
The Melanocortin System in Inflammatory Bowel Diseases: Insights into Its Mechanisms and Therapeutic Potentials

 ,
,  , , , ,
, , , ,  , and
, and 
Abstract
:1. Introduction
2. α-MSH as a Key Melanocortin in the Modulation of Inflammatory Processes
3. The Receptors of the Melanocortin System: The Basis for Their Role in Peripheral and Systemic Inflammation
3.1. MC1R
3.2. MC2R
3.3. MC3R
3.4. MC4R
3.5. MC5R
4. The Role of the Melanocortin System in IBD: What Evidence?
- MEDLINE: (melanocortin or MC1R or MC2R or MC3R or MC4R or MC5R or α-MSH or β-MSH or alpha MSH or beta MSH or KPV or MSH or melanocyte-stimulating hormone) and (Crohn’s disease or ulcerative colitis or inflammatory bowel disease or IBD or DSS or DNBS or TNBS);
- EMBASE: (“Melanocortin” or “MC1R” or “MC2R” or “MC3R” or “MC4R” or “MC5R” or “α-MSH” or “β-MSH” or “Alpha MSH” or “Beta MSH” or “KPV” or “MSH” or “melanocyte-stimulating hormone”) and (“Inflammatory bowel disease” or “Crohn” or “Ulcerative colitis” or “IBD” or “DSS” or “TNBS” or “DNBS”);
- Web of Science: (all = (melanocortin) or all = (MC1R) or all = (MC2R) or all = (MC3R) or all = (MC4R) or all = (MC5R) or all = (α-MSH) and all = (β-MSH) or all = (alpha MSH) or all = (beta MSH) or all = (KPV) or all = (MSH) or all = (melanocyte-stimulating hormone)) and (all = (inflammatory bowel disease) or all = (Crohn’s disease) or all = (ulcerative colitis) or all = (IBD) or all = (DSS) or all = (TNBS) or all = (DNBS)).
4.1. MC1R Mediates and Improves Intestinal Inflammation in Major Models of Experimental Colitis, and Some of Its Agonists Are Being Carefully Studied as Potential Therapeutic Agents in IBD
4.2. MC2R Is Involved in the Interaction between UVA and UVB and Murine DSS-Induced Colitis
4.3. Colic Expression of MC3R and MC4R Appears to Differ According to Disease Activity in IBD: Initial Experience and Scarce Evidence
4.4. Combining Recombinant Bacteria and α-MSH as a Strategy in Experimental Colitis
4.5. Exploiting the Similarity with KPV Tripeptide of the C-Terminal End of α-MSH: The Anti-Inflammatory Role of KPV-like Tripeptide KdPT
4.6. Not Only α-MSH: What Potential of β-MSH?
4.7. Prospects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Catania, A. The Melanocortin System in Leukocyte Biology. J. Leukoc. Biol. 2007, 81, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holder, J.R.; Haskell-Luevano, C. Melanocortin Ligands: 30 Years of Structure-Activity Relationship (SAR) Studies. Med. Res. Rev. 2004, 24, 325–356. [Google Scholar] [CrossRef]
- Yang, Y.; Harmon, C.M. Molecular Signatures of Human Melanocortin Receptors for Ligand Binding and Signaling. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 2436–2447. [Google Scholar] [CrossRef]
- Videira, I.F.d.S.; Moura, D.F.L.; Magina, S. Mechanisms Regulating Melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef] [Green Version]
- Lam, C.; Getting, S. Melanocortin Receptor Type 3 as a Potential Target for Anti-Inflammatory Therapy. CDTIA 2004, 3, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Catania, A.; Lonati, C.; Sordi, A.; Carlin, A.; Leonardi, P.; Gatti, S. The Melanocortin System in Control of Inflammation. Sci. World J. 2010, 10, 1840–1853. [Google Scholar] [CrossRef] [Green Version]
- Brzoska, T.; Luger, T.A.; Maaser, C.; Abels, C.; Böhm, M. Alpha-Melanocyte-Stimulating Hormone and Related Tripeptides: Biochemistry, Antiinflammatory and Protective Effects in Vitro and in Vivo, and Future Perspectives for the Treatment of Immune-Mediated Inflammatory Diseases. Endocr. Rev. 2008, 29, 581–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barberio, B.; Zamani, M.; Black, C.J.; Savarino, E.V.; Ford, A.C. Prevalence of Symptoms of Anxiety and Depression in Patients with Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis. Lancet Gastroenterol. Hepatol. 2021, 6, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Spina, A.; Mazzarella, C.; Dallio, M.; Romeo, M.; Pellegrino, R.; Durante, T.; Romano, M.; Loguercio, C.; Di Mauro, M.; Federico, A.; et al. The Lesson from the First Italian Lockdown: Impacts on Anxiety and Depressive Symptoms and Sleep Quality in Patients with Remission of Inflammatory Bowel Disease. Rev. Recent. Clin. Trials 2022, 17, 109–119. [Google Scholar] [CrossRef]
- Luo, J.; Xu, Z.; Noordam, R.; van Heemst, D.; Li-Gao, R. Depression and Inflammatory Bowel Disease: A Bidirectional Two-Sample Mendelian Randomization Study. J. Crohn’s Colitis 2022, 16, 633–642. [Google Scholar] [CrossRef]
- Chang, J.T. Pathophysiology of Inflammatory Bowel Diseases. N. Engl. J. Med. 2020, 383, 2652–2664. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F.; Pettersson, S.; Meyer zum Büschenfelde, K.H.; Strober, W. Local Administration of Antisense Phosphorothioate Oligonucleotides to the P65 Subunit of NF-Kappa B Abrogates Established Experimental Colitis in Mice. Nat. Med. 1996, 2, 998–1004. [Google Scholar] [CrossRef] [PubMed]
- Rogler, G.; Brand, K.; Vogl, D.; Page, S.; Hofmeister, R.; Andus, T.; Knuechel, R.; Baeuerle, P.A.; Schölmerich, J.; Gross, V. Nuclear Factor KappaB Is Activated in Macrophages and Epithelial Cells of Inflamed Intestinal Mucosa. Gastroenterology 1998, 115, 357–369. [Google Scholar] [CrossRef]
- Pallone, F.; Monteleone, G. Mechanisms of Tissue Damage in Inflammatory Bowel Disease. Curr. Opin. Gastroenterol. 2001, 17, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Raine, T.; Bonovas, S.; Burisch, J.; Kucharzik, T.; Adamina, M.; Annese, V.; Bachmann, O.; Bettenworth, D.; Chaparro-Sanchez, M.; Czuber-Dochan, W.; et al. ECCO Guidelines on Therapeutics in Ulcerative Colitis: Medical Treatment. J. Crohn’s Colitis 2022, 16, 2–17. [Google Scholar] [CrossRef]
- Harbord, M.; Annese, V.; Vavricka, S.R.; Allez, M.; Barreiro-de Acosta, M.; Boberg, K.M.; Burisch, J.; De Vos, M.; De Vries, A.-M.; Dick, A.D.; et al. The First European Evidence-Based Consensus on Extra-Intestinal Manifestations in Inflammatory Bowel Disease. J. Crohn’s Colitis 2016, 10, 239–254. [Google Scholar] [CrossRef]
- Torres, J.; Bonovas, S.; Doherty, G.; Kucharzik, T.; Gisbert, J.P.; Raine, T.; Adamina, M.; Armuzzi, A.; Bachmann, O.; Bager, P.; et al. ECCO Guidelines on Therapeutics in Crohn’s Disease: Medical Treatment. J. Crohn’s Colitis 2020, 14, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Kawai, T.; Akira, S. Signaling to NF-KappaB by Toll-like Receptors. Trends Mol. Med. 2007, 13, 460–469. [Google Scholar] [CrossRef]
- Lawrence, T. The Nuclear Factor NF-KappaB Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Thoma, A.; Lightfoot, A.P. NF-KB and Inflammatory Cytokine Signalling: Role in Skeletal Muscle Atrophy. Adv. Exp. Med. Biol. 2018, 1088, 267–279. [Google Scholar] [CrossRef]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB System. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Taylor, B.; Ourthiague, D.R.; Hoffmann, A. Distinct Single-Cell Signaling Characteristics Are Conferred by the MyD88 and TRIF Pathways during TLR4 Activation. Sci. Signal. 2015, 8, ra69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.-S.; Hayasaka, S.; Zhang, X.-Y.; Chi, Z.-L.; Hayasaka, Y. Effect of Alpha-Melanocyte-Stimulating Hormone on Interleukin 8 and Monocyte Chemotactic Protein 1 Expression in a Human Retinal Pigment Epithelial Cell Line. Ophthalmic Res. 2005, 37, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Haycock, J.W.; Wagner, M.; Morandini, R.; Ghanem, G.; Rennie, I.G.; Mac Neil, S. Alpha-Melanocyte-Stimulating Hormone Inhibits NF-KappaB Activation in Human Melanocytes and Melanoma Cells. J. Investig. Dermatol. 1999, 113, 560–566. [Google Scholar] [CrossRef]
- Buggy, J.J. Binding of Alpha-Melanocyte-Stimulating Hormone to Its G-Protein-Coupled Receptor on B-Lymphocytes Activates the Jak/STAT Pathway. Biochem. J. 1998, 331 Pt 1, 211–216. [Google Scholar] [CrossRef]
- Chang, S.H.; Jung, E.J.; Lim, D.G.; Park, Y.H.; Wee, Y.M.; Kim, J.H.; Kim, Y.H.; Choi, M.Y.; Koo, S.K.; Choi, K.D.; et al. Anti-Inflammatory Action of Alpha-Melanocyte Stimulating Hormone (Alpha-MSH) in Anti-CD3/CD28-Mediated Spleen and CD4(+)CD25(-) T Cells and a Partial Participation of IL-10. Immunol. Lett. 2008, 118, 44–48. [Google Scholar] [CrossRef]
- Nishida, T.; Taylor, A.W. Specific Aqueous Humor Factors Induce Activation of Regulatory T Cells. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2268–2274. [Google Scholar]
- Taylor, A.W.; Streilein, J.W.; Cousins, S.W. Alpha-Melanocyte-Stimulating Hormone Suppresses Antigen-Stimulated T Cell Production of Gamma-Interferon. Neuroimmunomodulation 1994, 1, 188–194. [Google Scholar] [CrossRef]
- Rodari, M.M.; Cerf-Bensussan, N.; Parlato, M. Dysregulation of the Immune Response in TGF-β Signalopathies. Front. Immunol. 2022, 13, 1066375. [Google Scholar] [CrossRef]
- Auriemma, M.; Brzoska, T.; Klenner, L.; Kupas, V.; Goerge, T.; Voskort, M.; Zhao, Z.; Sparwasser, T.; Luger, T.A.; Loser, K. α-MSH-Stimulated Tolerogenic Dendritic Cells Induce Functional Regulatory T Cells and Ameliorate Ongoing Skin Inflammation. J. Investig. Dermatol. 2012, 132, 1814–1824. [Google Scholar] [CrossRef] [Green Version]
- Saporiti, F.; Piacentini, L.; Alfieri, V.; Bono, E.; Ferrari, F.; Chiesa, M.; Colombo, G.I. Melanocortin-1 Receptor Positively Regulates Human Artery Endothelial Cell Migration. Cell. Physiol. Biochem. 2019, 52, 1339–1360. [Google Scholar] [CrossRef] [PubMed]
- Seaton, M.E.; Parent, B.A.; Sood, R.F.; Wurfel, M.M.; Muffley, L.A.; O’Keefe, G.E.; Gibran, N.S. Melanocortin-1 Receptor Polymorphisms and the Risk of Complicated Sepsis After Trauma: A Candidate Gene Association Study. Shock 2017, 47, 79–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, S.; Maisto, R.; Gesualdo, C.; Trotta, M.C.; Ferraraccio, F.; Kaneva, M.K.; Getting, S.J.; Surace, E.; Testa, F.; Simonelli, F.; et al. Corrigendum to “Activation of Melanocortin Receptors MC1 and MC5 Attenuates Retinal Damage in Experimental Diabetic Retinopathy”. Mediat. Inflamm. 2021, 2021, 1–2. [Google Scholar] [CrossRef]
- Maisto, R.; Gesualdo, C.; Trotta, M.C.; Grieco, P.; Testa, F.; Simonelli, F.; Barcia, J.M.; D’Amico, M.; Di Filippo, C.; Rossi, S. Melanocortin Receptor Agonists MCR1-5 Protect Photoreceptors from High-Glucose Damage and Restore Antioxidant Enzymes in Primary Retinal Cell Culture. J. Cell. Mol. Med. 2017, 21, 968–974. [Google Scholar] [CrossRef]
- Mykicki, N.; Herrmann, A.M.; Schwab, N.; Deenen, R.; Sparwasser, T.; Limmer, A.; Wachsmuth, L.; Klotz, L.; Köhrer, K.; Faber, C.; et al. Melanocortin-1 Receptor Activation Is Neuroprotective in Mouse Models of Neuroinflammatory Disease. Sci. Transl. Med. 2016, 8, 362ra146. [Google Scholar] [CrossRef] [Green Version]
- Catania, A. Neuroprotective Actions of Melanocortins: A Therapeutic Opportunity. Trends Neurosci. 2008, 31, 353–360. [Google Scholar] [CrossRef]
- Andersen, M.; Nagaev, I.; Meyer, M.K.; Nagaeva, O.; Wikberg, J.; Mincheva-Nilsson, L.; Andersen, G.N. Melanocortin 2, 3 and 4 Receptor Gene Expressions Are Downregulated in CD8+ T Cytotoxic Lymphocytes and CD19+ B Lymphocytes in Rheumatoid Arthritis Responding to TNF-α Inhibition. Scand. J. Immunol. 2017, 86, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Getting, S.J.; Perretti, M. MC3-R as a Novel Target for Antiinflammatory Therapy. Drug News Perspect. 2000, 13, 19–27. [Google Scholar] [PubMed]
- Getting, S.J.; Gibbs, L.; Clark, A.J.; Flower, R.J.; Perretti, M. POMC Gene-Derived Peptides Activate Melanocortin Type 3 Receptor on Murine Macrophages, Suppress Cytokine Release, and Inhibit Neutrophil Migration in Acute Experimental Inflammation. J. Immunol. 1999, 162, 7446–7453. [Google Scholar] [CrossRef] [PubMed]
- Getting, S.J.; Di Filippo, C.; Christian, H.C.; Lam, C.W.; Rossi, F.; D’Amico, M.; Perretti, M. MC-3 Receptor and the Inflammatory Mechanisms Activated in Acute Myocardial Infarct. J. Leukoc. Biol. 2004, 76, 845–853. [Google Scholar] [CrossRef]
- Getting, S.J.; Lam, C.W.; Chen, A.S.; Grieco, P.; Perretti, M. Melanocortin 3 Receptors Control Crystal-Induced Inflammation. FASEB J. 2006, 20, 2234–2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellacott, K.L.J.; Murphy, J.G.; Marks, D.L.; Cone, R.D. Obesity-Induced Inflammation in White Adipose Tissue Is Attenuated by Loss of Melanocortin-3 Receptor Signaling. Endocrinology 2007, 148, 6186–6194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuschillo, G.; Celentano, V.; Rottoli, M.; Sciaudone, G.; Gravina, A.G.; Pellegrino, R.; Marfella, R.; Romano, M.; Selvaggi, F.; Pellino, G. Influence of Diabetes Mellitus on Inflammatory Bowel Disease Course and Treatment Outcomes. A Systematic Review with Meta-Analysis. Dig. Liver Dis. 2023, 55, 580–586. [Google Scholar] [CrossRef]
- Mountjoy, K.G.; Mortrud, M.T.; Low, M.J.; Simerly, R.B.; Cone, R.D. Localization of the Melanocortin-4 Receptor (MC4-R) in Neuroendocrine and Autonomic Control Circuits in the Brain. Mol. Endocrinol. 1994, 8, 1298–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tracey, K.J. Physiology and Immunology of the Cholinergic Antiinflammatory Pathway. J. Clin. Investig. 2007, 117, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Zila, I.; Mokra, D.; Kopincova, J.; Kolomaznik, M.; Javorka, M.; Calkovska, A. Vagal-Immune Interactions Involved in Cholinergic Anti-Inflammatory Pathway. Physiol. Res. 2017, 66, S139–S145. [Google Scholar] [CrossRef]
- Giuliani, D.; Ottani, A.; Altavilla, D.; Bazzani, C.; Squadrito, F.; Guarini, S. Melanocortins and the Cholinergic Anti-Inflammatory Pathway. Adv. Exp. Med. Biol. 2010, 681, 71–87. [Google Scholar] [CrossRef]
- Tracey, K.J. The Inflammatory Reflex. Nature 2002, 420, 853–859. [Google Scholar] [CrossRef]
- Kamermans, A.; Verhoeven, T.; van Het Hof, B.; Koning, J.J.; Borghuis, L.; Witte, M.; van Horssen, J.; de Vries, H.E.; Rijnsburger, M. Setmelanotide, a Novel, Selective Melanocortin Receptor-4 Agonist Exerts Anti-Inflammatory Actions in Astrocytes and Promotes an Anti-Inflammatory Macrophage Phenotype. Front. Immunol. 2019, 10, 2312. [Google Scholar] [CrossRef]
- Flores-Bastías, O.; Gómez, G.I.; Orellana, J.A.; Karahanian, E. Activation of Melanocortin-4 Receptor by a Synthetic Agonist Inhibits Ethanolinduced Neuroinflammation in Rats. Curr. Pharm. Des. 2019, 25, 4799–4805. [Google Scholar] [CrossRef]
- Tao, Y.-X. The Melanocortin-4 Receptor: Physiology, Pharmacology, and Pathophysiology. Endocr. Rev. 2010, 31, 506–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huszar, D.; Lynch, C.A.; Fairchild-Huntress, V.; Dunmore, J.H.; Fang, Q.; Berkemeier, L.R.; Gu, W.; Kesterson, R.A.; Boston, B.A.; Cone, R.D.; et al. Targeted Disruption of the Melanocortin-4 Receptor Results in Obesity in Mice. Cell 1997, 88, 131–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loos, R.J.F.; Lindgren, C.M.; Li, S.; Wheeler, E.; Zhao, J.H.; Prokopenko, I.; Inouye, M.; Freathy, R.M.; Attwood, A.P.; Beckmann, J.S.; et al. Common Variants near MC4R Are Associated with Fat Mass, Weight and Risk of Obesity. Nat. Genet. 2008, 40, 768–775. [Google Scholar] [CrossRef] [Green Version]
- Cox, A.J.; West, N.P.; Cripps, A.W. Obesity, Inflammation, and the Gut Microbiota. Lancet Diabetes Endocrinol. 2015, 3, 207–215. [Google Scholar] [CrossRef]
- Asadi, A.; Shadab Mehr, N.; Mohamadi, M.H.; Shokri, F.; Heidary, M.; Sadeghifard, N.; Khoshnood, S. Obesity and Gut-Microbiota-Brain Axis: A Narrative Review. J. Clin. Lab. Anal. 2022, 36, e24420. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Kolb, R.; Sutterwala, F.S.; Zhang, W. Obesity and Cancer: Inflammation Bridges the Two. Curr. Opin. Pharmacol. 2016, 29, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Yeo, G.S.H.; Chao, D.H.M.; Siegert, A.-M.; Koerperich, Z.M.; Ericson, M.D.; Simonds, S.E.; Larson, C.M.; Luquet, S.; Clarke, I.; Sharma, S.; et al. The Melanocortin Pathway and Energy Homeostasis: From Discovery to Obesity Therapy. Mol. Metab. 2021, 48, 101206. [Google Scholar] [CrossRef]
- Haqq, A.M.; Chung, W.K.; Dollfus, H.; Haws, R.M.; Martos-Moreno, G.Á.; Poitou, C.; Yanovski, J.A.; Mittleman, R.S.; Yuan, G.; Forsythe, E.; et al. Efficacy and Safety of Setmelanotide, a Melanocortin-4 Receptor Agonist, in Patients with Bardet-Biedl Syndrome and Alström Syndrome: A Multicentre, Randomised, Double-Blind, Placebo-Controlled, Phase 3 Trial with an Open-Label Period. Lancet Diabetes Endocrinol. 2022, 10, 859–868. [Google Scholar] [CrossRef]
- Maisto, R.; Oltra, M.; Vidal-Gil, L.; Martínez-Gil, N.; Sancho-Pellúz, J.; Filippo, C.D.; Rossi, S.; D’Amico, M.; Barcia, J.M.; Romero, F.J. ARPE-19-Derived VEGF-Containing Exosomes Promote Neovascularization in HUVEC: The Role of the Melanocortin Receptor 5. Cell Cycle 2019, 18, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Trotta, M.C.; Maisto, R.; Alessio, N.; Hermenean, A.; D’Amico, M.; Di Filippo, C. The Melanocortin MC5R as a New Target for Treatment of High Glucose-Induced Hypertrophy of the Cardiac H9c2 Cells. Front. Physiol. 2018, 9, 1475. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Guan, X.; Zhou, R.; Gong, R. Melanocortin 5 Receptor Signaling Pathway in Health and Disease. Cell. Mol. Life Sci. 2020, 77, 3831–3840. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Biros, D.J.; Taylor, A.W. Injection of an Alpha-Melanocyte Stimulating Hormone Expression Plasmid Is Effective in Suppressing Experimental Autoimmune Uveitis. Int. Immunopharmacol. 2009, 9, 1079–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maaser, C. Crucial Role of the Melanocortin Receptor MC1R in Experimental Colitis. Gut 2006, 55, 1415–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
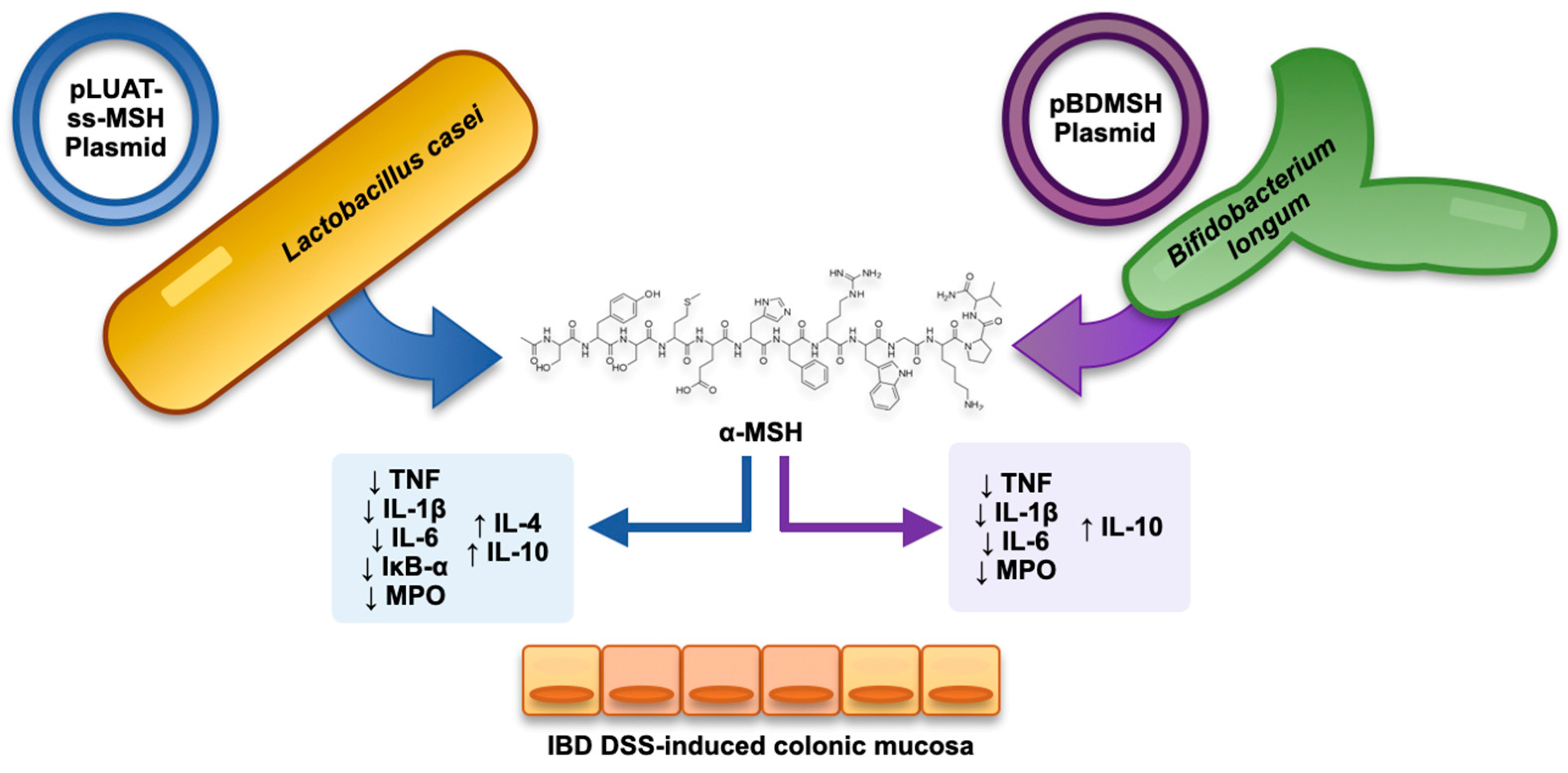
- Yoon, S.-W.; Lee, C.-H.; Kim, J.-Y.; Kim, J.-Y.; Sung, M.-H.; Poo, H. Lactobacillus Casei Secreting Alpha-MSH Induces the Therapeutic Effect on DSS-Induced Acute Colitis in Balb/c Mice. J. Microbiol. Biotechnol. 2008, 18, 1975–1983. [Google Scholar] [PubMed]
- Kannengiesser, K.; Maaser, C.; Heidemann, J.; Luegering, A.; Ross, M.; Brzoska, T.; Bohm, M.; Luger, T.A.; Domschke, W.; Kucharzik, T. Melanocortin-Derived Tripeptide KPV Has Anti-Inflammatory Potential in Murine Models of Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2008, 14, 324–331. [Google Scholar] [CrossRef]
- Bradamante, M.; Turčić, P.; Stambuk, N.; Konjevoda, P.; Aralica, G.; Alerić, I.; Kozmar, A. Cytoprotective Effects of β-Melanocortin in the Rat Gastrointestinal Tract. Molecules 2012, 17, 11680–11692. [Google Scholar] [CrossRef] [Green Version]
- Wei, P.; Yang, Y.; Ding, Q.; Li, X.; Sun, H.; Liu, Z.; Huang, J.; Gong, Y. Oral Delivery of Bifidobacterium Longum Expressing α-Melanocyte-Stimulating Hormone to Combat Ulcerative Colitis. J. Med. Microbiol. 2016, 65, 160–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiramoto, K.; Yamate, Y.; Sato, E.F. The Effects of Ultraviolet Eye Irradiation on Dextran Sodium Sulfate-Induced Ulcerative Colitis in Mice. Photochem. Photobiol. 2016, 92, 728–734. [Google Scholar] [CrossRef]
- Xiao, B.; Xu, Z.; Viennois, E.; Zhang, Y.; Zhang, Z.; Zhang, M.; Han, M.K.; Kang, Y.; Merlin, D. Orally Targeted Delivery of Tripeptide KPV via Hyaluronic Acid-Functionalized Nanoparticles Efficiently Alleviates Ulcerative Colitis. Mol. Ther. 2017, 25, 1628–1640. [Google Scholar] [CrossRef] [Green Version]
- Spana, C.; Taylor, A.W.; Yee, D.G.; Makhlina, M.; Yang, W.; Dodd, J. Probing the Role of Melanocortin Type 1 Receptor Agonists in Diverse Immunological Diseases. Front. Pharmacol. 2019, 9, 1535. [Google Scholar] [CrossRef]
- Gravina, A.G.; Panarese, I.; Trotta, M.C.; D’Amico, M.; Pellegrino, R.; Ferraraccio, F.; Galdiero, M.; Alfano, R.; Loguercio, C.; Romano, M.; et al. P072 Melanocortin MC3 and MC5 Receptors Expression Is Different According to Colonic Disease Activity in Crohn’s Disease and Ulcerative Colitis Biopsies: Results from an Immunohistochemical Observational Study. J. Crohn’s Colitis 2022, 16, i176–i177. [Google Scholar] [CrossRef]
- Dodd, J.; Jordan, R.; Makhlina, M.; Barnett, K.; Roffel, A.; Spana, C.; Obr, A.; Dhingra, P.; Kayne, P.S. A Novel Oral Formulation of the Melanocortin-1 Receptor Agonist PL8177 Resolves Inflammation in Preclinical Studies of Inflammatory Bowel Disease and Is Gut Restricted in Rats, Dogs, and Humans. Front. Immunol. 2023, 14, 1083333. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.L. The Hypothalamic-Pituitary-Adrenal Axis: A Brief History. Horm. Res. Paediatr. 2018, 89, 212–223. [Google Scholar] [CrossRef]
- Kojima, K.; Naruse, Y.; Iijima, N.; Wakabayashi, N.; Mitsufuji, S.; Ibata, Y.; Tanaka, M. HPA-Axis Responses during Experimental Colitis in the Rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R1348–R1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibolet, O.; Alper, R.; Ilan, Y.; Weidenfeld, J. Regulatory Role of the Pituitary-Adrenal Axis in Experimental Colitis: Effect of Adrenalectomy on the Clinical Course and the TH1/TH2 Immune Profile. Inflamm. Bowel Dis. 2005, 11, 1053–1059. [Google Scholar] [CrossRef]
- Rajora, N.; Boccoli, G.; Catania, A.; Lipton, J.M. Alpha-MSH Modulates Experimental Inflammatory Bowel Disease. Peptides 1997, 18, 381–385. [Google Scholar] [CrossRef]
- Oktar, B.K.; Ercan, F.; Yeğen, B.C.; Alican, I. The Effect of Alpha-Melanocyte Stimulating Hormone on Colonic Inflammation in the Rat. Peptides 2000, 21, 1271–1277. [Google Scholar] [CrossRef]
- San, T.; Oktar, B.K.; Salik, E.; Ercan, F.; Alican, I. The Effect of Alpha-Melanocyte Stimulating Hormone on Endotoxin-Induced Intestinal Injury. Peptides 2001, 22, 2077–2082. [Google Scholar] [CrossRef] [PubMed]
- Oktar, B.K.; Yüksel, M.; Alican, I. The Role of Cyclooxygenase Inhibition in the Effect of Alpha-Melanocyte-Stimulating Hormone on Reactive Oxygen Species Production by Rat Peritoneal Neutrophils. Prostaglandins Leukot. Essent. Fatty Acids 2004, 71, 1–5. [Google Scholar] [CrossRef]
- Hugot, J.P.; Chamaillard, M.; Zouali, H.; Lesage, S.; Cézard, J.P.; Belaiche, J.; Almer, S.; Tysk, C.; O’Morain, C.A.; Gassull, M.; et al. Association of NOD2 Leucine-Rich Repeat Variants with Susceptibility to Crohn’s Disease. Nature 2001, 411, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Ogura, Y.; Bonen, D.K.; Inohara, N.; Nicolae, D.L.; Chen, F.F.; Ramos, R.; Britton, H.; Moran, T.; Karaliuskas, R.; Duerr, R.H.; et al. A Frameshift Mutation in NOD2 Associated with Susceptibility to Crohn’s Disease. Nature 2001, 411, 603–606. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 Update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef] [PubMed]
- Luger, T.A.; Scholzen, T.E.; Brzoska, T.; Böhm, M. New Insights into the Functions of Alpha-MSH and Related Peptides in the Immune System. Ann. N. Y. Acad. Sci. 2003, 994, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Laroui, H.; Dalmasso, G.; Nguyen, H.T.T.; Yan, Y.; Sitaraman, S.V.; Merlin, D. Drug-Loaded Nanoparticles Targeted to the Colon with Polysaccharide Hydrogel Reduce Colitis in a Mouse Model. Gastroenterology 2010, 138, 843–853. [Google Scholar] [CrossRef]
- Dalmasso, G.; Charrier-Hisamuddin, L.; Nguyen, H.T.T.; Yan, Y.; Sitaraman, S.; Merlin, D. PepT1-Mediated Tripeptide KPV Uptake Reduces Intestinal Inflammation. Gastroenterology 2008, 134, 166–178. [Google Scholar] [CrossRef] [Green Version]
- Steel, A.; Nussberger, S.; Romero, M.F.; Boron, W.F.; Boyd, C.A.; Hediger, M.A. Stoichiometry and PH Dependence of the Rabbit Proton-Dependent Oligopeptide Transporter PepT1. J. Physiol. 1997, 498 Pt 3, 563–569. [Google Scholar] [CrossRef]
- Shen, H.; Smith, D.E.; Yang, T.; Huang, Y.G.; Schnermann, J.B.; Brosius, F.C. Localization of PEPT1 and PEPT2 Proton-Coupled Oligopeptide Transporter MRNA and Protein in Rat Kidney. Am. J. Physiol. 1999, 276, F658–F665. [Google Scholar] [CrossRef]
- Daniel, H. Molecular and Integrative Physiology of Intestinal Peptide Transport. Annu. Rev. Physiol. 2004, 66, 361–384. [Google Scholar] [CrossRef]
- Merlin, D.; Si-Tahar, M.; Sitaraman, S.V.; Eastburn, K.; Williams, I.; Liu, X.; Hediger, M.A.; Madara, J.L. Colonic Epithelial HPepT1 Expression Occurs in Inflammatory Bowel Disease: Transport of Bacterial Peptides Influences Expression of MHC Class 1 Molecules. Gastroenterology 2001, 120, 1666–1679. [Google Scholar] [CrossRef]
- Wallace, J.L.; MacNaughton, W.K.; Morris, G.P.; Beck, P.L. Inhibition of Leukotriene Synthesis Markedly Accelerates Healing in a Rat Model of Inflammatory Bowel Disease. Gastroenterology 1989, 96, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Peñate-Medina, T.; Damoah, C.; Benezra, M.; Will, O.; Kairemo, K.; Humbert, J.; Sebens, S.; Peñate-Medina, O. Alpha-MSH Targeted Liposomal Nanoparticle for Imaging in Inflammatory Bowel Disease (IBD). Curr. Pharm. Des. 2020, 26, 3840–3846. [Google Scholar] [CrossRef] [PubMed]
- Panaro, B.L.; Tough, I.R.; Engelstoft, M.S.; Matthews, R.T.; Digby, G.J.; Møller, C.L.; Svendsen, B.; Gribble, F.; Reimann, F.; Holst, J.J.; et al. The Melanocortin-4 Receptor Is Expressed in Enteroendocrine L Cells and Regulates the Release of Peptide YY and Glucagon-like Peptide 1 In Vivo. Cell Metab. 2014, 20, 1018–1029. [Google Scholar] [CrossRef] [Green Version]
- Wei, P.; Yang, Y.; Liu, Z.; Huang, J.; Gong, Y.; Sun, H. Oral Bifidobacterium Longum Expressing Alpha-Melanocyte-Stimulating Hormone to Fight Experimental Colitis. Drug Deliv. 2016, 23, 2058–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bettenworth, D.; Buyse, M.; Böhm, M.; Mennigen, R.; Czorniak, I.; Kannengiesser, K.; Brzoska, T.; Luger, T.A.; Kucharzik, T.; Domschke, W.; et al. The Tripeptide KdPT Protects from Intestinal Inflammation and Maintains Intestinal Barrier Function. Am. J. Pathol. 2011, 179, 1230–1242. [Google Scholar] [CrossRef]
- Böhm, M.; Luger, T. Are Melanocortin Peptides Future Therapeutics for Cutaneous Wound Healing? Exp. Dermatol. 2019, 28, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, S.H.; Lorenzetti, B.B.; Bristow, A.F.; Poole, S. Interleukin-1 Beta as a Potent Hyperalgesic Agent Antagonized by a Tripeptide Analogue. Nature 1988, 334, 698–700. [Google Scholar] [CrossRef]
- Follenfant, R.L.; Nakamura-Craig, M.; Henderson, B.; Higgs, G.A. Inhibition by Neuropeptides of Interleukin-1 Beta-Induced, Prostaglandin-Independent Hyperalgesia. Br. J. Pharmacol. 1989, 98, 41–43. [Google Scholar] [CrossRef]
- Mugridge, K.G.; Perretti, M.; Ghiara, P.; Parente, L. Alpha-Melanocyte-Stimulating Hormone Reduces Interleukin-1 Beta Effects on Rat Stomach Preparations Possibly through Interference with a Type I Receptor. Eur. J. Pharmacol. 1991, 197, 151–155. [Google Scholar] [CrossRef]
- Spencer, J.D.; Schallreuter, K.U. Regulation of Pigmentation in Human Epidermal Melanocytes by Functional High-Affinity Beta-Melanocyte-Stimulating Hormone/Melanocortin-4 Receptor Signaling. Endocrinology 2009, 150, 1250–1258. [Google Scholar] [CrossRef] [Green Version]
- Luger, T.A.; Scholzen, T.; Brzoska, T.; Becher, E.; Slominski, A.; Paus, R. Cutaneous Immunomodulation and Coordination of Skin Stress Responses by Alpha-Melanocyte-Stimulating Hormone. Ann. N. Y. Acad. Sci. 1998, 840, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Loram, L.C.; Culp, M.E.; Connolly-Strong, E.C.; Sturgill-Koszycki, S. Melanocortin Peptides: Potential Targets in Systemic Lupus Erythematosus. Inflammation 2015, 38, 260–271. [Google Scholar] [CrossRef]
- Wan, G.J.; Niewoehner, J.; Hayes, K. Acthar Gel (RCI): A Narrative Literature Review of Clinical and Economic Evidence. Clin. Outcomes Res. 2023, 15, 499–512. [Google Scholar] [CrossRef] [PubMed]
- Chandra, T.; Aggarwal, R. A Narrative Review of Acthar Gel for the Treatment of Myositis. Rheumatol. Ther. 2023, 10, 523–537. [Google Scholar] [CrossRef]
- Aggarwal, R.; Marder, G.; Koontz, D.C.; Nandkumar, P.; Qi, Z.; Oddis, C.V. Efficacy and Safety of Adrenocorticotropic Hormone Gel in Refractory Dermatomyositis and Polymyositis. Ann. Rheum. Dis. 2018, 77, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Montero-Melendez, T. ACTH: The Forgotten Therapy. Semin. Immunol. 2015, 27, 216–226. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Receptor | Ligands | Locations | Main Functions |
|---|---|---|---|
| MC1R | Nle-D-Phe47-α-MSH, α-MSH, β-MSH, γ-MSH, ACTH, agouti protein, PL-8177, PL-8331, ACTH(1-39) | Melanocytes, fibroblasts, immune system cells, glial cells, vascular endothelium, melanoma cells | Melanogenesis, inflammation regulation, vascular endothelial regulation, neuroprotection |
| MC2R | ACTH(1-24), D-Phe47-ACTH(1-24) | Adrenal cortex, adipocytes | Steroidogenesis |
| MC3R | Nle-D-Phe47-α-MSH, α-MSH, β-MSH, γ-MSH, ACTH, AGRP, SHU9119, ACTH(1-39) | CNS, B lymphocytes, macrophages, placenta, heart, intestine, colon | Energy homeostasis, inflammation regulation |
| MC4R | Nle-D-Phe47-α-MSH, α-MSH, β-MSH, γ-MSH, ACTH, agouti protein, AGRP, SHU9119, HS014, HS024, setmelanotide, ACTH(1-39) | CNS, colonic mucosa | Energy homeostasis, food behaviour, neuroprotection |
| MC5R | Nle-D-Phe47-α-MSH, α-MSH, β-MSH, γ-MSH, ACTH, JNJ-10229570, ACTH(1-39) | Ubiquitous | Inflammation regulation, glands secretion |
| First Author, Reference | Year | Main Melanocortin Studied | Inflammatory Bowel Disease Model | Main Results |
|---|---|---|---|---|
| Maaser et al. [64] | 2006 | MC1R mutant mice | DSS-induced colitis in C57BL/6 mice and bone marrow chimaeras, Citrobacter rodentium murine colitis. | Worse colitis parameters (more severe histological changes and worse weight loss) in MC1R mutant mice. Longer duration of Citrobacter rodentium colitis in MC1R mutant mice. |
| Yoon et al. [65] | 2008 | α-MSH-expressing recombinant Lactobacillus casei strains | DSS-induced colitis | Reduced myeloperoxidase activity, improved histological activity and weight loss. |
| Kannengiesser et al. [66] | 2008 | Melanocortin-derived tripeptide α-MSH(11-13) (KPV), MC1R mutant mice | DSS-induced colitis, CD45RBhi transfer colitis | Improvement with KPV treatment of DSS and CD45RBhi colitis regarding weight and histology. Less pronounced improvement in mice with DSS colitis and MC1R-mutated except for lower lethality. |
| Bradamante et al. [67] | 2012 | β-MSH | TNBS-induced colitis | Improved macroscopic picture and colonic necrosis of TNBS-treated mice. |
| Wei et al. [68] | 2016 | α-MSH-expressing recombinant Bifidobacterium longum strains | DSS-induced colitis | Reduced myeloperoxidase activity, TNF, IL-1β, IL-6 downregulation, and IL-10 upregulation. Improved histological activity. |
| Hiramoto et al. [69] | 2016 | MC2R | DSS-induced colitis in combination with ultraviolet A or B irradiation | Better outcomes in DSS colitis combined with ultraviolet A irradiation than B irradiation. Increased MC2R expression in DSS-induced colitis plus ultraviolet B. |
| Xiao et al. [70] | 2017 | Tripeptide KPV in hyaluronic acid-functionalised nanoparticles | DSS-induced murine colitis | Colonic release of KPV and internalisation in colonic cells with TNF downregulation. |
| Spana et al. [71] | 2019 | MC1R agonists (PL-8177, PL-8331) | Human whole blood stimulated by LPS, DNBS colitis Wistar rats | PL-8177 and PL-8331 inhibited TNF levels in whole human blood stimulated by LPS as positive controls (ACTH, α-MSH). PL-8177 improved DNBS colitis as the positive control (sulfasalazine). |
| Gravina et al. [72] | 2022 | MC3R, MC4R | Crohn’s disease and ulcerative colitis human specimen | MC3R and MC4R immunohistochemical expression in IBD colonic mucosa (higher in inflamed tracts versus healthy mucosa). |
| Dodd et al. [73] | 2023 | MC1R agonist PL-8177 | Murine DSS, DNBS colitis, oral PL-8177 pharmacokinetics in rat/dogs, oral PL-8177 in healthy male volunteers (phase 0 study) | DSS and DNBS colitis improvement, colonic bioavailability of PL-8177 in rats/dogs. No systemic circulation of PL-8177 in humans and faecal elimination. No severe adverse events. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gravina, A.G.; Pellegrino, R.; Durante, T.; Palladino, G.; Imperio, G.; D’Amico, G.; Trotta, M.C.; Dallio, M.; Romeo, M.; D’Amico, M.; et al. The Melanocortin System in Inflammatory Bowel Diseases: Insights into Its Mechanisms and Therapeutic Potentials. Cells 2023, 12, 1889. https://doi.org/10.3390/cells12141889
Gravina AG, Pellegrino R, Durante T, Palladino G, Imperio G, D’Amico G, Trotta MC, Dallio M, Romeo M, D’Amico M, et al. The Melanocortin System in Inflammatory Bowel Diseases: Insights into Its Mechanisms and Therapeutic Potentials. Cells. 2023; 12(14):1889. https://doi.org/10.3390/cells12141889
Chicago/Turabian StyleGravina, Antonietta Gerarda, Raffaele Pellegrino, Tommaso Durante, Giovanna Palladino, Giuseppe Imperio, Giovanbattista D’Amico, Maria Consiglia Trotta, Marcello Dallio, Mario Romeo, Michele D’Amico, and et al. 2023. "The Melanocortin System in Inflammatory Bowel Diseases: Insights into Its Mechanisms and Therapeutic Potentials" Cells 12, no. 14: 1889. https://doi.org/10.3390/cells12141889
APA StyleGravina, A. G., Pellegrino, R., Durante, T., Palladino, G., Imperio, G., D’Amico, G., Trotta, M. C., Dallio, M., Romeo, M., D’Amico, M., & Federico, A. (2023). The Melanocortin System in Inflammatory Bowel Diseases: Insights into Its Mechanisms and Therapeutic Potentials. Cells, 12(14), 1889. https://doi.org/10.3390/cells12141889