
The Effects of Taurocholic Acid on Biliary Damage and Liver Fibrosis Are Mediated by Calcitonin-Gene-Related Peptide Signaling

 ,
,  , , ,
, , ,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animal Models
2.3. Isolated Cholangiocytes
2.4. Measurement of CGRP in Serum and Immunoreactivity of CALCRL in Liver Sections
2.5. Evaluation of Liver Damage
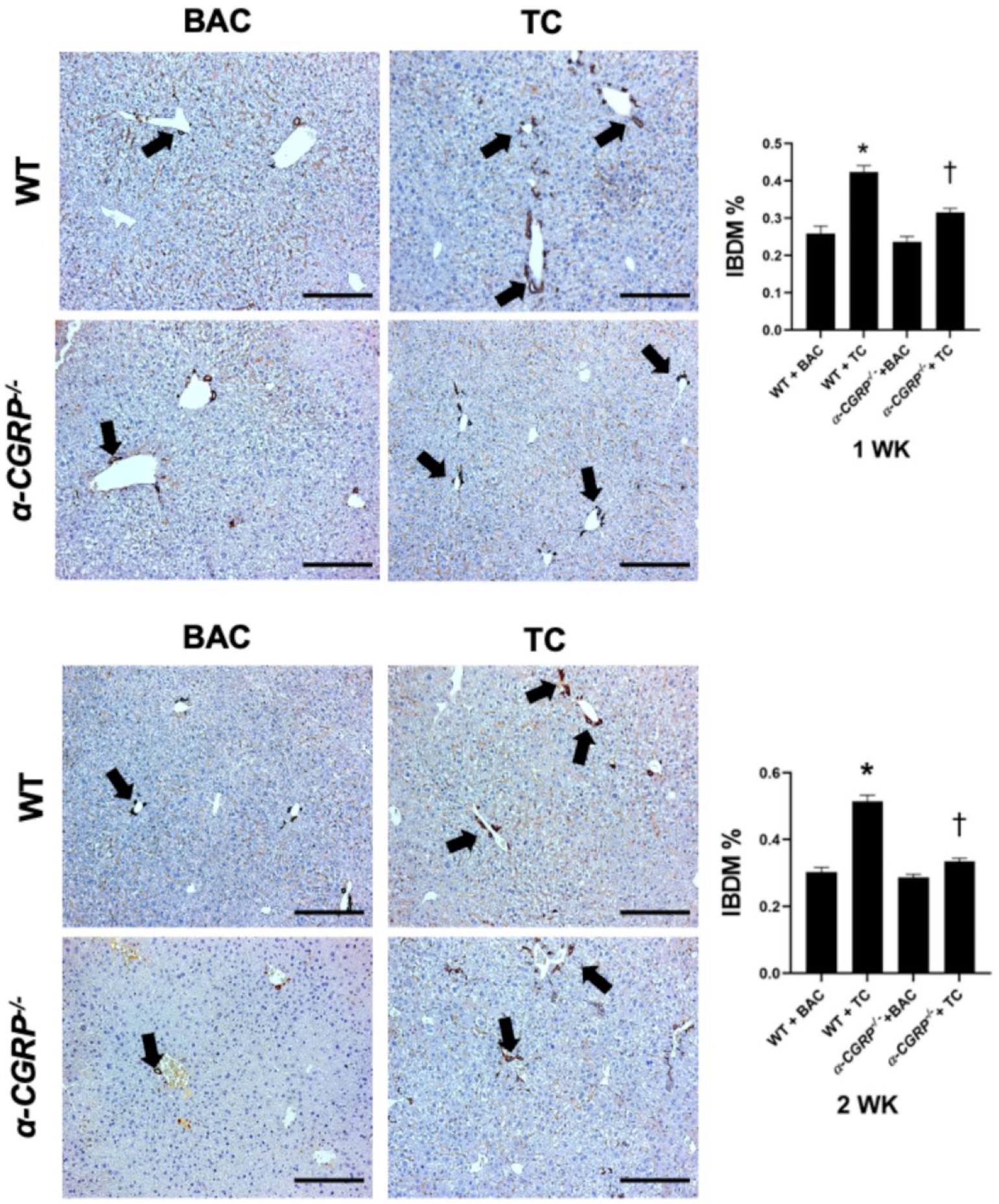
2.6. Evaluation of Intrahepatic Bile Duct Mass (IBDM)
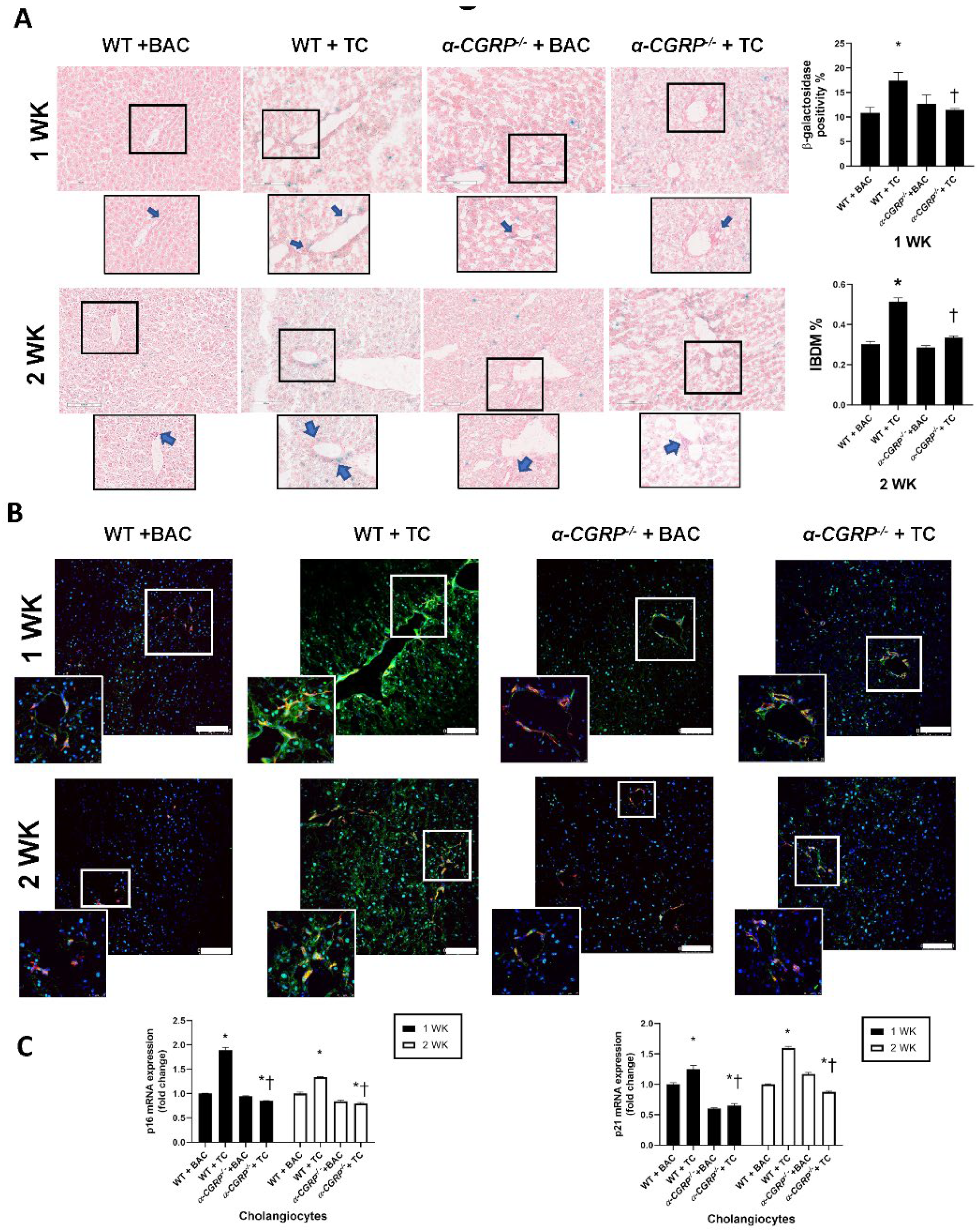
2.7. Evaluation of Biliary Senescence
2.8. Evaluation of Collagen Content
2.9. Measurement of cAMP/pERK/VEGF Signaling, Hepatic BA Composition, and Biliary Immunoreactivity/Expression of TGF-β1 and TGF-βRI
2.10. Statistical Analysis
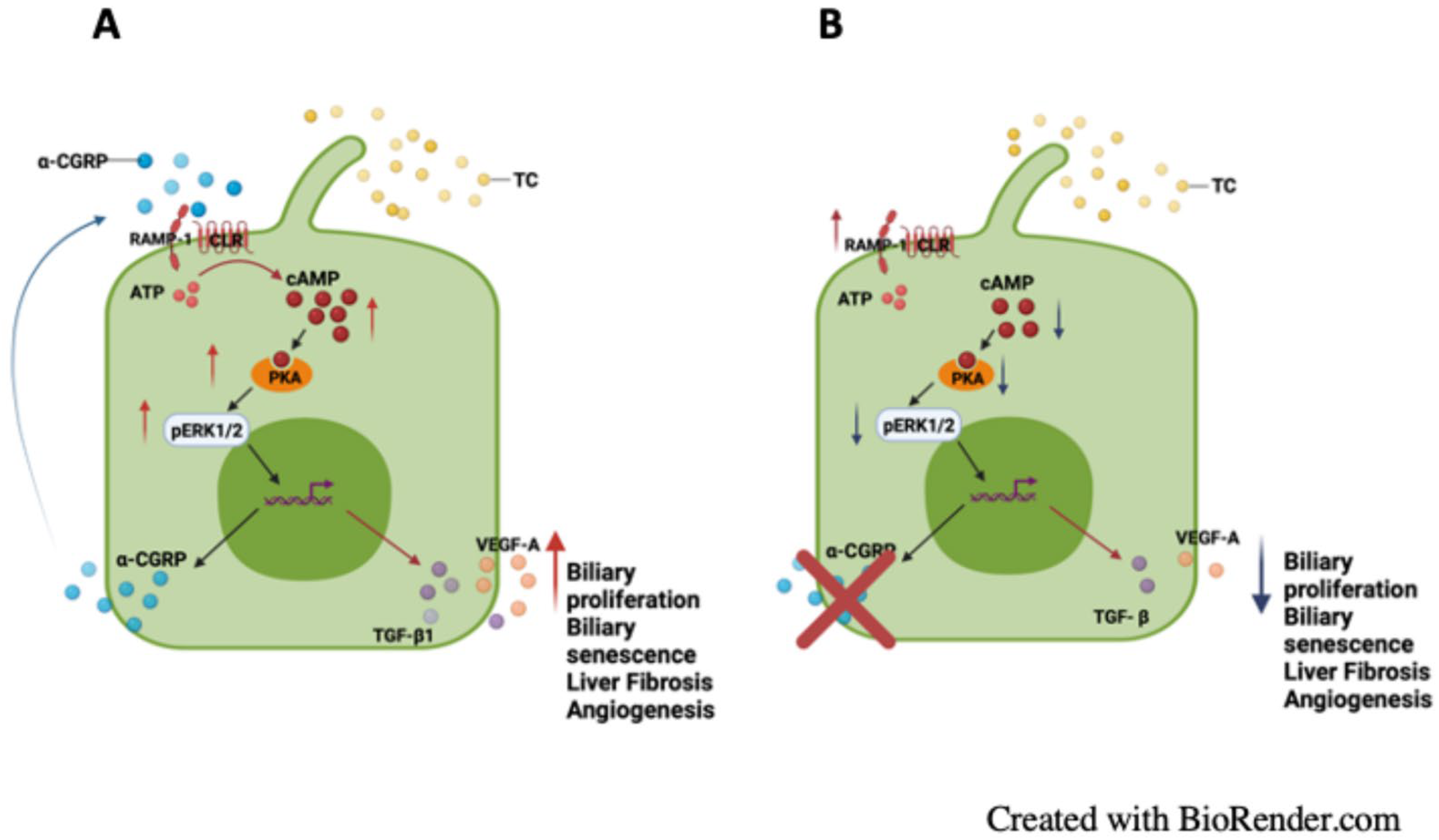
3. Results
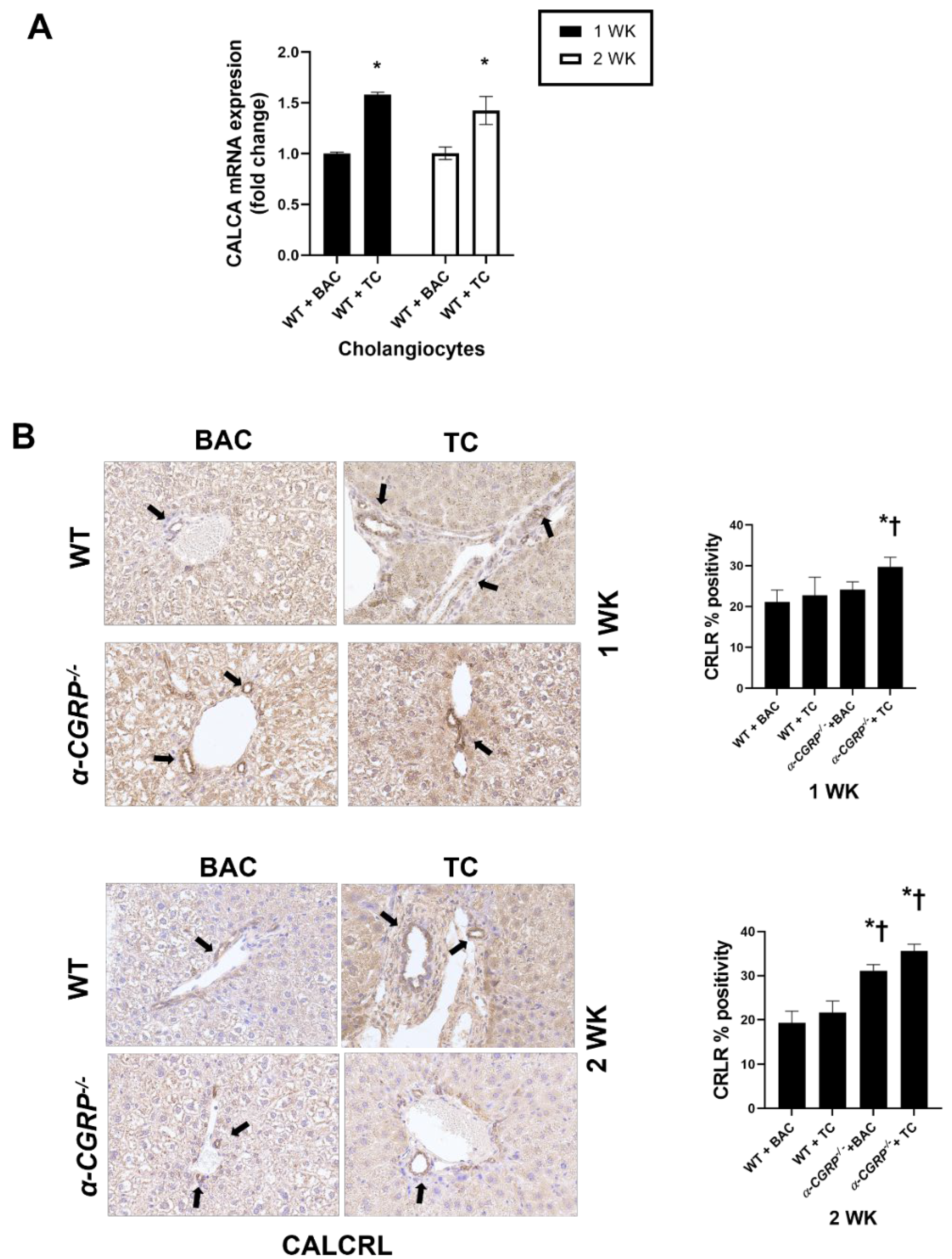
3.1. Measurement of CGRP Serum Levels and Immunoreactivity of CALCRL in Liver Sections
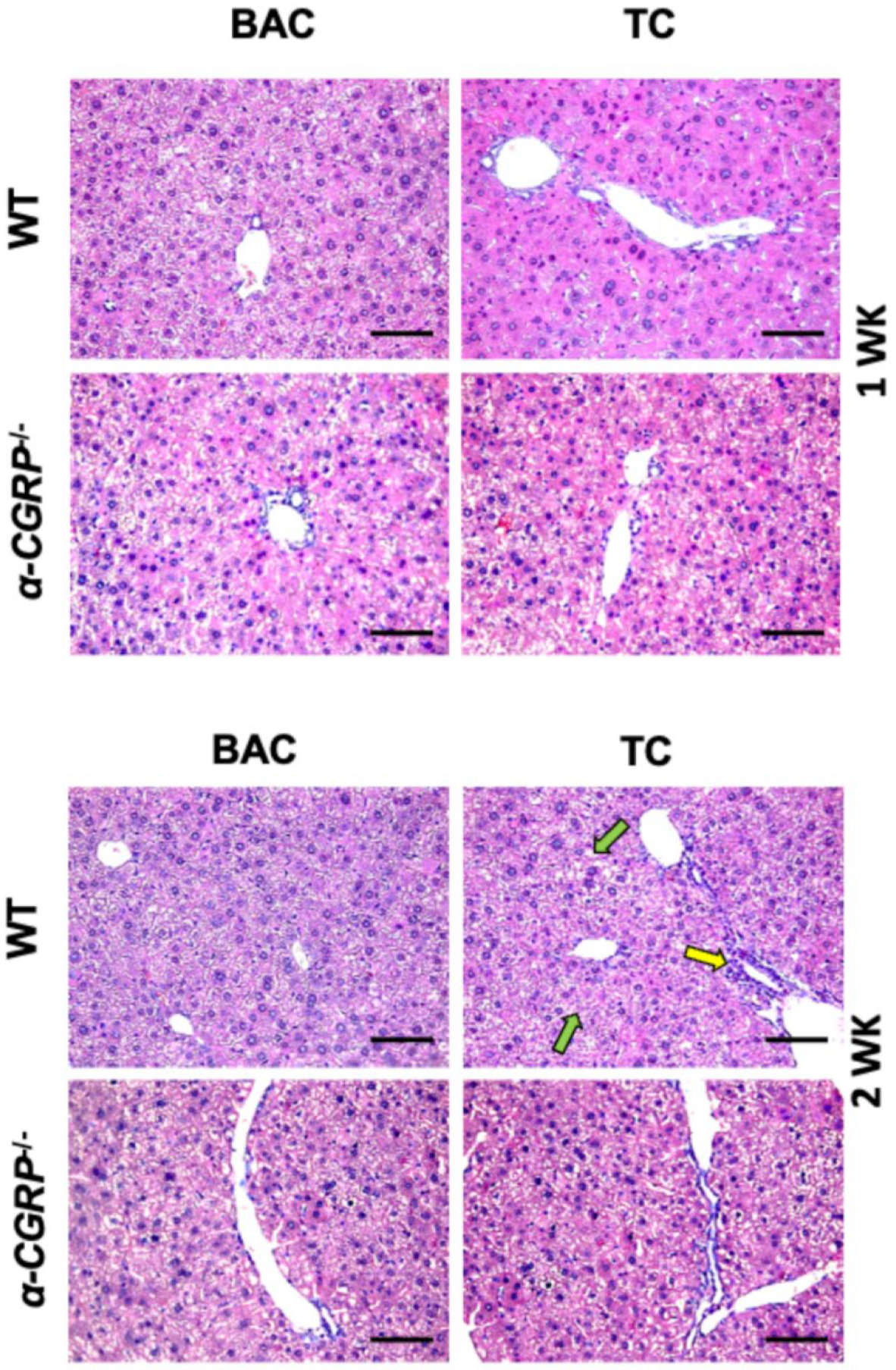
3.2. Evaluation of Liver Damage
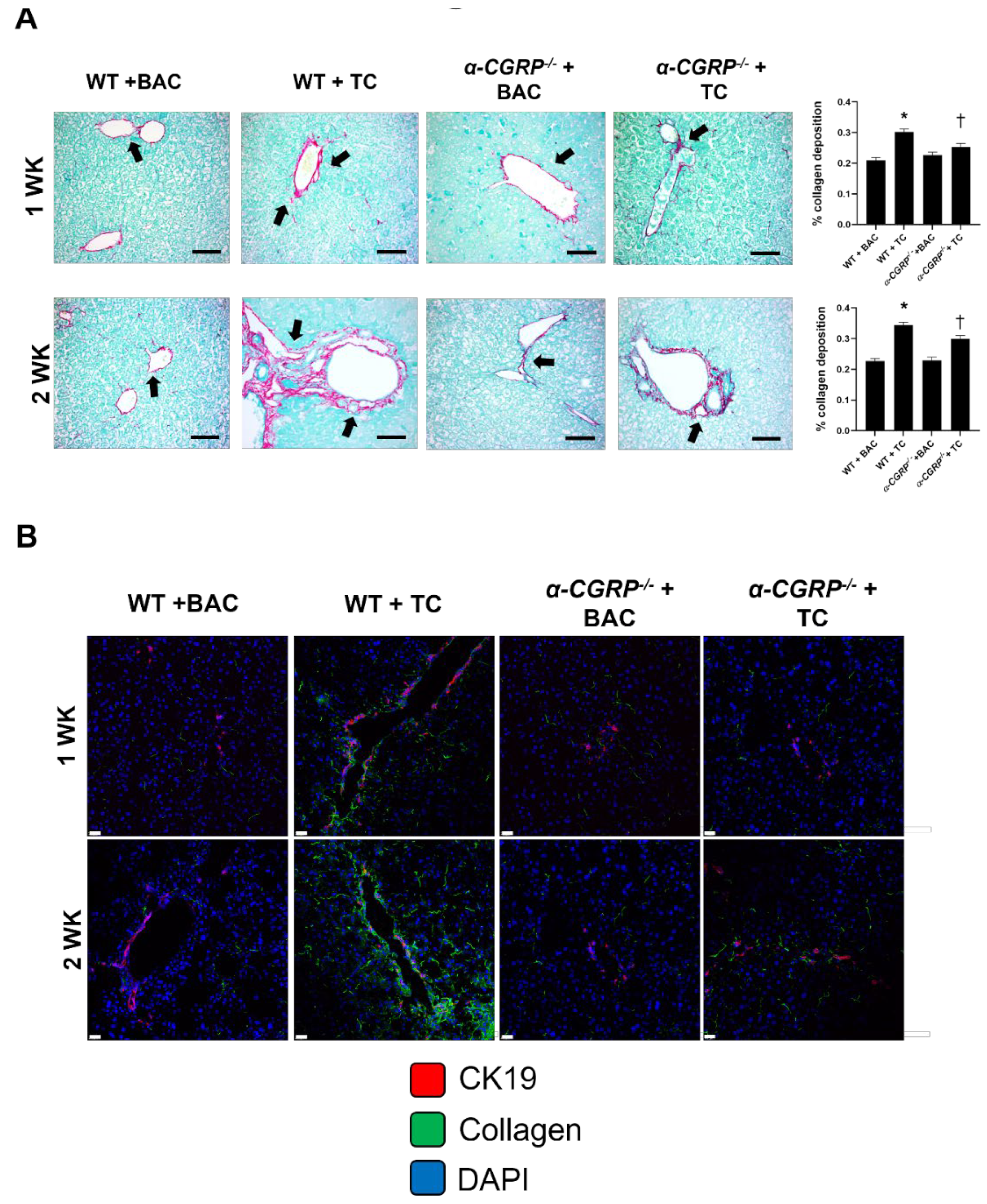
3.3. Evaluation of Collagen Deposition
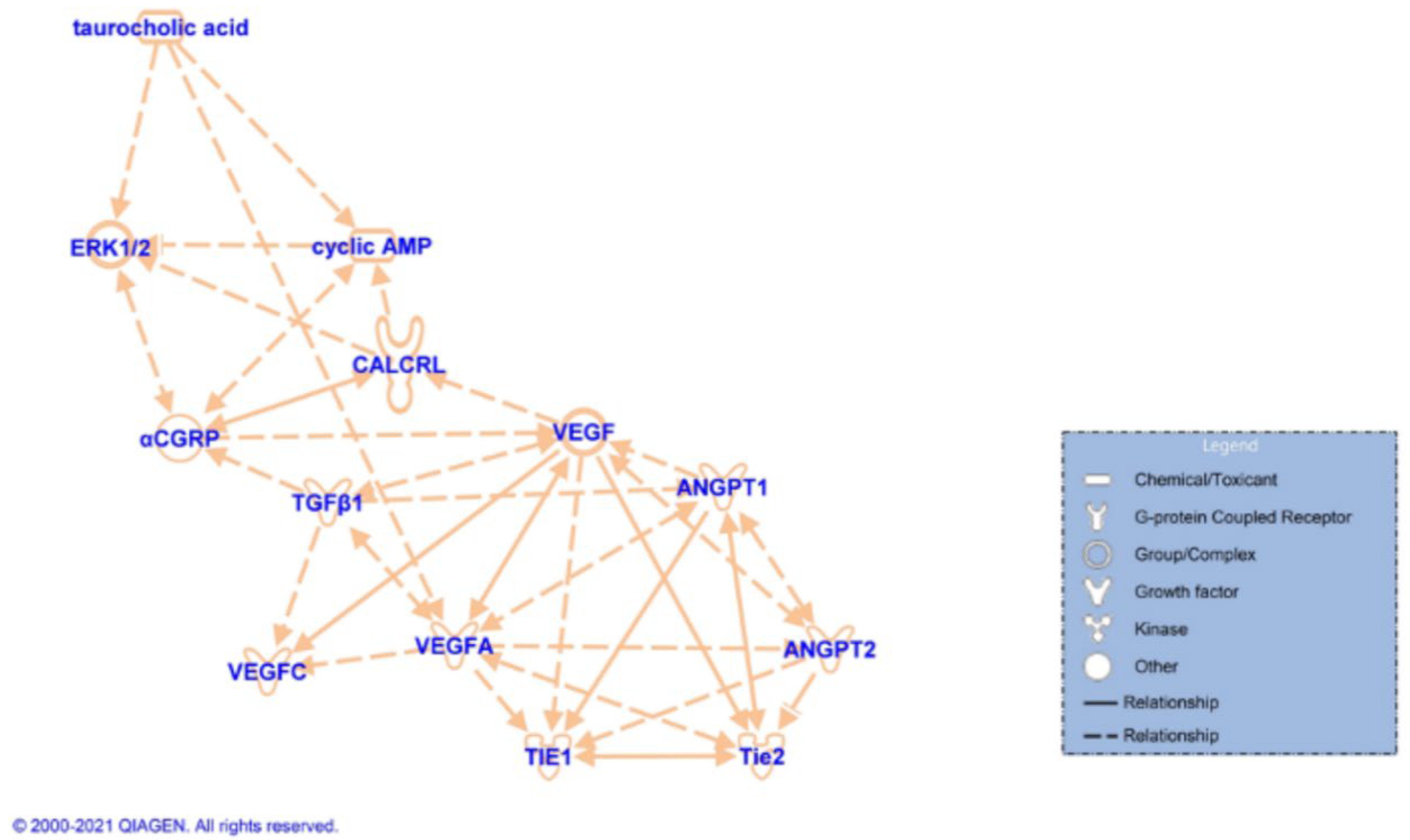
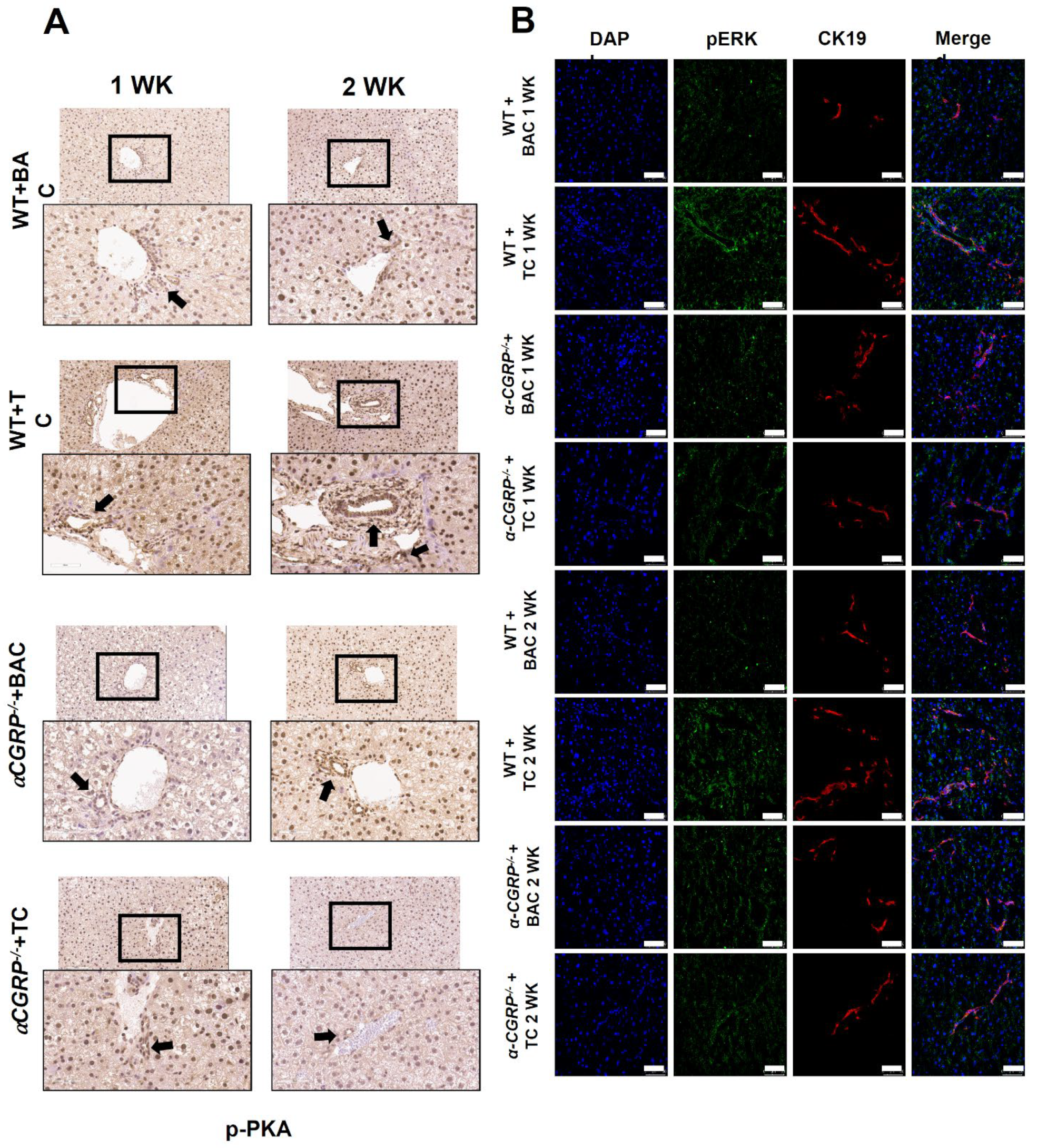
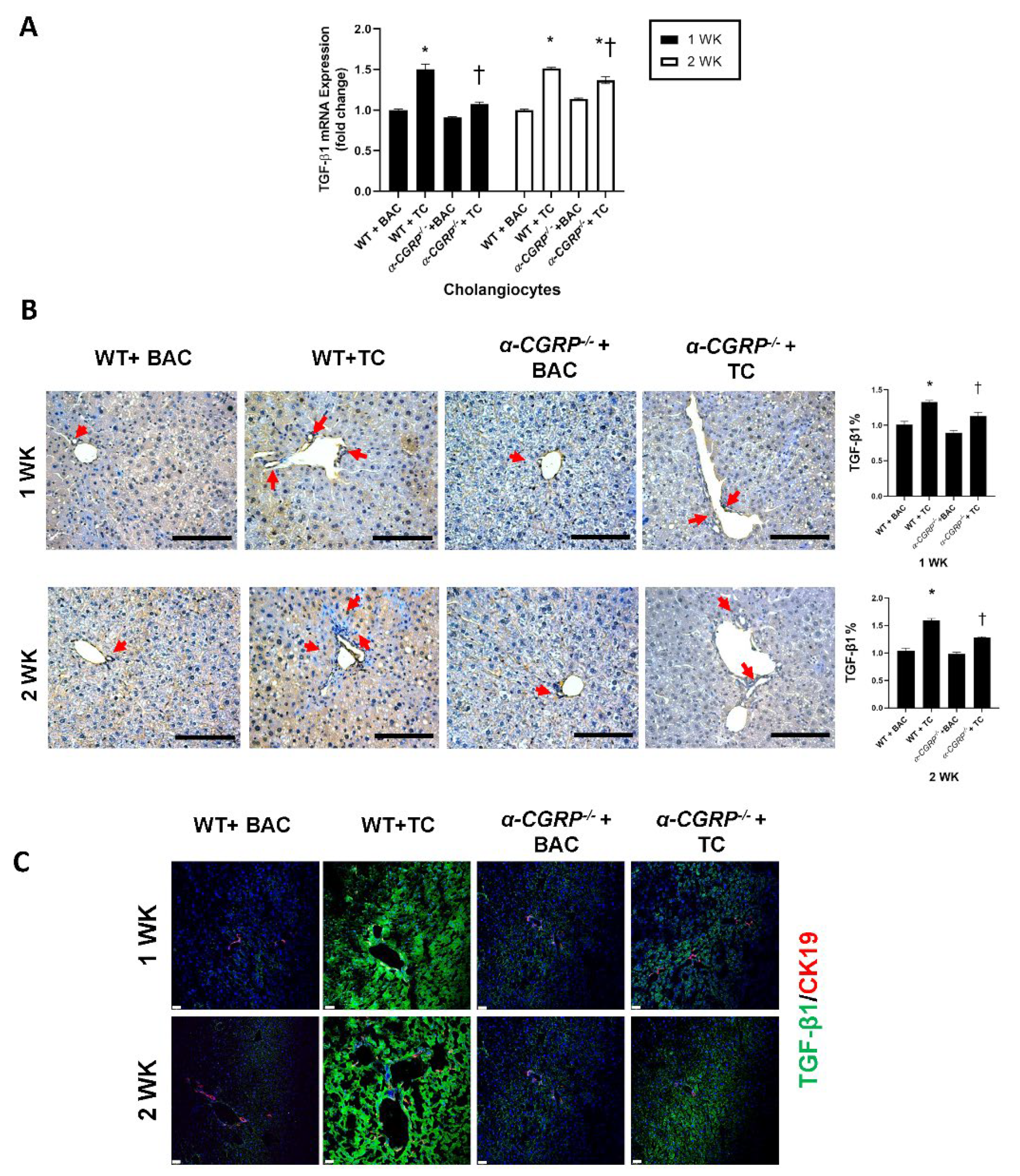
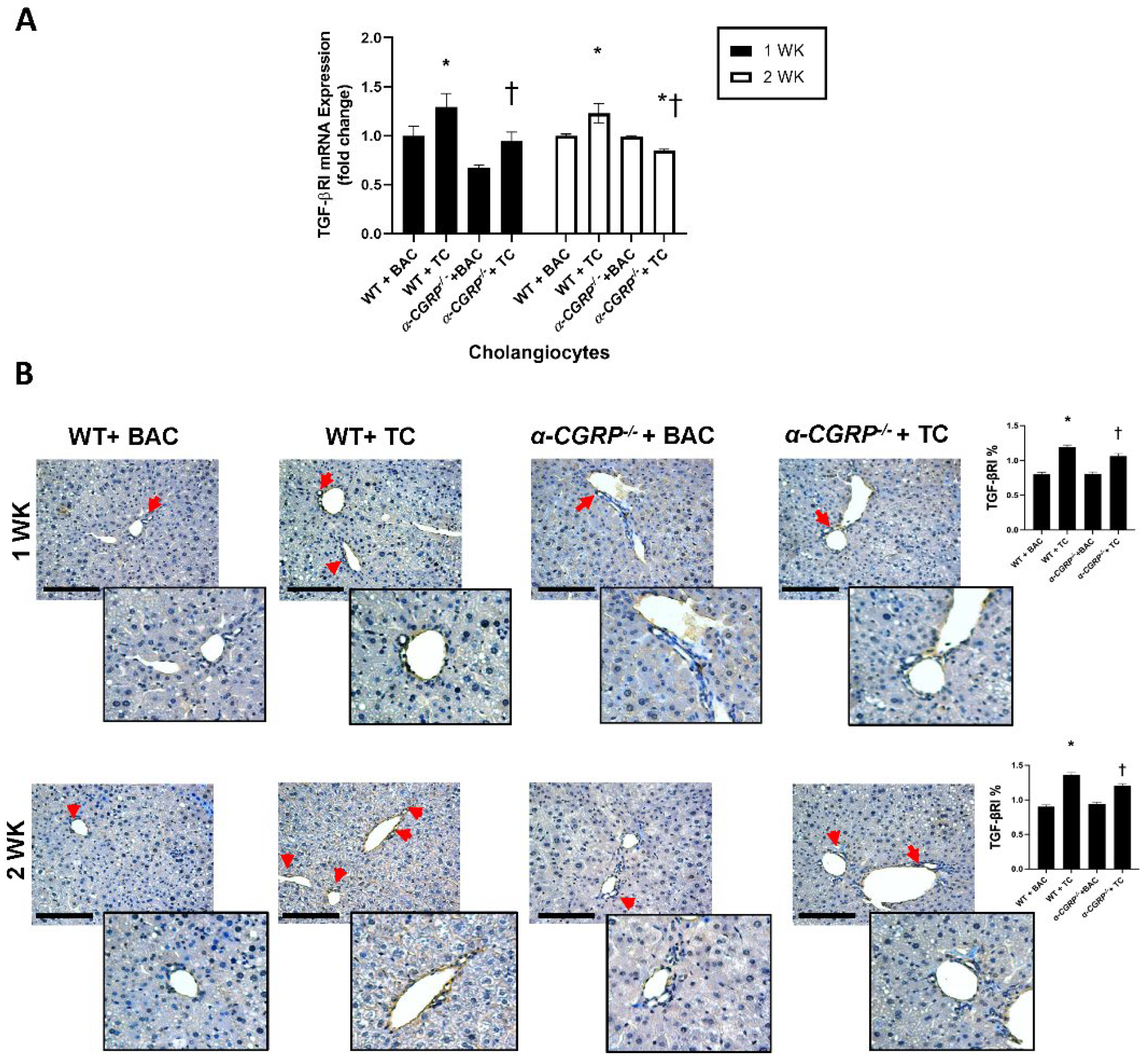
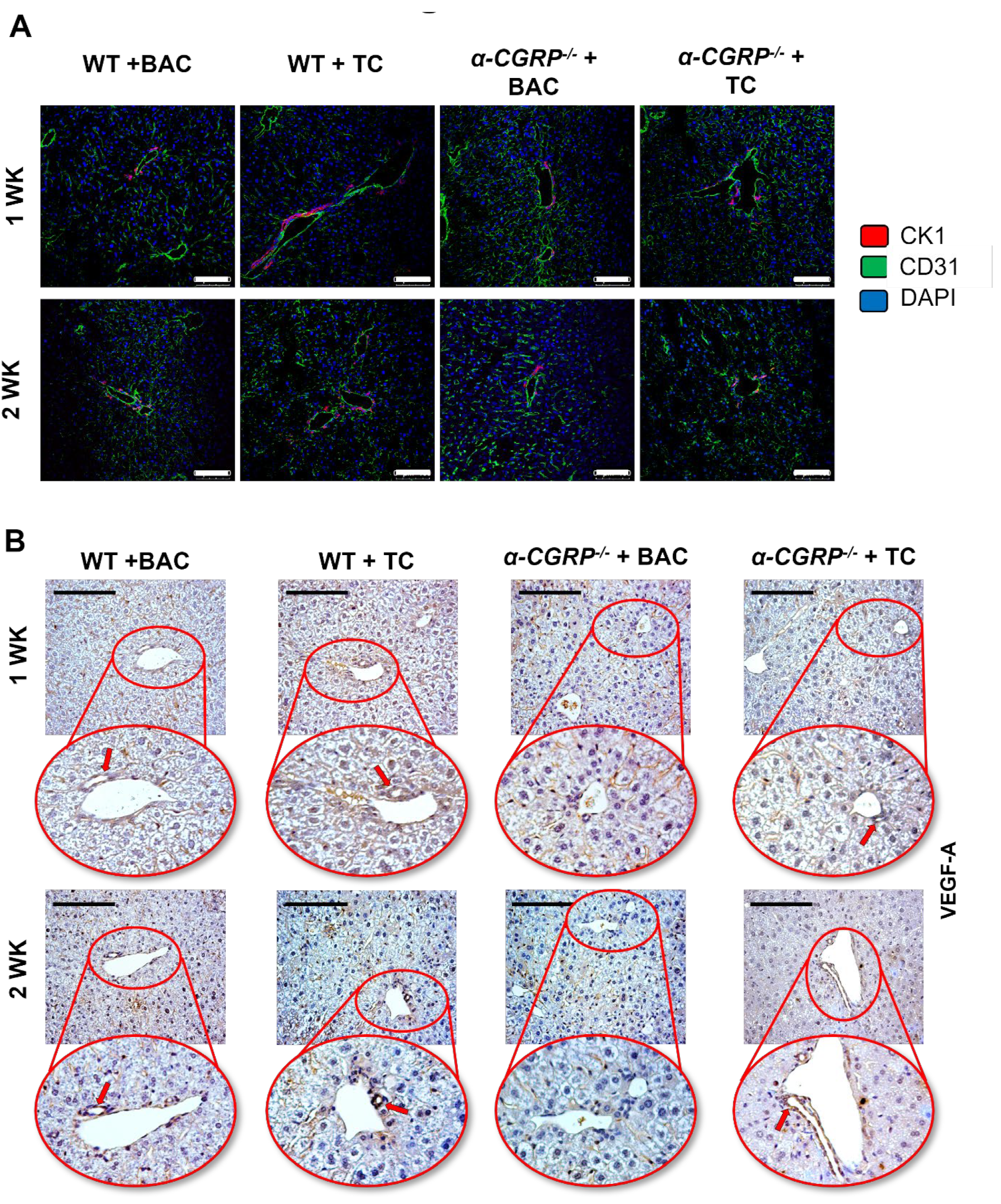
3.4. Measurement of Hepatic BA Composition, cAMP/pERK/VEGF Signaling, Hepatic BA Composition, and Biliary Immunoreactivity/Expression of TGF-β1, mdnTGF-βRI, and Angiogenic factors
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Kanno, N.; Lesage, G.; Glaser, S.; Alpini, G. Regulation of cholangiocyte bicarbonate secretion. Am. J. Physiol. Liver Physiol. 2001, 281, G612–G625. [Google Scholar] [CrossRef] [PubMed]
- Glaser, S.; Meng, F.; Han, Y.; Onori, P.; Chow, B.K.; Francis, H.; Venter, J.; McDaniel, K.; Marzioni, M.; Invernizzi, P.; et al. Secretin Stimulates Biliary Cell Proliferation by Regulating Expression of MicroRNA 125b and MicroRNA let7a in Mice. Gastroenterology 2014, 146, 1795.e12–1808.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Liu, R.; Yang, J.; Sun, L.; Zhang, L.; Jiang, Z.; Puri, P.; Gurley, E.C.; Lai, G.; Tang, Y.; et al. The Role of Long Noncoding Rna H19 in Gender Disparity of Cholestatic Liver Injury in Mul-tidrug Resistance 2 Gene Knockout Mice. Hepatology 2017, 66, 869–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Wu, N.; Meng, F.; Venter, J.; Giang, T.K.; Francis, H.; Kyritsi, K.; Wu, C.; Franchitto, A.; Alvaro, D.; et al. Knockout of Secretin Receptor Reduces Biliary Damage and Liver Fibrosis in Mdr2(−/−) Mice by Diminishing Senescence of Cholangiocytes. Lab. Invest. 2018, 98, 1449–1464. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Kennedy, L.; Hargrove, L.; Demieville, J.; Jones, H.; Madeka, T.; Karstens, A.; Chappell, K.; Alpini, G.; Sybenga, A.; et al. Ursodeoxycholate Inhibits Mast Cell Activation and Reverses Biliary Injury and Fibrosis in Mdr2(−/−) Mice and Human Primary Sclerosing Cholangitis. Lab. Invest. 2018, 98, 1465–1477. [Google Scholar] [CrossRef]
- Alpini, G.; Glaser, S.S.; Ueno, Y.; Rodgers, R.; Phinizy, J.L.; Francis, H.; Baiocchi, L.; Holcomb, L.A.; Caligiuri, A.; LeSage, G.D. Bile acid feeding induces cholangiocyte proliferation and secretion: Evidence for bile acid–regulated ductal secretion. Gastroenterology 1999, 116, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Alpini, G.; Ueno, Y.; Glaser, S.S.; Marzioni, M.; Phinizy, J.L.; Francis, H.; Lesage, G. Bile Acid Feeding Increased Pro-liferative Activity and Apical Bile Acid Transporter Expression in Both Small and Large Rat Cholangiocytes. Hepatology 2001, 34, 868–876. [Google Scholar] [CrossRef]
- Guicciardi, M.E.; Trussoni, C.E.; LaRusso, N.F.; Gores, G.J. The Spectrum of Reactive Cholangiocytes in Primary Sclerosing Cholangitis. Hepatology 2020, 71, 741–748. [Google Scholar] [CrossRef]
- Lleo, A.; Wang, G.Q.; Gershwin, M.E.; Hirschfield, G.M. Primary Biliary Cholangitis. Lancet 2020, 396, 1915–1926. [Google Scholar] [CrossRef]
- Kennedy, L.; Francis, H.; Invernizzi, P.; Venter, J.; Wu, N.; Carbone, M.; Gershwin, M.E.; Bernuzzi, F.; Franchitto, A.; Alvaro, D.; et al. Secretin/secretin receptor signaling mediates biliary damage and liver fibrosis in early-stage primary biliary cholangitis. FASEB J. 2019, 33, 10269–10279. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Marzioni, M.; Meng, F.; Francis, H.; Glaser, S.; Alpini, G. Ductular Reaction in Liver Diseases: Pathological Mechanisms and Translational Significances. Hepatology 2019, 69, 420–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, H.; Franchitto, A.; Ueno, Y.; Glaser, S.; DeMorrow, S.; Venter, J.; Gaudio, E.; Alvaro, D.; Fava, G.; Marzioni, M.; et al. H3 Histamine Receptor Agonist Inhibits Biliary Growth of Bdl Rats by Downregulation of the Camp-Dependent Pka/Erk1/2/Elk-1 Pathway. Lab. Invest. 2007, 87, 473–487. [Google Scholar] [CrossRef] [Green Version]
- Alpini, G.; Glaser, S.; Alvaro, D.; Ueno, Y.; Marzioni, M.; Francis, H.; Baiocchi, L.; Stati, T.; Barbaro, B.; Phinizy, J.L.; et al. Bile acid depletion and repletion regulate cholangiocyte growth and secretion by a phosphatidylinositol 3-kinase–dependent pathway in rats. Gastroenterology 2002, 123, 1226–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glaser, S.; Onori, P.; Gaudio, E.; Ueno, Y.; Pannarale, L.; Franchitto, A.; Francis, H.; Mancinelli, R.; Carpino, G.; Venter, J.; et al. Taurocholic acid prevents biliary damage induced by hepatic artery ligation in cholestatic rats. Dig. Liver Dis. 2010, 42, 709–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abenavoli, L.; Procopio, A.C.; Fagoonee, S.; Pellicano, R.; Carbone, M.; Luzza, F.; Invernizzi, P. Primary Biliary Cholangitis and Bile Acid Farnesoid X Receptor Agonists. Diseases 2020, 8, 20. [Google Scholar] [CrossRef]
- Etherington, R.E.; Millar, B.J.M.; Innes, B.A.; Jones, D.E.J.; Kirby, J.A.; Brain, J.G. Bile acid receptor agonists in primary biliary cholangitis: Regulation of the cholangiocyte secretome and downstream T cell differentiation. FASEB BioAdvances 2019, 1, 332–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glaser, S.S.; Ueno, Y.; DeMorrow, S.; Chiasson, V.L.; Katki, K.A.; Venter, J.; Francis, H.L.; Dickerson, I.M.; DiPette, D.J.; Supowit, S.C.; et al. Knockout of Alpha-Calcitonin Gene-Related Peptide Reduces Cholangiocyte Proliferation in Bile Duct Ligated Mice. Lab. Invest. 2007, 87, 914–926. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, M.G.; Mermod, J.-J.; Amara, S.; Swanson, L.W.; Sawchenko, P.E.; Rivier, J.; Vale, W.W.; Evans, R. Production of a novel neuropeptide encoded by the calcitonin gene via tissue-specific RNA processing. Nature 1983, 304, 129–135. [Google Scholar] [CrossRef]
- Laschinger, M.; Wang, Y.; Holzmann, G.; Wang, B.; Stöß, C.; Lu, M.; Brugger, M.; Schneider, A.; Knolle, P.; Wohlleber, D.; et al. The CGRP receptor component RAMP1 links sensory innervation with YAP activity in the regenerating liver. FASEB J. 2020, 34, 8125–8138. [Google Scholar] [CrossRef] [Green Version]
- Tiegs, G.; Bang, R.; Neuhuber, W.L. Requirement of Peptidergic Sensory Innervation for Disease Activity in Murine Models of Immune Hepatitis and Protection by Beta-Adrenergic Stimulation. J. Neuroimmunol. 1999, 96, 131–143. [Google Scholar] [CrossRef]
- Poyner, D.R. Calcitonin Gene-Related Peptide: Multiple Actions, Multiple Receptors. Pharmacol. Ther. 1992, 56, 23–51. [Google Scholar] [CrossRef]
- Conner, A.C.; Simms, J.; Howitt, S.G.; Wheatley, M.; Poyner, D.R. The Second Intracellular Loop of the Calcitonin Gene-related Peptide Receptor Provides Molecular Determinants for Signal Transduction and Cell Surface Expression. J. Biol. Chem. 2006, 281, 1644–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Y.; Ceci, L.; Wu, N.; Zhou, T.; Chen, L.; Venter, J.; Francis, H.; Bernuzzi, F.; Invernizzi, P.; Kyritsi, K.; et al. Knockout of Alpha-Calcitonin Gene-Related Peptide Attenuates Cholestatic Liver Injury by Differentially Regulating Cellular Senescence of Hepatic Stellate Cells and Cholangiocytes. Lab. Invest. 2019, 99, 764–776. [Google Scholar] [CrossRef]
- Francis, H.; Glaser, S.; Ueno, Y.; Lesage, G.; Marucci, L.; Benedetti, A.; Taffetani, S.; Marzioni, M.; Alvaro, D.; Venter, J.; et al. Camp Stimulates the Secretory and Proliferative Capacity of the Rat Intrahepatic Biliary Epithelium through Changes in the Pka/Src/Mek/Erk1/2 Pathway. J. Hepatol. 2004, 41, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hoff, A.O.; Wimalawansa, S.J.; Cote, G.J.; Gagel, R.F.; Westlund, K.N. Arthritic Calcitonin/Alpha Cal-citonin Gene-Related Peptide Knockout Mice Have Reduced Nociceptive Hypersensitivity. Pain 2001, 89, 265–273. [Google Scholar] [CrossRef]
- Glaser, S.; Gaudio, E.; Renzi, A.; Mancinelli, R.; Ueno, Y.; Venter, J.; White, M.; Kopriva, S.; Chiasson, V.; DeMorrow, S.; et al. Knockout of the neurokinin-1 receptor reduces cholangiocyte proliferation in bile duct-ligated mice. Am. J. Physiol. Liver Physiol. 2011, 301, G297–G305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishii, M.; Vroman, B.; LaRusso, N.F. Isolation and morphologic characterization of bile duct epithelial cells from normal rat liver. Gastroenterology 1989, 97, 1236–1247. [Google Scholar] [CrossRef]
- Permpoonputtana, K.; Porter, J.E.; Govitrapong, P. Calcitonin Gene-Related Peptide Mediates an Inflammatory Response in Schwann Cells Via Camp-Dependent Erk Signaling Cascade. Life Sci. 2016, 144, 19–25. [Google Scholar] [CrossRef]
- Zheng, S.; Li, W.; Xu, M.; Bai, X.; Zhou, Z.; Han, J.; Shyy, J.Y.-J.; Wang, X. Calcitonin gene-related peptide promotes angiogenesis via AMP-activated protein kinase. Am. J. Physiol. Physiol. 2010, 299, C1485–C1492. [Google Scholar] [CrossRef] [Green Version]
- Kakiyama, G.; Muto, A.; Takei, H.; Nittono, H.; Murai, T.; Kurosawa, T.; Hofmann, A.F.; Pandak, W.M.; Bajaj, J.S. A simple and accurate HPLC method for fecal bile acid profile in healthy and cirrhotic subjects: Validation by GC-MS and LC-MS. J. Lipid Res. 2014, 55, 978–990. [Google Scholar] [CrossRef] [Green Version]
- Basilio-De-Oliveira, R.P.; Pannain, V.L.N. Prognostic angiogenic markers (endoglin, VEGF, CD31) and tumor cell proliferation (Ki67) for gastrointestinal stromal tumors. World J. Gastroenterol. 2015, 21, 6924–6930. [Google Scholar] [CrossRef] [PubMed]
- DiPette, J.D.; Supowit, S.C. Calcitonin Gene-Related Peptide and Adrenomedullin. In Primer on the Autonomic Nervous System, 3rd ed; Academic Press: Cambridge, MA, USA, 2012; pp. 127–129. [Google Scholar]
- Alpini, G.; Lenzi, R.; Sarkozi, L.; Tavoloni, N. Biliary Physiology in Rats with Bile Ductular Cell Hyperplasia. Evidence for a Secretory Function of Proliferated Bile Ductules. J. Clin. Invest. 1988, 81, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, Y.; Francis, H.; Glaser, S.; DeMorrow, S.; Venter, J.; Benedetti, A.; Fava, G.; Marzioni, M.; Alpini, G. Taurocholic acid feeding prevents tumor necrosis factor-alpha-induced damage of cholangiocytes by a PI3K-mediated pathway. Exp. Biol. Med. 2007, 232, 942–949. [Google Scholar]
- Alpini, G.; Glaser, S.; Robertson, W.; Phinizy, J.L.; Rodgers, R.E.; Caligiuri, A.; LeSage, G. Bile Acids Stimulate Prolif-erative and Secretory Events in Large but Not Small Cholangiocytes. Am. J. Physiol. 1997, 273, G518–G529. [Google Scholar] [PubMed]
- Bijvelds, J.M.; Jorna, H.; Verkade, H.J.; Bot, A.G.; Hofmann, F.; Agellon, L.B.; Sinaasappel, M.; de Jonge, H.R. Activation of Cftr by Asbt-Mediated Bile Salt Absorption. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G870–G879. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wu, N.; Kennedy, L.; Francis, H.; Ceci, L.; Zhou, T.; Samala, N.; Kyritsi, K.; Wu, C.; Sybenga, A.; et al. Inhibition of Secretin/Secretin Receptor Axis Ameliorates Non-Alcoholic Fatty Liver Disease Phenotypes. Hepatology 2021, 74, 1845–1863. [Google Scholar] [CrossRef]
- Yin, P.; Wan, D.; Zhao, C.; Chen, J.; Zhao, X.; Wang, W.; Lu, X.; Yang, S.; Gu, J.; Xu, G. A metabonomic study of hepatitis B-induced liver cirrhosis and hepatocellular carcinoma by using RP-LC and HILIC coupled with mass spectrometry. Mol. BioSyst. 2009, 5, 868–876. [Google Scholar] [CrossRef]
- Wang, X.; Xie, G.; Zhao, A.; Zheng, X.; Huang, F.; Wang, Y.; Yao, C.; Jia, W.; Liu, P. Serum Bile Acids Are Associated with Pathological Progression of Hepatitis B-Induced Cirrhosis. J. Proteome Res. 2016, 15, 1126–1134. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Nakanuma, Y. Bile Acids and Deregulated Cholangiocyte Autophagy in Primary Biliary Cholangitis. Dig. Dis. 2017, 35, 210–216. [Google Scholar] [CrossRef]
- Sugata, K.; Kamimura, S.; Shijo, H. Differential effects between tauroursodeoxycholic and taurochenodeoxycholic acids in hepatic fibrosis: An assessment by primary cultured Ito and Kupffer cells from the rat liver. J. Gastroenterol. Hepatol. 1996, 11, 451–458. [Google Scholar] [CrossRef]
- Cho, G.J.; Lee, J.H.; Hong, S.H.; Lee, H.N.; Kim, C.M.; Kim, S.Y.; Yoon, K.J.; Oh, B.J.; Kim, J.H.; Jung, S.Y.; et al. Tauroursodeoxycholic Acid, a Bile Acid, Promotes Blood Vessel Repair by Recruiting Vascu-logenic Progenitor Cells. Stem Cells 2015, 33, 792–805. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.T.; Kanno, K.; Pham, Q.T.; Kikuchi, Y.; Kakimoto, M.; Kobayashi, T.; Otani, Y.; Kishikawa, N.; Miyauchi, M.; Arihiro, K.; et al. Senescent hepatic stellate cells caused by deoxycholic acid modulates malignant behavior of hepatocellular carcinoma. J. Cancer Res. Clin. Oncol. 2020, 146, 3255–3268. [Google Scholar] [CrossRef] [PubMed]
- Ye, H.-L.; Zhang, J.-W.; Chen, X.-Z.; Wu, P.-B.; Chen, L.; Zhang, G. Ursodeoxycholic acid alleviates experimental liver fibrosis involving inhibition of autophagy. Life Sci. 2020, 242, 117175. [Google Scholar] [CrossRef]
- Thakare, R.; AlAmoudi, J.A.; Gautam, N.; Rodrigues, A.D.; Alnouti, Y. Species differences in bile acids I. Plasma and urine bile acid composition. J. Appl. Toxicol. 2018, 38, 1323–1335. [Google Scholar] [CrossRef]
- García-Cañaveras, J.C.; Donato, M.T.; Castell, J.V.; Lahoz, A. Targeted Profiling of Circulating and Hepatic Bile Acids in Human, Mouse, and Rat Using a Uplc-Mrm-Ms-Validated Method. J. Lipid Res. 2012, 53, 2231–2241. [Google Scholar] [CrossRef] [Green Version]
- Li, W.-Q.; Tan, S.-L.; Li, X.-H.; Sun, T.-L.; Li, D.; Du, J.; Wei, S.-S.; Li, Y.-J.; Zhang, B.-K. Calcitonin gene-related peptide inhibits the cardiac fibroblasts senescence in cardiac fibrosis via up-regulating klotho expression. Eur. J. Pharmacol. 2018, 843, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Gaudio, E.; Barbaro, B.; Alvaro, D.; Glaser, S.; Francis, H.; Ueno, Y.; Meininger, C.; Franchitto, A.; Onori, P.; Marzioni, M.; et al. Vascular Endothelial Growth Factor Stimulates Rat Cholangiocyte Proliferation Via an Autocrine Mechanism. Gastroenterology 2006, 130, 1270–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Mourik, J.; Leeksma, O.C.; Reinders, J.H.; De Groot, P.G.; Zandbergen-Spaargaren, J. Vascular endothelial cells synthesize a plasma membrane protein indistinguishable from the platelet membrane glycoprotein IIa. J. Biol. Chem. 1985, 260, 11300–11306. [Google Scholar] [CrossRef]
- Giltay, J.C.; Brinkman, H.J.; Modderman, P.W.; Borne, A.E.V.D.; A Van Mourik, J. Human vascular endothelial cells express a membrane protein complex immunochemically indistinguishable from the platelet VLA-2 (glycoprotein Ia-IIa) complex. Blood 1989, 73, 1235–1241. [Google Scholar] [CrossRef]
- Gaudio, E.; Onori, P.; Pannarale, L.; Alvaro, D. Hepatic microcirculation and peribiliary plexus in experimental biliary cirrhosis: A morphological study. Gastroenterology 1996, 111, 1118–1124. [Google Scholar] [CrossRef]
- Gaudio, E.; Onori, P.; Franchitto, A.; Pannarale, L.; Alpini, G.; Alvaro, D. Hepatic microcirculation and cholangiocyte physiopathology. Ital. J. Anat. Embryol. 2005, 110, 71–75. [Google Scholar] [PubMed]
- Mancinelli, R.; Onori, P.; Gaudio, E.; Franchitto, A.; Carpino, G.; Ueno, Y.; Alvaro, D.; Annarale, L.P.; DeMorrow, S.; Francis, H. Taurocholate Feeding to Bile Duct Ligated Rats Prevents Caffeic Acid-Induced Bile Duct Damage by Changes in Cholangiocyte VEGF Expression. Exp. Biol. Med. 2009, 234, 462–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishima, T.; Ito, Y.; Hosono, K.; Tamura, Y.; Uchida, Y.; Hirata, M.; Suzsuki, T.; Amano, H.; Kato, S.; Kurihara, Y.; et al. Calcitonin gene-related peptide facilitates revascularization during hindlimb ischemia in mice. Am. J. Physiol. Circ. Physiol. 2011, 300, H431–H439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, S.P.; Kim, J. Exogenous CGRP upregulates profibrogenic growth factors through PKC/JNK signaling pathway in kidney proximal tubular cells. Cell Biol. Toxicol. 2018, 34, 251–262. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | WT+ BAC 1 wk | WT+ TC 1 wk | α-CGRP−/± BAC 1 wk | α-CGRP−/± TC 1 wk | WT+ BAC 2 wk | WT+ TC 2 wk | α-CGRP−/± BAC 2 wk | α-CGRP−/± TC 2 wk |
| Liver weight (g) | 1.4 ± 0.1 n = 8 | 1.5 ± 0.1 n = 17 | 1.6 ± 0.1 n = 9 | 1.4 ± 0.1 n = 10 | 1.3 ± 0.1 n = 10 | 1.6 ± 0.1 n = 11 | 1.4 ± 0.01 n = 7 | 1.3 ± 0.1 n = 14 |
| Body weight (g) | 27.7 ± 1.6 n = 8 | 24.8 ± 0.5 n = 17 | 25.4 ± 1.4 n = 9 | 22 ± 1.1 n = 10 | 27.6 ± 1.0 n = 10 | 25.0 ± 0.9 n = 11 | 26.8 ± 1.1 n = 7 | 23.4 ± 1.1 n = 14 |
| LW/BW (%) | 5.1 ± 0.3 n = 8 | 6.2 ± 0.2 n = 17 | 6.50 ± 0.7 n = 9 | 5.9 ± 0.3 n = 10 | 4.7 ± 0.3 n = 10 | 6.7 ± 0.6 * n = 11 | 5.3 ± 0.3 n = 7 | 5.7 ± 0.2 n = 14 |
| Inflammatory infiltrated area | 0.0 ± 0.0 n = 8 | 0.41 ± 0.06 n = 8 | 0.24 ± 0.05 n = 8 | 0.34 ± 0.05 n = 8 | 0.0 ± 0.0 n = 8 | 1.9 ± 0.16 * n = 8 | 0.25 ± 0.06 n = 8 | 0.62 ± 0.07 n = 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mancinelli, R.; Ceci, L.; Kennedy, L.; Francis, H.; Meadows, V.; Chen, L.; Carpino, G.; Kyritsi, K.; Wu, N.; Zhou, T.; et al. The Effects of Taurocholic Acid on Biliary Damage and Liver Fibrosis Are Mediated by Calcitonin-Gene-Related Peptide Signaling. Cells 2022, 11, 1591. https://doi.org/10.3390/cells11091591
Mancinelli R, Ceci L, Kennedy L, Francis H, Meadows V, Chen L, Carpino G, Kyritsi K, Wu N, Zhou T, et al. The Effects of Taurocholic Acid on Biliary Damage and Liver Fibrosis Are Mediated by Calcitonin-Gene-Related Peptide Signaling. Cells. 2022; 11(9):1591. https://doi.org/10.3390/cells11091591
Chicago/Turabian StyleMancinelli, Romina, Ludovica Ceci, Lindsey Kennedy, Heather Francis, Vik Meadows, Lixian Chen, Guido Carpino, Konstantina Kyritsi, Nan Wu, Tianhao Zhou, and et al. 2022. "The Effects of Taurocholic Acid on Biliary Damage and Liver Fibrosis Are Mediated by Calcitonin-Gene-Related Peptide Signaling" Cells 11, no. 9: 1591. https://doi.org/10.3390/cells11091591
APA StyleMancinelli, R., Ceci, L., Kennedy, L., Francis, H., Meadows, V., Chen, L., Carpino, G., Kyritsi, K., Wu, N., Zhou, T., Sato, K., Pannarale, L., Glaser, S., Chakraborty, S., Alpini, G., Gaudio, E., Onori, P., & Franchitto, A. (2022). The Effects of Taurocholic Acid on Biliary Damage and Liver Fibrosis Are Mediated by Calcitonin-Gene-Related Peptide Signaling. Cells, 11(9), 1591. https://doi.org/10.3390/cells11091591