Li, P.H.; Su, Q.; Guo, Y.H.; Lu, Y.R.; Zhong, Y.; Jia, Y.; Cheng, J.Q. miR-146a-5p improves the decidual cytokine microenvironment by regulating the toll-like receptor signaling pathway in unexplained spontaneous abortion. Int. Immunopharmacol. 2020, 89, 107066. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, J.; Huang, S. Patients with unexplained recurrent spontaneous abortion show decreased levels of microrna-146a-5p in the deciduae. Ann. Clin. Lab. Sci. 2018, 48, 177–182. [Google Scholar]
- Romero-Ruiz, A.; Avendaño, M.S.; Dominguez, F.; Lozoya, T.; Molina-Abril, H.; Sangiao-Alvarellos, S.; Gurrea, M.; Lara-Chica, M.; Fernandez-Sanchez, M.; Torres-Jimenez, E.; et al. Deregulation of miR-324/KISS1/kisspeptin in early ectopic pregnancy: Mechanistic findings with clinical and diagnostic implications. Am. J. Obstet. Gynecol. 2019, 220, 480.e1–480.e17. [Google Scholar] [CrossRef] [Green Version]
- Svobodová, I.; Korabečná, M.; Calda, P.; Břešťák, M.; Pazourková, E.; Pospíšilová, V.; Krkavcová, M.; Novotná, M.; Hořínek, A. Differentially expressed miRNAs in trisomy 21 placentas. Prenat. Diagn. 2016, 36, 775–784. [Google Scholar] [CrossRef]
- Rahman, M.L.; Liang, L.; Valeri, L.; Su, L.; Zhu, Z.; Gao, S.; Mostofa, G.; Qamruzzaman, Q.; Hauser, R.; Baccarelli, A.; et al. Regulation of birthweight by placenta-derived miRNAs: Evidence from an arsenic-exposed birth cohort in Bangladesh. Epigenetics 2018, 13, 573–590. [Google Scholar] [CrossRef]
- Zhang, J.T.; Cai, Q.Y.; Ji, S.S.; Zhang, H.X.; Wang, Y.H.; Yan, H.T.; Yang, X.J. Decreased miR-143 and increased miR-21 placental expression levels are associated with macrosomia. Mol. Med. Rep. 2016, 13, 3273–3280. [Google Scholar] [CrossRef]
- Kennedy, E.M.; Hermetz, K.; Burt, A.; Everson, T.M.; Deyssenroth, M.; Hao, K.; Chen, J.; Karagas, M.R.; Pei, D.; Koestler, D.C.; et al. Placental microRNA expression associates with birthweight through control of adipokines: Results from two independent cohorts. Epigenetics 2021, 16, 770–782. [Google Scholar] [CrossRef]
- Tagliaferri, S.; Cepparulo, P.; Vinciguerra, A.; Campanile, M.; Esposito, G.; Maruotti, G.M.; Zullo, F.; Annunziato, L.; Pignataro, G. miR-16-5p, miR-103-3p, and miR-27b-3p as Early Peripheral Biomarkers of Fetal Growth Restriction. Front. Pediatr. 2021, 9, 156. [Google Scholar] [CrossRef]
- Carreras-Badosa, G.; Bonmatí, A.; Ortega, F.J.; Mercader, J.M.; Guindo-Martínez, M.; Torrents, D.; Prats-Puig, A.; Martinez-Calcerrada, J.M.; de Zegher, F.; Ibáñez, L.; et al. Dysregulation of Placental miRNA in Maternal Obesity Is Associated with Pre- and Postnatal Growth. J. Clin. Endocrinol. Metab. 2017, 102, 2584–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.; Ni, X.; Mao, X.; Ying, H.; Du, Q. Cholestatic pregnancy is associated with reduced VCAM1 expression in vascular endothelial cell of placenta. Reprod. Toxicol. 2017, 74, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Bian, Y.; Xu, X.; Wang, X.; Zuo, C.; Meng, J.; Li, H.; Zhao, S.; Ning, Y.; Cao, Y.; et al. Downregulation of miR-29a/b/c in placenta accreta inhibits apoptosis of implantation site intermediate trophoblast cells by targeting MCL1. Placenta 2016, 48, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Meng, J.; Zuo, C.; Wang, S.; Li, H.; Zhao, S.; Huang, T.; Wang, X.; Yan, J. Downregulation of MicroRNA-125a in Placenta Accreta Spectrum Disorders Contributes Antiapoptosis of Implantation Site Intermediate Trophoblasts by Targeting MCLI. Reprod. Sci. 2019, 26, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Li, N.; Hou, R.; Qiao, C.; Liu, C. Development and validation of a four-microRNA signature for placenta accreta spectrum: An integrated competing endogenous RNA network analysis. Ann. Transl. Med. 2020, 8, 919. [Google Scholar] [CrossRef]
- Chen, S.; Pang, D.; Li, Y.; Zhou, J.; Liu, Y.; Yang, S.; Liang, K.; Yu, B. Serum miRNA biomarker discovery for placenta accreta spectrum. Placenta 2020, 101, 215–220. [Google Scholar] [CrossRef]
- Gysler, S.M.; Mulla, M.J.; Guerra, M.; Brosens, J.J.; Salmon, J.E.; Chamley, L.W.; Abrahams, V.M. Antiphospholipid antibody-induced miR-146a-3p drives trophoblast interleukin-8 secretion through activation of Toll-like receptor 8. Mol. Hum. Reprod. 2016, 22, 465–474. [Google Scholar] [CrossRef] [Green Version]
- McAninch, D.; Roberts, C.T.; Bianco-Miotto, T. Mechanistic Insight into Long Noncoding RNAs and the Placenta. Int. J. Mol. Sci. 2017, 18, 1371. [Google Scholar] [CrossRef] [Green Version]
- Majewska, M.; Lipka, A.; Paukszto, L.; Jastrzebski, J.P.; Gowkielewicz, M.; Jozwik, M.; Majewski, M.K. Preliminary [RNA]-Seq Analysis of Long Non-Coding [RNA]s Expressed in Human Term Placenta. Int. J. Mol. Sci. 2018, 19, 1894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chishima, T.; Iwakiri, J.; Hamada, M. Identification of Transposable Elements Contributing to Tissue-Specific Expression of Long Non-Coding RNAs. Genes 2018, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Nordin, M.; Bergman, D.; Halje, M.; Engström, W.; Ward, A. Epigenetic regulation of the Igf2/H19 gene cluster. Cell Prolif. 2014, 47, 189–199. [Google Scholar] [CrossRef]
- Yu, L.; Chen, M.; Zhao, D.; Yi, P.; Lu, L.; Han, J.; Zheng, X.; Zhou, Y.; Li, L. The H19 gene imprinting in normal pregnancy and pre-eclampsia. Placenta 2009, 30, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Gabory, A.; Ripoche, M.A.; Yoshimizu, T.; Dandolo, L. The H19 gene: Regulation and function of a non-coding RNA. Cytogenet. Genome Res. 2006, 113, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Gabory, A.; Jammes, H.; Dandolo, L. The H19 locus: Role of an imprinted non-coding RNA in growth and development. Bioessays 2010, 32, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol. 2012, 14, 659–665. [Google Scholar] [CrossRef]
- Tsunoda, Y.; Kudo, M.; Wada, R.; Ishino, K.; Kure, S.; Sakatani, T.; Takeshita, T.; Naito, Z. Expression level of long noncoding RNA H19 of normotensive placentas in late pregnancy relates to the fetal growth restriction. J. Obstet. Gynaecol. Res. 2020, 46, 1025–1034. [Google Scholar] [CrossRef]
- Lu, L.; Hou, Z.; Li, L.; Yang, Y.; Wang, X.; Zhang, B.; Ren, M.; Zhao, D.; Miao, Z.; Yu, L.; et al. Methylation pattern of H19 exon 1 is closely related to preeclampsia and trophoblast abnormalities. Int. J. Mol. Med. 2014, 34, 765–771. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Xia, Y.; Zhang, H.; Guo, H.; Feng, K.; Zhang, C. Overexpression of long non-coding RNA H19 promotes invasion and autophagy via the PI3K/AKT/mTOR pathways in trophoblast cells. Biomed. Pharmacother. 2018, 101, 691–697. [Google Scholar] [CrossRef]
- Fauque, P.; Ripoche, M.A.; Tost, J.; Journot, L.; Gabory, A.; Busato, F.; Le Digarcher, A.; Mondon, F.; Gut, I.; Jouannet, P.; et al. Modulation of imprinted gene network in placenta results in normal development of in vitro manipulated mouse embryos. Hum. Mol. Genet. 2010, 19, 1779–1790. [Google Scholar] [CrossRef] [Green Version]
- Sakian, S.; Louie, K.; Wong, E.C.; Havelock, J.; Kashyap, S.; Rowe, T.; Taylor, B.; Ma, S. Altered gene expression of H19 and IGF2 in placentas from ART pregnancies. Placenta 2015, 36, 1100–1105. [Google Scholar] [CrossRef]
- Marjonen, H.; Toivonen, M.; Lahti, L.; Kaminen-Ahola, N. Early prenatal alcohol exposure alters imprinted gene expression in placenta and embryo in a mouse model. PLoS ONE 2018, 13, e0197461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freschi, A.; Hur, S.K.; Valente, F.M.; Ideraabdullah, F.Y.; Sparago, A.; Gentile, M.T.; Oneglia, A.; Di Nucci, D.; Colucci-D’Amato, L.; Thorvaldsen, J.L.; et al. Tissue-specific and mosaic imprinting defects underlie opposite congenital growth disorders in mice. PLoS Genet. 2018, 14, e1007243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuckerwise, L.; Li, J.; Lu, L.; Men, Y.; Geng, T.; Buhimschi, C.S.; Buhimschi, I.A.; Bukowski, R.; Guller, S.; Paidas, M.; et al. H19 long noncoding RNA alters trophoblast cell migration and invasion by regulating TβR3 in placentae with fetal growth restriction. Oncotarget 2016, 7, 38398–38407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piyamongkol, W.; Suprasert, P. Allelic Characterization of IGF2 and H19 Gene Polymorphisms in Molar Tissues. Asian Pac. J. Cancer Prev. 2016, 17, 4405–4408. [Google Scholar] [PubMed]
- Su, R.; Wang, C.; Feng, H.; Lin, L.; Liu, X.; Wei, Y.; Yang, H. Alteration in Expression and Methylation of IGF2/H19 in Placenta and Umbilical Cord Blood Are Associated with Macrosomia Exposed to Intrauterine Hyperglycemia. PLoS ONE 2016, 11, e0148399. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Luo, X.; Gao, Q.; Wang, Y.; Gao, Q.; Long, W. Dysregulation of LncRNAs in Placenta and Pathogenesis of Preeclampsia. Curr. Drug Targets 2017, 18, 1165–1170. [Google Scholar] [CrossRef]
- Moradi, M.T.; Rahimi, Z.; Vaisi-Raygani, A. New insight into the role of long non-coding RNAs in the pathogenesis of preeclampsia. Hypertens. Pregnancy 2019, 38, 41–51. [Google Scholar] [CrossRef]
- Yang, X.; Meng, T. Long noncoding RNA in preeclampsia: Transcriptional noise or innovative indicators? BioMed Res. Int. 2019, 2019, 5437621. [Google Scholar] [CrossRef]
- Li, T.; Hu, D.; Gong, Y. Identification of potential lncRNAs and co-expressed mRNAs in gestational diabetes mellitus by RNA sequencing. J. Matern. Fetal Neonatal Med. 2021, 1–15. [Google Scholar] [CrossRef]
- Wu, K.; Liu, F.; Wu, W.; Chen, Y.; Wu, H.; Zhang, W. Long non-coding RNA HOX transcript antisense RNA (HOTAIR) suppresses the angiogenesis of human placentation by inhibiting vascular endothelial growth factor A expression. Reprod. Fertil. Dev. 2019, 31, 377–385. [Google Scholar] [CrossRef]
- Song, G.Y.; Na, Q.; Wang, D.; Qiao, C. Microarray expression profile of lncRNAs and mRNAs in the placenta of non-diabetic macrosomia. J. Dev. Orig. Health Dis. 2018, 9, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Meng, T.; Liu, X.; Sun, M.; Tong, C.; Liu, J.; Wang, H.; Du, J. Long non-coding RNA MALAT-1 is downregulated in preeclampsia and regulates proliferation, apoptosis, migration and invasion of JEG-3 trophoblast cells. Int. J. Clin. Exp. Path. 2015, 8, 12718. [Google Scholar]
- Feng, C.; Cheng, L.; Jin, J.; Liu, X.; Wang, F. Long non-coding RNA MALAT1 regulates trophoblast functions through VEGF/VEGFR1 signaling pathway. Arch. Gynecol. Obstet. 2021, 304, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, T.; Huang, S.; Zuo, Q.; Jiang, Z.; Yang, N.; Sun, L. LncRNA MALAT1 affects the migration and invasion of trophoblast cells by regulating FOS expression in early-onset preeclampsia. Pregnancy Hypertens. 2020, 21, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.Y.; Wang, X.H.; Liu, K.; Zhang, J.L. LncRNA MALAT1 regulates trophoblast cells migration and invasion via miR-206/IGF-1 axis. Cell Cycle 2020, 19, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Xu, Y.; Zou, Y.; Zuo, Q.; Huang, S.; Wang, S.; Lu, X.; He, X.; Wang, J.; Wang, T.; et al. Long Noncoding RNA 00473 Is Involved in Preeclampsia by LSD1 Binding-Regulated TFPI2 Transcription in Trophoblast Cells. Mol. Ther. Nucleic Acids 2018, 12, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.H.; Deng, W.B.; Liu, Y.F.; Liang, Y.X.; Fan, Z.M.; Gu, X.W.; Liu, J.L.; Sha, A.G.; Diao, H.L.; Yang, Z.M. Non-coding RNA LINC00473 mediates decidualization of human endometrial stromal cells in response to cAMP signaling. Sci. Rep. 2016, 6, 22744. [Google Scholar] [CrossRef] [Green Version]
- Chi, Z.; Gao, Q.; Sun, Y.; Zhou, F.; Wang, H.; Shu, X.; Zhang, M. LINC00473 downregulation facilitates trophoblast cell migration and invasion via the miR-15a-5p/LITAF axis in pre-eclampsia. Environ. Toxicol. 2021, 36, 1618–1627. [Google Scholar] [CrossRef]
- Liu, C.; Li, H.; Zhang, Y.; Ding, H. Long intergenic noncoding RNA 00473 promoting migration and invasion of trophoblastic cell line HTR-8/SVneo via regulating miR-424-5p-mediated wnt3a/β-catenin signaling pathway. J. Obstet. Gynaecol. Res. 2021, 47, 3034–3046. [Google Scholar] [CrossRef]
- Jiao, S.; Wang, S.Y.; Huang, Y. LncRNA PRNCR1 promoted the progression of eclampsia by regulating the MAPK signal pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 3635–3642. [Google Scholar] [CrossRef]
- Li, J.L.; Li, R.; Gao, Y.; Guo, W.C.; Shi, P.X.; Li, M. LncRNA CCAT1 promotes the progression of preeclampsia by regulating CDK4. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1216–1223. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Kuang, L.Y.; He, F.; Du, L.L.; Li, Q.L.; Sun, W.; Zhou, Y.M.; Li, X.M.; Li, X.Y.; Chen, D.J. The Role and Molecular Mechanism of Long Nocoding RNA-MEG3 in the Pathogenesis of Preeclampsia. Reprod. Sci. 2018, 25, 1619–1628. [Google Scholar] [CrossRef]
- Xu, Y.; Ge, Z.; Zhang, E.; Zuo, Q.; Huang, S.; Yang, N.; Wu, D.; Zhang, Y.; Chen, Y.; Xu, H.; et al. The lncRNA TUG1 modulates proliferation in trophoblast cells via epigenetic suppression of RND3. Cell Death Dis. 2017, 8, e3104. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, J.; Su, D.M.; Guan, L.N.; Mu, W.H.; Yu, M.; Ma, X.; Yang, R.J. lncRNA TUG1 modulates proliferation, apoptosis, invasion, and angiogenesis via targeting miR-29b in trophoblast cells. Hum. Genom. 2019, 13, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Wang, L.; Gao, M.; Guan, H. Long non-coding RNA TUG 1 regulates the migration and invasion of trophoblast-like cells through sponging miR-204-5p. Clin. Exp. Pharmacol. Physiol. 2019, 46, 380–388. [Google Scholar] [CrossRef]
- Zhang, W.; Zhou, Y.; Ding, Y. Lnc-DC mediates the over-maturation of decidual dendritic cells and induces the increase in Th1 cells in preeclampsia. Am. J. Reprod. Immunol. 2017, 77, e12647. [Google Scholar] [CrossRef]
- Song, X.; Rui, C.; Meng, L.; Zhang, R.; Shen, R.; Ding, H.; Li, J.; Li, J.; Long, W. Long non-coding RNA RPAIN regulates the invasion and apoptosis of trophoblast cell lines via complement protein C1q. Oncotarget 2017, 8, 7637–7646. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Chen, H.; Kong, W.; Zhang, Y.; Cao, L.; Gao, L.; Zhou, R. Down-regulated long non-coding RNA-ATB in preeclampsia and its effect on suppressing migration, proliferation, and tube formation of trophoblast cells. Placenta 2017, 49, 80–87. [Google Scholar] [CrossRef]
- Yin, Y.; Zhang, J.; Yu, H.; Liu, M.; Zheng, X.; Zhou, R. Effect of lncRNA-ATB/miR-651-3p/Yin Yang 1 pathway on trophoblast-endothelial cell interaction networks. J. Cell. Mol. Med. 2021, 25, 5391–5403. [Google Scholar] [CrossRef]
- Cao, C.; Li, J.; Li, J.; Liu, L.; Cheng, X.; Jia, R. Long Non-Coding RNA Uc.187 Is Upregulated in Preeclampsia and Modulates Proliferation, Apoptosis, and Invasion of HTR-8/SVneo Trophoblast Cells. J. Cell. Biochem. 2017, 118, 1462–1470. [Google Scholar] [CrossRef]
- Zuo, Q.; Huang, S.; Zou, Y.; Xu, Y.; Jiang, Z.; Zou, S.; Xu, H.; Sun, L. The Lnc RNA SPRY4-IT1 Modulates Trophoblast Cell Invasion and Migration by Affecting the Epithelial-Mesenchymal Transition. Sci. Rep. 2016, 6, 37183. [Google Scholar] [CrossRef] [PubMed]
- Oudejans, C.B.; Poutsma, A.; Michel, O.J.; Thulluru, H.K.; Mulders, J.; van de Vrugt, H.J.; Sistermans, E.A.; van Dijk, M. Noncoding RNA-regulated gain-of-function of STOX2 in Finnish pre-eclamptic families. Sci. Rep. 2016, 6, 32129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.J.; Ballabio, A.; Rupert, J.L.; Lafreniere, R.G.; Grompe, M.; Tonlorenzi, R.; Willard, H.F. A gene from the region of the human X inactivation centre is expressed exclusively from the inactive X chromosome. Nature 1991, 349, 38. [Google Scholar] [CrossRef] [PubMed]
- Erwin, J.A.; del Rosario, B.; Payer, B.; Lee, J.T. An ex vivo model for imprinting: Mutually exclusive binding of Cdx2 and Oct4 as a switch for imprinted and random X-inactivation. Genetics 2012, 192, 857–868. [Google Scholar] [CrossRef] [Green Version]
- Penkala, I.; Wang, J.; Syrett, C.M.; Goetzl, L.; López, C.B.; Anguera, M.C. lncRHOXF1, a Long Noncoding RNA from the X Chromosome That Suppresses Viral Response Genes during Development of the Early Human Placenta. Mol. Cell. Biol. 2016, 36, 1764–1775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Gong, Y. LncRNA-TCL6 promotes early abortion and inhibits placenta implantation via the EGFR pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7105–7112. [Google Scholar]
- Li, M.; Zhang, H.; Kong, Q. Long non-coding RNA IGF2-AS promotes trophoblast cell proliferation, migration, and invasion by regulating miR-520g/N-cadherin axis. J. Obstet. Gynaecol. Res. 2021, 47, 3047–3059. [Google Scholar] [CrossRef]
- Wang, Q.; Lu, X.; Li, C.; Zhang, W.; Lv, Y.; Wang, L.; Wu, L.; Meng, L.; Fan, Y.; Ding, H.; et al. Down-regulated long non-coding RNA PVT1 contributes to gestational diabetes mellitus and preeclampsia via regulation of human trophoblast cells. Biomed. Pharmacother. 2019, 120, 109501. [Google Scholar] [CrossRef]
- Gremlich, S.; Damnon, F.; Reymondin, D.; Braissant, O.; Schittny, J.; Baud, D.; Gerber, S.; Roth-Kleiner, M. The long non-coding RNA NEAT1 is increased in IUGR placentas, leading to potential new hypotheses of IUGR origin/development. Placenta 2014, 35, 44–49. [Google Scholar] [CrossRef]
- Xufei, F.; Xiujuan, Z.; Jianyi, L.; Liyan, Y.; Ting, Y.; Min, H. Up-regulation of LncRNA NEAT1 induces apoptosis of human placental trophoblasts. Free Radic. Res. 2020, 54, 678–686. [Google Scholar] [CrossRef]
- Quan, G.; Li, J. Circular RNAs: Biogenesis, expression and their potential roles in reproduction. J. Ovarian Res. 2018, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maass, P.G.; Glažar, P.; Memczak, S.; Dittmar, G.; Hollfinger, I.; Schreyer, L.; Sauer, A.V.; Toka, O.; Aiuti, A.; Luft, F.C.; et al. A map of human circular RNAs in clinically relevant tissues. J. Mol. Med. 2017, 95, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Feng, J.; Cheng, F.; Cui, X.; Gao, L.; Chen, Y.; Wang, F.; Zhong, T.; Li, Y.; Liu, L. Circular RNA expression profiles in placental villi from women with gestational diabetes mellitus. Biochem. Biophys. Res. Commun. 2018, 498, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; She, G.; Zhou, W.; Liu, K.; Miao, J.; Yu, B. Expression profile of circular RNAs in placentas of women with gestational diabetes mellitus. Endocr. J. 2019, 66, 431–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Wang, H.; Wu, X.; Long, W.; Zheng, F.; Kong, J.; Yu, B. The profile analysis of circular RNAs in human placenta of preeclampsia. Exp. Biol. Med. 2018, 243, 1109–1117. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Rao, H.; Chen, W.; Luo, X.; Tong, C.; Qi, H. Profiles of circular RNAs in human placenta and their potential roles related to preeclampsia. Biol. Reprod. 2018, 98, 705–712. [Google Scholar] [CrossRef] [Green Version]
- Ou, Y.; Liu, M.; Zhu, L.; Deng, K.; Chen, M.; Chen, H.; Zhang, J. The expression profile of circRNA and its potential regulatory targets in the placentas of severe pre-eclampsia. Taiwan. J. Obstet. Gynecol. 2019, 58, 769–777. [Google Scholar] [CrossRef]
- Ma, B.; Zhao, H.; Gong, L.; Xiao, X.; Zhou, Q.; Lu, H.; Cui, Y.; Xu, H.; Wu, S.; Tang, Y.; et al. Differentially expressed circular RNAs and the competing endogenous RNA network associated with preeclampsia. Placenta 2021, 103, 232–241. [Google Scholar] [CrossRef]
- Qian, Y.; Lu, Y.; Rui, C.; Qian, Y.; Cai, M.; Jia, R. Potential Significance of Circular RNA in Human Placental Tissue for Patients with Preeclampsia. Cell. Physiol. Biochem. 2016, 39, 1380–1390. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, X.; Gao, W.; Sun, M.; Chen, H.; Meng, T. Circular RNA VRK1 facilitates pre-eclampsia progression via sponging miR-221-3P to regulate PTEN/Akt. J. Cell. Mol. Med. 2021, 26, 1826–1841. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, H.; Zhang, Y.; Shi, J.; Chen, R.; Xiao, X. CircSFXN1 regulates the behaviour of trophoblasts and likely mediates preeclampsia. Placenta 2020, 101, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Gai, S.; Sun, L.; Wang, H.; Yang, P. Circular RNA hsa_circ_0007121 regulates proliferation, migration, invasion, and epithelial–mesenchymal transition of trophoblast cells by miR-182-5p/PGF axis in preeclampsia. Open Med. 2020, 15, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Niu, X.; Li, Q.; Zhao, Y.; Chen, X.; Sun, H. Circ_0085296 suppresses trophoblast cell proliferation, invasion, and migration via modulating miR-144/E-cadherin axis. Placenta 2020, 97, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Zhang, X.; Li, T.; Xie, R.; Zhou, J.; Luo, Y.; Yang, C. CircZDHHC20 represses the proliferation, migration and invasion in trophoblast cells by miR-144/GRHL2 axis. Cancer Cell Int. 2020, 20, 1–11. [Google Scholar] [CrossRef]
- Tang, R.; Zhang, Z.; Han, W. CircLRRK1 targets miR-223-3p to inhibit the proliferation, migration and invasion of trophoblast cells by regulating the PI3K/AKT signaling pathway. Placenta 2021, 104, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Ou, Y.; Zhu, L.; Wei, X.; Bai, S.; Chen, M.; Chen, H.; Zhang, J. Circular RNA circ_0111277 attenuates human trophoblast cell invasion and migration by regulating miR-494/HTRA1/Notch-1 signal pathway in pre-eclampsia. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, L.; Jia, J.; Ye, L.; Wang, Y.; Zhou, B.; Zhou, R. CircHIPK3 is decreased in preeclampsia and affects migration, invasion, proliferation, and tube formation of human trophoblast cells. Placenta 2019, 85, 1–8. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, H.; Yang, J.; Long, W.; Zhang, B.; Liu, J.; Yu, B. Down-regulated circPAPPA suppresses the proliferation and invasion of trophoblast cells via the miR-384/STAT3 pathway. Biosci. Rep. 2019, 39, BSR20191965. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.Y.; Zheng, L.L.; Huang, J.; Kong, H.F.; Chang, Y.J.; Wang, F.; Xin, H. CircTRNC18 inhibits trophoblast cell migration and epithelial–mesenchymal transition by regulating miR-762/Grhl2 pathway in pre-eclampsia. RNA Biol. 2019, 16, 1565–1573. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Yu, N.; Fan, L.; Chen, S.H.; Wu, J.L. Circ_0063517 acts as ceRNA, targeting the miR-31-5p-ETBR axis to regulate angiogenesis of vascular endothelial cells in preeclampsia. Life Sci. 2020, 244, 117306. [Google Scholar] [CrossRef]
- Deng, N.; Lei, D.; Huang, J.; Yang, Z.; Fan, C.; Wang, S. Circular RNA expression profiling identifies hsa_circ_0011460 as a novel molecule in severe preeclampsia. Pregnancy Hypertens. 2019, 17, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Ao, J.; Li, X.; Zhang, H.; Wu, J.; Cheng, W. Competing endogenous RNA expression profiling in pre-eclampsia identifies hsa_circ_0036877 as a potential novel blood biomarker for early pre-eclampsia. Clin. Epigenet. 2018, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, H.; Zhang, Y.; Shi, J.; Chen, R. circCRAMP1L is a novel biomarker of preeclampsia risk and may play a role in preeclampsia pathogenesis via regulation of the MSP/RON axis in trophoblasts. BMC Pregnancy Childbirth 2020, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, J.; Xu, Z.; Yang, J.; Xu, Y.; Liu, Y.; Li, B.; Xie, J.; Li, J. Circular RNA hsa_circ_0000848 promotes trophoblast cell migration and invasion and inhibits cell apoptosis by sponging hsa-miR-6768-5p. Front. Cell Dev. Biol. 2020, 8, 278. [Google Scholar] [CrossRef]
- Wang, H.; Luo, C.; Wu, X.; Zhang, J.; Xu, Z.; Liu, Y.; Li, B.; Li, J.; Xie, J. Circular RNA hsa_circ_0081343 promotes trophoblast cell migration and invasion and inhibits trophoblast apoptosis by regulating miR-210-5p/DLX3 axis. Reprod. Biol. Endocrinol. 2021, 19, 1–11. [Google Scholar] [CrossRef]
- Yao, P.; Hu, G.; Niu, H. Hsa_circ_0074371 Regulates Proliferation, Apoptosis, Migration, and Invasion via the miR-582-3p/LRP6 Axis in Trophoblast Cells. Biochem. Genet. 2021, 60, 267–285. [Google Scholar] [CrossRef]
- Wang, D.; Na, Q.; Song, G.; Wang, Y.; Wang, Y. The role of circRNA-SETD2/miR-519a/PTEN axis in fetal birth weight through regulating trophoblast proliferation. BioMed Res. Int. 2020, 2020, 9809632. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, G.; Tao, F.; Cao, Y.; Han, W.; Li, Q. circ-ZUFSP regulates trophoblasts migration and invasion through sponging miR-203 to regulate STOX1 expression. Biochem. Biophys. Res. Commun. 2020, 531, 472–479. [Google Scholar] [CrossRef]
- Umu, S.U.; Langseth, H.; Bucher-Johannessen, C.; Fromm, B.; Keller, A.; Meese, E.; Lauritzen, M.; Leithaug, M.; Lyle, R.; Rounge, T.B. A comprehensive profile of circulating RNAs in human serum. RNA Biol. 2018, 15, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, U.; Naderi-Meshkin, H.; Gupta, S.K.; Wolkenhauer, O.; Vera, J. The RNA world in the 21st century-a systems approach to finding non-coding keys to clinical questions. Brief. Bioinform. 2016, 17, 380–392. [Google Scholar] [CrossRef] [Green Version]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, S.; Weber, J.; Baxter, D.; Galas, D.J. Export of microRNAs and microRNA-protective protein by mammalian cells. Nucleic Acids Res. 2010, 38, 7248–7259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Rodosthenous, R.S.; Burris, H.H.; Sanders, A.P.; Just, A.C.; Dereix, A.E.; Svensson, K.; Solano, M.; Téllez-Rojo, M.M.; Wright, R.O.; Baccarelli, A.A. Second trimester extracellular microRNAs in maternal blood and fetal growth: An exploratory study. Epigenetics 2017, 12, 804–810. [Google Scholar] [CrossRef] [Green Version]
- Poon, L.L.; Leung, T.N.; Lau, T.K.; Lo, Y.M. Presence of fetal RNA in maternal plasma. Clin. Chem. 2000, 46, 1832–1834. [Google Scholar] [CrossRef] [Green Version]
- Tsui, N.B.Y.; Jiang, P.; Wong, Y.F.; Leung, T.Y.; Chan, K.C.A.; Chiu, R.W.K.; Sun, H.; Lo, Y.M.D. Maternal plasma RNA sequencing for genome-wide transcriptomic profiling and identification of pregnancy-associated transcripts. Clin. Chem. 2014, 60, 954–962. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, C.L.; Walker, S.P.; Tong, S. Measuring circulating placental RNAs to non-invasively assess the placental transcriptome and to predict pregnancy complications. Prenat. Diagn. 2016, 36, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Ngo, T.T.M.; Moufarrej, M.N.; Rasmussen, M.L.H.; Camunas-Soler, J.; Pan, W.; Okamoto, J.; Neff, N.F.; Liu, K.; Wong, R.J.; Downes, K.; et al. Noninvasive blood tests for fetal development predict gestational age and preterm delivery. Science 2018, 360, 1133–1136. [Google Scholar] [CrossRef] [Green Version]
- Traver, S.; Assou, S.; Scalici, E.; Haouzi, D.; Al-Edani, T.; Belloc, S.; Hamamah, S. Cell-free nucleic acids as non-invasive biomarkers of gynecological cancers, ovarian, endometrial and obstetric disorders and fetal aneuploidy. Hum. Reprod. Update 2014, 20, 905–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scalici, E.; Traver, S.; Mullet, T.; Molinari, N.; Ferrières, A.; Brunet, C.; Belloc, S.; Hamamah, S. Circulating microRNAs in follicular fluid, powerful tools to explore in vitro fertilization process. Sci. Rep. 2016, 6, 24976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Gu, W.W.; Gu, Y.; Yan, N.N.; Mao, Y.Y.; Zhen, X.X.; Wang, J.M.; Yang, J.; Shi, H.J.; Zhang, X.; et al. Association of the peripheral blood levels of circulating microRNAs with both recurrent miscarriage and the outcomes of embryo transfer in an in vitro fertilization process. J. Transl. Med. 2018, 16, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoffe, L.; Polsky, A.; Gilam, A.; Raff, C.; Mecacci, F.; Ognibene, A.; Crispi, F.; Gratacós, E.; Kanety, H.; Mazaki-Tovi, S.; et al. Early diagnosis of gestational diabetes mellitus using circulating microRNAs. Eur. J. Endocrinol. 2019, 181, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wu, J.; Zhao, Z.; Wang, J.; Chen, Z. Circulating LncRNA Serve as Fingerprint for Gestational Diabetes Mellitus Associated with Risk of Macrosomia. Cell. Physiol. Biochem. 2018, 48, 1012–1018. [Google Scholar] [CrossRef]
- Enquobahrie, D.A.; Wander, P.L.; Tadesse, M.G.; Qiu, C.; Holzman, C.; Williams, M.A. Maternal pre-pregnancy body mass index and circulating microRNAs in pregnancy. Obes. Res. Clin. Pract. 2017, 11, 464–474. [Google Scholar] [CrossRef]
- Adaikalakoteswari, A.; Vatish, M.; Alam, M.T.; Ott, S.; Kumar, S.; Saravanan, P. Low Vitamin B12 in Pregnancy Is Associated with Adipose-Derived Circulating miRs Targeting PPARγ and Insulin Resistance. J. Clin. Endocrinol. Metab. 2017, 102, 4200–4209. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, T.M.; Kaitu’u-Lino, T.J.; Walker, S.P.; Dane, K.M.; Lockie, E.B.; Tong, S.; Whitehead, C.L.; Hui, L. Variable effect of maternal oral glucose load on circulating cell-free placental mRNAs. J. Matern. Fetal Neonatal Med. 2017, 30, 501–503. [Google Scholar] [CrossRef]
- Gardiner, A.S.; Gutierrez, H.L.; Luo, L.; Davies, S.; Savage, D.D.; Bakhireva, L.N.; Perrone-Bizzozero, N.I. Alcohol Use During Pregnancy is Associated with Specific Alterations in MicroRNA Levels in Maternal Serum. Alcohol. Clin. Exp. Res. 2016, 40, 826–837. [Google Scholar] [CrossRef] [Green Version]
- Tseng, A.M.; Mahnke, A.H.; Wells, A.B.; Salem, N.A.; Allan, A.M.; Roberts, V.H.; Newman, N.; Walter, N.A.; Kroenke, C.D.; Grant, K.A.; et al. Maternal circulating miRNAs that predict infant FASD outcomes influence placental maturation. Life Sci. Alliance 2019, 2, e201800252. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, I.; Nahmias, N.; Novoselsky Persky, M.; Greenfield, C.; Goldman-Wohl, D.; Hurwitz, A.; Haimov-Kochman, R.; Yagel, S.; Imbar, T. Elevated circulating micro-ribonucleic acid (miRNA)-200b and miRNA-429 levels in anovulatory women. Fertil. Steril. 2017, 107, 269–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoffe, L.; Gilam, A.; Yaron, O.; Polsky, A.; Farberov, L.; Syngelaki, A.; Nicolaides, K.; Hod, M.; Shomron, N. Early Detection of Preeclampsia Using Circulating Small non-coding RNA. Sci. Rep. 2018, 8, 3401. [Google Scholar] [CrossRef] [PubMed]
- Munchel, S.; Rohrback, S.; Randise-Hinchliff, C.; Kinnings, S.; Deshmukh, S.; Alla, N.; Tan, C.; Kia, A.; Greene, G.; Leety, L.; et al. Circulating transcripts in maternal blood reflect a molecular signature of early-onset preeclampsia. Sci. Transl. Med. 2020, 12, eaaz0131. [Google Scholar] [CrossRef] [PubMed]
- Jairajpuri, D.S.; Malalla, Z.H.; Mahmood, N.; Almawi, W.Y. Circulating microRNA expression as predictor of preeclampsia and its severity. Gene 2017, 627, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.W.; Shim, J.I.; Shim, S.H.; Shin, Y.j.; Shim, S.H.; Chang, S.W.; Cha, D.H. Global gene expression analysis of cell-free RNA in amniotic fluid from women destined to develop preeclampsia. Medicine 2019, 98, e13971. [Google Scholar] [CrossRef]
- Gu, M.; Zheng, A.; Tu, W.; Zhao, J.; Li, L.; Li, M.; Han, S.; Hu, X.; Zhu, J.; Pan, Y.; et al. Circulating LncRNAs as Novel, Non-Invasive Biomarkers for Prenatal Detection of Fetal Congenital Heart Defects. Cell. Physiol. Biochem. 2016, 38, 1459–1471. [Google Scholar] [CrossRef]
- Biró, O.; Rigó, J.; Nagy, B. Noninvasive prenatal testing for congenital heart disease—Cell-free nucleic acid and protein biomarkers in maternal blood. J. Matern. Fetal Neonatal Med. 2018, 33, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, C.L.; McNamara, H.; Walker, S.P.; Alexiadis, M.; Fuller, P.J.; Vickers, D.K.; Hannan, N.J.; Hastie, R.; Tuohey, L.; Kaitu’u-Lino, T.J.; et al. Identifying late-onset fetal growth restriction by measuring circulating placental RNA in the maternal blood at 28 weeks’ gestation. Am. J. Obstet. Gynecol. 2016, 214, 521.e1–521.e8. [Google Scholar] [CrossRef]
- Carbone, I.F.; Conforti, A.; Picarelli, S.; Morano, D.; Alviggi, C.; Farina, A. Circulating nucleic acids in maternal plasma and serum in pregnancy complications: Are they really useful in clinical practice? a systematic review. Mol. Diagn. Ther. 2020, 24, 409–431. [Google Scholar] [CrossRef]
- Salomon, C.; Guanzon, D.; Scholz-Romero, K.; Longo, S.; Correa, P.; Illanes, S.E.; Rice, G.E. Placental Exosomes as Early Biomarker of Preeclampsia: Potential Role of Exosomal MicroRNAs Across Gestation. J. Clin. Endocrinol. Metab. 2017, 102, 3182–3194. [Google Scholar] [CrossRef]
- Wang, Y.; Du, X.; Wang, J. Transfer of miR-15a-5p by placental exosomes promotes pre-eclampsia progression by regulating PI3K/AKT signaling pathway via CDK1. Mol. Immunol. 2020, 128, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, Y.; Mouillet, J.F.; Coyne, C.B.; Sadovsky, Y. Review: Placenta-specific microRNAs in exosomes—Good things come in nano-packages. Placenta 2014, 35, S69–S73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, R.J.; Jensen, S.S.; Lim, J.W.E. Proteomic profiling of exosomes: Current perspectives. Proteomics 2008, 8, 4083–4099. [Google Scholar] [CrossRef]
- Sabapatha, A.; Gercel-Taylor, C.; Taylor, D.D. Specific isolation of placenta-derived exosomes from the circulation of pregnant women and their immunoregulatory consequences. Am. J. Reprod. Immunol. 2006, 56, 345–355. [Google Scholar] [CrossRef]
- Tong, M.; Abrahams, V.M.; Chamley, L.W. Immunological effects of placental extracellular vesicles. Immunol. Cell Biol. 2018, 96, 714–722. [Google Scholar] [CrossRef]
- Favaro, R.R.; Murrieta-Coxca, J.M.; Gutiérrez-Samudio, R.N.; Morales-Prieto, D.M.; Markert, U.R. Immunomodulatory properties of extracellular vesicles in the dialogue between placental and immune cells. Am. J. Reprod. Immunol. 2021, 85, e13383. [Google Scholar] [CrossRef]
- Tannetta, D.; Collett, G.; Vatish, M.; Redman, C.; Sargent, I. Syncytiotrophoblast extracellular vesicles—Circulating biopsies reflecting placental health. Placenta 2017, 52, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Chiarello, D.I.; Salsoso, R.; Toledo, F.; Mate, A.; Vázquez, C.M.; Sobrevia, L. Foetoplacental communication via extracellular vesicles in normal pregnancy and preeclampsia. Mol. Asp. Med. 2018, 60, 69–80. [Google Scholar] [CrossRef]
- Schmorl, G. Pathologisch-Anatomische Untersuchungen über Puerperal-Eklampsie; Vogel: Munich, Germany, 1893. [Google Scholar]
- Kumpel, B.; King, M.J.; Sooranna, S.; Jackson, D.; Eastlake, J.; Cheng, R.; Johnson, M. Phenotype and mRNA expression of syncytiotrophoblast microparticles isolated from human placenta. Ann. N. Y. Acad. Sci. 2008, 1137, 144–147. [Google Scholar] [CrossRef]
- Redman, C.W.G.; Tannetta, D.S.; Dragovic, R.A.; Gardiner, C.; Southcombe, J.H.; Collett, G.P.; Sargent, I.L. Review: Does size matter? Placental debris and the pathophysiology of pre-eclampsia. Placenta 2012, 33, S48–S54. [Google Scholar] [CrossRef] [PubMed]
- Chamley, L.W.; Holland, O.; Chen, Q.; Viall, C.; Stone, P.; Abumaree, M. Where is the maternofetal interface? Placenta 2014, 35, S74–S80. [Google Scholar] [CrossRef] [PubMed]
- Homer, H.; Rice, G.E.; Salomon, C. Review: Embryo- and endometrium-derived exosomes and their potential role in assisted reproductive treatments-liquid biopsies for endometrial receptivity. Placenta 2017, 54, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Condrat, C.E.; Varlas, V.N.; Duică, F.; Antoniadis, P.; Danila, C.A.; Cretoiu, D.; Suciu, N.; Crețoiu, S.M.; Voinea, S.C. Pregnancy-Related Extracellular Vesicles Revisited. Int. J. Mol. Sci. 2021, 22, 3904. [Google Scholar] [CrossRef]
- Nakahara, A.; Nair, S.; Ormazabal, V.; Elfeky, O.; Garvey, C.E.; Longo, S.; Salomon, C. Circulating placental extracellular vesicles and their potential roles during pregnancy. Ochsner J. 2020, 20, 439–445. [Google Scholar] [CrossRef]
- Biró, O.; Alasztics, B.; Molvarec, A.; Joó, J.; Nagy, B.; Rigó, J. Various levels of circulating exosomal total-miRNA and miR-210 hypoxamiR in different forms of pregnancy hypertension. Pregnancy Hypertens. 2017, 10, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Ouyang, Y.; Sadovsky, E.; Parks, W.T.; Chu, T.; Sadovsky, Y. Unique microRNA signals in plasma exosomes from pregnancies complicated by preeclampsia. Hypertension 2020, 75, 762–771. [Google Scholar] [CrossRef]
- Zabel, R.R.; Bär, C.; Ji, J.; Schultz, R.; Hammer, M.; Groten, T.; Schleussner, E.; Morales-Prieto, D.M.; Markert, U.R.; Favaro, R.R. Enrichment and characterization of extracellular vesicles from ex vivo one-sided human placenta perfusion. Am. J. Reprod. Immunol. 2021, 86, e13377. [Google Scholar] [CrossRef]
- Redman, C.W.G.; Sargent, I.L. Circulating microparticles in normal pregnancy and pre-eclampsia. Placenta 2008, 29 (Suppl. A), S73–S77. [Google Scholar] [CrossRef]
- Delorme-Axford, E.; Donker, R.B.; Mouillet, J.F.; Chu, T.; Bayer, A.; Ouyang, Y.; Wang, T.; Stolz, D.B.; Sarkar, S.N.; Morelli, A.E.; et al. Human placental trophoblasts confer viral resistance to recipient cells. Proc. Natl. Acad. Sci. USA 2013, 110, 12048–12053. [Google Scholar] [CrossRef] [Green Version]
- Schuster, J.; Cheng, S.B.; Padbury, J.; Sharma, S. Placental extracellular vesicles and pre-eclampsia. Am. J. Reprod. Immunol. 2021, 85, e13297. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Matsubara, Y.; Uchikura, Y.; Sugiyama, T. Pathophysiology of Preeclampsia: The Role of Exosomes. Int. J. Mol. Sci. 2021, 22, 2572. [Google Scholar] [CrossRef] [PubMed]
- Cronqvist, T.; Tannetta, D.; Mörgelin, M.; Belting, M.; Sargent, I.; Familari, M.; Hansson, S.R. Syncytiotrophoblast derived extracellular vesicles transfer functional placental miRNAs to primary human endothelial cells. Sci. Rep. 2017, 7, 4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Li, Y.; Li, R.; Diao, Z.; Yany, M.; Wu, M.; Sun, H.; Yan, G.; Hu, Y. Placenta-associated serum exosomal miR-155 derived from patients with preeclampsia inhibits eNOS expression in human umbilical vein endothelial cells. Int. J. Mol. Med. 2018, 41, 1731–1739. [Google Scholar] [CrossRef] [Green Version]
- Cronqvist, T.; Saljé, K.; Familari, M.; Guller, S.; Schneider, H.; Gardiner, C.; Sargent, I.L.; Redman, C.W.; Mörgelin, M.; Åkerström, B.; et al. Syncytiotrophoblast vesicles show altered micro-RNA and haemoglobin content after ex-vivo perfusion of placentas with haemoglobin to mimic preeclampsia. PLoS ONE 2014, 9, e90020. [Google Scholar] [CrossRef]
- Truong, G.; Guanzon, D.; Kinhal, V.; Elfeky, O.; Lai, A.; Longo, S.; Nuzhat, Z.; Palma, C.; Scholz-Romero, K.; Menon, R.; et al. Oxygen tension regulates the miRNA profile and bioactivity of exosomes released from extravillous trophoblast cells—Liquid biopsies for monitoring complications of pregnancy. PLoS ONE 2017, 12, e0174514. [Google Scholar] [CrossRef] [Green Version]
- Salomon, C.; Scholz-Romero, K.; Sarker, S.; Sweeney, E.; Kobayashi, M.; Correa, P.; Longo, S.; Duncombe, G.; Mitchell, M.D.; Rice, G.E.; et al. Gestational Diabetes Mellitus Is Associated with Changes in the Concentration and Bioactivity of Placenta-Derived Exosomes in Maternal Circulation Across Gestation. Diabetes 2016, 65, 598–609. [Google Scholar] [CrossRef] [Green Version]
- Gillet, V.; Ouellet, A.; Stepanov, Y.; Rodosthenous, R.S.; Croft, E.K.; Brennan, K.; Abdelouahab, N.; Baccarelli, A.; Takser, L. miRNA profiles in extracellular vesicles from serum early in pregnancies complicated by gestational diabetes mellitus. J. Clin. Endocrinol. Metab. 2019, 104, 5157–5169. [Google Scholar] [CrossRef]
- Nair, S.; Jayabalan, N.; Guanzon, D.; Palma, C.; Scholz-Romero, K.; Elfeky, O.; Zuñiga, F.; Ormazabal, V.; Diaz, E.; Rice, G.E.; et al. Human placental exosomes in gestational diabetes mellitus carry a specific set of miRNAs associated with skeletal muscle insulin sensitivity. Clin. Sci. 2018, 132, 2451–2467. [Google Scholar] [CrossRef]
- Fallen, S.; Baxter, D.; Wu, X.; Kim, T.K.; Shynlova, O.; Lee, M.Y.; Scherler, K.; Lye, S.; Hood, L.; Wang, K. Extracellular vesicle RNAs reflect placenta dysfunction and are a biomarker source for preterm labour. J. Cell. Mol. Med. 2018, 22, 2760–2773. [Google Scholar] [CrossRef] [Green Version]
- Ying, X.; Jin, X.; Zhu, Y.; Liang, M.; Chang, X.; Zheng, L. Exosomes released from decidual macrophages deliver miR-153-3p, which inhibits trophoblastic biological behavior in unexplained recurrent spontaneous abortion. Int. Immunopharmacol. 2020, 88, 106981. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, Y.; Bayer, A.; Chu, T.; Tyurin, V.A.; Kagan, V.E.; Morelli, A.E.; Coyne, C.B.; Sadovsky, Y. Isolation of human trophoblastic extracellular vesicles and characterization of their cargo and antiviral activity. Placenta 2016, 47, 86–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, T.F.; Donaldson, B.; Bouqueau, M.; Kampmann, B.; Holder, B. Macrophage- but not monocyte-derived extracellular vesicles induce placental pro-inflammatory responses. Placenta 2018, 69, 92–95. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tool | Descripton | Citation |
|---|---|---|
| (a) Homology-based tools for ncRNA identification | ||
| Blast/RNAcentral | sequence based search | |
| Infernal/Rfam | prediction based on covariances of secondary structures | |
| (b) Tools for de novo identification of ncRNAs | ||
| RNAz | predicting structurally conserved and thermodynamically stable RNA secondary structures in multiple sequence (genome) alignments | [22] |
| QRNA | prediction based on comparative genome sequence analysis | [23] |
| RNAsamba | tool to predict the coding potential of RNA molecules from sequence information using a neural network-based that models both the whole sequence and the ORF to identify patterns that distinguish coding from non-coding transcripts | [24] |
| FEELnc | alignment-free program that accurately annotates lncRNAs based on a Random Forest model trained with general features such as multi k-mer frequencies and relaxed open reading frames | [25] |
| LGC | discriminating lncRNAs from protein-coding RNAs across diverse species that range from plants to mammals | [26] |
| CPAT | novel alignment-free method: recognizes coding and noncoding transcripts from a large pool of candidates | [27] |
| COME | identification and characterization of novel lncRNAs | [28] |
| PLEK | algorithm to distinguish lncRNAs from messenger RNAs (mRNAs), in the absence of genomic sequences or annotations | [29] |
| PhyloCSF | comparative genomics method that analyzes a multispecies nucleotide sequence alignment to determine whether it is likely to represent a conserved protein-coding region, based on a formal statistical comparison of phylogenetic codon models | [30] |
| (c) Identification of ncRNAs with the help of transcriptomic data | ||
| lncRScan-SVM | tool for predicting the lncRNAs (classifying protein coding and lncRNA transcripts using support vector machine) | [31] |
| slncky | lncRNA discovery tool that produces a high-quality set of lncRNAs from RNA-sequencing data and further uses evolutionary constraint to prioritize lncRNAs that are likely to be functionally important | [32] |
| CNCI | effective for classifying incomplete transcripts and sense–antisense pairs (highly accurate classification of transcripts assembled from whole-transcriptome sequencing data in a cross-species manner, that demonstrated gene evolutionary divergence between vertebrates, and invertebrates, or between plants, and provided a long non-coding RNA catalog of orangutan) | [33] |
| CREMA | tool that can be used to rank long non-protein coding RNA predictions for use in conjunction with gene expression studies | [34] |
| miRNA | Function | Citation |
|---|---|---|
| let-7d | down-regulation inhibits the proliferation and invasion of trophoblast cells | [71] |
| miR-15b | inhibits trophoblast cell invasion and endothelial cell tube formation by suppressing the expression of argonaute 2 | [83] |
| miR-17, miR-20a, miR-20b | are differentially expressed in PE, regulating EPHB4 and ephrin-B2 expression in trophoblast and endothelial cells via the same “seed” sequence | [98] |
| mir-20b | may contribute to PE through inhibiting proliferation, invasion and migration of placental trophoblast cells by targeting MCL-1 | [99] |
| miR-22 | up-regulation is modulating production of androgen and estrogen and up-regulated in PE placenta | [100] |
| miR-30a-3p | expression is significantly increased in PE and might be involved in the pathogenesis by targeting IGF-1 and regulating the invasion and apoptosis of trophoblast cells | [78] |
| miR-34a | hypo-methylation of the miR-34a promoter is associated with PE and PE severity | [101] |
| regulates trophoblast invasion through the Notch signal transduction | [102] | |
| contributes to trophoblast cell apoptosis in PE by targeting BCL-2 | [87] | |
| down-regulation miR-34a-5p improves invasion and migration of trophoblast cells by targetting SMAD4 | [103] | |
| miR-93 | inhibits MMP-2 and reduces migration and invasion of immortalized trophoblast cells | [76] |
| miR-128a | induces apoptosis of HTR-8/SVneo cells and thus may contribute to PE | [86] |
| miR-181a-5p | is increased in both the plasma and placenta of severe PE patients and suppresses the invasion and migration of HTR-8/SVneo cells by directly targeting IGF2BP2 | [74] |
| up-regulation induces apoptosis, and suppresses invasion in HTR-8/SVneo and JAR cells. | [104] | |
| miR-134 | down-regulates ITGB1 and inhibits infiltration of trophoblast cells in placenta of patients with PE | [79] |
| miR-135a-5p | promotes migration and invasion of trophoblast cells through targeting -TrCP | [105] |
| miR-137 | reduces the proliferation and migration of trophoblast cells by targeting ERR | [72] |
| miR-141 | up-regulated in PE and regulates trophoblast (JEG-3 and HTR-8/SVneo) proliferation and invasion and intercellular communication vie EVs | [18] |
| hypoxia-induced microRNA-141 regulates trophoblast apoptosis, invasion, and vascularization by blocking CXCL12/CXCR2/4 signal transduction | [106] | |
| miR-141-5p regulates ATF2 via effecting MAPK1/ERK2 signaling to promote preeclampsia | [107] | |
| miR-144 | may play an important role in the pathogenesis of PE through targeting PTEN in trophoblastic cells | [108] |
| MicroRNA-144-3p may participate in the pathogenesis of preeclampsia by targeting Cox-2 | [109] | |
| miR-195 | could promote cell invasion via directly targeting ActRIIB in trophoblast cells | [82] |
| is suggested to regulate PE by affecting placental proliferation, apoptosis, and angiogenesis | [110] | |
| miR-200 | miR-200c,-20a and -20b are involved in hydrogen sulfide stimulation of VEGF in trophoblasts | [111] |
| miR-203 | significantly increased in PE placenta inhibiting vascular endothelial growth factor A (VEGFA) | [112] |
| miR-218 | contributes to PE by targeting LASP1 to inhibit trophoblast invasion | [81] |
| miR-299 | up-regulation suppresses the invasion and migration of HTR-8/SVneo trophoblast cells partly via targeting HDAC2 | [75] |
| miR-320a | overexpression in PE placenta inhibits trophoblast cell invasion by targeting estrogen-related receptor-gamma (ERR), but not migration or proliferation | [80] |
| upregulation inhibits proliferation and invasion of trophoblast cells by targeting IL-4 | [113] | |
| miR-454 | promotes the proliferation and invasion of trophoblast cells by inhibiting EPHB4 expression | [73] |
| promotes the proliferation and invasion of trophoblast cells by downregulation of ALK7 | [114] | |
| miR-520c-3p | is suppressing inflammasome activation and inflammatory cascade by down-regulating NLRP3 | [115] |
| miR-520g | is suppressing the migration and invasion of trophoblast via at least partial inhibition of MMP2 translation inhibition | [77] |
| miR-4421 | is highly expressed in PE, which may promote the progression of PE by down-regulating the expression of CYP11B2 | [116] |
| miR-125b | involved in early PE development through regulation of Trop-2 expression | [117] |
| LncRNA | Function | Citation |
|---|---|---|
| Malat-1 | down-regulated in preeclampsia, regulates proliferation, apoptosis, migration and invasion of JEG-3 choriocarcinoma cells | [173,174] |
| affects the migration and invasion of trophoblast cell by regulating FOS expression | [175] | |
| regulates trophoblast cells migration and invasion via miR-206/IGF-1 axis | [176] | |
| LOC391533, LOC284100, CEACAMP8 | dysregulation seems to be associated with preeclampsia | [15] |
| linc00473 | down-regulated in the placenta of patients with severe PE. knockdown in trophoblast cell lines significantly inhibites cell proliferation and promotes apoptosis, whereas overexpression stimulates trophoblast proliferation. Further, linc00473 inhibited the expression of tissue factor pathway inhibitor 2 (TFPI2) through binding to lysine-specific demethylase 1 (LSD1) | [177] |
| mediates decidualization of human endometrial stromal cells in response to cAMP signaling | [178] | |
| downregulation facilitates trophoblast cell migration and invasion via the miR-15a-5p/LITAF axis in pre-eclampsia, | [179] | |
| Linc00473 mediates regulation of Wnt/-catenin signaling pathway by miR-424-5p and affects invasion and migration of the trophoblastic cell line HTR-8/SVneo | [180] | |
| PRNCR1 | promotes the progression of PE by modulating the MAPK signaling pathway | [181] |
| CCAT1 | is highly expressed in PE and can promote the progression of PE by inhibiting the expression of CDK4 | [182] |
| MEG3 | is lower expressed in the placenta of patients with PE, and its regulation of trophoblast cell epithelial-mesenchymal transition via the TGF- pathway inhibitor SMAD7 may be the molecular mechanism involved in the pathogenesis of PE | [183] |
| TUG1 | is modulating proliferation in trophoblast cells via epigenetic suppression of RND3 | [184] |
| is modulating proliferation, apoptosis, invasion, and angiogenesis via targeting miR-29b in trophoblast cells | [185] | |
| is regulating the migration and invasion of trophoblast-like cells through sponging miR-204-5p | [186] | |
| lnc-DCs | overexpression in dendritic cells mediates their maturation by phosphorylating STAT3 and induces the over-maturation of decidual dendritic cells in PE and leads to an increase in Th1 cells | [187] |
| RPAIN | regulates the invasion and apoptosis of trophoblast cell lines via complement protein C1q | [188] |
| ATB | down-regulated in PE placentas which was found to decrease migration, proliferation, and tube-formation of HTR-8/SVneo cells | [189] |
| functions as a competitive endogenous RNA of miR-651-3p to regulate YY1 on progress of spiral artery remodelling | [190] | |
| uc.187 | is up-regulated in preeclampsia and modulates proliferation, apoptosis, and invasion of HTR-8/SVneo cells | [191] |
| SPRY4-IT1 | modulates trophoblast cell invasion and migration by affecting the epithelial-mesenchymal transition | [192] |
| up-regulation modulates proliferation, migration, apoptosis, and network formation in HTR-8SV/neo cells | [16] | |
| NN | An lncRNA within intron 3 of the STOX2 gene which seems to regulate an essential trophoblast differentiation pathway | [193] |
| XIST | has a role in X chromosome inactivation in females, a process that is paternal specific in the trophoblast and random in the fetus | [194,195] |
| lncRHOXF1 | is the first example of an lncRNA from the X chromosome that regulates the host response to viral infections in human placental progenitor cells | [196] |
| LncRNA-TCL6 | plays a role in early abortion by inhibiting placental implantation via the EGFR pathway | [197] |
| LncRNaIGF2-AS | plays a role in recurrent miscarriage by regulating trophoblast functions | [198] |
| PVT1 | is down-regulated in GDM and PE | [199] |
| HOTAIR | plays an important role in suppressing angiogenesis of the human placenta by inhibiting the expression of VEGFA | [171] |
| NEAT1 | is increased in intrauterine growth retardation (IUGR) placentas but the pathomechanism is not yet clear; up-regulation is inducing apoptosis in HTR-8/SVneo cells | [200,201] |
| Disease | Circulating ncRNA | Citation |
|---|---|---|
| Early pregnancy loss | up-regulated hsa-let- 7c, hsa-miR-122 and down-regulated hsa-miR- 135a in plasma | [42] |
| Recurrent miscarriage | miR-27a-3p, miR-29a-3p, miR-100-5p and miR-127-3p are increased and miR-486-5p decreased in plasma | [244] |
| Fetal growth restriction | up-regulated miR-16-5p, miR-103-3p, miR-107-3p, and miR-27b-3p in plasma | [141] |
| Fetal congenital heart defects | lncRNAs ENST00000436681, ENST00000422826 are up-regulated and AA584040, AA709223 and BX478947 down-regulated in plasma | [257] |
| Placenta accreta spectrum | down-regulated miR-139-3p, miR-196a-5p, miR-518a-3p, and miR-671-3p in serum | [147] |
| Gestational diabetes mellitus | miR-223 and miR-23a are up-regulated in plasma | [245] |
| Preeclampsia | hsa-circ-0036877 is up-regulated in blood | [223] |
| Preeclampsia | circCRAMP1L circulating levels are significantly lower in plasma | [224] |
| Preeclampsia | miR-215, miR-155, miR-650, miR-210, miR-21 are up-regulated, and miR-18a, miR-19b1 down-regulated in plasma | [255] |
| Preeclampsia | hsa-miR-486-1-5p and hsa-miR-486-2-5p are up-regulated in exosomes | [261] |
| Preeclampsia | hsa-miR-210 is up-regulated in exosomes | [258] |
| Preeclampsia | miR-15a-5p is up-regulated in exosomes | [262] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Žarković, M.; Hufsky, F.; Markert, U.R.; Marz, M. The Role of Non-Coding RNAs in the Human Placenta. Cells 2022, 11, 1588. https://doi.org/10.3390/cells11091588
Žarković M, Hufsky F, Markert UR, Marz M. The Role of Non-Coding RNAs in the Human Placenta. Cells. 2022; 11(9):1588. https://doi.org/10.3390/cells11091588
Chicago/Turabian StyleŽarković, Milena, Franziska Hufsky, Udo R. Markert, and Manja Marz. 2022. "The Role of Non-Coding RNAs in the Human Placenta" Cells 11, no. 9: 1588. https://doi.org/10.3390/cells11091588
APA StyleŽarković, M., Hufsky, F., Markert, U. R., & Marz, M. (2022). The Role of Non-Coding RNAs in the Human Placenta. Cells, 11(9), 1588. https://doi.org/10.3390/cells11091588