

Methylglyoxal Impairs the Pro-Angiogenic Ability of Mouse Adipose-Derived Stem Cells (mADSCs) via a Senescence-Associated Mechanism

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. mADSCs Isolation and Culture
2.3. mADSCs Characterization
- (a)
- Immunophenotype
- (b)
- Adipogenic differentiation
- (c)
- Oil Red-O staining
- (d)
- Osteogenic differentiation
- (e)
- Alizarin Red S staining
2.4. Cell Culture Procedure
2.5. Cell Viability
2.6. Population Doubling Time (PDT)
2.7. Western Blot Analysis
2.8. RNA Isolation, Reverse Transcription and Real-Time PCR
2.9. Migration and Co-Culture Assays
2.10. Detection of Senescence-Associated Beta-Galactosidase (SA-βgal) Activity
2.11. Multiplex Immunoassay
2.12. Statistical Procedures
3. Results
3.1. Characterization of Isolated mADSCs
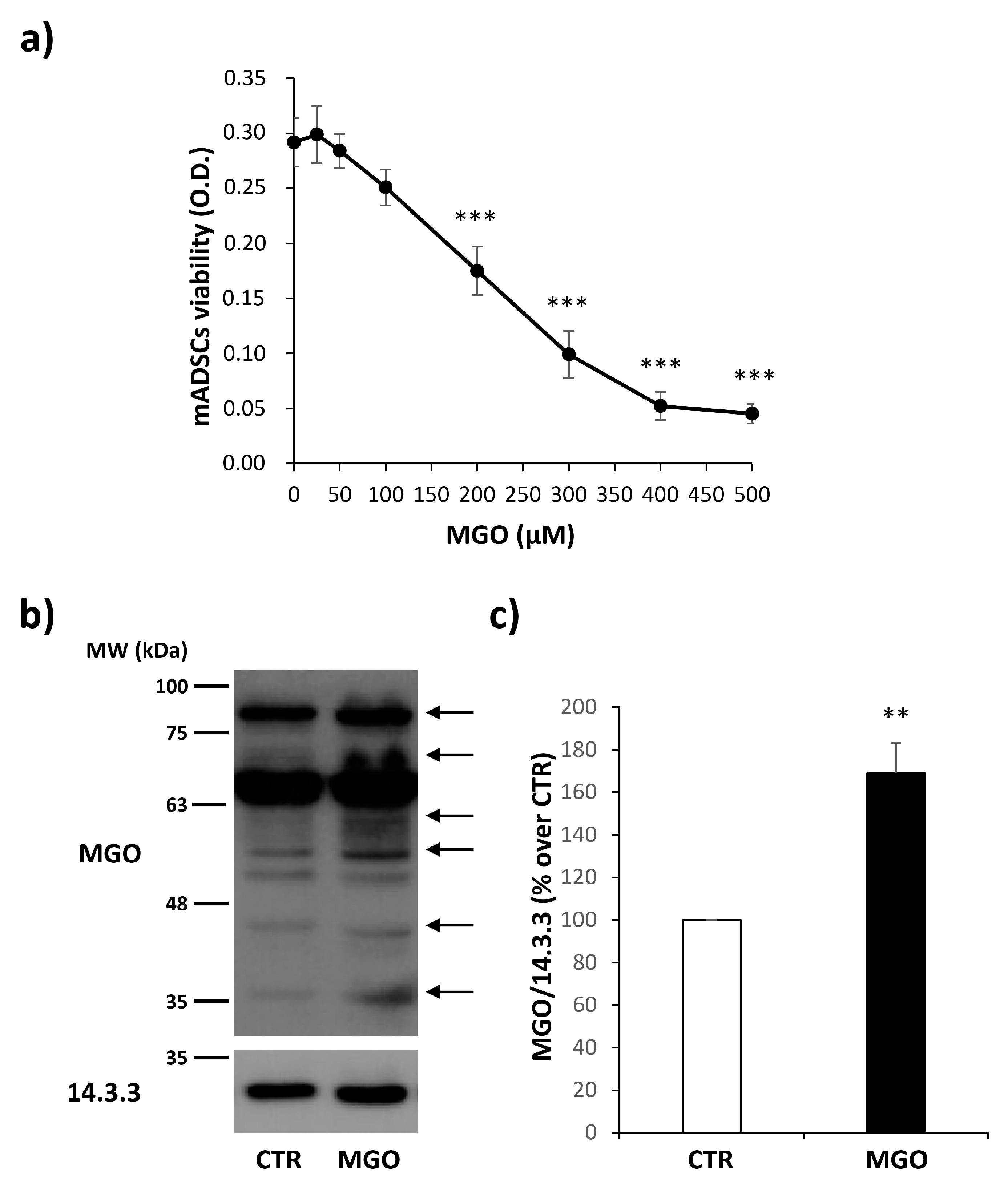
3.2. MGO Treatment Does Not Impair mADSC Viability and Induces an Accumulation of MGO-Adducts
3.3. MGO Treatment Impairs the Pro-Angiogenic Ability of mADSCs
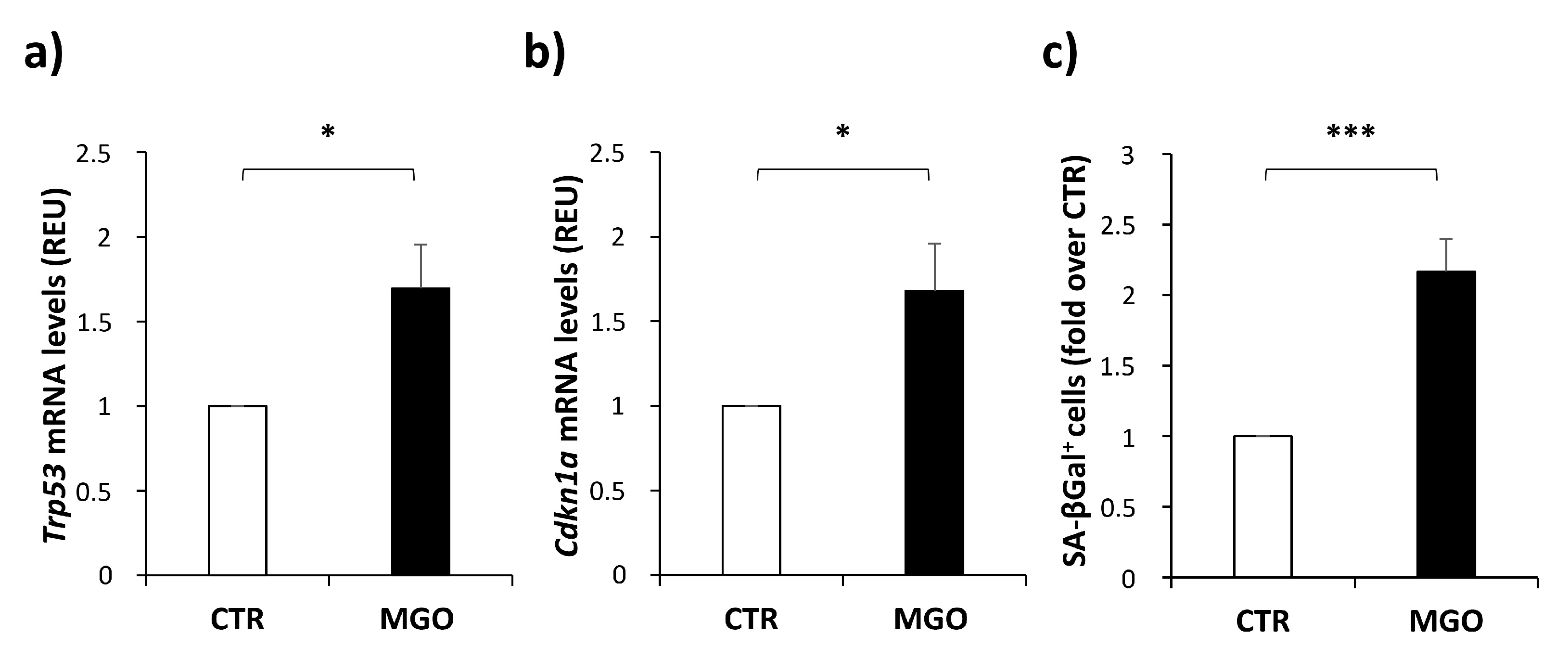
3.4. MGO Treatment Induces Senescence in mADSCs
3.5. MGO Treatment Impairs the Soluble Factor Dependent Pro-Angiogenic Ability of mADSCs
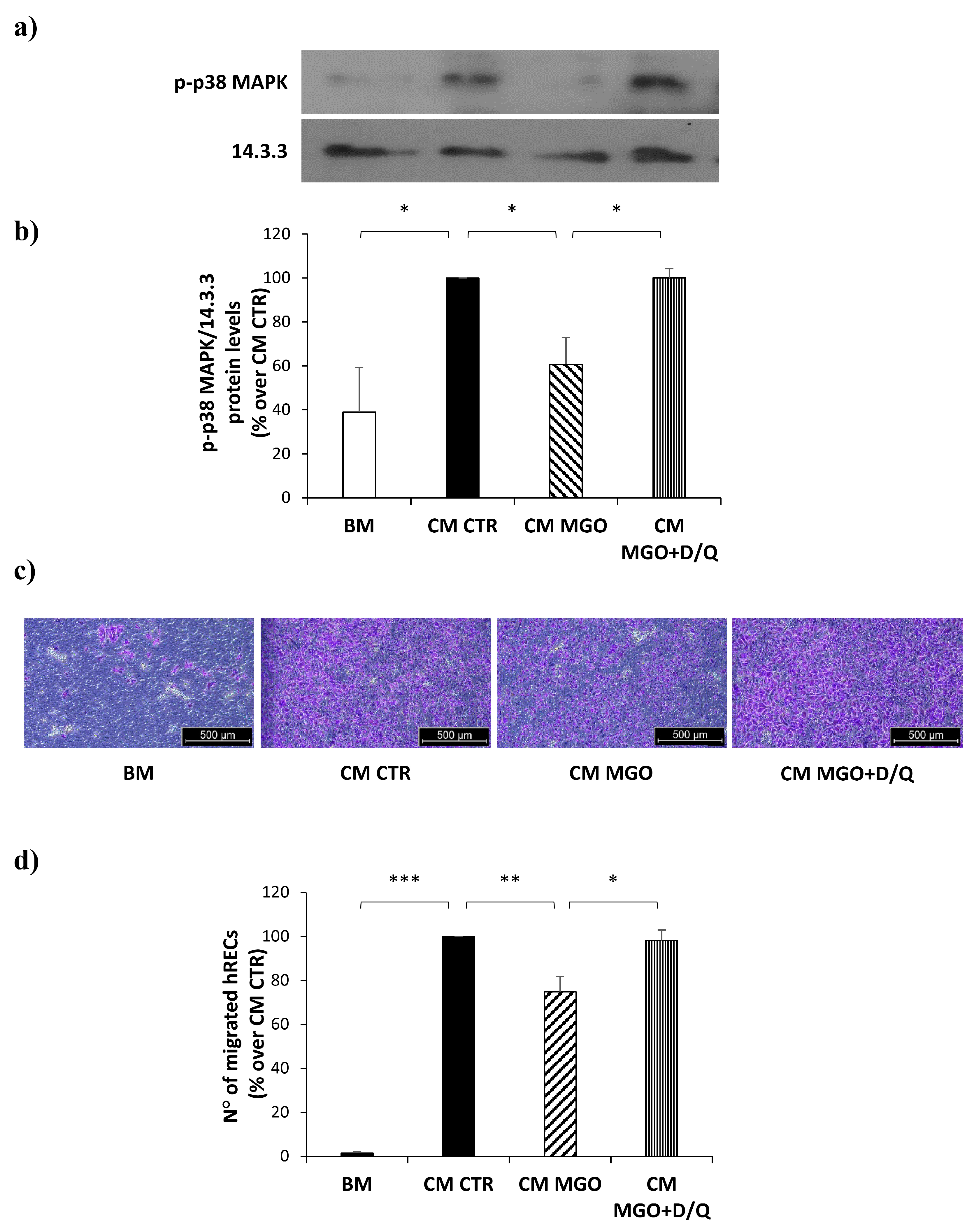
3.6. p38 MAPK Activation Is Reduced by CM from MGO-Treated mADSCs in hRECs
3.7. Senolytic Treatment of mADSCs Exposed to MGO Restores Their Ability to Reduce Migration of hRECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yin, M.; Zhang, Y.; Yu, H.; Li, X. Role of Hyperglycemia in the Senescence of Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, 9, 665412. [Google Scholar] [CrossRef] [PubMed]
- Trzyna, A.; Banas-Zabczyk, A. Adipose-Derived Stem Cells Secretome and Its Potential Application in “Stem Cell-Free Therapy”. Biomolecules 2021, 11, 878. [Google Scholar] [CrossRef] [PubMed]
- Maacha, S.; Sidahmed, H.; Jacob, S.; Gentilcore, G.; Calzone, R.; Grivel, J.C.; Cugno, C. Paracrine Mechanisms of Mesenchymal Stromal Cells in Angiogenesis. Stem. Cells Int. 2020, 2020, 4356359. [Google Scholar] [CrossRef]
- Hutchings, G.; Janowicz, K.; Moncrieff, L.; Dompe, C.; Strauss, E.; Kocherova, I.; Nawrocki, M.J.; Kruszyna, L.; Wasiatycz, G.; Antosik, P.; et al. The Proliferation and Differentiation of Adipose-Derived Stem Cells in Neovascularization and Angiogenesis. Int. J. Mol. Sci. 2020, 21, 3790. [Google Scholar] [CrossRef]
- Krawczenko, A.; Klimczak, A. Adipose Tissue-Derived Mesenchymal Stem/Stromal Cells and Their Contribution to Angiogenic Processes in Tissue Regeneration. Int. J. Mol. Sci. 2022, 23, 2425. [Google Scholar] [CrossRef]
- Dzhoyashvili, N.A.; Efimenko, A.Y.; Kochegura, T.N.; Kalinina, N.I.; Koptelova, N.V.; Sukhareva, O.Y.; Shestakova, M.V.; Akchurin, R.S.; Tkachuk, V.A.; Parfyonova, Y.V. Disturbed angiogenic activity of adipose-derived stromal cells obtained from patients with coronary artery disease and diabetes mellitus type 2. J. Transl. Med. 2014, 12, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, O.; Usui, S.; Takashima, S.I.; Nomura, A.; Yamaguchi, K.; Takeda, Y.; Goten, C.; Hamaoka, T.; Ootsuji, H.; Murai, H.; et al. Diabetes impairs the angiogenic capacity of human adipose-derived stem cells by reducing the CD271(+) subpopulation in adipose tissue. Biochem. Biophys. Res. Commun. 2019, 517, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kim, Y.H.; Jun, Y.J.; Yoo, G.; Rhie, J.W. The effect of diabetes on the wound healing potential of adipose-tissue derived stem cells. Int. Wound J. 2016, 13 (Suppl. S1), 33–41. [Google Scholar] [CrossRef] [Green Version]
- Allaman, I.; Belanger, M.; Magistretti, P.J. Methylglyoxal, the dark side of glycolysis. Front. Neurosci. 2015, 9, 23. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, N.; Thornalley, P.J. Dicarbonyls linked to damage in the powerhouse: Glycation of mitochondrial proteins and oxidative stress. Biochem. Soc. Trans. 2008, 36, 1045–1050. [Google Scholar] [CrossRef] [Green Version]
- Nigro, C.; Leone, A.; Fiory, F.; Prevenzano, I.; Nicolo, A.; Mirra, P.; Beguinot, F.; Miele, C. Dicarbonyl Stress at the Crossroads of Healthy and Unhealthy Aging. Cells 2019, 8, 749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prevenzano, I.; Leone, A.; Longo, M.; Nicolo, A.; Cabaro, S.; Collina, F.; Panarese, I.; Botti, G.; Formisano, P.; Napoli, R.; et al. Glyoxalase 1 knockdown induces age-related beta-cell dysfunction and glucose intolerance in mice. EMBO Rep. 2022, 23, e52990. [Google Scholar] [CrossRef]
- Nigro, C.; Leone, A.; Raciti, G.A.; Longo, M.; Mirra, P.; Formisano, P.; Beguinot, F.; Miele, C. Methylglyoxal-Glyoxalase 1 Balance: The Root of Vascular Damage. Int. J. Mol. Sci. 2017, 18, 188. [Google Scholar] [CrossRef] [Green Version]
- Nigro, C.; Leone, A.; Longo, M.; Prevenzano, I.; Fleming, T.H.; Nicolo, A.; Parrillo, L.; Spinelli, R.; Formisano, P.; Nawroth, P.P.; et al. Methylglyoxal accumulation de-regulates HoxA5 expression, thereby impairing angiogenesis in glyoxalase 1 knock-down mouse aortic endothelial cells. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 73–85. [Google Scholar] [CrossRef]
- Waqas, K.; Muller, M.; Koedam, M.; El Kadi, Y.; Zillikens, M.C.; van der Eerden, B.C.J. Methylglyoxal—An advanced glycation end products (AGEs) precursor—Inhibits differentiation of human MSC-derived osteoblasts in vitro independently of receptor for AGEs (RAGE). Bone 2022, 164, 116526. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Yang, X.; Qin, J.; Ye, K.; Wang, X.; Shi, H.; Jiang, M.; Liu, X.; Lu, X. Glyoxalase-1 Overexpression Reverses Defective Proangiogenic Function of Diabetic Adipose-Derived Stem Cells in Streptozotocin-Induced Diabetic Mice Model of Critical Limb Ischemia. Stem. Cells Transl. Med. 2017, 6, 261–271. [Google Scholar] [CrossRef] [Green Version]
- D’Esposito, V.; Lecce, M.; Marenzi, G.; Cabaro, S.; Ambrosio, M.R.; Sammartino, G.; Misso, S.; Migliaccio, T.; Liguoro, P.; Oriente, F.; et al. Platelet-rich plasma counteracts detrimental effect of high-glucose concentrations on mesenchymal stem cells from Bichat fat pad. J. Tissue Eng. Regen. Med. 2020, 14, 701–713. [Google Scholar] [CrossRef]
- Lee, M.S.; Youn, C.; Kim, J.H.; Park, B.J.; Ahn, J.; Hong, S.; Kim, Y.D.; Shin, Y.K.; Park, S.G. Enhanced Cell Growth of Adipocyte-Derived Mesenchymal Stem Cells Using Chemically-Defined Serum-Free Media. Int. J. Mol. Sci. 2017, 18, 1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debacq-Chainiaux, F.; Erusalimsky, J.D.; Campisi, J.; Toussaint, O. Protocols to detect senescence-associated beta-galactosidase (SA-betagal) activity, a biomarker of senescent cells in culture and in vivo. Nat. Protoc. 2009, 4, 1798–1806. [Google Scholar] [CrossRef]
- Agareva, M.; Stafeev, I.; Michurina, S.; Sklyanik, I.; Shestakova, E.; Ratner, E.; Hu, X.; Menshikov, M.; Shestakova, M.; Parfyonova, Y. Type 2 Diabetes Mellitus Facilitates Shift of Adipose-Derived Stem Cells Ex Vivo Differentiation toward Osteogenesis among Patients with Obesity. Life 2022, 12, 688. [Google Scholar] [CrossRef]
- Legzdina, D.; Romanauska, A.; Nikulshin, S.; Kozlovska, T.; Berzins, U. Characterization of Senescence of Culture-expanded Human Adipose-derived Mesenchymal Stem Cells. Int. J. Stem. Cells 2016, 9, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Shi, G.; Lei, X.; Huang, Y.; Li, X.; Bai, L.; Qin, C. Age-related alteration in characteristics, function, and transcription features of ADSCs. Stem. Cell Res. Ther. 2021, 12, 473. [Google Scholar] [CrossRef]
- Kwon, H.M.; Hur, S.M.; Park, K.Y.; Kim, C.K.; Kim, Y.M.; Kim, H.S.; Shin, H.C.; Won, M.H.; Ha, K.S.; Kwon, Y.G.; et al. Multiple paracrine factors secreted by mesenchymal stem cells contribute to angiogenesis. Vascul. Pharmacol. 2014, 63, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Ma, J.; Li, S.; Liu, W. Applicability of adipose-derived mesenchymal stem cells in treatment of patients with type 2 diabetes. Stem. Cell Res. Ther. 2019, 10, 274. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, R.; Yan, S.F.; Schmidt, A.M. Methylglyoxal comes of AGE. Cell 2006, 124, 258–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.H.; Yang, H.M.; Park, J.B.; Choi, G.S.; Joh, J.W.; Kwon, C.H.; Chun, J.M.; Lee, S.K.; Kim, S.J. Isolation and characterization of mouse mesenchymal stem cells. Transplant. Proc. 2008, 40, 2649–2654. [Google Scholar] [CrossRef]
- Taha, M.F.; Hedayati, V. Isolation, identification and multipotential differentiation of mouse adipose tissue-derived stem cells. Tissue Cell 2010, 42, 211–216. [Google Scholar] [CrossRef]
- Abdallah, B.M.; Alzahrani, A.M.; Abdel-Moneim, A.M.; Ditzel, N.; Kassem, M. A simple and reliable protocol for long-term culture of murine bone marrow stromal (mesenchymal) stem cells that retained their in vitro and in vivo stemness in long-term culture. Biol. Proced. Online 2019, 21, 3. [Google Scholar] [CrossRef]
- Fosmark, D.S.; Torjesen, P.A.; Kilhovd, B.K.; Berg, T.J.; Sandvik, L.; Hanssen, K.F.; Agardh, C.D.; Agardh, E. Increased serum levels of the specific advanced glycation end product methylglyoxal-derived hydroimidazolone are associated with retinopathy in patients with type 2 diabetes mellitus. Metabolism 2006, 55, 232–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabbani, N.; Thornalley, P.J. Methylglyoxal, glyoxalase 1 and the dicarbonyl proteome. Amino Acids 2012, 42, 1133–1142. [Google Scholar] [CrossRef]
- Rabbani, N.; Thornalley, P.J. Dicarbonyl stress in cell and tissue dysfunction contributing to ageing and disease. Biochem. Biophys. Res. Commun. 2015, 458, 221–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnellan, L.; Young, C.; Simpson, B.S.; Acland, M.; Dhillon, V.S.; Costabile, M.; Fenech, M.; Hoffmann, P.; Deo, P. Proteomic Analysis of Methylglyoxal Modifications Reveals Susceptibility of Glycolytic Enzymes to Dicarbonyl Stress. Int. J. Mol. Sci. 2022, 23, 3689. [Google Scholar] [CrossRef]
- Koch, A.E.; Distler, O. Vasculopathy and disordered angiogenesis in selected rheumatic diseases: Rheumatoid arthritis and systemic sclerosis. Arthritis Res. Ther. 2007, 9 (Suppl. S2), S3. [Google Scholar] [CrossRef] [Green Version]
- Vulesevic, B.; McNeill, B.; Giacco, F.; Maeda, K.; Blackburn, N.J.; Brownlee, M.; Milne, R.W.; Suuronen, E.J. Methylglyoxal-Induced Endothelial Cell Loss and Inflammation Contribute to the Development of Diabetic Cardiomyopathy. Diabetes 2016, 65, 1699–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigro, C.; Raciti, G.A.; Leone, A.; Fleming, T.H.; Longo, M.; Prevenzano, I.; Fiory, F.; Mirra, P.; D’Esposito, V.; Ulianich, L.; et al. Methylglyoxal impairs endothelial insulin sensitivity both in vitro and in vivo. Diabetologia 2014, 57, 1485–1494. [Google Scholar] [CrossRef]
- Liu, H.; Yu, S.; Zhang, H.; Xu, J. Angiogenesis impairment in diabetes: Role of methylglyoxal-induced receptor for advanced glycation endproducts, autophagy and vascular endothelial growth factor receptor 2. PLoS ONE 2012, 7, e46720. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Lin, S.; Ao, Q.; He, X. The co-culture of ASCs and EPCs promotes vascularized bone regeneration in critical-sized bone defects of cranial bone in rats. Stem. Cell Res. Ther. 2020, 11, 338. [Google Scholar] [CrossRef]
- Lupo, G.; Agafonova, A.; Cosentino, A.; Giurdanella, G.; Mannino, G.; Lo Furno, D.; Romano, I.R.; Giuffrida, R.; D’Angeli, F.; Anfuso, C.D. Protective Effects of Human Pericyte-like Adipose-Derived Mesenchymal Stem Cells on Human Retinal Endothelial Cells in an In Vitro Model of Diabetic Retinopathy: Evidence for Autologous Cell Therapy. Int. J. Mol. Sci. 2023, 24, 913. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Lie, P.; Miao, T.; Yu, M.; Lu, Q.; Feng, T.; Li, J.; Zu, T.; Liu, X.; Li, H. Conditioned medium from umbilical cord mesenchymal stem cells induces migration and angiogenesis. Mol. Med. Rep. 2015, 12, 20–30. [Google Scholar] [CrossRef] [Green Version]
- Collawn, S.S.; Mobley, J.A.; Banerjee, N.S.; Chow, L.T. Conditioned Media From Adipose-Derived Stromal Cells Accelerates Healing in 3-Dimensional Skin Cultures. Ann. Plast. Surg. 2016, 76, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Dong, R.; Chang, M.; Liang, X.; Wang, H.C. Adipose-Derived Stem Cells for the Treatment of Diabetic Wound: From Basic Study to Clinical Application. Front. Endocrinol. 2022, 13, 882469. [Google Scholar] [CrossRef]
- Suvakov, S.; Cubro, H.; White, W.M.; Butler Tobah, Y.S.; Weissgerber, T.L.; Jordan, K.L.; Zhu, X.Y.; Woollard, J.R.; Chebib, F.T.; Milic, N.M.; et al. Targeting senescence improves angiogenic potential of adipose-derived mesenchymal stem cells in patients with preeclampsia. Biol. Sex. Differ. 2019, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Ratushnyy, A.; Ezdakova, M.; Buravkova, L. Secretome of Senescent Adipose-Derived Mesenchymal Stem Cells Negatively Regulates Angiogenesis. Int. J. Mol. Sci. 2020, 21, 1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Hong, Y.; Zhang, H.; Li, X. Mesenchymal Stem Cell Senescence and Rejuvenation: Current Status and Challenges. Front. Cell Dev. Biol. 2020, 8, 364. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Pirtskhalava, T.; Farr, J.N.; Weigand, B.M.; Palmer, A.K.; Weivoda, M.M.; Inman, C.L.; Ogrodnik, M.B.; Hachfeld, C.M.; Fraser, D.G.; et al. Senolytics improve physical function and increase lifespan in old age. Nat. Med. 2018, 24, 1246–1256. [Google Scholar] [CrossRef]
- Wiley, C.D.; Campisi, J. The metabolic roots of senescence: Mechanisms and opportunities for intervention. Nat. Metab. 2021, 3, 1290–1301. [Google Scholar] [CrossRef]
- Fallah, A.; Sadeghinia, A.; Kahroba, H.; Samadi, A.; Heidari, H.R.; Bradaran, B.; Zeinali, S.; Molavi, O. Therapeutic targeting of angiogenesis molecular pathways in angiogenesis-dependent diseases. Biomed. Pharmacother. 2019, 110, 775–785. [Google Scholar] [CrossRef]
- Shen, S.; Wang, F.; Fernandez, A.; Hu, W. Role of platelet-derived growth factor in type II diabetes mellitus and its complications. Diab. Vasc. Dis. Res. 2020, 17, 1479164120942119. [Google Scholar] [CrossRef]
- Corre, I.; Paris, F.; Huot, J. The p38 pathway, a major pleiotropic cascade that transduces stress and metastatic signals in endothelial cells. Oncotarget 2017, 8, 55684–55714. [Google Scholar] [CrossRef]
- Yoshizuka, N.; Chen, R.M.; Xu, Z.; Liao, R.; Hong, L.; Hu, W.Y.; Yu, G.; Han, J.; Chen, L.; Sun, P. A novel function of p38-regulated/activated kinase in endothelial cell migration and tumor angiogenesis. Mol. Cell Biol. 2012, 32, 606–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Lv, Z.; Chen, Q.; Guo, M.; Wang, X.; Huang, F. Vascular endothelial growth factor over-expressed mesenchymal stem cells-conditioned media ameliorate palmitate-induced diabetic endothelial dysfunction through PI-3K/AKT/m-TOR/eNOS and p38/MAPK signaling pathway. Biomed. Pharmacother. 2018, 106, 491–498. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′ to 3′) |
|---|---|
| Cyclophilin A: | Forward GCAGACAAAGTTCCAAAGACAG Reverse CACCCTGGCACATGAATCC |
| Fatty acid binding protein 4 (Fabp4 or Ap2): | Forward TCTCACCTGGAAGACAGCTCC Reverse GCTGATGATCATGTTGGGCTTGG |
| Peroxisome proliferator-activated receptor gamma 2 (Pparγ2): | Forward CAGTGGAGACCGCCCAGGCT Reverse TGGAGCAGGGGGTGAAGGCT |
| Glucose transporter member 4 (Glut4): | Forward CAATGTCTTGGCCGTGTTGG Reverse GCCCTGATGTTAGCCCTGAG |
| Adiponectin (AdipoQ): | Forward CTGACGACACCAAAAGGGCTC Reverse TCCAACCTGCACAAGTTCCC |
| Transformation-related protein 53 (Trp53): | Forward CCTCTCCCCCGCAAAAGAAA Reverse GACTCCTCTGTAGCATGGGC |
| Cyclin-dependent kinase inhibitor 1A (Cdkn1a): | Forward GCAGACCAGCCTGACAGATTT Reverse CTGACCCACAGCAGAAGAGG |
| Runt-related transcription factor 2 (Runx2): | Forward AGTCCATGCAGGAATATTTAAGGC Reverse CCAAAAGAAGCTTTGCTGACA |
| Msh homeobox 2 (Msx2): | Forward CCAGACATATGAGCCCCACC Reverse ACAGGTACTGTTTCTGGCGG |
| Osteopontin (Ocp): | Forward: CCGAGGTGATAGCTTGGCTT Reverse: ACAGGGATGACATCGAGGGA |
| Osteocalcin (Ocn): | Forward: GGTAGTGAACAGACTCCGGC Reverse: GGGCAGCACAGGTCCTAAAT |
| Variables | CTR | MGO |
|---|---|---|
| IL-6 (pg/mL × 1/µg of proteins) | 0.20 ± 0.03 | 0.32 ± 0.04 * |
| MCP-1(MCAF) (pg/mL × 1/µg of proteins) | 52.15 ± 4.40 | 80.29 ± 8.44 * |
| IL-12p40 (pg/mL × 1/µg of proteins) | 0.24 ± 0.03 | 0.35 ± 0.04 * |
| IL-1β (pg/mL × 1/µg of proteins) | 0.009 ± 0.003 | 0.01 ± 0.0005 |
| IL-5 (pg/mL × 1/µg of proteins) | 0.04 ± 0.01 | 0.036 ± 0.01 |
| Eotaxin (pg/mL × 1/µg of proteins) | 1.16 ± 0.29 | 1.80 ± 0.46 |
| G-CSF (pg/mL × 1/µg of proteins) | 0.12 ± 0.07 | 0.17 ± 0.08 |
| GM-CSF (pg/mL × 1/µg of proteins) | 0.12 ± 0.05 | 0.08 ± 0.04 |
| IFN-ƴ (pg/mL × 1/µg of proteins) | 0.03 ± 0.01 | 0.03 ± 0.01 |
| KC (pg/mL × 1/µg of proteins) | 3.40 ± 0.80 | 4.33 ± 0.81 |
| MIP-1β (pg/mL × 1/µg of proteins) | 0.19 ± 0.04 | 0.21 ± 0.05 |
| RANTES (pg/mL × 1/µg of proteins) | 0.32 ± 0.08 | 0.41 ± 0.18 |
| TNF-α (pg/mL × 1/µg of proteins) | 0.05 ± 0.003 | 0.04 ± 0.01 |
| Variables | CTR | MGO |
|---|---|---|
| VEGF (pg/mL × 1/µg of proteins) | 30.88 ± 3.02 | 18.34 ± 0.67 ** |
| PDGF-BB (pg/mL × 1/µg of proteins) | 0.31 ± 0.05 | 0.05 ± 0.06 * |
| Basic FGF (pg/mL × 1/µg of proteins) | 1.13 ± 0.87 | 0.83 ± 0.67 |
| M-CSF (pg/mL × 1/µg of proteins) | 4.90 ± 0.94 | 3.83 ± 0.43 |
| MIP2 (pg/mL × 1/µg of proteins) | 0.07 ± 0.02 | 0.04 ± 0.01 |
| Variables | CTR | MGO | MGO + D/Q |
|---|---|---|---|
| VEGF (pg/mL × 1/µg of proteins) | 31.28 ± 1.59 | 16.27 ± 1.42 *** | 31.90 ± 0.78 ### |
| PDGF-BB (pg/mL × 1/µg of proteins) | 0.54 ± 0.12 | 0.18 ± 0.07 * | 0.55 ± 0.05 ## |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leone, A.; Nicolò, A.; Prevenzano, I.; Zatterale, F.; Longo, M.; Desiderio, A.; Spinelli, R.; Campitelli, M.; Conza, D.; Raciti, G.A.; et al. Methylglyoxal Impairs the Pro-Angiogenic Ability of Mouse Adipose-Derived Stem Cells (mADSCs) via a Senescence-Associated Mechanism. Cells 2023, 12, 1741. https://doi.org/10.3390/cells12131741
Leone A, Nicolò A, Prevenzano I, Zatterale F, Longo M, Desiderio A, Spinelli R, Campitelli M, Conza D, Raciti GA, et al. Methylglyoxal Impairs the Pro-Angiogenic Ability of Mouse Adipose-Derived Stem Cells (mADSCs) via a Senescence-Associated Mechanism. Cells. 2023; 12(13):1741. https://doi.org/10.3390/cells12131741
Chicago/Turabian StyleLeone, Alessia, Antonella Nicolò, Immacolata Prevenzano, Federica Zatterale, Michele Longo, Antonella Desiderio, Rosa Spinelli, Michele Campitelli, Domenico Conza, Gregory Alexander Raciti, and et al. 2023. "Methylglyoxal Impairs the Pro-Angiogenic Ability of Mouse Adipose-Derived Stem Cells (mADSCs) via a Senescence-Associated Mechanism" Cells 12, no. 13: 1741. https://doi.org/10.3390/cells12131741
APA StyleLeone, A., Nicolò, A., Prevenzano, I., Zatterale, F., Longo, M., Desiderio, A., Spinelli, R., Campitelli, M., Conza, D., Raciti, G. A., Beguinot, F., Nigro, C., & Miele, C. (2023). Methylglyoxal Impairs the Pro-Angiogenic Ability of Mouse Adipose-Derived Stem Cells (mADSCs) via a Senescence-Associated Mechanism. Cells, 12(13), 1741. https://doi.org/10.3390/cells12131741