
Current Strategies for Exosome Cargo Loading and Targeting Delivery



Abstract
:1. Introduction
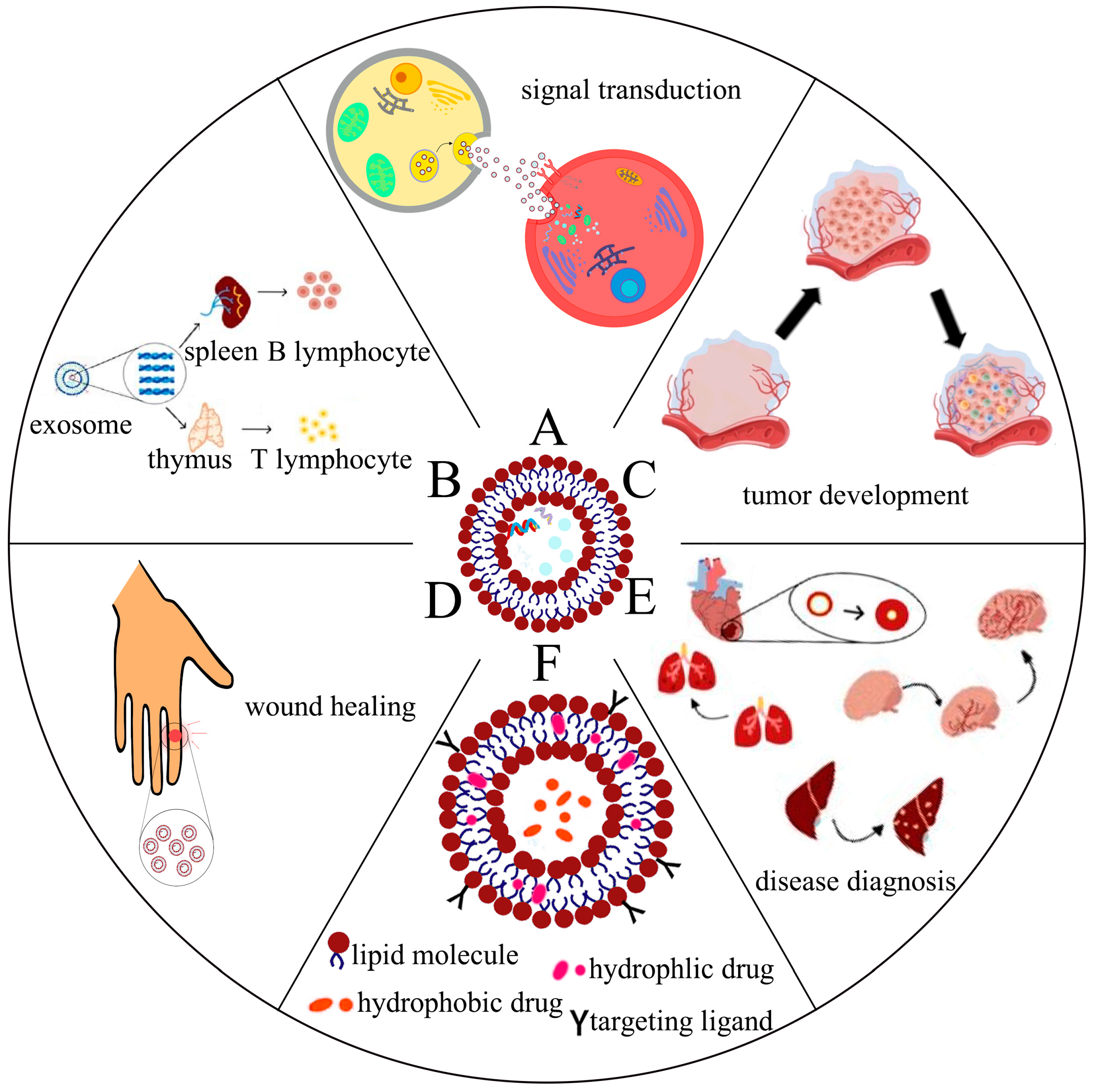
2. Biological Functions of Exosomes
2.1. Exosomes Are Involved in Intercellular Signaling
2.2. Exosomes Are Involved in Antigen Presentation and Regulation of Immune Responses
2.3. Exosomes Are Involved in Tumor Development
2.4. The Role of Exosomes for Wound Healing
2.5. The Role of Exosomes in Disease Diagnosis
2.6. The Role of Exosomes in Drug Delivery
3. Advantages of Exosomes as Drug Carriers
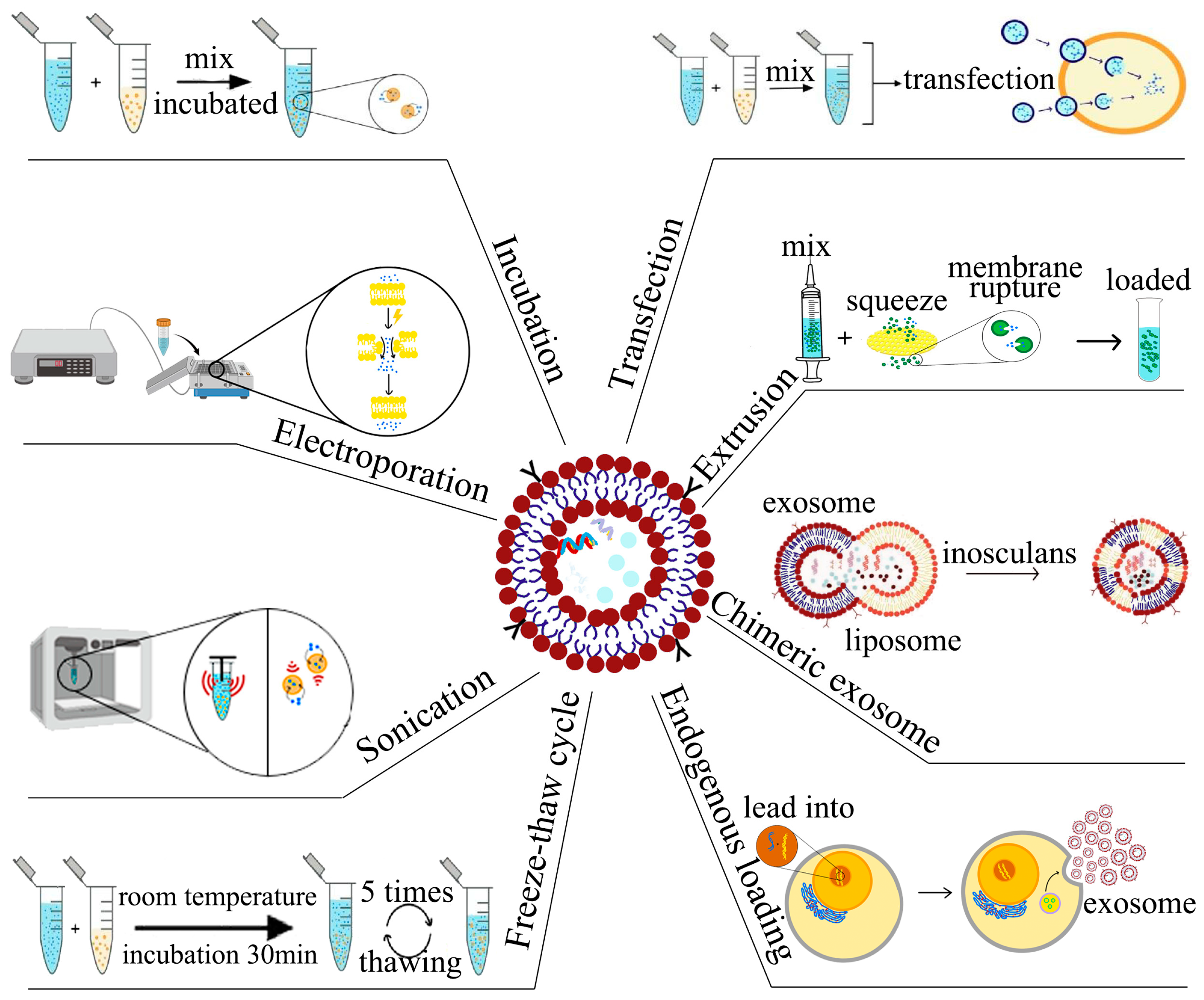
4. Strategies for Loading of Small-Molecule Drugs
4.1. Incubation for Loading of Small-Molecule Drugs
4.2. Electroporation for Loading of Small-Molecule Drugs
4.3. Sonication for Loading of Small-Molecule Drugs
4.4. Extrusion for Loading of Small-Molecule Drugs
4.5. Chimeric Exosome Method for Loading of Small-Molecule Drugs
4.6. Endogenous Loading for Small-Molecule Drugs
5. Loading Strategies for Nucleic Acid Drugs
5.1. Incubation for Loading of Nucleic Acid Drugs
5.2. Electroporation for Loading of Nucleic Acid Drugs
5.3. Sonication for Loading Nucleic Acid Drugs
5.4. Transfection for Loading Nucleic Acid Drugs
6. Loading Strategies for Protein Drugs
6.1. Incubation for Loading Protein Drugs
6.2. Electroporation for Loading Protein Drugs
6.3. Sonication for Loading of Protein Drugs
6.4. Freeze–Thaw for Loading Protein Drugs
6.5. Transfection for Loading of Protein Drugs
6.6. Supplemental Loading Strategies for Protein Drugs
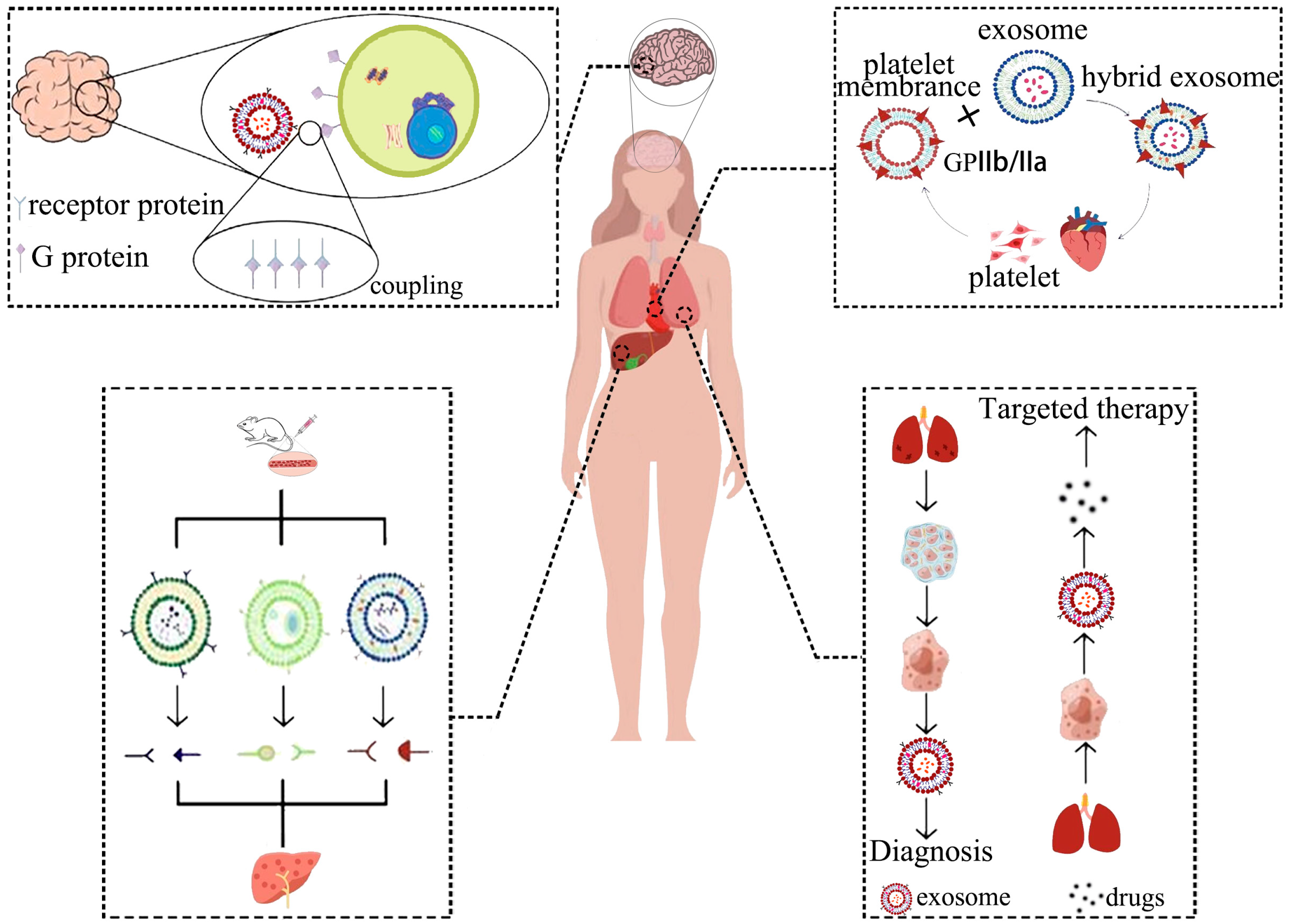
7. Engineered Exosome Targeted Delivery Methods
7.1. Brain Targeting Exosomes
7.2. Lung Targeting Exosomes
7.3. Cardiac Targeting Exosomes
7.4. Hepatic Targeting Exosomes
8. Prospects for Clinical Applications
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Zhang, K.; Tan, S.; Gao, F.; Zhang, Y.; Xu, W.; Wang, H.; Gu, D.; Zhu, L.; Li, S.; et al. Exosomal circLPAR1 functions in colorectal cancer diagnosis and tumorigenesis through suppressing BRD4 via METTL3–eIF3h interaction. Mol. Cancer 2022, 21, 49. [Google Scholar] [CrossRef] [PubMed]
- Sedej, I.; Štalekar, M.; Tušek Žnidarič, M.; Goričar, K.; Kojc, N.; Kogovšek, P.; Dolžan, V.; Arnol, M.; Lenassi, M. Extracellular vesicle-bound DNA in urine is indicative of kidney allograft injury. J. Extracell. Vesicles 2022, 11, e12268. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Chen, J.; Jiang, T.; Yan, C.; Kang, Y.; Zhang, M.; Xiang, K.; Guo, J.; Jiang, G.; Wang, C.; et al. Milk-derived exosomes carrying siRNA-KEAP1 promote diabetic wound healing by improving oxidative stress. Drug Deliv. Transl. Res. 2023, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gebara, N.; Scheel, J.; Skovronova, R.; Grange, C.; Marozio, L.; Gupta, S.; Giorgione, V.; Caicci, F.; Benedetto, C.; Khalil, A.; et al. Single extracellular vesicle analysis in human amniotic fluid shows evidence of phenotype alterations in preeclampsia. J. Extracell. Vesicles 2022, 11, e12217. [Google Scholar] [CrossRef]
- Cai, J.; Gong, L.; Li, G.; Guo, J.; Yi, X.; Wang, Z. Exosomes in ovarian cancer ascites promote epithelial–mesenchymal transition of ovarian cancer cells by delivery of miR-6780b-5p. Cell Death Dis. 2021, 12, 210. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef]
- Vietri, M.; Radulovic, M.; Stenmark, H. The many functions of ESCRTs. Nat. Rev. Mol. Cell Biol. 2020, 21, 25–42. [Google Scholar] [CrossRef]
- Gurung, S.; Perocheau, D.; Touramanidou, L.; Baruteau, J. The exosome journey: From biogenesis to uptake and intracellular signalling. Cell Commun. Signal. 2021, 19, 47. [Google Scholar] [CrossRef]
- Isaac, R.; Reis, F.C.G.; Ying, W.; Olefsky, J.M. Exosomes as mediators of intercellular crosstalk in metabolism. Cell Metab. 2021, 33, 1744–1762. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.V.; Da, R.S.A.; Ramalho, J.; Maximo, C.C.; Cardoso, M.H.; Pintado, P.; Carvalho, A.S.; Beck, H.C.; Matthiesen, R.; Zuzarte, M.; et al. LAMP2A regulates the loading of proteins into exosomes. Sci. Adv. 2022, 8, m1140. [Google Scholar] [CrossRef]
- Gomzikova, M.O.; James, V.; Rizvanov, A.A. Mitochondria Donation by Mesenchymal Stem Cells: Current Understanding and Mitochondria Transplantation Strategies. Front. Cell Dev. Biol. 2021, 9, 653322. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Uchiyama, J.; Imami, K.; Ishihama, Y.; Kageyama, R.; Kobayashi, T. Novel Roles of Small Extracellular Vesicles in Regulating the Quiescence and Proliferation of Neural Stem Cells. Front. Cell Dev. Biol. 2021, 9, 762293. [Google Scholar] [CrossRef]
- Peruzzotti-Jametti, L.; Bernstock, J.D.; Willis, C.M.; Manferrari, G.; Rogall, R.; Fernandez-Vizarra, E.; Williamson, J.C.; Braga, A.; van den Bosch, A.; Leonardi, T.; et al. Neural stem cells traffic functional mitochondria via extracellular vesicles. PLoS Biol. 2021, 19, e3001166. [Google Scholar] [CrossRef]
- Hu, K.; McKay, P.F.; Samnuan, K.; Najer, A.; Blakney, A.K.; Che, J.; O’Driscoll, G.; Cihova, M.; Stevens, M.M.; Shattock, R.J. Presentation of antigen on extracellular vesicles using transmembrane domains from viral glycoproteins for enhanced immunogenicity. J. Extracell. Vesicles 2022, 11, e12199. [Google Scholar] [CrossRef]
- Li, M.; Soder, R.; Abhyankar, S.; Abdelhakim, H.; Braun, M.W.; Trinidad, C.V.; Pathak, H.B.; Pessetto, Z.; Deighan, C.; Ganguly, S.; et al. WJMSC-derived small extracellular vesicle enhance T cell suppression through PD-L1. J. Extracell. Vesicles 2021, 10, e12067. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Su, C. Design strategies and application progress of therapeutic exosomes. Theranostics 2019, 9, 1015–1028. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Chen, L.; Zhang, H.; Qiu, X.; Huang, Q.; Wan, F.; Le, Z.; Geng, S.; Zhang, A.; Qiu, S.; et al. Tumor-intrinsic YTHDF1 drives immune evasion and resistance to immune checkpoint inhibitors via promoting MHC-I degradation. Nat. Commun. 2023, 14, 265. [Google Scholar] [CrossRef]
- Bai, K.; Lee, C.; Liu, X.; Li, J.; Cao, D.; Zhang, L.; Hu, D.; Li, H.; Hou, Y.; Xu, Y.; et al. Human placental exosomes induce maternal systemic immune tolerance by reprogramming circulating monocytes. J. Nanobiotechnol. 2022, 20, 86. [Google Scholar] [CrossRef]
- Meng, W.; Hao, Y.; He, C.; Li, L.; Zhu, G. Exosome-orchestrated hypoxic tumor microenvironment. Mol. Cancer 2019, 18, 57. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Wang, X.; Zhai, S.; Shi, M.; Peng, C.; Deng, X.; Fu, D.; Wang, J.; Shen, B. Hypoxia-induced exosomal circPDK1 promotes pancreatic cancer glycolysis via c-myc activation by modulating miR-628-3p/BPTF axis and degrading BIN1. J. Hematol. Oncol. 2022, 15, 128. [Google Scholar] [CrossRef] [PubMed]
- Mashouri, L.; Yousefi, H.; Aref, A.R.; Ahadi, A.M.; Molaei, F.; Alahari, S.K. Exosomes: Composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol. Cancer 2019, 18, 75. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, Y.; Liu, L.; Si, C.; Xu, Y.; Wu, X.; Wang, C.; Sun, Z.; Kang, Q. Exosomal circTUBGCP4 promotes vascular endothelial cell tipping and colorectal cancer metastasis by activating Akt signaling pathway. J. Exp. Clin. Cancer Res. 2023, 42, 46. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Liu, W.; Wei, W.; Zhao, Y.; Zhuang, P.; Wang, X.; Wang, Y.; Hu, Y.; Dai, H. Photothermal Hydrogel Encapsulating Intelligently Bacteria-Capturing Bio-MOF for Infectious Wound Healing. ACS Nano 2022, 16, 19491–19508. [Google Scholar] [CrossRef]
- Sadeghi, A.; Zare-Gachi, M.; Najjar-Asl, M.; Rajabi, S.; Fatemi, M.J.; Forghani, S.F.; Daemi, H.; Pezeshki-Modaress, M. Hybrid gelatin-sulfated alginate scaffolds as dermal substitutes can dramatically accelerate healing of full-thickness diabetic wounds. Carbohydr. Polym. 2023, 302, 120404. [Google Scholar] [CrossRef]
- Nourian, D.A.; Mirahmadi, B.F.; Chehelgerdi, M.; Raeisi, D.S. Skin tissue engineering: Wound healing based on stem-cell-based therapeutic strategies. Stem Cell Res. Ther. 2019, 10, 111. [Google Scholar] [CrossRef]
- Hade, M.D.; Suire, C.N.; Mossell, J.; Suo, Z. Extracellular vesicles: Emerging frontiers in wound healing. Med. Res. Rev. 2022, 42, 2102–2125. [Google Scholar] [CrossRef]
- Li, Q.; Hu, W.; Huang, Q.; Yang, J.; Li, B.; Ma, K.; Wei, Q.; Wang, Y.; Su, J.; Sun, M.; et al. MiR146a-loaded engineered exosomes released from silk fibroin patch promote diabetic wound healing by targeting IRAK1. Signal Transduct. Target. Ther. 2023, 8, 62. [Google Scholar] [CrossRef]
- Miyazaki, K.; Wada, Y.; Okuno, K.; Murano, T.; Morine, Y.; Ikemoto, T.; Saito, Y.; Ikematsu, H.; Kinugasa, Y.; Shimada, M.; et al. An exosome-based liquid biopsy signature for pre-operative identification of lymph node metastasis in patients with pathological high-risk T1 colorectal cancer. Mol. Cancer 2023, 22, 2. [Google Scholar] [CrossRef]
- Jang, Y.O.; Ahn, H.; Dao, T.N.T.; Hong, J.; Shin, W.; Lim, Y.; Chung, S.J.; Lee, J.; Liu, H.; Koo, B.; et al. Magnetic transferrin nanoparticles (MTNs) assay as a novel isolation approach for exosomal biomarkers in neurological diseases. Biomater. Res. 2023, 27, 12. [Google Scholar] [CrossRef]
- Vázquez Mera, S.; Martelo Vidal, L.; Miguéns Suárez, P.; Saavedra Nieves, P.; Arias, P.; González Fernández, C.; Mosteiro Añón, M.; Corbacho Abelaira, M.D.; Blanco Aparicio, M.; Méndez Brea, P.; et al. Serum exosome inflamma-miRs are surrogate biomarkers for asthma phenotype and severity. Allergy 2023, 78, 141–155. [Google Scholar] [CrossRef]
- Xiang, Z.; Jiang, B.; Li, W.; Zhai, G.; Zhou, H.; Wang, Y.; Wu, J. The diagnostic and prognostic value of serum exosome-derived carbamoyl phosphate synthase 1 in HEV-related acute liver failure patients. J. Med. Virol. 2022, 94, 5015–5025. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, H.; Fu, J.; Wu, X.; Liang, X.; Liu, X.; Wu, X.; Cao, L.; Xu, Z.; Dong, M. Microfluidic-based exosome isolation and highly sensitive aptamer exosome membrane protein detection for lung cancer diagnosis. Biosens. Bioelectron. 2022, 214, 114487. [Google Scholar] [CrossRef]
- Wang, H.; Lu, Z.; Zhao, X. Tumorigenesis, diagnosis, and therapeutic potential of exosomes in liver cancer. J. Hematol. Oncol. 2019, 12, 133. [Google Scholar] [CrossRef]
- Tang, X.; Guo, T.; Gao, X.; Wu, X.; Xing, X.; Ji, J.; Li, Z. Exosome-derived noncoding RNAs in gastric cancer: Functions and clinical applications. Mol. Cancer 2021, 20, 99. [Google Scholar] [CrossRef]
- Tenchov, R.; Sasso, J.M.; Wang, X.; Liaw, W.; Chen, C.; Zhou, Q.A. Exosomes—Nature’s Lipid Nanoparticles, a Rising Star in Drug Delivery and Diagnostics. ACS Nano 2022, 16, 17802–17846. [Google Scholar] [CrossRef]
- Li, M.Y.; Liu, L.Z.; Dong, M. Progress on pivotal role and application of exosome in lung cancer carcinogenesis, diagnosis, therapy and prognosis. Mol. Cancer 2021, 20, 22. [Google Scholar] [CrossRef]
- Mi, B.; Chen, L.; Xiong, Y.; Yang, Y.; Panayi, A.C.; Xue, H.; Hu, Y.; Yan, C.; Hu, L.; Xie, X.; et al. Osteoblast/Osteoclast and Immune Cocktail Therapy of an Exosome/Drug Delivery Multifunctional Hydrogel Accelerates Fracture Repair. ACS Nano 2022, 16, 771–782. [Google Scholar] [CrossRef]
- Yerneni, S.S.; Yalcintas, E.P.; Smith, J.D.; Averick, S.; Campbell, P.G.; Ozdoganlar, O.B. Skin-targeted delivery of extracellular vesicle-encapsulated curcumin using dissolvable microneedle arrays. Acta Biomater. 2022, 149, 198–212. [Google Scholar] [CrossRef]
- Terstappen, G.C.; Meyer, A.H.; Bell, R.D.; Zhang, W. Strategies for delivering therapeutics across the blood-brain barrier. Nat. Rev. Drug Discov. 2021, 20, 362–383. [Google Scholar] [CrossRef]
- Zhan, Q.; Yi, K.; Qi, H.; Li, S.; Li, X.; Wang, Q.; Wang, Y.; Liu, C.; Qiu, M.; Yuan, X.; et al. Engineering blood exosomes for tumor-targeting efficient gene/chemo combination therapy. Theranostics 2020, 10, 7889–7905. [Google Scholar] [CrossRef]
- Subhan, M.A.; Torchilin, V.P. Efficient nanocarriers of siRNA therapeutics for cancer treatment. Transl. Res. 2019, 214, 62–91. [Google Scholar] [CrossRef]
- Murphy, D.E.; de Jong, O.G.; Brouwer, M.; Wood, M.J.; Lavieu, G.; Schiffelers, R.M.; Vader, P. Extracellular vesicle-based therapeutics: Natural versus engineered targeting and trafficking. Exp. Mol. Med. 2019, 51, 1–12. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Peng, Y.; Du, Y.; Yang, Z.; Qi, X. Dendritic cell derived exosomes loaded neoantigens for personalized cancer immunotherapies. J. Control. Release 2023, 353, 423–433. [Google Scholar] [CrossRef]
- Zhu, S.; Huang, H.; Liu, D.; Wen, S.; Shen, L.; Lin, Q. Augmented cellular uptake and homologous targeting of exosome-based drug loaded IOL for posterior capsular opacification prevention and biosafety improvement. Bioact. Mater. 2022, 15, 469–481. [Google Scholar] [CrossRef]
- Zhao, S.; Mi, Y.; Zheng, B.; Wei, P.; Gu, Y.; Zhang, Z.; Xu, Y.; Cai, S.; Li, X.; Li, D. Highly-metastatic colorectal cancer cell released miR-181a-5p-rich extracellular vesicles promote liver metastasis by activating hepatic stellate cells and remodelling the tumour microenvironment. J. Extracell. Vesicles 2022, 11, e12186. [Google Scholar] [CrossRef]
- Rehman, F.U.; Liu, Y.; Yang, Q.; Yang, H.; Liu, R.; Zhang, D.; Muhammad, P.; Liu, Y.; Hanif, S.; Ismail, M.; et al. Heme Oxygenase-1 targeting exosomes for temozolomide resistant glioblastoma synergistic therapy. J. Control. Release 2022, 345, 696–708. [Google Scholar] [CrossRef]
- Cully, M. Exosome-based candidates move into the clinic. Nat. Rev. Drug Discov. 2021, 20, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Qu, M.; Lin, Q.; Huang, L.; Fu, Y.; Wang, L.; He, S.; Fu, Y.; Yang, S.; Zhang, Z.; Zhang, L.; et al. Dopamine-loaded blood exosomes targeted to brain for better treatment of Parkinson’s disease. J. Control. Release 2018, 287, 156–166. [Google Scholar] [CrossRef]
- Zhang, K.; Dong, C.; Chen, M.; Yang, T.; Wang, X.; Gao, Y.; Wang, L.; Wen, Y.; Chen, G.; Wang, X.; et al. Extracellular vesicle-mediated delivery of miR-101 inhibits lung metastasis in osteosarcoma. Theranostics 2020, 10, 411–425. [Google Scholar] [CrossRef]
- Cui, J.; Wang, X.; Li, J.; Zhu, A.; Du, Y.; Zeng, W.; Guo, Y.; Di, L.; Wang, R. Immune Exosomes Loading Self-Assembled Nanomicelles Traverse the Blood–Brain Barrier for Chemo-immunotherapy against Glioblastoma. ACS Nano 2023, 17, 1464–1484. [Google Scholar] [CrossRef]
- Guo, J.; Wang, F.; Hu, Y.; Luo, Y.; Wei, Y.; Xu, K.; Zhang, H.; Liu, H.; Bo, L.; Lv, S.; et al. Exosome-based bone-targeting drug delivery alleviates impaired osteoblastic bone formation and bone loss in inflammatory bowel diseases. Cell Rep. Med. 2023, 4, 100881. [Google Scholar] [CrossRef]
- Wang, C.; Li, N.; Li, Y.; Hou, S.; Zhang, W.; Meng, Z.; Wang, S.; Jia, Q.; Tan, J.; Wang, R.; et al. Engineering a HEK-293T exosome-based delivery platform for efficient tumor-targeting chemotherapy/internal irradiation combination therapy. J. Nanobiotechnol. 2022, 20, 247. [Google Scholar] [CrossRef]
- Al Faruque, H.; Choi, E.; Kim, J.; Kim, E. Enhanced effect of autologous EVs delivering paclitaxel in pancreatic cancer. J. Control. Release 2022, 347, 330–346. [Google Scholar] [CrossRef]
- Yang, X.; Shi, G.; Guo, J.; Wang, C.; He, Y. Exosome-encapsulated antibiotic against intracellular infections of methicillin-resistant Staphylococcus aureus. Int. J. Nanomed. 2018, 13, 8095–8104. [Google Scholar] [CrossRef]
- Wu, G.; Zhang, J.; Zhao, Q.; Zhuang, W.; Ding, J.; Zhang, C.; Gao, H.; Pang, D.W.; Pu, K.; Xie, H.Y. Molecularly Engineered Macrophage-Derived Exosomes with Inflammation Tropism and Intrinsic Heme Biosynthesis for Atherosclerosis Treatment. Angew. Chem. 2020, 132, 4097–4103. [Google Scholar] [CrossRef]
- Lennaard, A.J.; Mamand, D.R.; Wiklander, R.J.; El, A.S.; Wiklander, O. Optimised Electroporation for Loading of Extracellular Vesicles with Doxorubicin. Pharmaceutics 2021, 14, 38. [Google Scholar] [CrossRef]
- Jia, G.; Han, Y.; An, Y.; Ding, Y.; He, C.; Wang, X.; Tang, Q. NRP-1 targeted and cargo-loaded exosomes facilitate simultaneous imaging and therapy of glioma in vitro and in vivo. Biomaterials 2018, 178, 302–316. [Google Scholar] [CrossRef]
- Le Saux, S.; Aarrass, H.; Lai-Kee-Him, J.; Bron, P.; Armengaud, J.; Miotello, G.; Bertrand-Michel, J.; Dubois, E.; George, S.; Faklaris, O.; et al. Post-production modifications of murine mesenchymal stem cell (mMSC) derived extracellular vesicles (EVs) and impact on their cellular interaction. Biomaterials 2020, 231, 119675. [Google Scholar] [CrossRef]
- Wang, P.; Wang, H.; Huang, Q.; Peng, C.; Yao, L.; Chen, H.; Qiu, Z.; Wu, Y.; Wang, L.; Chen, W. Exosomes from M1-Polarized Macrophages Enhance Paclitaxel Antitumor Activity by Activating Macrophages-Mediated Inflammation. Theranostics 2019, 9, 1714–1727. [Google Scholar] [CrossRef]
- Du, J.; Wan, Z.; Wang, C.; Lu, F.; Wei, M.; Wang, D.; Hao, Q. Designer exosomes for targeted and efficient ferroptosis induction in cancer via chemo-photodynamic therapy. Theranostics 2021, 11, 8185–8196. [Google Scholar] [CrossRef]
- Haney, M.J.; Klyachko, N.L.; Harrison, E.B.; Zhao, Y.; Kabanov, A.V.; Batrakova, E.V. TPP1 Delivery to Lysosomes with Extracellular Vesicles and their Enhanced Brain Distribution in the Animal Model of Batten Disease. Adv. Healthc. Mater. 2019, 8, 1801271. [Google Scholar] [CrossRef]
- Sun, L.; Fan, M.; Huang, D.; Li, B.; Xu, R.; Gao, F.; Chen, Y. Clodronate-loaded liposomal and fibroblast-derived exosomal hybrid system for enhanced drug delivery to pulmonary fibrosis. Biomaterials 2021, 271, 120761. [Google Scholar] [CrossRef]
- Zhang, L.; Fan, C.; Hao, W.; Zhuang, Y.; Liu, X.; Zhao, Y.; Chen, B.; Xiao, Z.; Chen, Y.; Dai, J. NSCs Migration Promoted and Drug Delivered Exosomes-Collagen Scaffold via a Bio-Specific Peptide for One-Step Spinal Cord Injury Repair. Adv. Healthc. Mater. 2021, 10, 2001896. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, L.; Guo, H.; Chen, L.; Huang, X. Dapagliflozin-Loaded Exosome Mimetics Facilitate Diabetic Wound Healing by HIF-1α-Mediated Enhancement of Angiogenesis. Adv. Healthc. Mater. 2022, 12, 2202751. [Google Scholar] [CrossRef]
- Duan, L.; Xu, L.M.; Xu, X.; Qin, Z.A.; Zhou, X.Y.; Xiao, Y.; Liang, Y.J.; Xia, J. Exosome-mediated delivery of gene vectors for gene therapy. Nanoscale 2021, 13, 1387–1397. [Google Scholar] [CrossRef]
- Rayamajhi, S.; Nguyen, T.D.T.; Marasini, R.; Aryal, S. Macrophage-derived exosome-mimetic hybrid vesicles for tumor targeted drug delivery. Acta Biomater. 2019, 94, 482–494. [Google Scholar] [CrossRef]
- Lin, Y.; Wu, J.; Gu, W.; Huang, Y.; Tong, Z.; Huang, L.; Tan, J. Exosome-Liposome Hybrid Nanoparticles Deliver CRISPR/Cas9 System in MSCs. Adv. Sci. 2018, 5, 1700611. [Google Scholar] [CrossRef]
- Chan, M.H.; Chang, Z.X.; Huang, C.F.; Lee, L.J.; Liu, R.S.; Hsiao, M. Integrated therapy platform of exosomal system: Hybrid inorganic/organic nanoparticles with exosomes for cancer treatment. Nanoscale Horiz. 2022, 7, 352–367. [Google Scholar] [CrossRef]
- Li, L.; He, D.; Guo, Q.; Zhang, Z.; Ru, D.; Wang, L.; Gong, K.; Liu, F.; Duan, Y.; Li, H. Exosome-liposome hybrid nanoparticle codelivery of TP and miR497 conspicuously overcomes chemoresistant ovarian cancer. J. Nanobiotechnol. 2022, 20, 50. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Cheng, L.; Lu, Y.; Zhang, X.; Wang, Y.; Deng, J.; Zhou, J.; Liu, B.; Liu, J. Thermosensitive Exosome–Liposome Hybrid Nanoparticle-Mediated Chemoimmunotherapy for Improved Treatment of Metastatic Peritoneal Cancer. Adv. Sci. 2020, 7, 2000515. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wu, D.; Ma, X.; Wang, J.; Hou, W.; Zhang, W. Exosomes as drug carriers for cancer therapy and challenges regarding exosome uptake. Biomed. Pharmacother. 2020, 128, 110237. [Google Scholar] [CrossRef]
- Sancho-Albero, M.; Encabo-Berzosa, M.D.M.; Beltrán-Visiedo, M.; Fernández-Messina, L.; Sebastián, V.; Sánchez-Madrid, F.; Arruebo, M.; Santamaría, J.; Martín-Duque, P. Efficient encapsulation of theranostic nanoparticles in cell-derived exosomes: Leveraging the exosomal biogenesis pathway to obtain hollow gold nanoparticle-hybrids. Nanoscale 2019, 11, 18825–18836. [Google Scholar] [CrossRef]
- Ma, Q.; Fan, Q.; Han, X.; Dong, Z.; Xu, J.; Bai, J.; Tao, W.; Sun, D.; Wang, C. Platelet-derived extracellular vesicles to target plaque inflammation for effective anti-atherosclerotic therapy. J. Control. Release 2021, 329, 445–453. [Google Scholar] [CrossRef]
- Murphy, D.E.; de Jong, O.G.; Evers, M.J.W.; Nurazizah, M.; Schiffelers, R.M.; Vader, P. Natural or Synthetic RNA Delivery: A Stoichiometric Comparison of Extracellular Vesicles and Synthetic Nanoparticles. Nano Lett. 2021, 21, 1888–1895. [Google Scholar] [CrossRef]
- Herrmann, I.K.; Wood, M.; Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat. Nanotechnol. 2021, 16, 748–759. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, H.; Kim, V.N. Sequence determinant of small RNA production by DICER. Nature 2023, 615, 323–330. [Google Scholar] [CrossRef]
- Gong, C.; Tian, J.; Wang, Z.; Gao, Y.; Wu, X.; Ding, X.; Qiang, L.; Li, G.; Han, Z.; Yuan, Y.; et al. Functional exosome-mediated co-delivery of doxorubicin and hydrophobically modified microRNA 159 for triple-negative breast cancer therapy. J. Nanobiotechnol. 2019, 17, 93. [Google Scholar] [CrossRef]
- Haraszti, R.A.; Miller, R.; Didiot, M.; Biscans, A.; Alterman, J.F.; Hassler, M.R.; Roux, L.; Echeverria, D.; Sapp, E.; DiFiglia, M.; et al. Optimized Cholesterol-siRNA Chemistry Improves Productive Loading onto Extracellular Vesicles. Mol. Ther. 2018, 26, 1973–1982. [Google Scholar] [CrossRef]
- Davis, S.M.; Hariharan, V.N.; Lo, A.; Turanov, A.A.; Echeverria, D.; Sousa, J.; McHugh, N.; Biscans, A.; Alterman, J.F.; Karumanchi, S.A.; et al. Chemical optimization of siRNA for safe and efficient silencing of placental sFLT1. Mol. Ther. Nucleic Acids 2022, 29, 135–149. [Google Scholar] [CrossRef]
- Hade, M.D.; Suire, C.N.; Suo, Z. An Effective Peptide-Based Platform for Efficient Exosomal Loading and Cellular Delivery of a microRNA. ACS Appl. Mater. Interfaces 2023, 15, 3851–3866. [Google Scholar] [CrossRef]
- Rong, Y.; Wang, Z.; Tang, P.; Wang, J.; Ji, C.; Chang, J.; Zhu, Y.; Ye, W.; Bai, J.; Liu, W.; et al. Engineered extracellular vesicles for delivery of siRNA promoting targeted repair of traumatic spinal cord injury. Bioact. Mater. 2023, 23, 328–342. [Google Scholar] [CrossRef] [PubMed]
- Reshke, R.; Taylor, J.A.; Savard, A.; Guo, H.; Rhym, L.H.; Kowalski, P.S.; Trung, M.T.; Campbell, C.; Little, W.; Anderson, D.G.; et al. Reduction of the therapeutic dose of silencing RNA by packaging it in extracellular vesicles via a pre-microRNA backbone. Nat. Biomed. Eng. 2020, 4, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Zhu, Y.; Ali, D.J.; Tian, T.; Xu, H.; Si, K.; Sun, B.; Chen, B.; Xiao, Z. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J. Nanobiotechnol. 2020, 18, 10. [Google Scholar] [CrossRef]
- Kim, G.; Kim, M.; Lee, Y.; Byun, J.W.; Hwang, D.W.; Lee, M. Systemic delivery of microRNA-21 antisense oligonucleotides to the brain using T7-peptide decorated exosomes. J. Control. Release 2020, 317, 273–281. [Google Scholar] [CrossRef]
- Zhao, Z.; Shuang, T.; Gao, Y.; Lu, F.; Zhang, J.; He, W.; Qu, L.; Chen, B.; Hao, Q. Targeted delivery of exosomal miR-484 reprograms tumor vasculature for chemotherapy sensitization. Cancer Lett. 2022, 530, 45–58. [Google Scholar] [CrossRef]
- Yang, Z.; Shi, J.; Xie, J.; Wang, Y.; Sun, J.; Liu, T.; Zhao, Y.; Zhao, X.; Wang, X.; Ma, Y.; et al. Large-scale generation of functional mRNA-encapsulating exosomes via cellular nanoporation. Nat. Biomed. Eng. 2020, 4, 69–83. [Google Scholar] [CrossRef]
- Tao, H.; Xu, H.; Zuo, L.; Li, C.; Qiao, G.; Guo, M.; Zheng, L.; Leitgeb, M.; Lin, X. Exosomes-coated bcl-2 siRNA inhibits the growth of digestive system tumors both in vitro and in vivo. Int. J. Biol. Macromol. 2020, 161, 470–480. [Google Scholar] [CrossRef]
- Born, L.J.; Chang, K.H.; Shoureshi, P.; Lay, F.; Bengali, S.; Hsu, A.T.W.; Abadchi, S.N.; Harmon, J.W.; Jay, S.M. HOTAIR-Loaded Mesenchymal Stem/Stromal Cell Extracellular Vesicles Enhance Angiogenesis and Wound Healing. Adv. Healthc. Mater. 2022, 11, 2002070. [Google Scholar] [CrossRef]
- Yu, X.; Bai, Y.; Han, B.; Ju, M.; Tang, T.; Shen, L.; Li, M.; Yang, L.; Zhang, Z.; Hu, G.; et al. Extracellular vesicle-mediated delivery of circDYM alleviates CUS-induced depressive-like behaviours. J. Extracell. Vesicles 2022, 11, e12185. [Google Scholar] [CrossRef] [PubMed]
- Abreu, R.C.; Ramos, C.V.; Becher, C.; Lino, M.; Jesus, C.; Costa Martins, P.A.; Martins, P.A.T.; Moreno, M.J.; Fernandes, H.; Ferreira, L. Exogenous loading of miRNAs into small extracellular vesicles. J. Extracell. Vesicles 2021, 10, e12111. [Google Scholar] [CrossRef]
- Xu, L.; Faruqu, F.N.; Lim, Y.M.; Lim, K.Y.; Liam-Or, R.; Walters, A.A.; Lavender, P.; Fear, D.; Wells, C.M.; Tzu-Wen Wang, J.; et al. Exosome-mediated RNAi of PAK4 prolongs survival of pancreatic cancer mouse model after loco-regional treatment. Biomaterials 2021, 264, 120369. [Google Scholar] [CrossRef]
- McCann, J.; Sosa Miranda, C.D.; Guo, H.; Reshke, R.; Savard, A.; Zardini Buzatto, A.; Taylor, J.A.; Li, L.; Gibbings, D.J. Contaminating transfection complexes can masquerade as small extracellular vesicles and impair their delivery of RNA. J. Extracell. Vesicles 2022, 11, e12220. [Google Scholar] [CrossRef]
- McConnell, R.E.; Youniss, M.; Gnanasambandam, B.; Shah, P.; Zhang, W.; Finn, J.D. Transfection reagent artefact likely accounts for some reports of extracellular vesicle function. J. Extracell. Vesicles 2022, 11, e12253. [Google Scholar] [CrossRef]
- Lagassé, H.A.D.; McCormick, Q.; Sauna, Z.E. Secondary failure: Immune responses to approved protein therapeutics. Trends Mol. Med. 2021, 27, 1074–1083. [Google Scholar] [CrossRef]
- Gouveia, M.G.; Wesseler, J.P.; Ramaekers, J.; Weder, C.; Scholten, P.; Bruns, N. Polymersome-based protein drug delivery—Quo vadis? Chem. Soc. Rev. 2023, 52, 728–778. [Google Scholar] [CrossRef]
- Wan, T.; Zhong, J.; Pan, Q.; Zhou, T.; Ping, Y.; Liu, X. Exosome-mediated delivery of Cas9 ribonucleoprotein complexes for tissue-specific gene therapy of liver diseases. Sci. Adv. 2022, 8, p9435. [Google Scholar] [CrossRef]
- Yao, X.; Lyu, P.; Yoo, K.; Yadav, M.K.; Singh, R.; Atala, A.; Lu, B. Engineered extracellular vesicles as versatile ribonucleoprotein delivery vehicles for efficient and safe CRISPR genome editing. J. Extracell. Vesicles 2021, 10, e12076. [Google Scholar] [CrossRef]
- Haney, M.J.; Klyachko, N.L.; Zhao, Y.; Gupta, R.; Plotnikova, E.G.; He, Z.; Patel, T.; Piroyan, A.; Sokolsky, M.; Kabanov, A.V.; et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J. Control. Release 2015, 207, 18–30. [Google Scholar] [CrossRef]
- Fuhrmann, G.; Chandrawati, R.; Parmar, P.A.; Keane, T.J.; Maynard, S.A.; Bertazzo, S.; Stevens, M.M. Engineering Extracellular Vesicles with the Tools of Enzyme Prodrug Therapy. Adv. Mater. 2018, 30, 1706616. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Morales, B.; Antunes-Ricardo, M.; González-Valdez, J. Exosome-Mediated Insulin Delivery for the Potential Treatment of Diabetes Mellitus. Pharmaceutics 2021, 13, 1870. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Yang, X.; He, C.; Chen, Y.; Li, C.; Li, S.; Chen, Y.; Wu, Y.; Xiang, Z.; Kang, J.; et al. Rejuvenation of tendon stem/progenitor cells for functional tendon regeneration through platelet-derived exosomes loaded with recombinant Yap1. Acta Biomater. 2023, 161, 80–99. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Tan, J.; Wu, C.; Zhang, J.; Liu, T.; Fan, C.; Li, J.; Zhang, Y. Extracellular vesicles engineered with valency-controlled DNA nanostructures deliver CRISPR/Cas9 system for gene therapy. Nucleic Acids Res. 2020, 48, 8870–8882. [Google Scholar] [CrossRef]
- Görgens, A.; Corso, G.; Hagey, D.W.; Jawad Wiklander, R.; Gustafsson, M.O.; Felldin, U.; Lee, Y.; Bostancioglu, R.B.; Sork, H.; Liang, X.; et al. Identification of storage conditions stabilizing extracellular vesicles preparations. J. Extracell. Vesicles 2022, 11, e12238. [Google Scholar] [CrossRef]
- Hajipour, H.; Farzadi, L.; Roshangar, L.; Latifi, Z.; Kahroba, H.; Shahnazi, V.; Hamdi, K.; Ghasemzadeh, A.; Fattahi, A.; Nouri, M. A human chorionic gonadotropin (hCG) delivery platform using engineered uterine exosomes to improve endometrial receptivity. Life Sci. 2021, 275, 119351. [Google Scholar] [CrossRef]
- Zou, X.; Yuan, M.; Zhang, T.; Zheng, N.; Wu, Z. EVs Containing Host Restriction Factor IFITM3 Inhibited ZIKV Infection of Fetuses in Pregnant Mice through Trans-placenta Delivery. Mol. Ther. 2021, 29, 176–190. [Google Scholar] [CrossRef]
- Chen, R.; Huang, H.; Liu, H.; Xi, J.; Ning, J.; Zeng, W.; Shen, C.; Zhang, T.; Yu, G.; Xu, Q.; et al. Friend or Foe? Evidence Indicates Endogenous Exosomes Can Deliver Functional gRNA and Cas9 Protein. Small 2019, 15, 1902686. [Google Scholar] [CrossRef]
- Majeau, N.; Fortin-Archambault, A.; Gérard, C.; Rousseau, J.; Yaméogo, P.; Tremblay, J.P. Serum extracellular vesicles for delivery of CRISPR-CAS9 ribonucleoproteins to modify the dystrophin gene. Mol. Ther. 2022, 30, 2429–2442. [Google Scholar] [CrossRef]
- Yim, N.; Ryu, S.; Choi, K.; Lee, K.R.; Lee, S.; Choi, H.; Kim, J.; Shaker, M.R.; Sun, W.; Park, J.; et al. Exosome engineering for efficient intracellular delivery of soluble proteins using optically reversible protein–protein interaction module. Nat. Commun. 2016, 7, 12277. [Google Scholar] [CrossRef]
- Choi, H.; Kim, Y.; Mirzaaghasi, A.; Heo, J.; Kim, Y.N.; Shin, J.H.; Kim, S.; Kim, N.H.; Cho, E.S.; In Yook, J.; et al. Exosome-based delivery of super-repressor IκBα relieves sepsis-associated organ damage and mortality. Sci. Adv. 2020, 6, z6980. [Google Scholar] [CrossRef] [PubMed]
- Pandit, R.; Chen, L.; Götz, J. The blood-brain barrier: Physiology and strategies for drug delivery. Adv. Drug Deliv. Rev. 2020, 165–166, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Venkat, P.; Cui, C.; Chopp, M.; Zacharek, A.; Wang, F.; Landschoot-Ward, J.; Shen, Y.; Chen, J. MiR-126 Mediates Brain Endothelial Cell Exosome Treatment–Induced Neurorestorative Effects after Stroke in Type 2 Diabetes Mellitus Mice. Stroke 2019, 50, 2865–2874. [Google Scholar] [CrossRef]
- Ge, X.; Guo, M.; Hu, T.; Li, W.; Huang, S.; Yin, Z.; Li, Y.; Chen, F.; Zhu, L.; Kang, C.; et al. Increased Microglial Exosomal miR-124-3p Alleviates Neurodegeneration and Improves Cognitive Outcome after rmTBI. Mol. Ther. 2020, 28, 503–522. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Huang, Y.; Liu, Y.; Zhang, X.; Wang, Q.; Liu, Y.; Ma, X.; Long, X.; Ruan, Y.; Lei, H.; et al. Astrocyte-derived exosomal lncRNA 4933431K23Rik modulates microglial phenotype and improves post-traumatic recovery via SMAD7 regulation. Mol. Ther. 2023, 31, 1313–1331. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, M.J.; Xie, J.; Ceroi, A.; Balusu, S.; Castelein, J.; Van Wonterghem, E.; Van Imschoot, G.; Ward, A.; Menheniott, T.R.; Gustafsson, O.; et al. Choroid plexus-derived extracellular vesicles exhibit brain targeting characteristics. Biomaterials 2022, 290, 121830. [Google Scholar] [CrossRef]
- Morad, G.; Carman, C.V.; Hagedorn, E.J.; Perlin, J.R.; Zon, L.I.; Mustafaoglu, N.; Park, T.; Ingber, D.E.; Daisy, C.C.; Moses, M.A. Tumor-Derived Extracellular Vesicles Breach the Intact Blood–Brain Barriervia Transcytosis. ACS Nano 2019, 13, 13853–13865. [Google Scholar] [CrossRef]
- Tian, T.; Liang, R.; Erel-Akbaba, G.; Saad, L.; Obeid, P.J.; Gao, J.; Chiocca, E.A.; Weissleder, R.; Tannous, B.A. Immune Checkpoint Inhibition in GBM Primed with Radiation by Engineered Extracellular Vesicles. ACS Nano 2022, 16, 1940–1953. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, C.; Xia, N.; Tian, H.; Li, D.; Lin, J.; Mei, X.; Wu, C. Berberine-loaded M2 macrophage-derived exosomes for spinal cord injury therapy. Acta Biomater. 2021, 126, 211–223. [Google Scholar] [CrossRef]
- Pham, T.C.; Jayasinghe, M.K.; Pham, T.T.; Yang, Y.; Wei, L.; Usman, W.M.; Chen, H.; Pirisinu, M.; Gong, J.; Kim, S.; et al. Covalent conjugation of extracellular vesicles with peptides and nanobodies for targeted therapeutic delivery. J. Extracell. Vesicles 2021, 10, e12057. [Google Scholar] [CrossRef]
- Khalaj, K.; Figueira, R.L.; Antounians, L.; Lauriti, G.; Zani, A. Systematic review of extracellular vesicle-based treatments for lung injury: Are EVs a potential therapy for COVID-19? J. Extracell. Vesicles 2020, 9, 1795365. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Su, P.; Pan, T.; Zhou, X.; Li, H.; Huang, H.; Wang, A.; Wang, F.; Huang, J.; Yan, H.; et al. Engineering Extracellular Vesicles Enriched with Palmitoylated ACE2 as COVID-19 Therapy. Adv. Mater. 2021, 33, 2103471. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M. GPIIb/IIIa-GPVI–commanded platelet patrol. Blood 2022, 140, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wang, X.; Li, Z.; Zhu, D.; Cores, J.; Wang, Z.; Li, J.; Mei, X.; Cheng, X.; Su, T.; et al. Platelet membrane and stem cell exosome hybrids enhance cellular uptake and targeting to heart injury. Nano Today 2021, 39, 101210. [Google Scholar] [CrossRef]
- Li, Q.; Huang, Z.; Wang, Q.; Gao, J.; Chen, J.; Tan, H.; Li, S.; Wang, Z.; Weng, X.; Yang, H.; et al. Targeted immunomodulation therapy for cardiac repair by platelet membrane engineering extracellular vesicles via hitching peripheral monocytes. Biomaterials 2022, 284, 121529. [Google Scholar] [CrossRef]
- Zhang, G.; Huang, X.; Xiu, H.; Sun, Y.; Chen, J.; Cheng, G.; Song, Z.; Peng, Y.; Shen, Y.; Wang, J.; et al. Extracellular vesicles: Natural liver-accumulating drug delivery vehicles for the treatment of liver diseases. J. Extracell. Vesicles 2020, 10, e12030. [Google Scholar] [CrossRef]
- He, Y.; Rodrigues, R.M.; Wang, X.; Seo, W.; Ma, J.; Hwang, S.; Fu, Y.; Trojnar, E.; Matyas, C.; Zhao, S.; et al. Neutrophil-to-hepatocyte communication via LDLR-dependent miR-223-enriched extracellular vesicle transfer ameliorates nonalcoholic steatohepatitis. J. Clin. Investig. 2021, 131, e141513. [Google Scholar] [CrossRef]
- Zuo, B.; Zhang, Y.; Zhao, K.; Wu, L.; Qi, H.; Yang, R.; Gao, X.; Geng, M.; Wu, Y.; Jing, R.; et al. Universal immunotherapeutic strategy for hepatocellular carcinoma with exosome vaccines that engage adaptive and innate immune responses. J. Hematol. Oncol. 2022, 15, 46. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Load Strategy | Loaded Drug | Exosome Source | Method for Loading Drug into Exosome | Load Efficiency | Ref. | |
|---|---|---|---|---|---|---|
| Incubation | Small molecule | DOX | HEK-293T | 4 °C, 2 h | 11.73% | [54] |
| PTX | Human pancreatic ductal carcinoma cell | Room temperature, 1 h | 4.2 ± 0.63% | [55] | ||
| Linezolid | Macrophage | 37 °C, 1 h | 5.06 ± 0.45% | [56] | ||
| Nucleic acid | miR-159 | Macrophage | 37 °C, 1.5 h | 5.33% | [79] | |
| siRNA | Mesenchymal stem cell | 37 °C, 1 h | 43% | [80] | ||
| miR-21-5p | Mesenchymal stem cell | Room temperature, 8 h | 1600 copies of miRNA/exosome | [82] | ||
| Protein | catalase | Macrophage | Room temperature, 18 h | 4.9 ± 0.5% | [100] | |
| β-glucuronidase | human mesenchymal stem cells | Room temperature, 10 min | NS | [101] | ||
| Electroporation | Small molecule | DOX | Lens epithelial cell | 250 V, 350 μF, 4.5 ms | 72% | [46] |
| HAL | M2 macrophage | 100 V, 200 Ω, 100 μF | 25.14% | [57] | ||
| Curcumin and SPIONs | Mouse macrophage cell line Raw264.7 | 400 V, 150 μF, 1 ms | NS | [59] | ||
| Nucleic acid | siRNA | Induced neural stem cell | 400 mV, 125 μF, 10–15 ms | 17% | [83] | |
| miR-21 inhibitor oligonucleotides | HEK-293T | 1000 V, 10 ms | 0.5% | [85] | ||
| miRNA-21 | 293T | 400 V | 1.68 ± 0.23% | [86] | ||
| miR-484 | HEK-293T | 700 V, 150 mF | NS | [87] | ||
| Protein | Insulin | Hepatocellular carcinoma cell | 200 V, 50 μF | 50.75 ± 1.2% | [102] | |
| Primary dermal fibroblast | 57.42 ± 5.47% | |||||
| Pancreatic β-cell | 49.70 ± 4.32% | |||||
| Recombinant Yap1 protein | Platelet | 200 V, 500 μF, 26 ms | 40.65 ± 3.79% | [103] | ||
| Cas9 RNP | Hepatic stellate cell | NS | 20% | [98] | ||
| Sonication | Small molecule | PTX | M1 macrophage | 20% amplitude, 6 cycles of 30 s on/off for 3 min with a 2 min cooling period between each cycle | 19.55 ± 2.48% | [61] |
| Curcumin | Macrophage | 20% amplitude, six cycles of 30 s on/off for 3 min with a 2 min cooling period between each cycle | 0.56 ± 0.01 μg curcumin (1 μg albumin-EVs) | [40] | ||
| Er and RB | HEK-293T | 20% amplitude, 6 cycles, 10 s on/off, 3 min duration, 2 min cooling period between each cycle | 84% (RB), 60% (Er) | [62] | ||
| Nucleic acid | Bcl-2 siRNA | Milk | 30 W, sonicated for 5 s, and then stopped for 2 s for 2 min | 66.9 ± 4.5% | [89] | |
| siRNA-Keap1 | Milk | 20% amplitude, six 30-s on/off cycles, and a cooling time of 2 min between each cycle | 24% | [4] | ||
| Protein | Catalase | Macrophage | 500 V, 2 kHz, 20% power, 4 s pulses/2 s pauses, repeat 6 times | 26.1 ± 1.2% | [100] | |
| TPP1 | Macrophage | Sonication in water bath at room temperature for 30 min | 70 μg TPP1/1011 exosomes | [63] | ||
| Cas9 RNP | HEK-293T | 2 kHz, 5% power, 2 s pulse/1 s pause and repeated 20 cycles | 15.34% | [104] | ||
| Extrusion | Small molecule | PTX | huc-MSCs | 100 nm pore size extrusion 11 times | 14.23 ± 0.25% | [65] |
| DA | Induced pluripotent stem cell differentiated from endothelial cell | 400 and 200 nm polycarbonate membranes extrusion 10–20 times | 45% | [66] | ||
| Protein | Catalase | Macrophage | 200 nm pore size | 22.2 ± 3.1% | [100] | |
| Freeze–Thaw Cycle | Protein | Catalase | Macrophage | −80 °C fast freeze, thaw at room temperature, repeat cycle 3 times | 14.7 ± 1.1% | [100] |
| hCG | Uterine fluid | −70 °C fast freeze, thaw at room temperature, repeat cycle 5 times | 14.02 ± 5.46% | [106] | ||
| Cas9 RNP | HEK-293T | Freeze in liquid nitrogen, thaw at room temperature, repeat cycle 3 times | 37.62% | [104] | ||
| Chimeric Exosome Method | Small molecule | TP | Cisplatin-resistant human ovarian cancer cell line | Vortex and sonication (33% amplitude, 2 s pulsed on/off, for 3 min) | 78 ± 3% | [71] |
| DTX | Fibroblast | Freeze in liquid nitrogen for 5 min, thaw at room temperature for 15 min, repeat 3 times | 4.3% | [72] | ||
| CLD | Fibroblast | Vortex and sonication (30% amplitude, 30 s pulse on/off, for 2 min) | 70.6 ± 2.7% | [64] | ||
| NIN | Fibroblast | Mixed with EL-CLD in chloroform | 90% | [64] | ||
| Endogenous loading | Small molecule | PEG-HGNs | Murine melanoma cell | Drug was added to the cells and cultured for 24 h | 50% | [74] |
| MCC950 | Platelet | Incubation at 4 °C for 12 h | 7.1% | [75] | ||
| Transfection | Nucleic acid | HOTAIR | BDMSCs | pCMV-HOTAIR plasmid, P3000 reagent | Cargo expression increased 974-fold | [90] |
| circDYM | HEK-293T | circDYM-GFP lentivirus, Lipofectamine 2000 | 34.75 copies/exosome | [91] | ||
| miRNA | Human umbilical cord blood-derived mononuclear cell, human urine, foetal bovine serum | Exo-Fect | Above 50% | [92] | ||
| Protein | IFITM3 | 293T | IFITM3 expression vector, PEI transfection reagent | NS | [107] | |
| Cas9 protein | HeLa cell, HuH7 cell | HPV-specific and HBV-specific CRISPR/Cas9 expression plasmid | NS | [108] | ||
| CRISPR RNP | Serum | Lipofectamine CRISPRMAX | 96% | [109] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, H.; Guo, S.; Ren, X.; Wu, Z.; Liu, S.; Yao, X. Current Strategies for Exosome Cargo Loading and Targeting Delivery. Cells 2023, 12, 1416. https://doi.org/10.3390/cells12101416
Zeng H, Guo S, Ren X, Wu Z, Liu S, Yao X. Current Strategies for Exosome Cargo Loading and Targeting Delivery. Cells. 2023; 12(10):1416. https://doi.org/10.3390/cells12101416
Chicago/Turabian StyleZeng, Haifeng, Shaoshen Guo, Xuancheng Ren, Zhenkun Wu, Shuwen Liu, and Xingang Yao. 2023. "Current Strategies for Exosome Cargo Loading and Targeting Delivery" Cells 12, no. 10: 1416. https://doi.org/10.3390/cells12101416
APA StyleZeng, H., Guo, S., Ren, X., Wu, Z., Liu, S., & Yao, X. (2023). Current Strategies for Exosome Cargo Loading and Targeting Delivery. Cells, 12(10), 1416. https://doi.org/10.3390/cells12101416