
Macromolecule Translocation across the Intestinal Mucosa of HIV-Infected Patients by Transcytosis and through Apoptotic Leaks

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Macromolecule Permeability of the Duodenal and Sigmoid Mucosa
2.3. Cell Culture Experiments
2.4. Apoptosis Assays and Inhibitor Studies
2.5. Western Blots
2.6. Statistical Analysis
3. Results
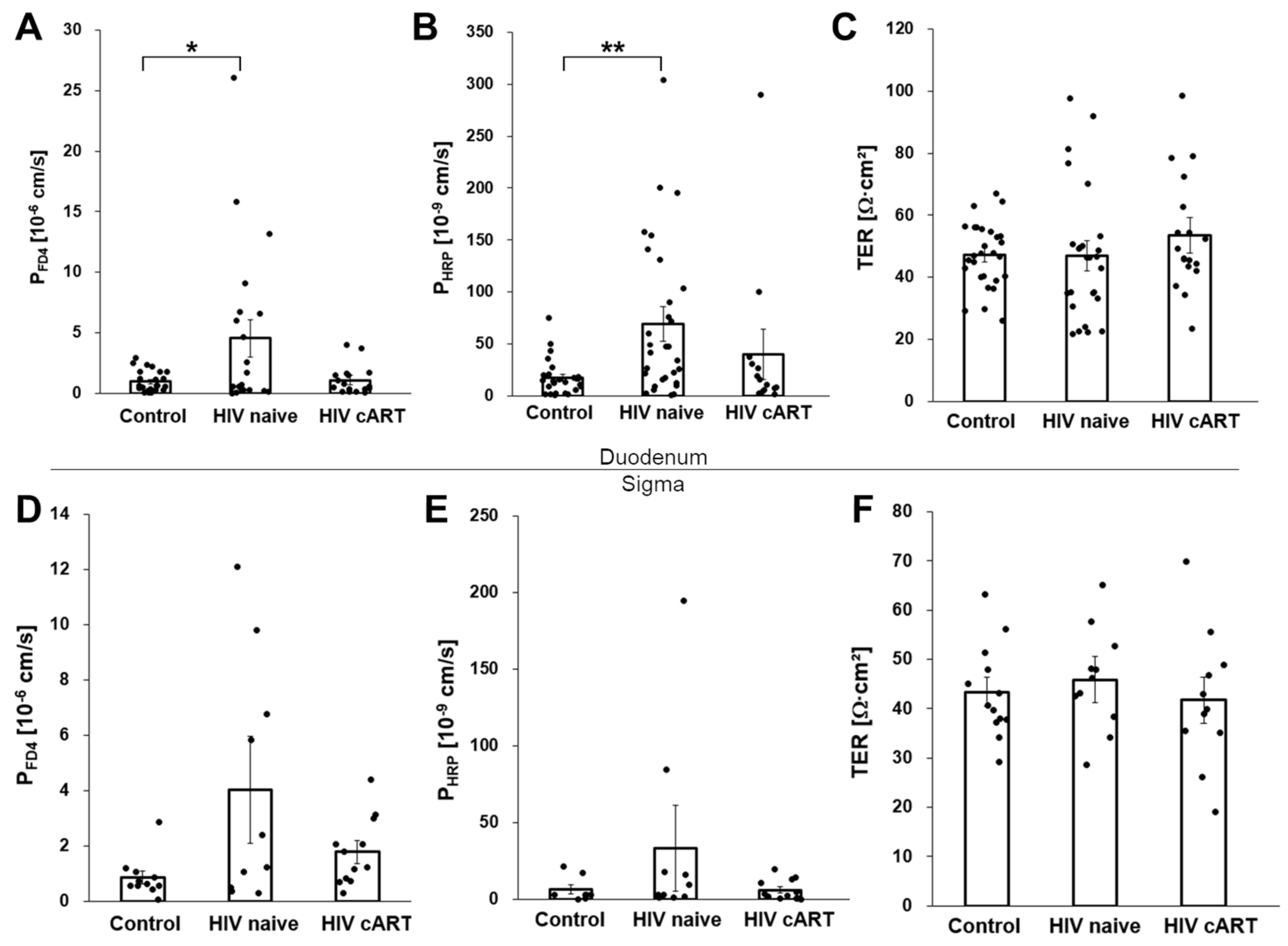
3.1. Increased Macromolecule Translocation in Untreated HIV-Infection
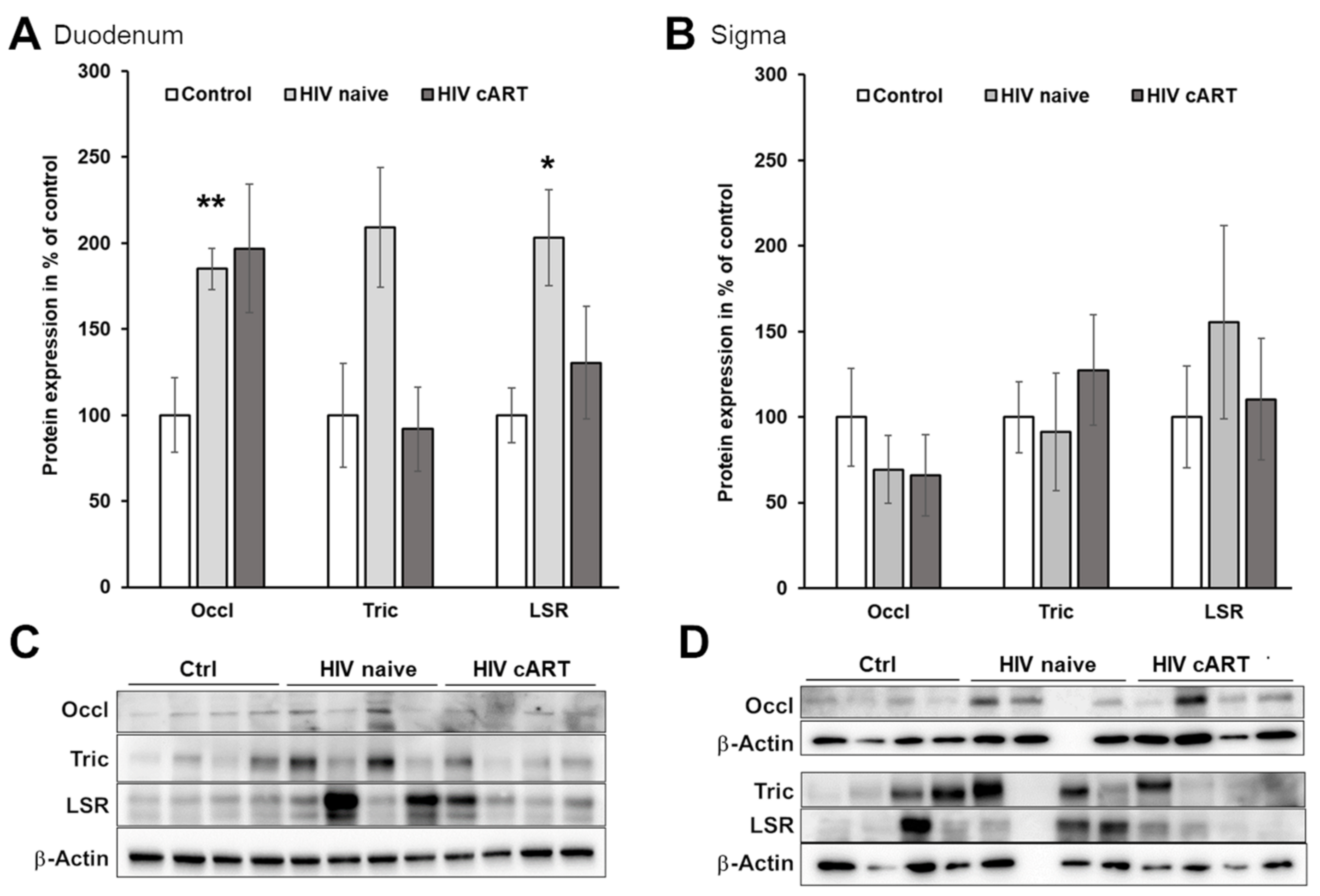
3.2. The Paracellular Pathway as a Potential Route for Macromolecule Translocation
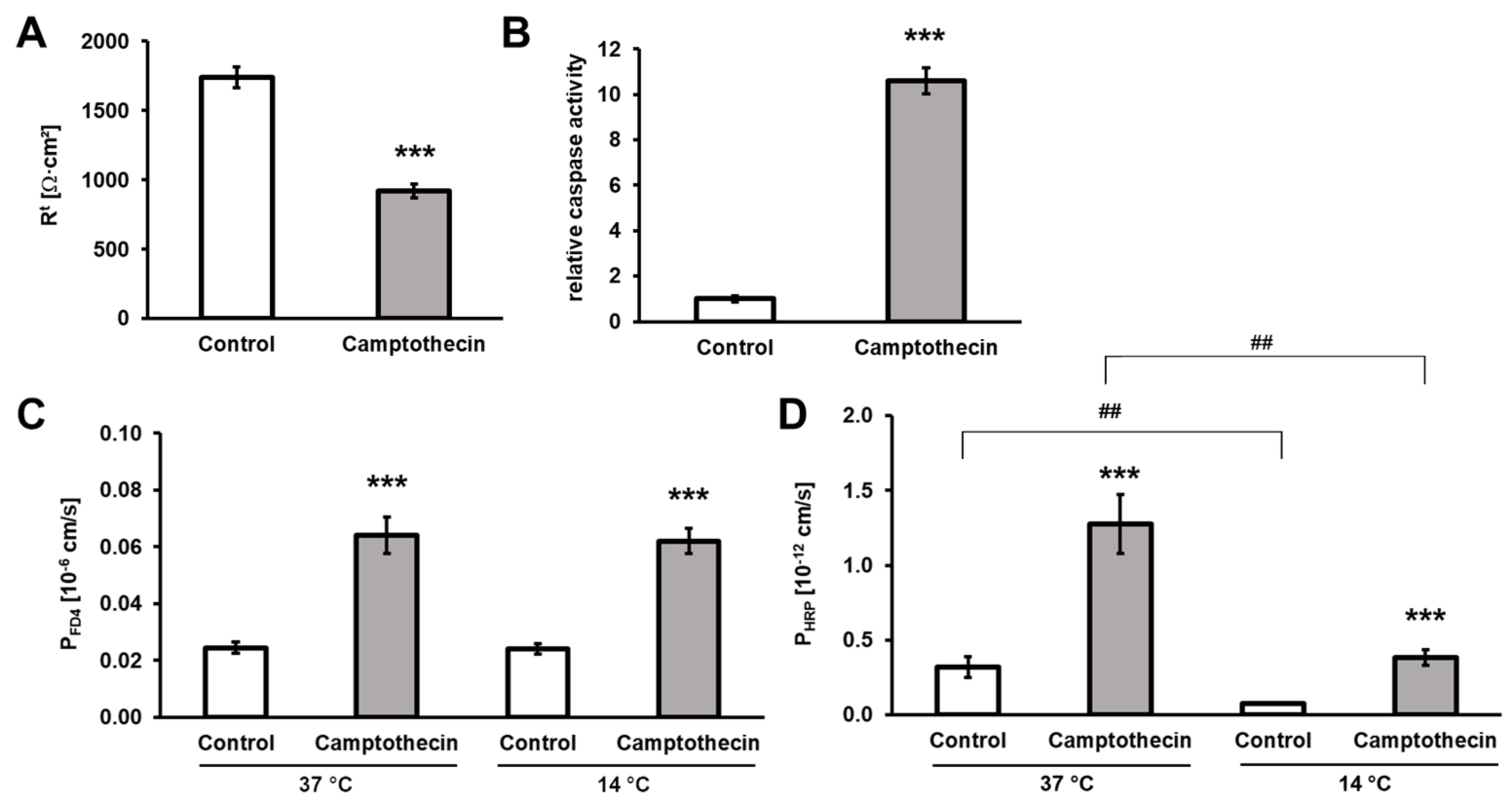
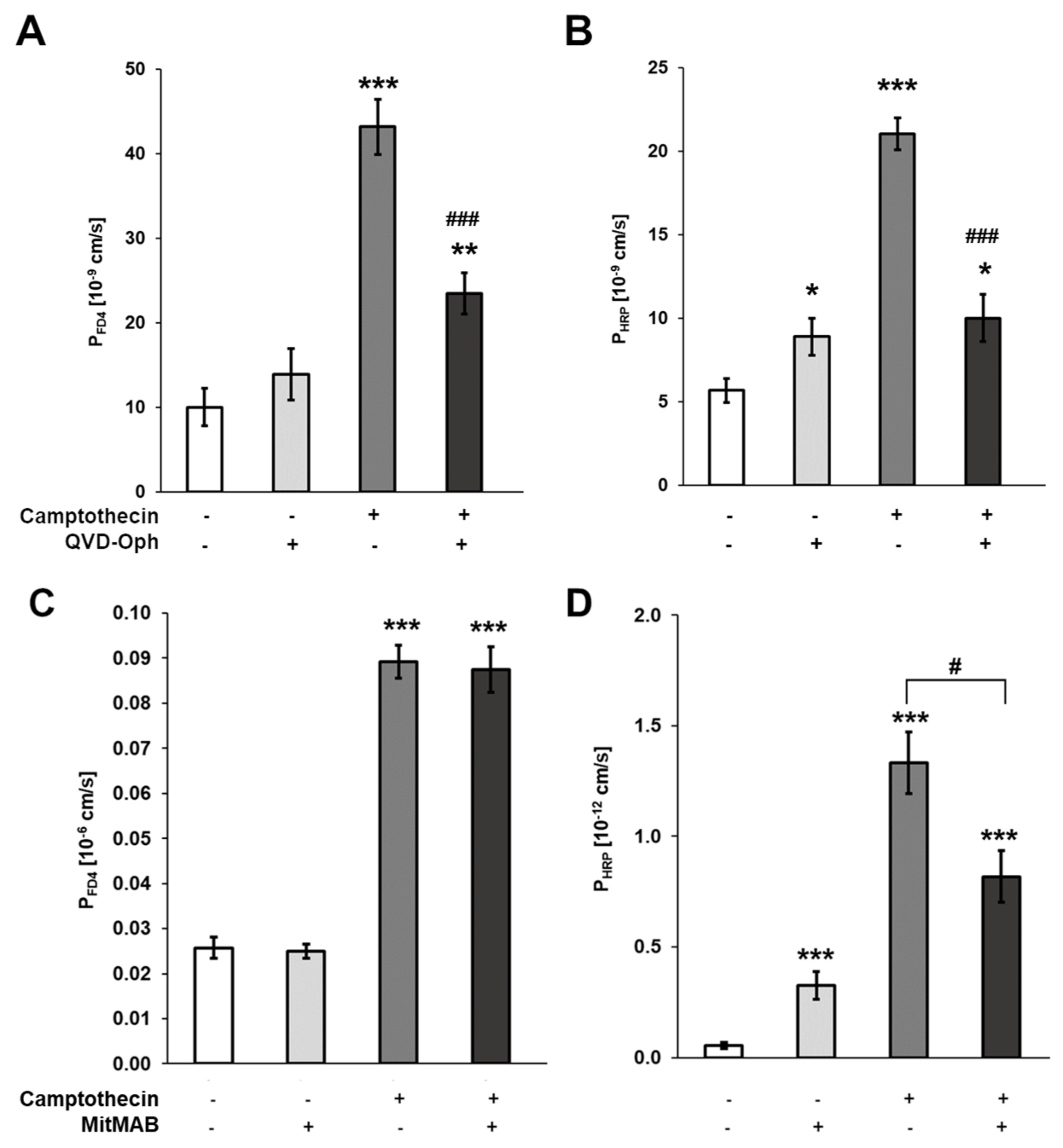
3.3. Contribution of Enterocyte Apoptosis to Macromolecule Translocation
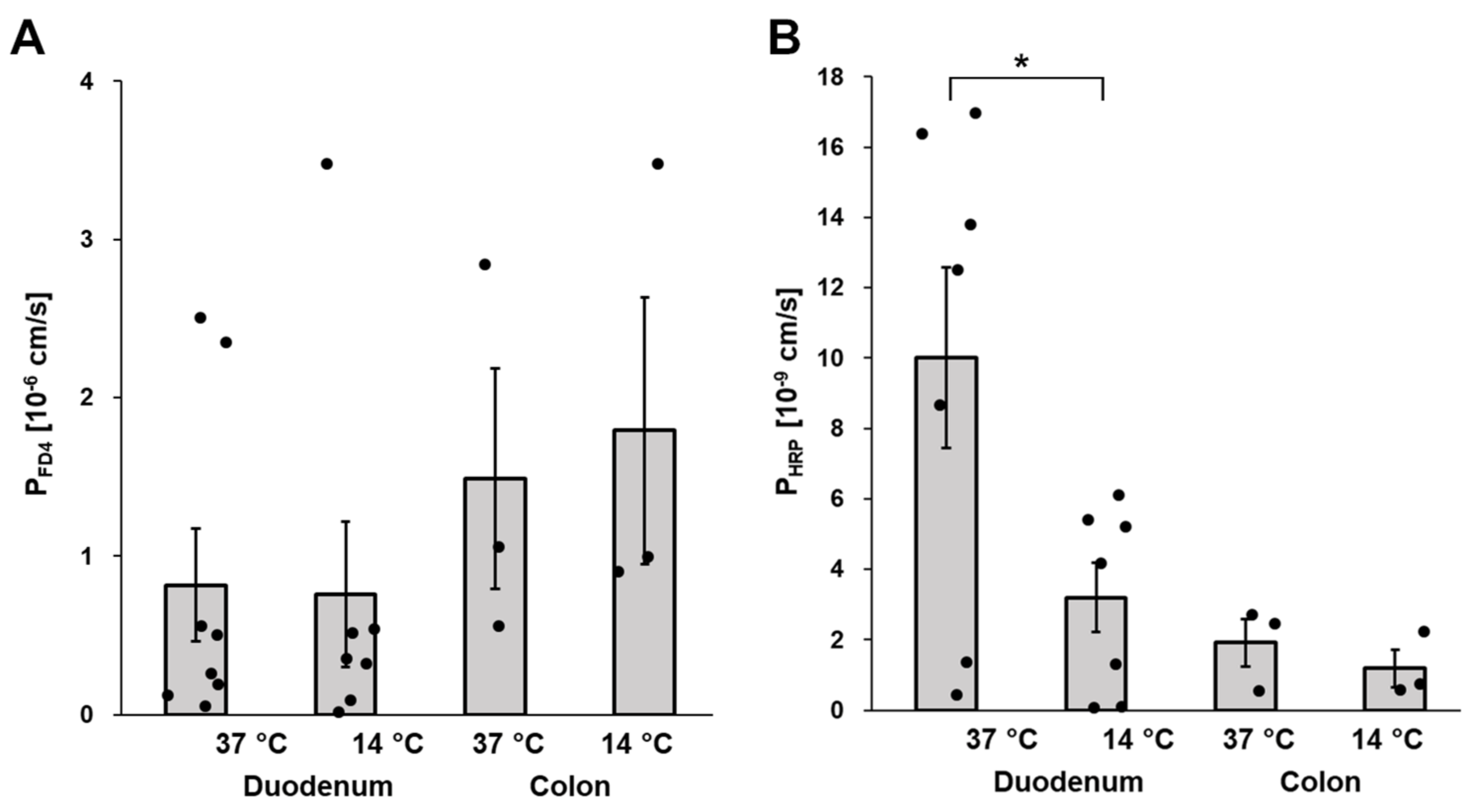
3.4. Macromolecule Translocation by Transcytosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ancuta, P.; Kamat, A.; Kunstman, K.J.; Kim, E.Y.; Autissier, P.; Wurcel, A.; Zaman, T.; Stone, D.; Mefford, M.; Morgello, S.; et al. Microbial translocation is associated with increased monocyte activation and dementia in AIDS patients. PLoS ONE 2008, 3, e2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 2006, 12, 1365–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, G.; Bellistri, G.M.; Borghi, E.; Tincati, C.; Ferramosca, S.; La Francesca, M.; Morace, G.; Gori, A.; Monforte, A.D. Microbial translocation is associated with sustained failure in CD4+ T-cell reconstitution in HIV-infected patients on long-term highly active antiretroviral therapy. AIDS 2008, 22, 2035–2038. [Google Scholar] [CrossRef] [PubMed]
- Cassol, E.; Malfeld, S.; Mahasha, P.; van der Merwe, S.; Cassol, S.; Seebregts, C.; Alfano, M.; Poli, G.; Rossouw, T. Persistent microbial translocation and immune activation in HIV-1-infected South Africans receiving combination antiretroviral therapy. J. Infect. Dis. 2010, 202, 723–733. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Lederman, M.M.; Hunt, P.; Sieg, S.F.; Haley, K.; Rodriguez, B.; Landay, A.; Martin, J.; Sinclair, E.; Asher, A.I.; et al. Plasma levels of bacterial DNA correlate with immune activation and the magnitude of immune restoration in persons with antiretroviral-treated HIV infection. J. Infect. Dis. 2009, 199, 1177–1185. [Google Scholar] [CrossRef]
- Morris, A.; Hillenbrand, M.; Finkelman, M.; George, M.P.; Singh, V.; Kessinger, C.; Lucht, L.; Busch, M.; McMahon, D.; Weinman, R.; et al. Serum (1-->3)-beta-D-glucan levels in HIV-infected individuals are associated with immunosuppression, inflammation, and cardiopulmonary function. J. Acquir. Immune Defic. Syndr. 2012, 61, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Brenchley, J.M.; Price, D.A.; Douek, D.C. HIV disease: Fallout from a mucosal catastrophe? Nat. Immunol. 2006, 7, 235–239. [Google Scholar] [CrossRef]
- Klatt, N.R.; Funderburg, N.T.; Brenchley, J.M. Microbial translocation, immune activation, and HIV disease. Trends Microbiol. 2013, 21, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Somsouk, M.; Estes, J.D.; Deleage, C.; Dunham, R.M.; Albright, R.; Inadomi, J.M.; Martin, J.N.; Deeks, S.G.; McCune, J.M.; Hunt, P.W. Gut epithelial barrier and systemic inflammation during chronic HIV infection. AIDS 2015, 29, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Chitre, A.S.; Kattah, M.G.; Rosli, Y.Y.; Pao, M.; Deswal, M.; Deeks, S.G.; Hunt, P.W.; Abdel-Mohsen, M.; Montaner, L.J.; Kim, C.C.; et al. A20 upregulation during treated HIV disease is associated with intestinal epithelial cell recovery and function. PLoS Pathog. 2018, 14, e1006806. [Google Scholar] [CrossRef] [Green Version]
- Tugizov, S. Human immunodeficiency virus-associated disruption of mucosal barriers and its role in HIV transmission and pathogenesis of HIV/AIDS disease. Tissue Barriers 2016, 4, e1159276. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, S.; George, M.D.; Reay, E.; Guadalupe, M.; Flamm, J.; Prindiville, T.; Dandekar, S. Rapid onset of intestinal epithelial barrier dysfunction in primary human immunodeficiency virus infection is driven by an imbalance between immune response and mucosal repair and regeneration. J. Virol. 2008, 82, 538–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allam, O.; Samarani, S.; Mehraj, V.; Jenabian, M.A.; Tremblay, C.; Routy, J.P.; Amre, D.; Ahmad, A. HIV induces production of IL-18 from intestinal epithelial cells that increases intestinal permeability and microbial translocation. PLoS ONE 2018, 13, e0194185. [Google Scholar] [CrossRef] [PubMed]
- Nazli, A.; Chan, O.; Dobson-Belaire, W.N.; Ouellet, M.; Tremblay, M.J.; Gray-Owen, S.D.; Arsenault, A.L.; Kaushic, C. Exposure to HIV-1 directly impairs mucosal epithelial barrier integrity allowing microbial translocation. PLoS Pathog. 2010, 6, e1000852. [Google Scholar] [CrossRef]
- Chung, C.Y.; Alden, S.L.; Funderburg, N.T.; Fu, P.; Levine, A.D. Progressive proximal-to-distal reduction in expression of the tight junction complex in colonic epithelium of virally-suppressed HIV+ individuals. PLoS Pathog. 2014, 10, e1004198. [Google Scholar] [CrossRef]
- Hofer, U.; Schlaepfer, E.; Baenziger, S.; Nischang, M.; Regenass, S.; Schwendener, R.; Kempf, W.; Nadal, D.; Speck, R.F. Inadequate clearance of translocated bacterial products in HIV-infected humanized mice. PLoS Pathog. 2010, 6, e1000867. [Google Scholar] [CrossRef]
- Guerville, M.; Boudry, G. Gastrointestinal and hepatic mechanisms limiting entry and dissemination of lipopolysaccharide into the systemic circulation. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G1–G15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoultz, I.; Keita, A.V. The Intestinal Barrier and Current Techniques for the Assessment of Gut Permeability. Cells 2020, 9, 1909. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [Green Version]
- Al-Sadi, R.; Khatib, K.; Guo, S.; Ye, D.; Youssef, M.; Ma, T. Occludin regulates macromolecule flux across the intestinal epithelial tight junction barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G1054–G1064. [Google Scholar] [CrossRef] [Green Version]
- Richter, J.F.; Hildner, M.; Schmauder, R.; Turner, J.R.; Schumann, M.; Reiche, J. Occludin knockdown is not sufficient to induce transepithelial macromolecule passage. Tissue Barriers 2019, 7, 1612661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cording, J.; Arslan, B.; Staat, C.; Dithmer, S.; Krug, S.M.; Krüger, A.; Berndt, P.; Günther, R.; Winkler, L.; Blasig, I.E.; et al. Trictide, a tricellulin-derived peptide to overcome cellular barriers. Ann. N. Y. Acad. Sci. 2017, 1405, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.M.; Amasheh, S.; Richter, J.F.; Milatz, S.; Gunzel, D.; Westphal, J.K.; Huber, O.; Schulzke, J.D.; Fromm, M. Tricellulin forms a barrier to macromolecules in tricellular tight junctions without affecting ion permeability. Mol. Biol. Cell 2009, 20, 3713–3724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krug, S.M.; Hayaishi, T.; Iguchi, D.; Watari, A.; Takahashi, A.; Fromm, M.; Nagahama, M.; Takeda, H.; Okada, Y.; Sawasaki, T.; et al. Angubindin-1, a novel paracellular absorption enhancer acting at the tricellular tight junction. J. Control. Release Off. J. Control. Release Soc. 2017, 260, 1–11. [Google Scholar] [CrossRef]
- Epple, H.J.; Schneider, T.; Troeger, H.; Kunkel, D.; Allers, K.; Moos, V.; Amasheh, M.; Loddenkemper, C.; Fromm, M.; Zeitz, M.; et al. Impairment of the intestinal barrier is evident in untreated but absent in suppressively treated HIV-infected patients. Gut 2009, 58, 220–227. [Google Scholar] [CrossRef]
- Epple, H.J.; Kreusel, K.M.; Hanski, C.; Schulzke, J.D.; Riecken, E.O.; Fromm, M. Differential stimulation of intestinal mucin secretion by cholera toxin and carbachol. Pflug. Arch. 1997, 433, 638–647. [Google Scholar] [CrossRef]
- Maidana, D.E.; Tsoka, P.; Tian, B.; Dib, B.; Matsumoto, H.; Kataoka, K.; Lin, H.; Miller, J.W.; Vavvas, D.G. A Novel ImageJ Macro for Automated Cell Death Quantitation in the Retina. Investig. Ophthalmol. Vis. Sci. 2015, 56, 6701–6708. [Google Scholar] [CrossRef] [Green Version]
- Kuller, L.H.; Tracy, R.; Belloso, W.; De Wit, S.; Drummond, F.; Lane, H.C.; Ledergerber, B.; Lundgren, J.; Neuhaus, J.; Nixon, D.; et al. Inflammatory and coagulation biomarkers and mortality in patients with HIV infection. PLoS Med. 2008, 5, e203. [Google Scholar] [CrossRef]
- Masuda, S.; Oda, Y.; Sasaki, H.; Ikenouchi, J.; Higashi, T.; Akashi, M.; Nishi, E.; Furuse, M. LSR defines cell corners for tricellular tight junction formation in epithelial cells. J. Cell Sci. 2011, 124, 548–555. [Google Scholar] [CrossRef] [Green Version]
- Kitajiri, S.; Katsuno, T.; Sasaki, H.; Ito, J.; Furuse, M.; Tsukita, S. Deafness in occludin-deficient mice with dislocation of tricellulin and progressive apoptosis of the hair cells. Biol. Open 2014, 3, 759–766. [Google Scholar] [CrossRef] [Green Version]
- Epple, H.J.; Allers, K.; Troger, H.; Kuhl, A.; Erben, U.; Fromm, M.; Zeitz, M.; Loddenkemper, C.; Schulzke, J.D.; Schneider, T. Acute HIV infection induces mucosal infiltration with CD4+ and CD8+ T cells, epithelial apoptosis, and a mucosal barrier defect. Gastroenterology 2010, 139, 1289–1300. [Google Scholar] [CrossRef] [PubMed]
- Tincati, C.; Douek, D.C.; Marchetti, G. Gut barrier structure, mucosal immunity and intestinal microbiota in the pathogenesis and treatment of HIV infection. AIDS Res. Ther. 2016, 13, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soderholm, J.D.; Streutker, C.; Yang, P.C.; Paterson, C.; Singh, P.K.; McKay, D.M.; Sherman, P.M.; Croitoru, K.; Perdue, M.H. Increased epithelial uptake of protein antigens in the ileum of Crohn’s disease mediated by tumour necrosis factor alpha. Gut 2004, 53, 1817–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hundorfean, G.; Zimmer, K.P.; Strobel, S.; Gebert, A.; Ludwig, D.; Buning, J. Luminal antigens access late endosomes of intestinal epithelial cells enriched in MHC I and MHC II molecules: In vivo study in Crohn’s ileitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G798–G808. [Google Scholar] [CrossRef] [Green Version]
- Tomita, M.; Ohkubo, R.; Hayashi, M. Lipopolysaccharide transport system across colonic epithelial cells in normal and infective rat. Drug Metab. Pharmacokinet. 2004, 19, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Kotler, D.P. HIV enteropathy and aging: Gastrointestinal immunity, mucosal epithelial barrier, and microbial translocation. Curr. Opin. HIV AIDS 2014, 9, 309–316. [Google Scholar] [CrossRef]
- Buret, A.G.; Bhargava, A. Modulatory mechanisms of enterocyte apoptosis by viral, bacterial and parasitic pathogens. Crit. Rev. Microbiol. 2014, 40, 1–17. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Controls | HIV Naive | HIV cART | |
|---|---|---|---|
| Duodenum | |||
| N | 20 | 19 | 10 |
| Sex (m/w) | 10/10 | 14/5 | 8/1 |
| Age (years) | 49.5 (±2.3) | 43.5 (±2.5) | 57.1 (±1.7) |
| CD4+ T cells (cells/µL) | ND | 239 (±20) | 371 (±1.5) |
| Viral load (log10 copies/mL) | NA | 5.3 (±2.5) | <LOD |
| Colon | |||
| N | 10 | 5 | 9 |
| Sex (m/w) | 7/6 | 4/1 | 9/0 |
| Age (years) | 51.3 (±2.7) | 31.5 (±2.4) | 64.6 (±0.9) |
| CD4+ T cells (cells/µL) | ND | 130 (±13) | 563 (±12) |
| Viral load (log10 copies/mL) | NA | 5.2 (±2.7) | <LOD |
| n | m | Apoptosis % | SEM | p (to Ctrl) | p (to HIV) | |
|---|---|---|---|---|---|---|
| Duodenum | ||||||
| Ctrl | 14 | 33,990 | 1.52 | 0.23 | - | - |
| HIV naive | 10 | 24,458 | 3.25 | 0.70 | 0.0278 | - |
| HIV cART | 10 | 21,357 | 2.07 | 0.25 | 0.1249 | 0.1298 |
| Colon | ||||||
| Ctrl | 20 | 37,391 | 2.11 | 0.43 | - | - |
| HIV naive | 19 | 48,837 | 4.13 | 0.68 | 0.0310 | - |
| HIV cART | 17 | 39,882 | 1.87 | 0.47 | 0.7085 | 0.0115 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krug, S.M.; Grünhagen, C.; Allers, K.; Bojarski, C.; Seybold, J.; Schneider, T.; Schulzke, J.-D.; Epple, H.-J. Macromolecule Translocation across the Intestinal Mucosa of HIV-Infected Patients by Transcytosis and through Apoptotic Leaks. Cells 2023, 12, 1887. https://doi.org/10.3390/cells12141887
Krug SM, Grünhagen C, Allers K, Bojarski C, Seybold J, Schneider T, Schulzke J-D, Epple H-J. Macromolecule Translocation across the Intestinal Mucosa of HIV-Infected Patients by Transcytosis and through Apoptotic Leaks. Cells. 2023; 12(14):1887. https://doi.org/10.3390/cells12141887
Chicago/Turabian StyleKrug, Susanne M., Carolin Grünhagen, Kristina Allers, Christian Bojarski, Joachim Seybold, Thomas Schneider, Jörg-Dieter Schulzke, and Hans-Jörg Epple. 2023. "Macromolecule Translocation across the Intestinal Mucosa of HIV-Infected Patients by Transcytosis and through Apoptotic Leaks" Cells 12, no. 14: 1887. https://doi.org/10.3390/cells12141887
APA StyleKrug, S. M., Grünhagen, C., Allers, K., Bojarski, C., Seybold, J., Schneider, T., Schulzke, J. -D., & Epple, H. -J. (2023). Macromolecule Translocation across the Intestinal Mucosa of HIV-Infected Patients by Transcytosis and through Apoptotic Leaks. Cells, 12(14), 1887. https://doi.org/10.3390/cells12141887