
MicroRNAs Regulate Ca2+ Homeostasis in Murine Embryonic Stem Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Maintenance
2.2. Microarray Analysis
2.3. Fura-2 Live Cell Imaging
2.4. Annexin V Staining
2.5. Western Blotting
2.6. RNA Extraction and qPCR
2.7. Plasmids and Transfection
2.8. Statistical Analysis
3. Results
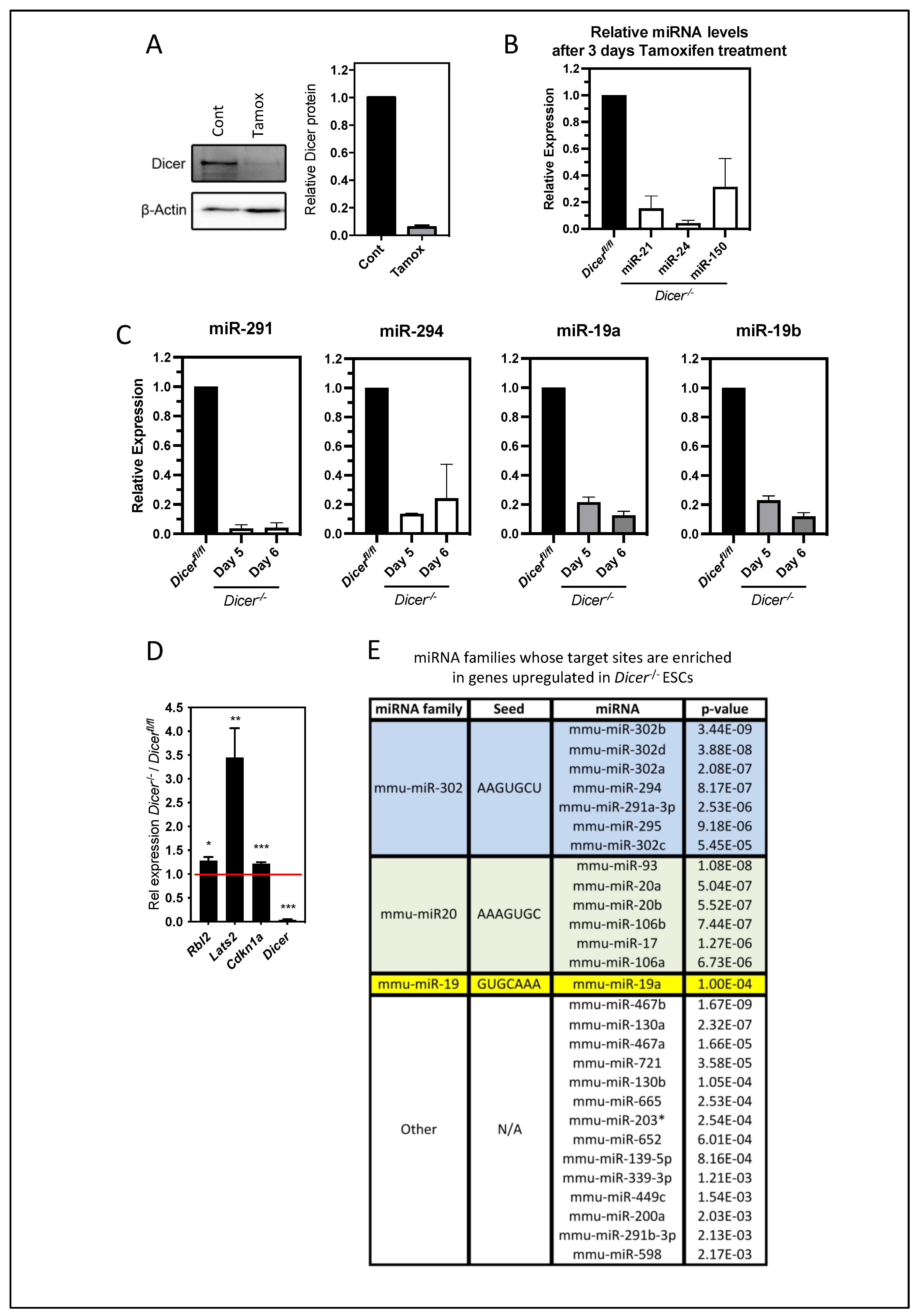
3.1. The Deletion of Dicer in ESCs Results in the Upregulation of Genes Involved in Ca2+ Signaling
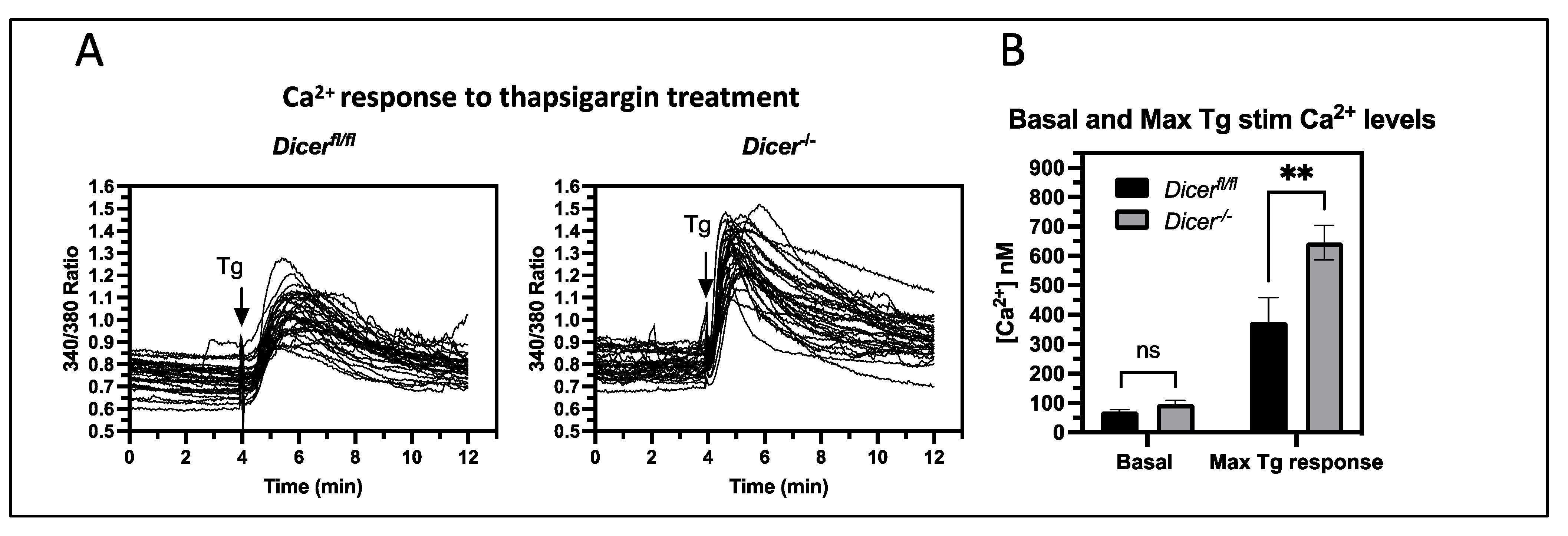
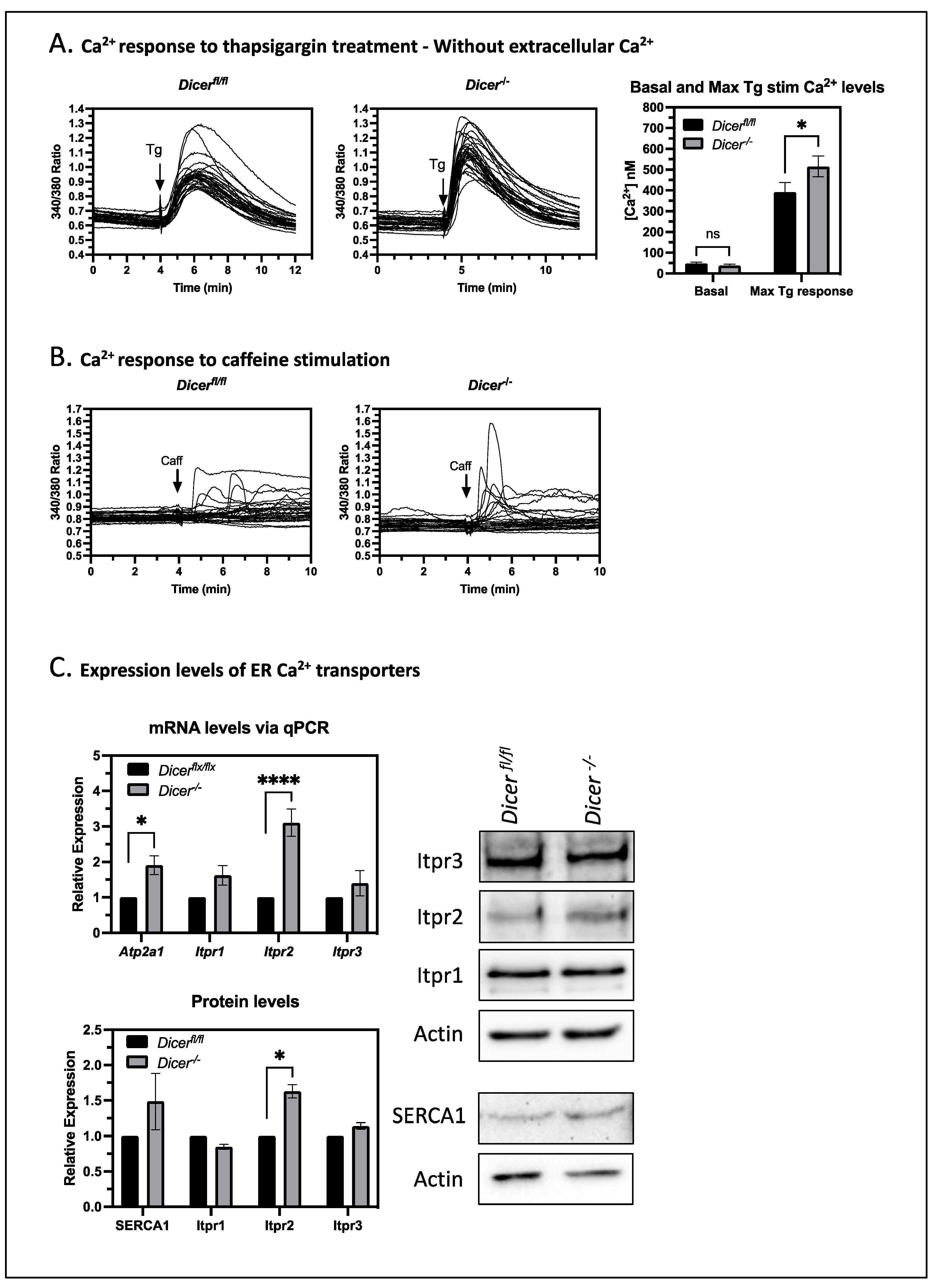
3.2. Dicer−/− ESCs Display Increased Cytoplasmic Ca2+ Levels upon Thapsigargin Treatment
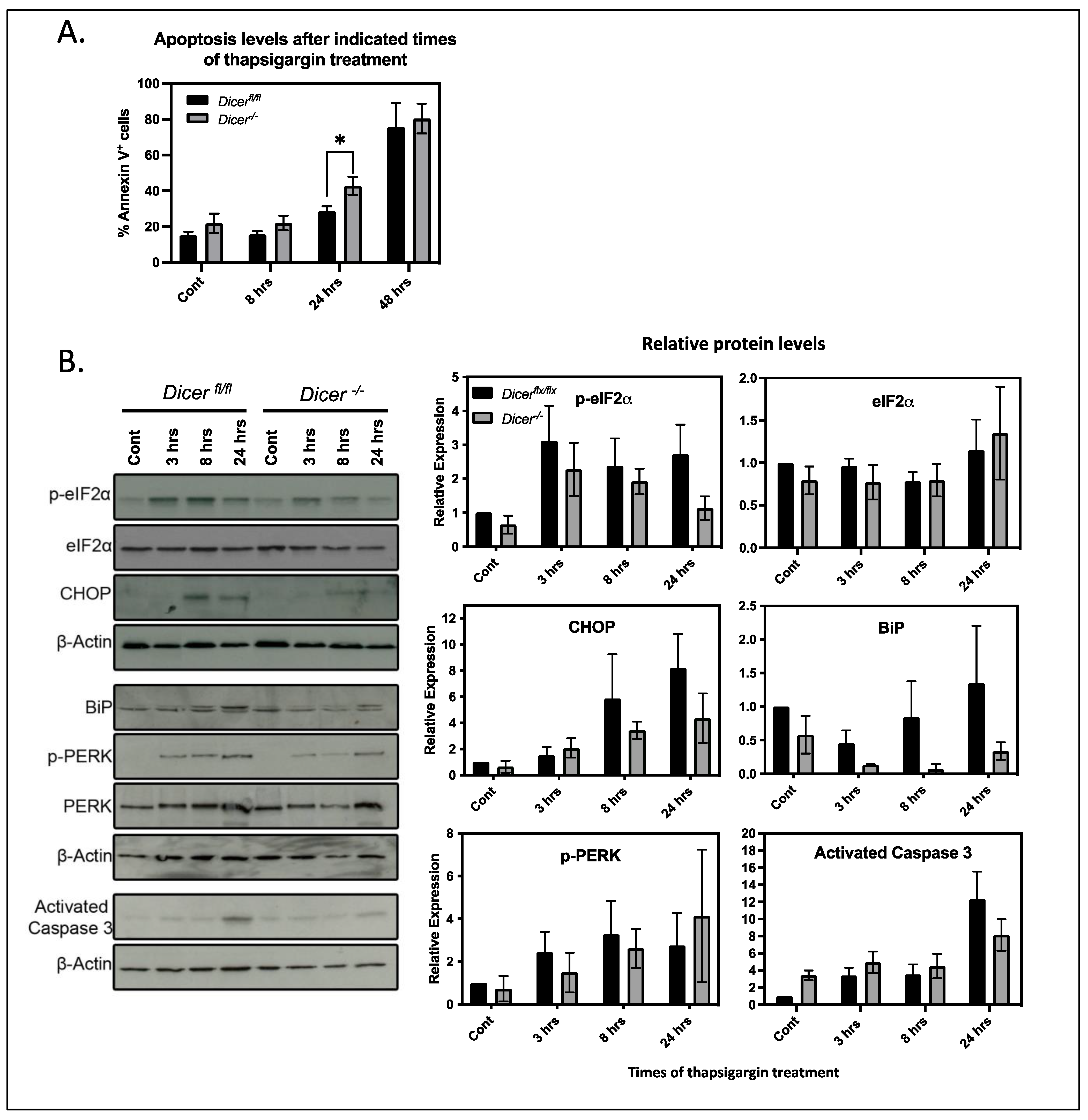
3.3. miRNA Regulation of Ca2+ Homeostasis Plays Virtually No Role in the Stress Response of ESCs
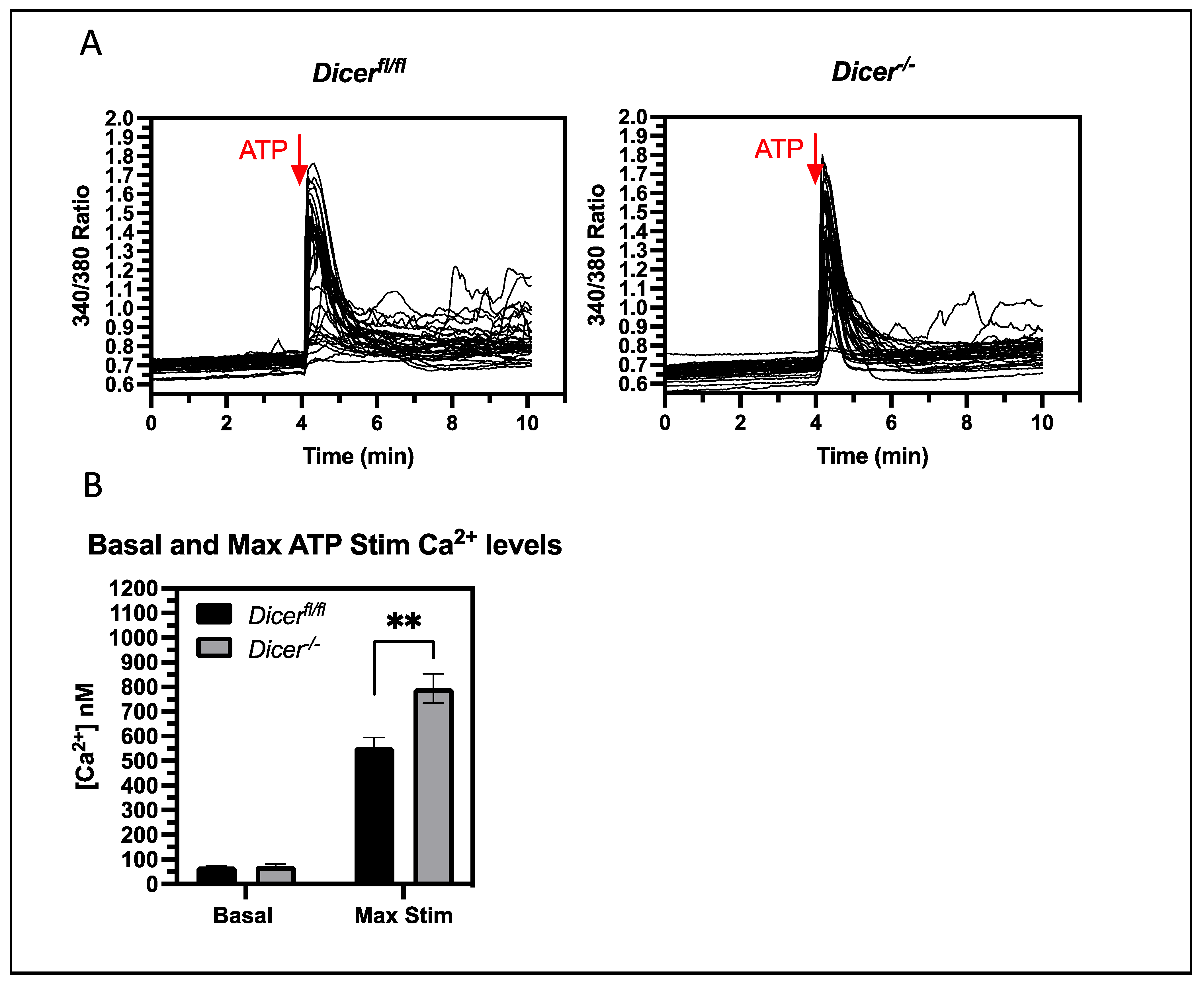
3.4. miRNAs Regulate Cytoplasmic Ca2+ Levels upon the Stimulation of Purinergic Receptors
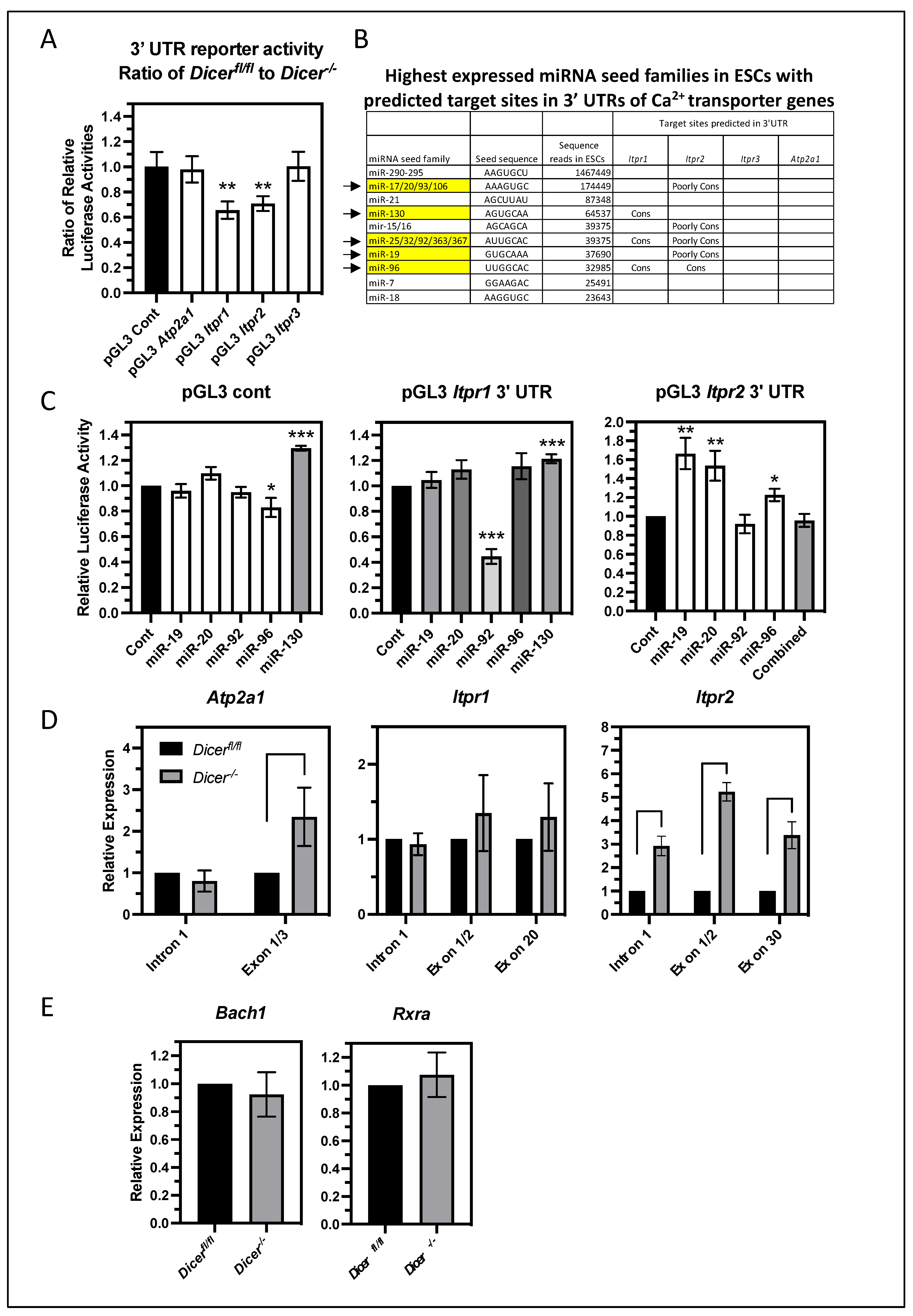
3.5. The miRNA Regulation of Ca2+ Homeostasis Correlates with an Increased Expression of the IP3 Receptor 2 (Itpr2)
3.6. miRNAs Primarily Regulate the Level of Itpr2 Expression Indirectly in ESCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jonas, S.; Izaurralde, E. Towards a Molecular Understanding of MicroRNA-Mediated Gene Silencing. Nat. Rev. Genet. 2015, 16, 421–433. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a Big Role in Gene Regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef]
- Bernstein, E.; Kim, S.Y.; Carmell, M.A.; Murchison, E.P.; Alcorn, H.; Li, M.Z.; Mills, A.A.; Elledge, S.J.; Anderson, K.V.; Hannon, G.J. Dicer Is Essential for Mouse Development. Nat. Genet. 2003, 35, 215–217. [Google Scholar] [CrossRef]
- Spruce, T.; Pernaute, B.; Di-Gregorio, A.; Cobb, B.S.; Merkenschlager, M.; Manzanares, M.; Rodriguez, T.A. An Early Developmental Role for MiRNAs in the Maintenance of Extraembryonic Stem Cells in the Mouse Embryo. Dev. Cell 2010, 19, 207–219. [Google Scholar] [CrossRef] [Green Version]
- Murchison, E.P.; Partridge, J.F.; Tam, O.H.; Cheloufi, S.; Hannon, G.J. Characterization of Dicer-Deficient Murine Embryonic Stem Cells. Proc. Natl. Acad. Sci. USA 2005, 102, 12135–12140. [Google Scholar] [CrossRef]
- Kanellopoulou, C.; Muljo, S.A.; Kung, A.L.; Ganesan, S.; Drapkin, R.; Jenuwein, T.; Livingston, D.M.; Rajewsky, K. Dicer-Deficient Mouse Embryonic Stem Cells Are Defective in Differentiation and Centromeric Silencing. Genes Dev. 2005, 19, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Benetti, R.; Gonzalo, S.; Jaco, I.; Muñoz, P.; Gonzalez, S.; Schoeftner, S.; Murchison, E.; Andl, T.; Chen, T.; Klatt, P.; et al. A Mammalian MicroRNA Cluster Controls DNA Methylation and Telomere Recombination via Rbl2-Dependent Regulation of DNA Methyltransferases. Nat. Struct. Mol. Biol. 2008, 15, 268–279. [Google Scholar] [CrossRef]
- Sinkkonen, L.; Hugenschmidt, T.; Berninger, P.; Gaidatzis, D.; Mohn, F.; Artus-Revel, C.G.; Zavolan, M.; Svoboda, P.; Filipowicz, W. MicroRNAs Control de Novo DNA Methylation through Regulation of Transcriptional Repressors in Mouse Embryonic Stem Cells. Nat. Struct. Mol. Biol. 2008, 15, 259–267. [Google Scholar] [CrossRef]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 Coding Regions Modulate Embryonic Stem Cell Differentiation. Nature 2008, 455, 1124–1128. [Google Scholar] [CrossRef]
- Melton, C.; Judson, R.L.; Blelloch, R. Opposing MicroRNA Families Regulate Self-Renewal in Mouse Embryonic Stem Cells. Nature 2010, 463, 621–626. [Google Scholar] [CrossRef]
- Ma, Y.; Yao, N.; Liu, G.; Dong, L.; Liu, Y.; Zhang, M.; Wang, F.; Wang, B.; Wei, X.; Dong, H.; et al. Functional Screen Reveals Essential Roles of MiR-27a/24 in Differentiation of Embryonic Stem Cells. EMBO J. 2015, 34, 361–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Baskerville, S.; Shenoy, A.; Babiarz, J.E.; Baehner, L.; Blelloch, R. Embryonic Stem Cell–Specific MicroRNAs Regulate the G1-S Transition and Promote Rapid Proliferation. Nat. Genet. 2008, 40, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Guo, W.-T.; Tian, S.; He, X.; Wang, X.-W.; Liu, X.; Gu, K.-L.; Ma, X.; Huang, D.; Hu, L.; et al. MiR-290/371-Mbd2-Myc Circuit Regulates Glycolytic Metabolism to Promote Pluripotency. EMBO J. 2015, 34, 609–623. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.X.Y.; Ravi, A.; Calabrese, J.M.; Medeiros, L.A.; Kirak, O.; Dennis, L.M.; Jaenisch, R.; Burge, C.B.; Sharp, P.A. A Latent Pro-Survival Function for the Mir-290-295 Cluster in Mouse Embryonic Stem Cells. PLoS Genet. 2011, 7, e1002054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, B.; Marcais, A.; Dharmalingam, G.; Carroll, T.; Kanellopoulou, C.; Graumann, J.; Nesterova, T.B.; Bermange, A.; Brazauskas, P.; Xella, B.; et al. MicroRNAs of the MiR-290-295 Family Maintain Bivalency in Mouse Embryonic Stem Cells. Stem Cell Rep. 2016, 6, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Hao, B.; Webb, S.E.; Miller, A.L.; Yue, J. The Role of Ca2+ Signaling on the Self-Renewal and Neural Differentiation of Embryonic Stem Cells (ESCs). Cell Calcium 2016, 59, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhu, L.; Yang, A.; Lin, J.; Tang, F.; Jin, S.; Wei, Z.; Li, J.; Jin, Y. Calcineurin-NFAT Signaling Critically Regulates Early Lineage Specification in Mouse Embryonic Stem Cells and Embryos. Cell Stem Cell 2011, 8, 46–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, S.E.; Miller, A.L. Calcium Signalling during Embryonic Development. Nat. Rev. Mol. Cell Biol. 2003, 4, 539–551. [Google Scholar] [CrossRef]
- Finger, F.; Hoppe, T. MicroRNAs Meet Calcium: Joint Venture in ER Proteostasis. Sci. Signal. 2014, 7, re11. [Google Scholar] [CrossRef] [Green Version]
- Nesterova, T.B.; Popova, B.C.; Cobb, B.S.; Norton, S.; Senner, C.E.; Tang, Y.A.; Spruce, T.; Rodriguez, T.A.; Sado, T.; Merkenschlager, M.; et al. Dicer Regulates Xist Promoter Methylation in ES Cells Indirectly through Transcriptional Control of Dnmt3a. Epigenetics Chromatin 2008, 1, 2–21. [Google Scholar] [CrossRef] [Green Version]
- Medina, I.; Carbonell, J.; Pulido, L.; Madeira, S.C.; Goetz, S.; Conesa, A.; Tárraga, J.; Pascual-Montano, A.; Nogales-Cadenas, R.; Santoyo, J.; et al. Babelomics: An Integrative Platform for the Analysis of Transcriptomics, Proteomics and Genomic Data with Advanced Functional Profiling. Nucleic Acids Res. 2010, 38, W210–W213. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Malgaroli, A.; Milani, D.; Meldolesi, J.; Pozzan, T. Fura-2 Measurement of Cytosolic Free Ca2+ in Monolayers and Suspensions of Various Types of Animal Cells. J. Cell Biol. 1987, 105, 2145–2155. [Google Scholar] [CrossRef]
- Pernaute, B.; Spruce, T.; Smith, K.M.; Sánchez-Nieto, J.M.; Manzanares, M.; Cobb, B.; Rodriguez, T.A. MicroRNAs Control the Apoptotic Threshold in Primed Pluripotent Stem Cells through Regulation of BIM. Genes Dev. 2014, 28, 1873–1878. [Google Scholar] [CrossRef] [Green Version]
- Babiarz, J.E.; Ruby, J.G.; Wang, Y.; Bartel, D.P.; Blelloch, R. Mouse ES Cells Express Endogenous ShRNAs, SiRNAs, and Other Microprocessor-Independent, Dicer-Dependent Small RNAs. Genes Dev. 2008, 22, 2773–2785. [Google Scholar] [CrossRef] [Green Version]
- Yanagida, E.; Shoji, S.; Hirayama, Y.; Yoshikawa, F.; Otsu, K.; Uematsu, H.; Hiraoka, M.; Furuichi, T.; Kawano, S. Functional Expression of Ca2+ Signaling Pathways in Mouse Embryonic Stem Cells. Cell Calcium 2004, 36, 135–146. [Google Scholar] [CrossRef]
- Pinton, P.; Giorgi, C.; Siviero, R.; Zecchini, E.; Rizzuto, R. Calcium and Apoptosis: ER-Mitochondria Ca2+ Transfer in the Control of Apoptosis. Oncogene 2008, 27, 6407–6418. [Google Scholar] [CrossRef] [Green Version]
- Anderson, P.; Kedersha, N. Stressful Initiations. J. Cell Sci. 2002, 115, 3227–3234. [Google Scholar] [CrossRef]
- Holcik, M.; Sonenberg, N. Translational Control in Stress and Apoptosis. Nat. Rev. Mol. Cell Biol. 2005, 6, 318–327. [Google Scholar] [CrossRef]
- Emde, A.; Hornstein, E. MiRNAs at the Interface of Cellular Stress and Disease. EMBO J. 2014, 33, 1428–1437. [Google Scholar] [CrossRef] [Green Version]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates That Thousands of Human Genes Are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Warnatz, H.-J.; Schmidt, D.; Manke, T.; Piccini, I.; Sultan, M.; Borodina, T.; Balzereit, D.; Wruck, W.; Soldatov, A.; Vingron, M.; et al. The BTB and CNC Homology 1 (BACH1) Target Genes Are Involved in the Oxidative Stress Response and in Control of the Cell Cycle*. J. Biol. Chem. 2011, 286, 23521–23532. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Warnier, M.; Raynard, C.; Ferrand, M.; Kirsh, O.; Defossez, P.; Martin, N.; Bernard, D. The Nuclear Receptor RXRA Controls Cellular Senescence by Regulating Calcium Signaling. Aging Cell 2018, 17, e12831. [Google Scholar] [CrossRef]
- Drawnel, F.M.; Wachten, D.; Molkentin, J.D.; Maillet, M.; Aronsen, J.M.; Swift, F.; Sjaastad, I.; Liu, N.; Catalucci, D.; Mikoshiba, K.; et al. Mutual Antagonism between IP3RII and MiRNA-133a Regulates Calcium Signals and Cardiac Hypertrophy. J. Cell Biol. 2012, 199, 783–798. [Google Scholar] [CrossRef]
- Diener, C.; Hart, M.; Alansary, D.; Poth, V.; Walch-Rückheim, B.; Menegatti, J.; Grässer, F.; Fehlmann, T.; Rheinheimer, S.; Niemeyer, B.A.; et al. Modulation of Intracellular Calcium Signaling by MicroRNA-34a-5p. Cell Death Dis. 2018, 9, 1008. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium Signalling: Dynamics, Homeostasis and Remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [Green Version]
- Leclerc, C.; Néant, I.; Moreau, M. The Calcium: An Early Signal That Initiates the Formation of the Nervous System during Embryogenesis. Front. Mol. Neurosci. 2012, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Papanayotou, C.; Almeida, I.D.; Liao, P.; Oliveira, N.M.M.; Lu, S.-Q.; Kougioumtzidou, E.; Zhu, L.; Shaw, A.; Sheng, G.; Streit, A.; et al. Calfacilitin Is a Calcium Channel Modulator Essential for Initiation of Neural Plate Development. Nat. Commun. 2013, 4, 1837. [Google Scholar] [CrossRef]
- Tyser, R.; Miranda, A.; Chen, C.; Davidson, S.M. Calcium Handling Precedes Cardiac Differentiation to Initiate the First Heartbeat. eLife 2016, 5, e17113. [Google Scholar] [CrossRef] [PubMed]
- Song, M.-S.; Rossi, J.J. Molecular Mechanisms of Dicer: Endonuclease and Enzymatic Activity. Biochem. J. 2017, 474, 1603–1618. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reid, K.M.; Sanchez-Nieto, J.M.; Terrasse, S.; Faccenda, D.; Pernaute, B.; Campanella, M.; Rodriguez, T.A.; Cobb, B.S. MicroRNAs Regulate Ca2+ Homeostasis in Murine Embryonic Stem Cells. Cells 2023, 12, 1957. https://doi.org/10.3390/cells12151957
Reid KM, Sanchez-Nieto JM, Terrasse S, Faccenda D, Pernaute B, Campanella M, Rodriguez TA, Cobb BS. MicroRNAs Regulate Ca2+ Homeostasis in Murine Embryonic Stem Cells. Cells. 2023; 12(15):1957. https://doi.org/10.3390/cells12151957
Chicago/Turabian StyleReid, Kimberley M., Juan Miguel Sanchez-Nieto, Sandra Terrasse, Danilo Faccenda, Barbara Pernaute, Michelangelo Campanella, Tristan A. Rodriguez, and Bradley S. Cobb. 2023. "MicroRNAs Regulate Ca2+ Homeostasis in Murine Embryonic Stem Cells" Cells 12, no. 15: 1957. https://doi.org/10.3390/cells12151957
APA StyleReid, K. M., Sanchez-Nieto, J. M., Terrasse, S., Faccenda, D., Pernaute, B., Campanella, M., Rodriguez, T. A., & Cobb, B. S. (2023). MicroRNAs Regulate Ca2+ Homeostasis in Murine Embryonic Stem Cells. Cells, 12(15), 1957. https://doi.org/10.3390/cells12151957