
Analytical Models for Measuring the Mechanical Properties of Yeast



Abstract
:1. Introduction
2. Mathematical Models for Measuring Mechanical Properties by Micromanipulation
3. Mathematical Models for Measuring Mechanical Properties by the AFM Method
4. Mathematical Models for Measuring Mechanical Properties by the SICM Method
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, Q.; Xia, Y.; Sandig, M.; Yang, J. Characterization of Cell Elasticity Correlated with Cell Morphology by Atomic Force Microscope. J. Biomech. 2012, 45, 304–309. [Google Scholar] [CrossRef]
- Hansma, H.G. Surface Biology of DNA by Atomic Force Microscopy. Annu. Rev. Phys. Chem. 2001, 52, 71–92. [Google Scholar] [CrossRef]
- Mashmoushy, H.; Zhang, Z.; Thomas, C.R. Micromanipulation Measurement of the Mechanical Properties of Baker’s Yeast Cells. Biotechnol. Tech. 1998, 12, 925–929. [Google Scholar] [CrossRef]
- Savin, N.A.; Kolmogorov, V.S.; Timoshenko, R.V.; Klyachko, N.L.; Parkhomenko, Y.N.; Salikhov, S.V.; Majouga, A.G.; Erofeev, A.S.; Gorelkin, P.V. Application of Nanotechnologies in Studying Yeast Structure in Candida. Nanobiotechnol. Rep. 2021, 16, 450–472. [Google Scholar] [CrossRef]
- Gaboriaud, F.; Dufrêne, Y.F. Atomic Force Microscopy of Microbial Cells: Application to Nanomechanical Properties, Surface Forces and Molecular Recognition Forces. Colloids Surf. B Biointerfaces 2007, 54, 10–19. [Google Scholar] [CrossRef]
- Savin, N.; Kolmogorov, V.; Timoshenko, R.; Vaneev, A.; Iakovlev, A.; Suchalko, O.; Grammatikova, N.; Levshin, I.; Klyachko, N.; Parkhomenko, Y.; et al. Application of the Scanning Ion-Conductance Microscopy (SICM) in Study of Voriconazole Impact on Candida Parapsilosis Surface Structure. M&M 2021, 27, 494–495. [Google Scholar] [CrossRef]
- Levshin, I.B.; Simonov, A.Y.; Lavrenov, S.N.; Panov, A.A.; Grammatikova, N.E.; Alexandrov, A.A.; Ghazy, E.S.M.O.; Savin, N.A.; Gorelkin, P.V.; Erofeev, A.S.; et al. Antifungal Thiazolidines: Synthesis and Biological Evaluation of Mycosidine Congeners. Pharmaceuticals 2022, 15, 563. [Google Scholar] [CrossRef]
- Savin, N.; Erofeev, A.; Kolmogorov, V.; Salikhov, S.; Efremov, Y.; Timashev, P.; Grammatikova, N.; Levshin, I.; Edwards, C.; Korchev, Y.; et al. Scanning Ion-Conductance Microscopy Technique for Studying the Topography and Mechanical Properties of Candida Parapsilosis Yeast Microorganisms. Biomater. Sci. 2023, 11, 611–617. [Google Scholar] [CrossRef]
- Savin, N.; Kolmogorov, V.; Iakovlev, A.; Timoshenko, R.; Gorelkin, P.; Erofeev, A.; Korchev, Y.E. Antimicrobial Activity of Antifungal Drugs on Candida Parapsilosis Studied by Scanning Ionconductance Microscopy (SICM). Biophys. J. 2021, 120, 361a. [Google Scholar] [CrossRef]
- Rheinlaender, J.; Geisse, N.A.; Proksch, R.; Schäffer, T.E. Comparison of Scanning Ion Conductance Microscopy with Atomic Force Microscopy for Cell Imaging. Langmuir 2011, 27, 697–704. [Google Scholar] [CrossRef]
- Müller, D.J.; Dumitru, A.C.; Lo Giudice, C.; Gaub, H.E.; Hinterdorfer, P.; Hummer, G.; De Yoreo, J.J.; Dufrêne, Y.F.; Alsteens, D. Atomic Force Microscopy-Based Force Spectroscopy and Multiparametric Imaging of Biomolecular and Cellular Systems. Chem. Rev. 2021, 121, 11701–11725. [Google Scholar] [CrossRef]
- Shi, S.; Wu, T.; Zheng, P. Direct Measurements of the Cobalt-Thiolate Bonds Strength in Rubredoxin by Single-Molecule Force Spectroscopy. ChemBioChem 2022, 23, e202200165. [Google Scholar] [CrossRef]
- Cui, S.; Yu, J.; Kühner, F.; Schulten, K.; Gaub, H.E. Double-Stranded DNA Dissociates into Single Strands When Dragged into a Poor Solvent. J. Am. Chem. Soc. 2007, 129, 14710–14716. [Google Scholar] [CrossRef]
- Pukhova, V.; Banfi, F.; Ferrini, G. Complex Force Dynamics in Atomic Force Microscopy Resolved by Wavelet Transforms. Nanotechnology 2013, 24, 505716. [Google Scholar] [CrossRef]
- Pukhova, V.; Banfi, F.; Ferrini, G. Energy Dissipation in Multifrequency Atomic Force Microscopy. Beilstein J. Nanotechnol. 2014, 5, 494–500. [Google Scholar] [CrossRef]
- Dazzi, A.; Prazeres, R.; Glotin, F.; Ortega, J.M. Analysis of Nano-Chemical Mapping Performed by an AFM-Based (“AFMIR”) Acousto-Optic Technique. Ultramicroscopy 2007, 107, 1194–1200. [Google Scholar] [CrossRef]
- Clède, S.; Lambert, F.; Sandt, C.; Kascakova, S.; Unger, M.; Harté, E.; Plamont, M.-A.; Saint-Fort, R.; Deniset-Besseau, A.; Gueroui, Z.; et al. Detection of an Estrogen Derivative in Two Breast Cancer Cell Lines Using a Single Core Multimodal Probe for Imaging (SCoMPI) Imaged by a Panel of Luminescent and Vibrational Techniques. Analyst 2013, 138, 5627. [Google Scholar] [CrossRef] [Green Version]
- Ruggeri, F.S.; Habchi, J.; Chia, S.; Horne, R.I.; Vendruscolo, M.; Knowles, T.P.J. Infrared Nanospectroscopy Reveals the Molecular Interaction Fingerprint of an Aggregation Inhibitor with Single Aβ42 Oligomers. Nat. Commun. 2021, 12, 688. [Google Scholar] [CrossRef]
- Schulte, A.; Schuhmann, W. Single-Cell Microelectrochemistry. Angew. Chem. Int. Ed. 2007, 46, 8760–8777. [Google Scholar] [CrossRef]
- Fan, Y.; Han, C.; Zhang, B. Recent Advances in the Development and Application of Nanoelectrodes. Analyst 2016, 141, 5474–5487. [Google Scholar] [CrossRef] [Green Version]
- Bonfanti, A.; Fouchard, J.; Khalilgharibi, N.; Charras, G.; Kabla, A. A Unified Rheological Model for Cells and Cellularised Materials. R. Soc. Open Sci. 2020, 7, 190920. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.H.; Maragò, O.M.; Volpe, G. Optical Tweezers; Cambridge University Press: Cambridge, UK, 2015; ISBN 9781107051164. [Google Scholar]
- Ashkin, A. Atomic-Beam Deflection by Resonance-Radiation Pressure. Phys. Rev. Lett. 1970, 25, 1321–1324. [Google Scholar] [CrossRef]
- Maragò, O.M.; Jones, P.H.; Gucciardi, P.G.; Volpe, G.; Ferrari, A.C. Optical Trapping and Manipulation of Nanostructures. Nat. Nanotechnol. 2013, 8, 807–819. [Google Scholar] [CrossRef] [Green Version]
- Ando, J.; Bautista, G.; Smith, N.; Fujita, K.; Daria, V.R. Optical Trapping and Surgery of Living Yeast Cells Using a Single Laser. Rev. Sci. Instrum. 2008, 79, 103705. [Google Scholar] [CrossRef] [Green Version]
- Volpe, G.; Maragò, O.M.; Rubinsztein-Dunlop, H.; Pesce, G.; Stilgoe, A.B.; Volpe, G.; Tkachenko, G.; Truong, V.G.; Chormaic, S.N.; Kalantarifard, F.; et al. Roadmap for Optical Tweezers. J. Phys. Photonics 2023, 5, 022501. [Google Scholar] [CrossRef]
- Ashkin, A.; Dziedzic, J.M.; Yamane, T. Optical Trapping and Manipulation of Single Cells Using Infrared Laser Beams. Nature 1987, 330, 769–771. [Google Scholar] [CrossRef]
- Ashkin, A.; Dziedzic, J.M. Optical Trapping and Manipulation of Viruses and Bacteria. Science 1987, 235, 1517–1520. [Google Scholar] [CrossRef]
- Lee, S.-H. Optimal Integration of Wide Field Illumination and Holographic Optical Tweezers for Multimodal Microscopy with Ultimate Flexibility and Versatility. Opt. Express 2018, 26, 8049. [Google Scholar] [CrossRef]
- Preston, R.D. Polysaccharide Conformation and Cell Wall Function. Annu. Rev. Plant Physiol. 1979, 30, 55–78. [Google Scholar] [CrossRef]
- Stenson, J.D.; Hartley, P.; Wang, C.; Thomas, C.R. Determining the Mechanical Properties of Yeast Cell Walls. Biotechnol. Prog. 2011, 27, 505–512. [Google Scholar] [CrossRef]
- Zhang, Z.; Ferenczi, M.A.; Lush, A.C.; Thomas, C.R. A Novel Micromanipulation Technique for Measuring the Bursting Strength of Single Mammalian Cells. Appl. Microbiol. Biotechnol. 1991, 36, 208–210. [Google Scholar] [CrossRef]
- Peeters, E.A.G.; Bouten, C.V.C.; Oomens, C.W.J.; Baaijens, F.P.T. Monitoring the Biomechanical Response of Individual Cells under Compression: A New Compression Device. Med. Biol. Eng. Comput. 2003, 41, 498–503. [Google Scholar] [CrossRef]
- WANG, C.X. Modelling the Mechanical Properties of Single Suspension-Cultured Tomato Cells. Ann. Bot. 2004, 93, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Stenson, J.D.; Thomas, C.R.; Hartley, P. Modelling the Mechanical Properties of Yeast Cells. Chem. Eng. Sci. 2009, 64, 1892–1903. [Google Scholar] [CrossRef]
- Feng, W.W.; Yang, W.-H. On the Contact Problem of an Inflated Spherical Nonlinear Membrane. J. Appl. Mech. 1973, 40, 209–214. [Google Scholar] [CrossRef]
- Banavar, S.; Gomez, C.; Trogdon, M.; Petzold, L.; Yi, T.; Campas, O. Mechanical Feedback Coordinates Cell Wall Expansion and Assembly in Yeast Mating Morphogenesis. Biophys. J. 2018, 114, 326a. [Google Scholar]
- Mercadé-Prieto, R.; Thomas, C.R.; Zhang, Z. Mechanical Double Layer Model for Saccharomyces Cerevisiae Cell Wall. Eur. Biophys. J. 2013, 42, 613–620. [Google Scholar] [CrossRef]
- Lardner, T.J.; Pujara, P. Compression of Spherical Cells. In Mechanics Today; Elsevier: Amsterdam, The Netherlands, 1980; pp. 161–176. [Google Scholar]
- Cheng, L.Y. Deformation Analyses in Cell and Developmental Biology. Part II—Mechanical Experiments on Cells. J. Biomech. Eng. 1987, 109, 18–24. [Google Scholar] [CrossRef]
- Cheng, L.Y. Deformation Analyses in Cell and Developmental Biology. Part I—Formal Methodology. J. Biomech. Eng. 1987, 109, 10–17. [Google Scholar] [CrossRef]
- Smith, A.E.; Moxham, K.E.; Middelberg, A.P.J. On Uniquely Determining Cell–Wall Material Properties with the Compression Experiment. Chem. Eng. Sci. 1998, 53, 3913–3922. [Google Scholar] [CrossRef]
- Skalak, R.; Tozeren, A.; Zarda, R.P.; Chien, S. Strain Energy Function of Red Blood Cell Membranes. Biophys. J. 1973, 13, 245–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WANG, C.X.; PRITCHARD, J.; THOMAS, C.R. Investigation of the Mechanics of Single Tomato Fruit Cells. J. Texture Stud. 2006, 37, 597–606. [Google Scholar] [CrossRef]
- Malvern, L.E. Introduction to the Mechanics of a Continuous Medium Prentice-Hall; Prentice Hall: Englewood Cliffs, NJ, USA, 1969. [Google Scholar]
- Zhao, L.; Schaefer, D.; Xu, H.; Modi, S.J.; LaCourse, W.R.; Marten, M.R. Elastic Properties of the Cell Wall of Aspergillus Nidulans Studied with Atomic Force Microscopy. Biotechnol. Prog. 2008, 21, 292–299. [Google Scholar] [CrossRef]
- Geitmann, A.; Ortega, J.K.E. Mechanics and Modeling of Plant Cell Growth. Trends Plant Sci. 2009, 14, 467–478. [Google Scholar] [CrossRef]
- Van De Fliert, B.W.; Howell, P.D.; Ockenden, J.R. Pressure-Driven Flow of a Thin Viscous Sheet. J. Fluid. Mech. 1995, 292, 359–376. [Google Scholar] [CrossRef]
- Klis, F.M.; de Koster, C.G.; Brul, S. Cell Wall-Related Bionumbers and Bioestimates of Saccharomyces Cerevisiae and Candida Albicans. Eukaryot. Cell 2014, 13, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Goldenbogen, B.; Giese, W.; Hemmen, M.; Uhlendorf, J.; Herrmann, A.; Klipp, E. Dynamics of Cell Wall Elasticity Pattern Shapes the Cell during Yeast Mating Morphogenesis. Open Biol. 2016, 6, 160136. [Google Scholar] [CrossRef] [Green Version]
- Binnig, G.; Quate, C.F.; Gerber, C. Atomic Force Microscope. Phys. Rev. Lett. 1986, 56, 930–933. [Google Scholar] [CrossRef] [Green Version]
- Hertz, H. Ueber den kontakt elastischer koerper. J. Reine Angew. Math. 1881, 92, 156–171. [Google Scholar]
- Derjaguin, B.V.; Muller, V.M.; Toporov, Y.P. Effect of Contact Deformations on the Adhesion of Particles. J. Colloid. Interface Sci. 1975, 53, 314–326. [Google Scholar] [CrossRef]
- Vella, D.; Ajdari, A.; Vaziri, A.; Boudaoud, A. The Indentation of Pressurized Elastic Shells: From Polymeric Capsules to Yeast Cells. J. R. Soc. Interface 2012, 9, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Emad, A.; Heinz, W.F.; Antonik, M.D.; D’Costa, N.P.; Nageswaran, S.; Schoenenberger, C.-A.; Hoh, J.H. Relative Microelastic Mapping of Living Cells by Atomic Force Microscopy. Biophys. J. 1998, 74, 1564–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timoshenko, S.; Woinowsky-Krieger, S. Theory of Plates and Shells, 2nd ed.; McGraw-Hill: New York, NY, USA, 1970. [Google Scholar]
- Garcia, P.D.; Garcia, R. Determination of the Elastic Moduli of a Single Cell Cultured on a Rigid Support by Force Microscopy. Biophys. J. 2018, 114, 2923–2932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Pinto, R.; Gong, H.; Vahabikashi, A.; Johnson, M. The Effect of the Endothelial Cell Cortex on Atomic Force Microscopy Measurements. Biophys. J. 2013, 105, 300–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico, F.; Roca-Cusachs, P.; Gavara, N.; Farré, R.; Rotger, M.; Navajas, D. Probing Mechanical Properties of Living Cells by Atomic Force Microscopy with Blunted Pyramidal Cantilever Tips. Phys. Rev. E 2005, 72, 021914. [Google Scholar] [CrossRef] [Green Version]
- Briscoe, B.J.; Sebastian, K.S.; Adams, M.J. The Effect of Indenter Geometry on the Elastic Response to Indentation. J. Phys. D Appl. Phys. 1994, 27, 1156–1162. [Google Scholar] [CrossRef]
- Efremov, Y.M.; Wang, W.-H.; Hardy, S.D.; Geahlen, R.L.; Raman, A. Measuring Nanoscale Viscoelastic Parameters of Cells Directly from AFM Force-Displacement Curves. Sci. Rep. 2017, 7, 1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, P.; Takahashi, R.; Kuribayashi-Shigetomi, K.; Subagyo, A.; Sueoka, K.; Maloney, J.M.; Van Vliet, K.J.; Okajima, T. Temporal Variation in Single-Cell Power-Law Rheology Spans the Ensemble Variation of Cell Population. Biophys. J. 2017, 113, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Efremov, Y.M.; Bagrov, D.V.; Kirpichnikov, M.P.; Shaitan, K.V. Application of the Johnson–Kendall–Roberts Model in AFM-Based Mechanical Measurements on Cells and Gel. Colloids Surf. B Biointerfaces 2015, 134, 131–139. [Google Scholar] [CrossRef]
- Vinckier, A.; Semenza, G. Measuring Elasticity of Biological Materials by Atomic Force Microscopy. FEBS Lett. 1998, 430, 12–16. [Google Scholar] [CrossRef]
- Johnson, K.L.; Kendall, K.; Roberts, A.D. Surface Energy and the Contact of Elastic Solids. Proc. R. Soc. Lond. A. Math. Phys. Sci. 1971, 324, 301–313. [Google Scholar] [CrossRef]
- Mahaffy, R.E.; Park, S.; Gerde, E.; Käs, J.; Shih, C.K. Quantitative Analysis of the Viscoelastic Properties of Thin Regions of Fibroblasts Using Atomic Force Microscopy. Biophys. J. 2004, 86, 1777–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, R.A.; Vitorino, M.V.; Godinho, C.P.; Bourbon-Melo, N.; Robalo, T.T.; Fernandes, F.; Rodrigues, M.S.; Sá-Correia, I. Yeast Adaptive Response to Acetic Acid Stress Involves Structural Alterations and Increased Stiffness of the Cell Wall. Sci. Rep. 2021, 11, 12652. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.A.; Godinho, C.P.; Vitorino, M.V.; Robalo, T.T.; Fernandes, F.; Rodrigues, M.S.; Sá-Correia, I. Crosstalk between Yeast Cell Plasma Membrane Ergosterol Content and Cell Wall Stiffness under Acetic Acid Stress Involving Pdr18. J. Fungi 2022, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, T.; Ishii, Y.; Ishikawa, Y.; Matsumoto, T.; Sato, M. Experimental and Numerical Analyses of Local Mechanical Properties Measured by Atomic Force Microscopy for Sheared Endothelial Cells. Biomed. Mater. Eng. 2002, 12, 319–327. [Google Scholar]
- MORLEY, L.S.D. The Thin-Walled Circular Cylinder Subjected to Concentrated Radial Loads. Q. J. Mech. Appl. Math. 1960, 13, 24–37. [Google Scholar] [CrossRef]
- Bui, V.C.; Kim, Y.U.; Choi, S.S. Physical Characteristics of Saccharomyces Cerevisiae. Surf. Interface Anal. 2008, 40, 1323–1327. [Google Scholar] [CrossRef]
- Dague, E.; Bitar, R.; Ranchon, H.; Durand, F.; Yken, H.M.; François, J.M. An Atomic Force Microscopy Analysis of Yeast Mutants Defective in Cell Wall Architecture. Yeast 2010, 27, 673–684. [Google Scholar] [CrossRef]
- Svaldo Lanero, T.; Cavalleri, O.; Krol, S.; Rolandi, R.; Gliozzi, A. Mechanical Properties of Single Living Cells Encapsulated in Polyelectrolyte Matrixes. J. Biotechnol. 2006, 124, 723–731. [Google Scholar] [CrossRef]
- Chaudhari, R.D.; Stenson, J.D.; Overton, T.W.; Thomas, C.R. Effect of Bud Scars on the Mechanical Properties of Saccharomyces Cerevisiae Cell Walls. Chem. Eng. Sci. 2012, 84, 188–196. [Google Scholar] [CrossRef] [Green Version]
- Cao, G.; Chandra, N. Evaluation of Biological Cell Properties Using Dynamic Indentation Measurement. Phys. Rev. E 2010, 81, 021924. [Google Scholar] [CrossRef] [Green Version]
- Sirghi, L. Atomic Force Microscopy Indentation of Living Cells; Formatex Research Center: Badajoz, Spain, 2010. [Google Scholar]
- Arfsten, J.; Leupold, S.; Bradtmöller, C.; Kampen, I.; Kwade, A. Atomic Force Microscopy Studies on the Nanomechanical Properties of Saccharomyces Cerevisiae. Colloids Surf. B Biointerfaces 2010, 79, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Mercadé-Prieto, R.; York, D.; Preece, J.A.; Zhang, Z. Structure and Mechanical Properties of Consumer-Friendly PMMA Microcapsules. Ind. Eng. Chem. Res. 2013, 52, 11253–11265. [Google Scholar] [CrossRef]
- Dimitriadis, E.K.; Horkay, F.; Maresca, J.; Kachar, B.; Chadwick, R.S. Determination of Elastic Moduli of Thin Layers of Soft Material Using the Atomic Force Microscope. Biophys. J. 2002, 82, 2798–2810. [Google Scholar] [CrossRef] [Green Version]
- Sneddon, I.N. The Relation between Load and Penetration in the Axisymmetric Boussinesq Problem for a Punch of Arbitrary Profile. Int. J. Eng. Sci. 1965, 3, 47–57. [Google Scholar] [CrossRef]
- Harding, J.W.; Sneddon, I.N. The Elastic Stresses Produced by the Indentation of the Plane Surface of a Semi-Infinite Elastic Solid by a Rigid Punch. Math. Proc. Camb. Philos. Soc. 1945, 41, 16–26. [Google Scholar] [CrossRef]
- Johnson, K.L. Contact Mechanics; Cambridge University Press: Cambridge, UK, 1985; ISBN 9780521255769. [Google Scholar]
- Pharr, G.M.; Oliver, W.C.; Brotzen, F.R. On the Generality of the Relationship among Contact Stiffness, Contact Area, and Elastic Modulus during Indentation. J. Mater. Res. 1992, 7, 613–617. [Google Scholar] [CrossRef]
- Gavara, N.; Chadwick, R.S. Determination of the Elastic Moduli of Thin Samples and Adherent Cells Using Conical Atomic Force Microscope Tips. Nat. Nanotechnol. 2012, 7, 733–736. [Google Scholar] [CrossRef] [Green Version]
- Costa, K.D.; Yin, F.C.P. Analysis of Indentation: Implications for Measuring Mechanical Properties With Atomic Force Microscopy. J. Biomech. Eng. 1999, 121, 462–471. [Google Scholar] [CrossRef]
- Williams, G.; Watts, D.C. Non-Symmetrical Dielectric Relaxation Behaviour Arising from a Simple Empirical Decay Function. Trans. Faraday Soc. 1970, 66, 80. [Google Scholar] [CrossRef]
- Ossola, D.; Dorwling-Carter, L.; Dermutz, H.; Behr, P.; Vörös, J.; Zambelli, T. Simultaneous Scanning Ion Conductance Microscopy and Atomic Force Microscopy with Microchanneled Cantilevers. Phys. Rev. Lett. 2015, 115, 238103. [Google Scholar] [CrossRef]
- Sánchez, D.; Johnson, N.; Li, C.; Novak, P.; Rheinlaender, J.; Zhang, Y.; Anand, U.; Anand, P.; Gorelik, J.; Frolenkov, G.I.; et al. Noncontact Measurement of the Local Mechanical Properties of Living Cells Using Pressure Applied via a Pipette. Biophys. J. 2008, 95, 3017–3027. [Google Scholar] [CrossRef] [Green Version]
- Rheinlaender, J.; Schäffer, T.E. Mapping the Mechanical Stiffness of Live Cells with the Scanning Ion Conductance Microscope. Soft Matter 2013, 9, 3230. [Google Scholar] [CrossRef]
- Clarke, R.W.; Novak, P.; Zhukov, A.; Tyler, E.J.; Cano-Jaimez, M.; Drews, A.; Richards, O.; Volynski, K.; Bishop, C.; Klenerman, D. Low Stress Ion Conductance Microscopy of Sub-Cellular Stiffness. Soft Matter 2016, 12, 7953–7958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korchev, Y.E.; Milovanovic, M.; Bashford, C.L.; Bennett, D.C.; Sviderskaya, E.V.; Vodyanoy, I.; Lab, M.J. Specialized Scanning Ion-conductance Microscope for Imaging of Living Cells. J. Microsc. 1997, 188, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, M.; Orsini, P.; Pellegrini, M.; Baschieri, P.; Dinelli, F.; Petracchi, D.; Tognoni, E.; Ascoli, C. Weak Hydrostatic Forces in Far-Scanning Ion Conductance Microscopy Used to Guide Neuronal Growth Cones. Neurosci. Res. 2011, 69, 234–240. [Google Scholar] [CrossRef]
- Kolmogorov, V.S.; Erofeev, A.S.; Barykin, E.P.; Timoshenko, R.V.; Lopatukhina, E.V.; Kozin, S.A.; Salikhov, S.V.; Klyachko, N.L.; Mitkevich, V.A.; Edwards, C.R.W.; et al. Scanning Ion-Conductance Microscopy for Studying β-Amyloid Aggregate Formation on Living Cell Surface. bioRxiv 2022. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Model | Function | An Object | Description |
|---|---|---|---|---|
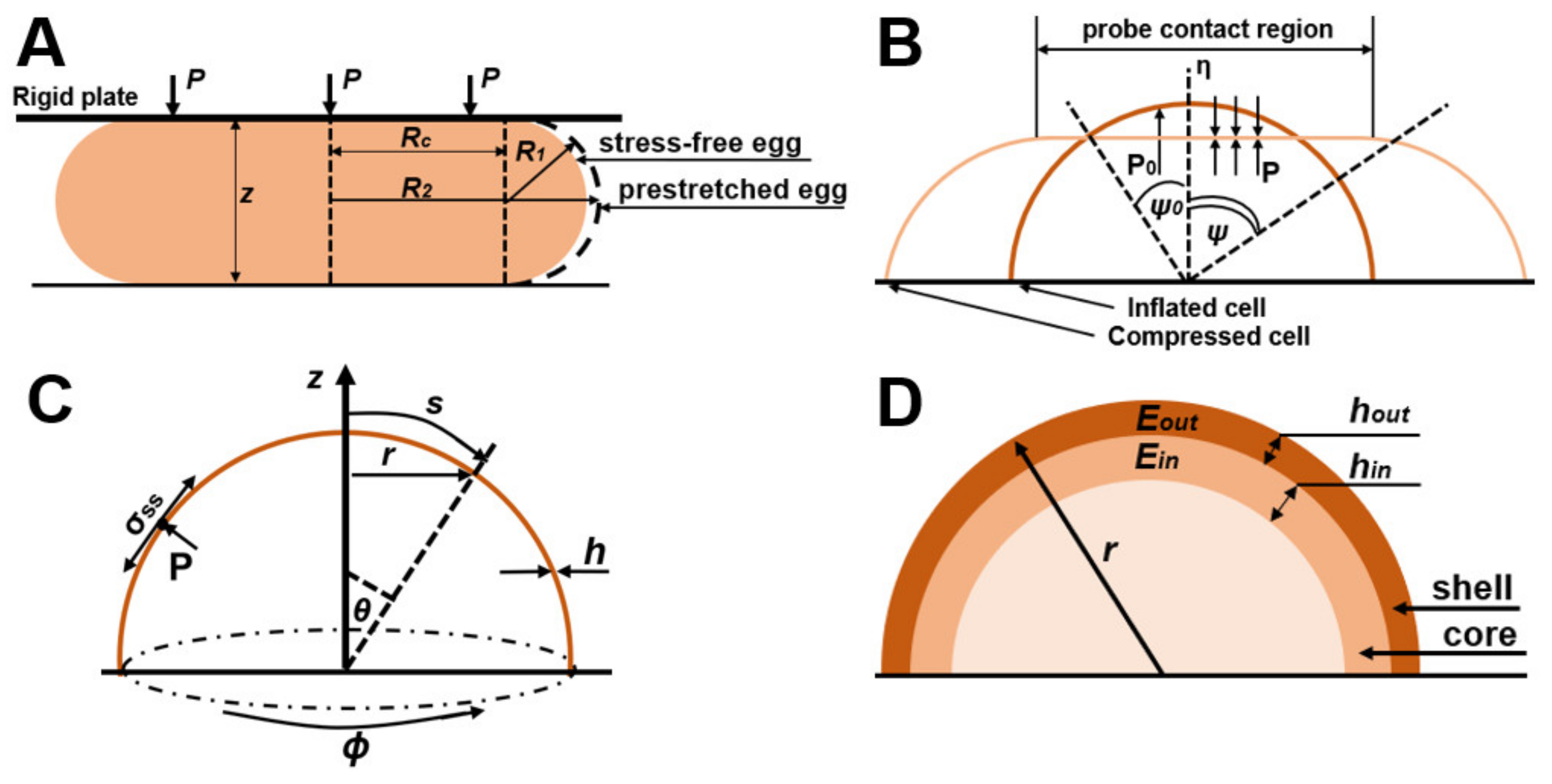
| J.D. Stenson et al. [31,35] | Sea urchin egg model | Infinitely small deformation in Equations (10)–(12), final deformation in Equations (13)–(15), Hankey’s deformation in Equations (16)–(18) | yeast cell wall | In the model, cells are thin-walled, liquid-filled spheres; the desired characteristics depend on the Poisson’s ratio and the thickness of the cell wall. It is possible to neglect cell wall permeability at high strain rates. Fixing the initial stretch factor leads to an inaccurate estimate of the elastic modulus. |
| Feng and Yang [36] | Model of compression of hollow spheres filled with gas | Equations (4)–(7) constitutive equations for contact and non-contact regions | cell wall of tomato cells | The cell wall in this model is divided into areas in contact and areas not in contact with compressive forces. |
| Banavar et al. [37] | Shell theory | Local normal balance of forces of the cell wall in Equation (19) Stresses in the cell wall according to Equations (20) and (21) | growing cell wall dynamics | The growing cell wall behaves like an inhomogeneous viscous liquid with a spatially changing viscosity that increases with distance from the growth apex |
| Mercade’-Prieto et al. [38] | Core-shell model | wall stiffness F/r(Eh)out | cell wall | The model gives an estimate of the overall stiffness of the cell wall (Figure 1D). |
| Source | Model | Function | An Object | Description |
|---|---|---|---|---|
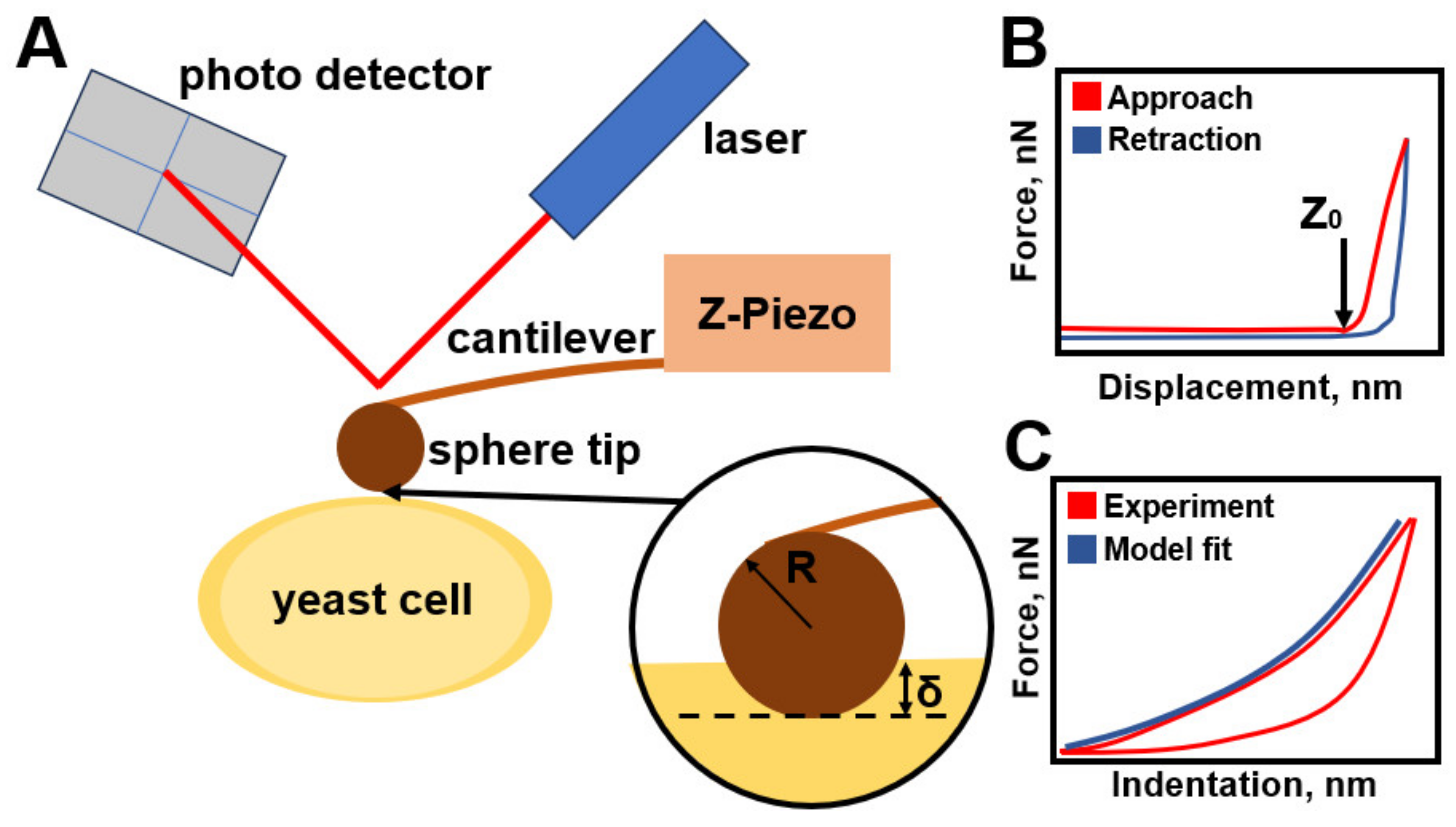
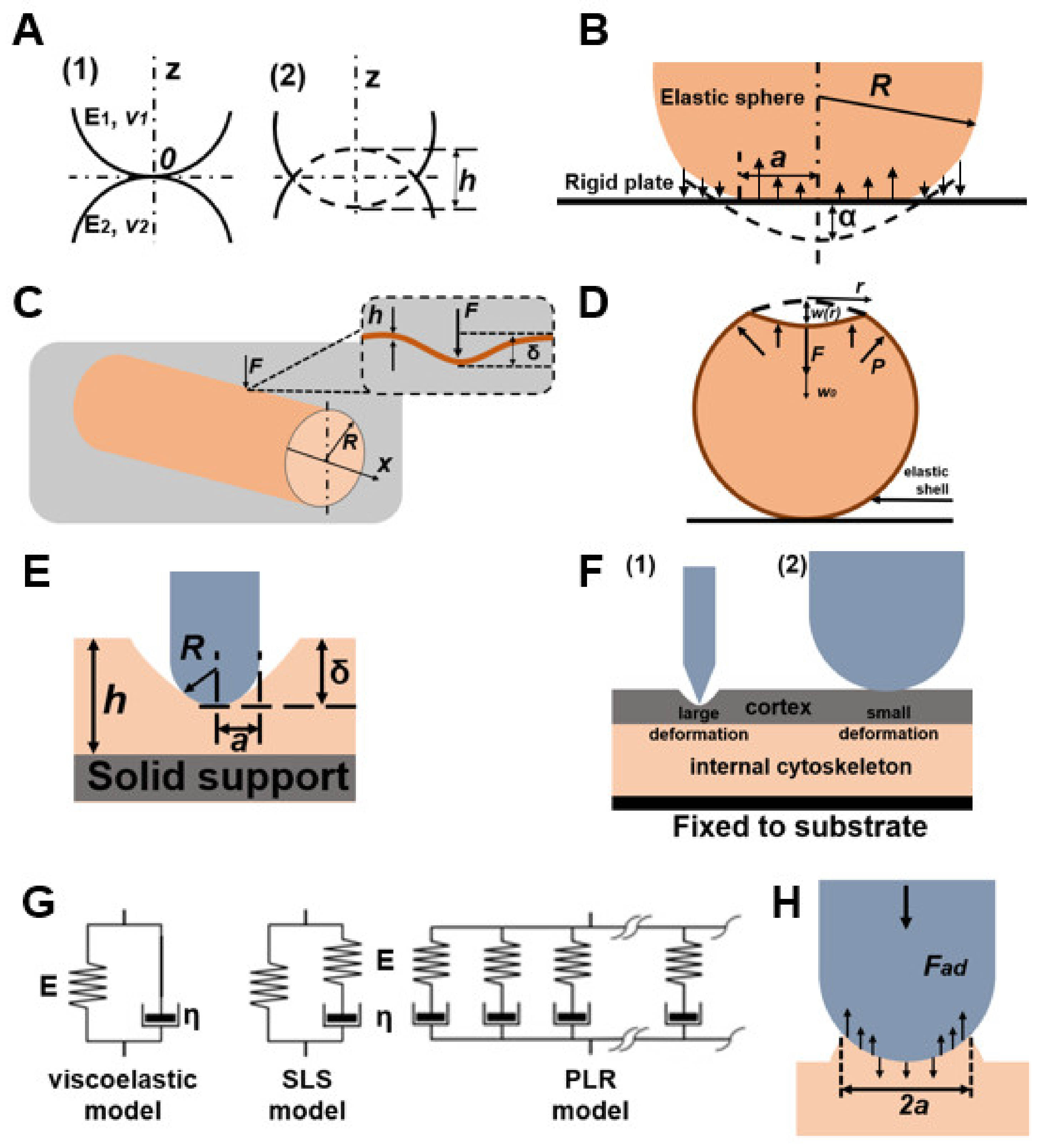
| H. Hertz [52] | Hertz Model | Cantilever Force Equation (22), effective Young’s modulus Equation (23) (When the material of the tip is significantly harder than the material of the sample, Equation (24)) | homogeneous smooth bodies | The model is used under the assumptions that the indenter shape is parabolic, and the sample thickness is much greater than the indentation depth. The model does not allow the probe to stick to the sample. |
| B. Derjaguin [53] | DTM model | cell wall | The model is applicable in the presence of long-range surface forces outside the area of contact between the probe and the sample and is valid in the event of weak adhesion between the nanoindenter and the outer surface of the sample. Its use is a priority for objects with low cohesion and a small radius of curvature. | |
| Zhao et al. [46] | Cylindrical shell model | The modulus of elasticity of the cell wall in Equation (28) | cell wall | In the technique, F and δ are linearly dependent on each other, while the cell wall elasticity constant kw depends on the mechanical properties and dimensions of the cell wall but does not depend on the internal pressure of the cell. |
| Vella et al. [54] | Elastic shell model | internal pressure in yeast cells | Young’s modulus is an order of magnitude higher than the values obtained using the Hertz model. | |
| Mercade’-Prieto et al. [38] | Single layer sphere | The values of F and Eh are calculated from Equations (28) and (29) | cell wall | Corrected values of the Young’s modulus are higher than using Hertz–Sneddon analysis but lower than using micromanipulation compression. |
| Mercade’-Prieto et al. [38] | Double layer model | Force profile at small deformations in Equation (31). | cell wall | The model of a two-layer cell wall suggests the possibility of estimating the elastic modulus by AFM only for the outer layer. |
| E. A-Hassan, S.P. Timoshenko [55,56] | Theory of elastic shells | Young’s modulus is estimated from the ratio between the effective Young’s modulus, shell thickness and bending modulus | cells | Cells in the model are represented as shells filled with liquid. |
| P. Garcia & R. Garcia [57] | Non-Hertz model | In the case of a paraboloid probe, the force is expressed by Equation (33). | mammalian cells attached to a solid support | The cell’s Young’s modulus depends on the solid substrate, and the bottom effect artifact is determined by the ratio between the contact radius and cell thickness. The model is applicable when the indentation is less than or equal to the tip radius. |
| R. Vargas-Pinto et al. [58] | Hertz Model and Contact Model | The force, in the case of a spherical tip, is expressed by Equation (29) In the case of a sharp tip, the model is used Rico et al. [59] and Briscoe et al. [60], where the force is expressed in Equation (34) | mammalian cells with cortex | Combining the models resolved the issue of inaccuracy in determining the rigidity of the cage. Sharp probes examine the cortical layer, and spherical probes record the rigidity of the cortical layer together with the cytoskeleton. In the model presented, the elastic component and the active stress component are combined into an effective elastic response for ease of calculation. |
| Y. Efremov et al. [61] | Elastic-Viscoelastic Compliance | Ting’s solution for indentation of a viscoelastic sample with a rigid spherical tip Equations (35) and (36). | living cells and hydrogels | It reflects the approach-retraction hysteresis well but requires an appropriate choice of the viscoelastic function. |
| Y. Efremov et al. [61] P. Cai et al. [62] | Standard Linear Solid-State Rheology and Power Rheology | Relaxation time Equations (35) and (36), Kohlrausch–Williams–Watts function Equation (37). | living cells | The standard linear rigid body model is a combination of a spring and damper, in which the spring is parallel to the Maxwell element. |
| Y. Efremov et al. [63] | Johnson-Kendall-Roberts model | The indentation depth, contact radius, and maximum adhesive force are presented in Equations (38)–(40), respectively) | living cells and hydrogels | The model fits the retraction part well with force-distance curves. |
| Source | Model | Function | An Object | Description |
|---|---|---|---|---|
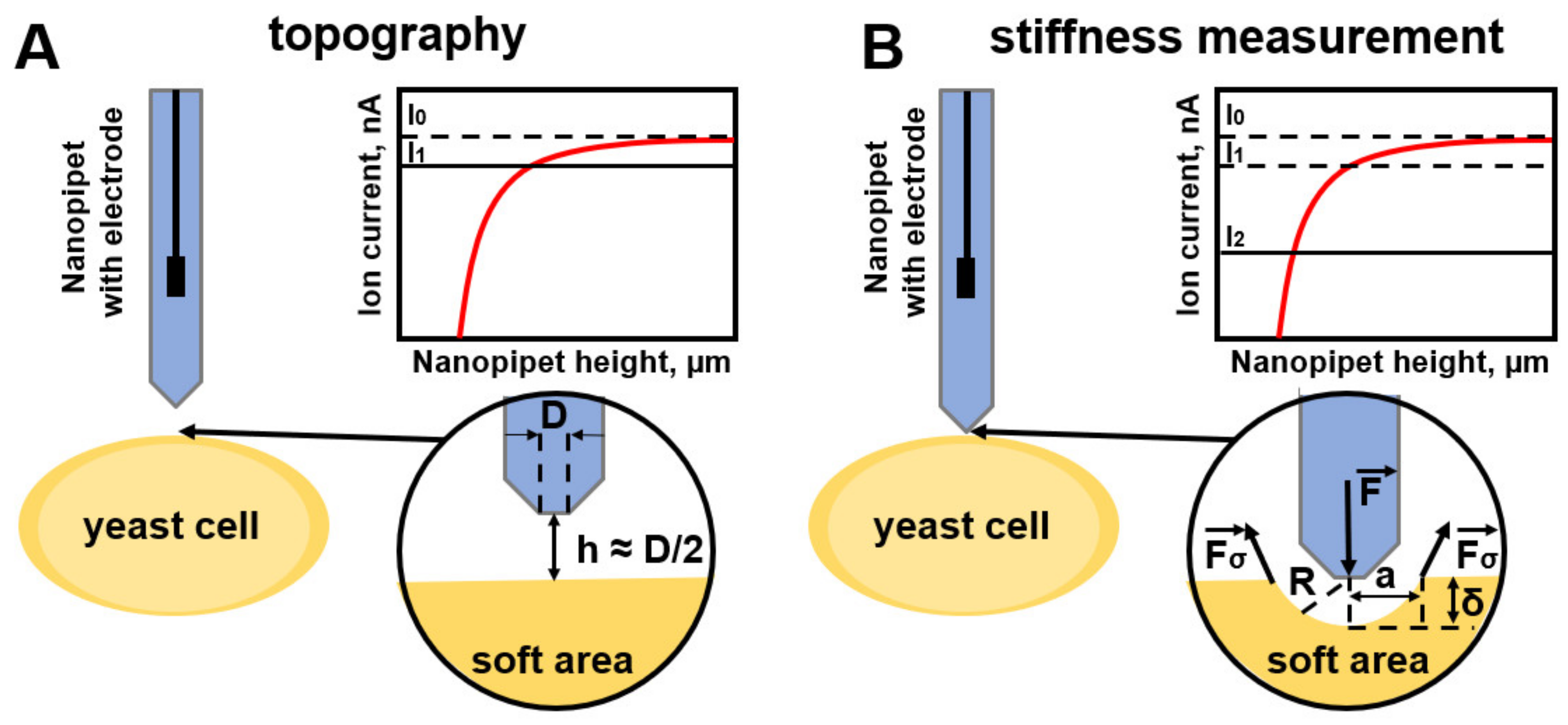
| D. Sanchez et al. [88], Rheinlaender, J., & Schäffer [89] | Hydrodynamic model | The force exerted on a flat surface in Equation (43). Young’s modulus of the sample in Equation (41). | cell membrane | To obtain the mechanical properties of the cell, hydrostatic pressure is applied through a nanopipette, which can lead to a mechanical response of the cell. |
| R. Clarke et al. [90] | Internal colloidal pressure model | The modulus of elasticity of the cell wall in Equation (28) | cells with glycocalyx and cells without glycocalyx | Indentation is performed by means of internal colloidal pressure between the cell surface and the surface of the nanopipette tip, which significantly reduces the invasiveness of the method. |
| Kolmogorov et al. [61], Savin N. et al. [8] | Hertz Model | The internal force is presented in Equation (51). | Mammalian cells [61], yeast cells [8] | The technique is based on the deformation of a double electric layer of decan-saline solution with a nanopipette. The displacement from the tip surface to the cell surface is minimized. However, there is no method for obtaining viscoelastic properties in all presented SICM models. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savin, N.; Erofeev, A.; Gorelkin, P. Analytical Models for Measuring the Mechanical Properties of Yeast. Cells 2023, 12, 1946. https://doi.org/10.3390/cells12151946
Savin N, Erofeev A, Gorelkin P. Analytical Models for Measuring the Mechanical Properties of Yeast. Cells. 2023; 12(15):1946. https://doi.org/10.3390/cells12151946
Chicago/Turabian StyleSavin, Nikita, Alexander Erofeev, and Petr Gorelkin. 2023. "Analytical Models for Measuring the Mechanical Properties of Yeast" Cells 12, no. 15: 1946. https://doi.org/10.3390/cells12151946
APA StyleSavin, N., Erofeev, A., & Gorelkin, P. (2023). Analytical Models for Measuring the Mechanical Properties of Yeast. Cells, 12(15), 1946. https://doi.org/10.3390/cells12151946