

Utility of the Zebrafish Model for Studying Neuronal and Behavioral Disturbances Induced by Embryonic Exposure to Alcohol, Nicotine, and Cannabis


Abstract
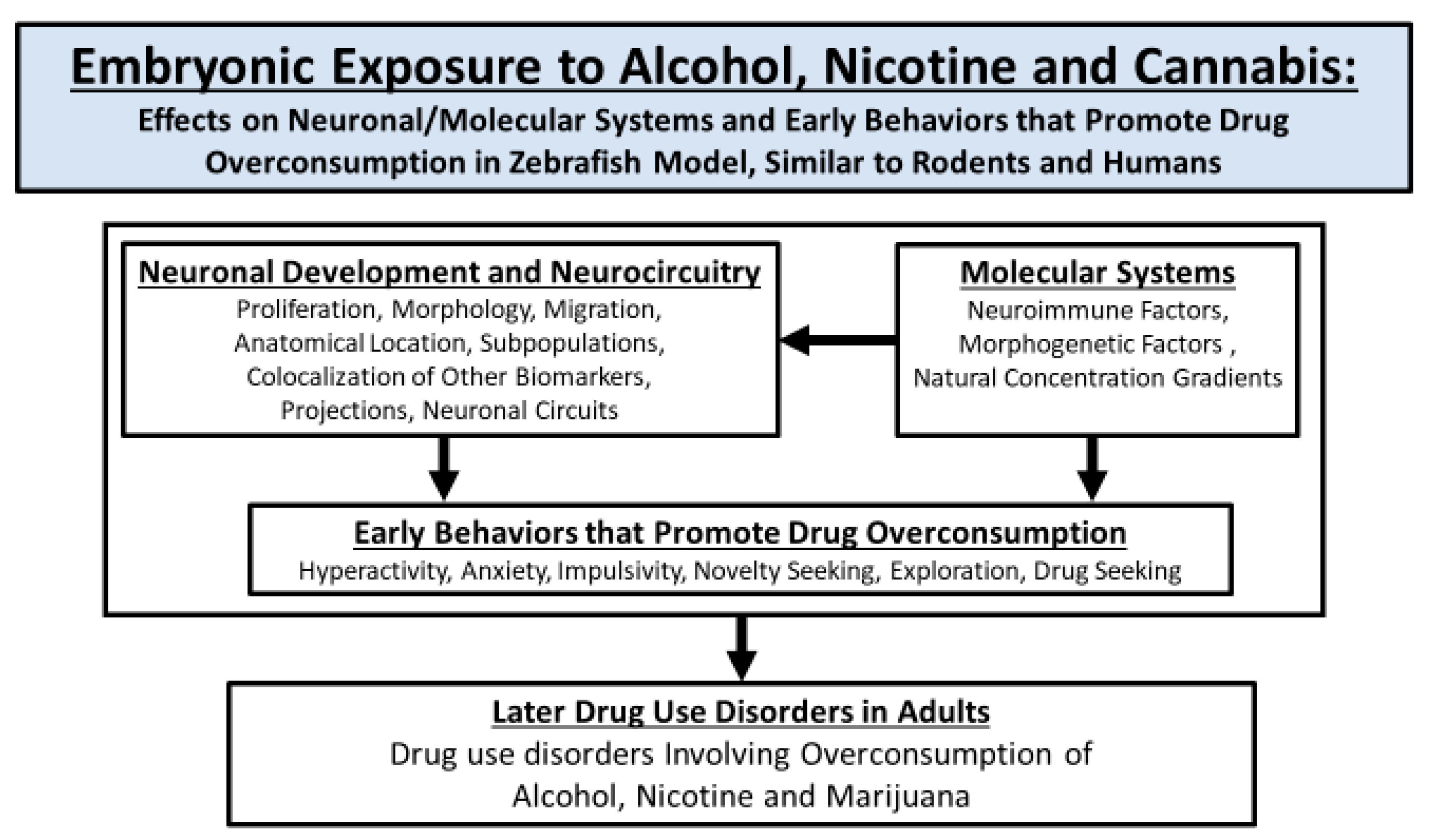
:1. Introduction
2. Human Studies of Prenatal Exposure to Drugs of Abuse
2.1. Advantages and Limitations of Human Studies
2.2. Behavioral Disturbances Observed in Human Studies
2.3. Neuronal Disturbances Observed in Human Studies
3. Rodent Studies of Prenatal Exposure to Drugs of Abuse
3.1. Advantages and Limitations of Rodent Studies
3.2. Behavioral Disturbances Observed in Rodent Studies
3.3. Neuronal Disturbances Observed in Rodent Studies
4. Zebrafish Studies of Embryonic Exposure to Drugs of Abuse
4.1. Advantages and Limitations of Zebrafish Studies
4.2. Behavioral Disturbances Observed in Zebrafish Studies
4.3. Neuronal Disturbances Observed in Zebrafish Studies
4.3.1. Neuropeptide Neuronal Disturbances Observed in Zebrafish
4.3.2. Neuronal Morphology and Circuitry Disturbances Observed in Zebrafish
4.4. Mechanisms Underlying Neuronal and Behavioral Disturbances in Zebrafish
4.4.1. Brain Mechanisms Underlying Behavioral Disturbances in Zebrafish
4.4.2. Brain Mechanisms Underlying Neuronal Disturbances in Zebrafish
4.5. Direct Relation of Disturbances in Neurons to Changes in Zebrafish Behavior
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Wendell, A.D. Overview and epidemiology of substance abuse in pregnancy. Clin. Obs. Gynecol. 2013, 56, 91–96. [Google Scholar] [CrossRef]
- Forray, A. Substance use during pregnancy. F1000Res 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Collier, A.D.; Halkina, V.; Min, S.S.; Roberts, M.Y.; Campbell, S.D.; Camidge, K.; Leibowitz, S.F. Embryonic Ethanol Exposure Affects the Early Development, Migration, and Location of Hypocretin/Orexin Neurons in Zebrafish. Alcohol. Clin. Exp. Res. 2019, 43, 1702–1713. [Google Scholar] [CrossRef]
- Collier, A.D.; Yasmin, N.; Karatayev, O.; Abdulai, A.R.; Yu, B.; Fam, M.; Campbell, S.; Leibowitz, S.F. Embryonic ethanol exposure and optogenetic activation of hypocretin neurons stimulate similar behaviors early in life associated with later alcohol consumption. Sci. Rep. 2023, in press. [Google Scholar]
- Collier, A.D.; Yasmin, N.; Chang, G.Q.; Karatayev, O.; Khalizova, N.; Fam, M.; Abdulai, A.R.; Yu, B.; Leibowitz, S.F. Embryonic ethanol exposure induces ectopic Hcrt and MCH neurons outside hypothalamus in rats and zebrafish: Role in ethanol-induced behavioural disturbances. Addict. Biol. 2022, 27, e13238. [Google Scholar] [CrossRef] [PubMed]
- Collier, A.D.; Yasmin, N.; Karatayev, O.; Abdulai, A.R.; Yu, B.; Khalizova, N.; Fam, M.; Leibowitz, S.F. Neuronal chemokine concentration gradients mediate effects of embryonic ethanol exposure on ectopic hypocretin/orexin neurons and behavior in zebrafish. Sci. Rep. 2023, 13, 1447. [Google Scholar] [CrossRef]
- Yasmin, N.; Collier, A.D.; Karatayev, O.; Abdulai, A.R.; Yu, B.; Fam, M.; Khalizova, N.; Leibowitz, S.F. Subpopulations of hypocretin/orexin neurons differ in measures of their cell proliferation, dynorphin co-expression, projections, and response to embryonic ethanol exposure. Sci. Rep. 2023, 13, 8448. [Google Scholar] [CrossRef]
- Yao, H.; Wu, W.; Cerf, I.; Zhao, H.W.; Wang, J.; Negraes, P.D.; Muotri, A.R.; Haddad, G.G. Methadone interrupts neural growth and function in human cortical organoids. Stem Cell Res. 2020, 49, 102065. [Google Scholar] [CrossRef]
- Etemadi-Aleagha, A.; Akhgari, M. Psychotropic drug abuse in pregnancy and its impact on child neurodevelopment: A review. World J. Clin. Pediatr. 2022, 11, 1–13. [Google Scholar] [CrossRef]
- Duko, B.; Ayano, G.; Pereira, G.; Betts, K.; Alati, R. Prenatal tobacco use and the risk of mood disorders in offspring: A systematic review and meta-analysis. Soc. Psychiatry Psychiatr. Epidemiol. 2020, 55, 1549–1562. [Google Scholar] [CrossRef]
- Lees, B.; Mewton, L.; Jacobus, J.; Valadez, E.A.; Stapinski, L.A.; Teesson, M.; Tapert, S.F.; Squeglia, L.M. Association of Prenatal Alcohol Exposure With Psychological, Behavioral, and Neurodevelopmental Outcomes in Children From the Adolescent Brain Cognitive Development Study. Am. J. Psychiatry 2020, 177, 1060–1072. [Google Scholar] [CrossRef]
- Ross, E.J.; Graham, D.L.; Money, K.M.; Stanwood, G.D. Developmental consequences of fetal exposure to drugs: What we know and what we still must learn. Neuropsychopharmacology 2015, 40, 61–87. [Google Scholar] [CrossRef]
- Maggs, J.L.; Schulenberg, J.E. Initiation and course of alcohol consumption among adolescents and young adults. Recent. Dev. Alcohol. 2005, 17, 29–47. [Google Scholar] [CrossRef]
- Groenman, A.P.; Janssen, T.W.P.; Oosterlaan, J. Childhood Psychiatric Disorders as Risk Factor for Subsequent Substance Abuse: A Meta-Analysis. J. Am. Acad. Child. Adolesc. Psychiatry 2017, 56, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.R.; Felton, J.W.; Risco, C.M.; Lejuez, C.W.; MacPherson, L. Brief report: The interaction of impulsivity with risk-taking is associated with early alcohol use initiation. J. Adolesc. 2014, 37, 1253–1256. [Google Scholar] [CrossRef]
- Watts, A.L.; Wood, P.K.; Jackson, K.M.; Lisdahl, K.M.; Heitzeg, M.M.; Gonzalez, R.; Tapert, S.F.; Barch, D.M.; Sher, K.J. Incipient alcohol use in childhood: Early alcohol sipping and its relations with psychopathology and personality. Dev. Psychopathol. 2021, 33, 1338–1350. [Google Scholar] [CrossRef] [PubMed]
- Marmorstein, N.R.; White, H.R.; Loeber, R.; Stouthamer-Loeber, M. Anxiety as a predictor of age at first use of substances and progression to substance use problems among boys. J. Abnorm. Child. Psychol. 2010, 38, 211–224. [Google Scholar] [CrossRef]
- Fatseas, M.; Serre, F.; Swendsen, J.; Auriacombe, M. Effects of anxiety and mood disorders on craving and substance use among patients with substance use disorder: An ecological momentary assessment study. Drug Alcohol. Depend. 2018, 187, 242–248. [Google Scholar] [CrossRef]
- Morganstern, I.; Chang, G.Q.; Chen, Y.W.; Barson, J.R.; Zhiyu, Y.; Hoebel, B.G.; Leibowitz, S.F. Role of melanin-concentrating hormone in the control of ethanol consumption: Region-specific effects revealed by expression and injection studies. Physiol. Behav. 2010, 101, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Dodge, N.C.; Jacobson, J.L.; Jacobson, S.W. Effects of Fetal Substance Exposure on Offspring Substance Use. Pediatr. Clin. North. Am. 2019, 66, 1149–1161. [Google Scholar] [CrossRef]
- Goldschmidt, L.; Cornelius, M.D.; Day, N.L. Prenatal cigarette smoke exposure and early initiation of multiple substance use. Nicotine Tob. Res. 2012, 14, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Sonon, K.E.; Richardson, G.A.; Cornelius, J.R.; Kim, K.H.; Day, N.L. Prenatal marijuana exposure predicts marijuana use in young adulthood. Neurotoxicol Teratol. 2015, 47, 10–15. [Google Scholar] [CrossRef]
- Goldschmidt, L.; Richardson, G.A.; De Genna, N.M.; Cornelius, M.D.; Day, N.L. Prenatal alcohol exposure and offspring alcohol use and misuse at 22 years of age: A prospective longitudinal study. Neurotoxicol Teratol. 2019, 71, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Poon, K.; Leibowitz, S.F. Consumption of Substances of Abuse during Pregnancy Increases Consumption in Offspring: Possible Underlying Mechanisms. Front. Nutr. 2016, 3, 11. [Google Scholar] [CrossRef]
- Faas, A.E.; Sponton, E.D.; Moya, P.R.; Molina, J.C. Differential responsiveness to alcohol odor in human neonates: Effects of maternal consumption during gestation. Alcohol. 2000, 22, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Hannigan, J.H.; Chiodo, L.M.; Sokol, R.J.; Janisse, J.; Delaney-Black, V. Prenatal alcohol exposure selectively enhances young adult perceived pleasantness of alcohol odors. Physiol. Behav. 2015, 148, 71–77. [Google Scholar] [CrossRef]
- Bonomo, Y.A.; Bowes, G.; Coffey, C.; Carlin, J.B.; Patton, G.C. Teenage drinking and the onset of alcohol dependence: A cohort study over seven years. Addiction 2004, 99, 1520–1528. [Google Scholar] [CrossRef] [PubMed]
- Rockville, M. NSDUH Series H-46; HHS: Rockville, MD, USA, 2013; HHS Publication No. (SMA) 13-4795. [Google Scholar]
- Karkhanis, A.; Holleran, K.M.; Jones, S.R. Dynorphin/Kappa Opioid Receptor Signaling in Preclinical Models of Alcohol, Drug, and Food Addiction. Int. Rev. Neurobiol. 2017, 136, 53–88. [Google Scholar] [CrossRef] [PubMed]
- Lalanne, L.; Ayranci, G.; Kieffer, B.L.; Lutz, P.E. The kappa opioid receptor: From addiction to depression, and back. Front. Psychiatry 2014, 5, 170. [Google Scholar] [CrossRef]
- Wang, X.; Dow-Edwards, D.; Anderson, V.; Minkoff, H.; Hurd, Y. Discrete opioid gene expression impairment in the human fetal brain associated with maternal marijuana use. Pharmacogenomics J. 2006, 6, 255–264. [Google Scholar] [CrossRef]
- Zhou, Y.; Kreek, M.J. Involvement of Activated Brain Stress Responsive Systems in Excessive and "Relapse" Alcohol Drinking in Rodent Models: Implications for Therapeutics. J. Pharmacol. Exp. Ther. 2018, 366, 9–20. [Google Scholar] [CrossRef]
- Anderson, R.I.; Becker, H.C. Role of the Dynorphin/Kappa Opioid Receptor System in the Motivational Effects of Ethanol. Alcohol. Clin. Exp. Res. 2017, 41, 1402–1418. [Google Scholar] [CrossRef]
- DiNieri, J.A.; Wang, X.; Szutorisz, H.; Spano, S.M.; Kaur, J.; Casaccia, P.; Dow-Edwards, D.; Hurd, Y.L. Maternal cannabis use alters ventral striatal dopamine D2 gene regulation in the offspring. Biol. Psychiatry 2011, 70, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Thannickal, T.C.; John, J.; Shan, L.; Swaab, D.F.; Wu, M.-F.; Ramanathan, L.; McGregor, R.; Chew, K.-T.; Cornford, M.; Yamanaka, A. Opiates increase the number of hypocretin-producing cells in human and mouse brain and reverse cataplexy in a mouse model of narcolepsy. Sci. Transl. Med. 2018, 10, eaao4953. [Google Scholar] [CrossRef]
- Gyawali, U.; James, M.H. Sleep disturbance in substance use disorders: The orexin (hypocretin) system as an emerging pharmacological target. Neuropyschopharm 2023, 48, 228–229. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Zhu, Y.; Qin, J. Human brain organoid-on-a-chip to model prenatal nicotine exposure. Lab. Chip 2018, 18, 851–860. [Google Scholar] [CrossRef] [PubMed]
- Ware, A.L.; Long, X.; Lebel, C. Functional connectivity of the attention networks is altered and relates to neuropsychological outcomes in children with prenatal alcohol exposure. Dev. Cogn. Neurosci. 2021, 48, 100951. [Google Scholar] [CrossRef] [PubMed]
- Gross, L.A.; Moore, E.M.; Wozniak, J.R.; Coles, C.D.; Kable, J.A.; Sowell, E.R.; Jones, K.L.; Riley, E.P.; Mattson, S.N.; CIFASD. Neural correlates of verbal memory in youth with heavy prenatal alcohol exposure. Brain Imaging Behav. 2018, 12, 806–822. [Google Scholar] [CrossRef]
- Bailey, C.D.C.; Gerlai, R.; Cameron, N.M.; Marcolin, M.L.; McCormick, C.M. Preclinical methodological approaches investigating of the effects of alcohol on perinatal and adolescent neurodevelopment. Neurosci. Biobehav. Rev. 2020, 116, 436–451. [Google Scholar] [CrossRef]
- Ellenbroek, B.; Youn, J. Rodent models in neuroscience research: Is it a rat race? Dis. Model. Mech. 2016, 9, 1079–1087. [Google Scholar] [CrossRef]
- Clancy, B.; Darlington, R.B.; Finlay, B.L. Translating developmental time across mammalian species. Neuroscience 2001, 105, 7–17. [Google Scholar] [CrossRef]
- Winzer-Serhan, U.H. Long-term consequences of maternal smoking and developmental chronic nicotine exposure. Front. Biosci. 2008, 13, 636–649. [Google Scholar] [CrossRef]
- Bayer, S.A. Timetable of neurogenesis in the human brain based on experimentally determined patterns in the rat. Neurotoxicol 1993, 14, 83–144. [Google Scholar]
- Polli, F.S.; Kohlmeier, K.A. Prenatal Nicotine Exposure in Rodents: Why Are There So Many Variations in Behavioral Outcomes? Nicotine Tob. Res. 2020, 22, 1694–1710. [Google Scholar] [CrossRef]
- Marquardt, K.; Brigman, J.L. The impact of prenatal alcohol exposure on social, cognitive and affective behavioral domains: Insights from rodent models. Alcohol. 2016, 51, 1–15. [Google Scholar] [CrossRef]
- Ciafre, S.; Ferraguti, G.; Greco, A.; Polimeni, A.; Ralli, M.; Ceci, F.M.; Ceccanti, M.; Fiore, M. Alcohol as an early life stressor: Epigenetics, metabolic, neuroendocrine and neurobehavioral implications. Neurosci. Biobehav. Rev. 2020, 118, 654–668. [Google Scholar] [CrossRef] [PubMed]
- Motamedi, S.; Amleshi, R.S.; Javar, B.A.; Shams, P.; Kohlmeier, K.A.; Shabani, M. Cannabis during pregnancy: A way to transfer an impairment to later life. Birth Defects Res. 2023, 115, 1327–1344. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.Q.; Karatayev, O.; Halkina, V.; Edelstien, J.; Ramirez, E.; Leibowitz, S.F. Hypothalamic CCL2/CCR2 Chemokine System: Role in Sexually Dimorphic Effects of Maternal Ethanol Exposure on Melanin-Concentrating Hormone and Behavior in Adolescent Offspring. J. Neurosci. 2018, 38, 9072–9090. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.R.; Johnson, J.M.; Varlinskaya, E.I. Increased ethanol intake is associated with social anxiety in offspring exposed to ethanol on gestational day 12. Behav. Brain Res. 2020, 393, 112766. [Google Scholar] [CrossRef]
- Kim, P.; Park, J.H.; Choi, C.S.; Choi, I.; Joo, S.H.; Kim, M.K.; Kim, S.Y.; Kim, K.C.; Park, S.H.; Kwon, K.J.; et al. Effects of ethanol exposure during early pregnancy in hyperactive, inattentive and impulsive behaviors and MeCP2 expression in rodent offspring. Neurochem. Res. 2013, 38, 620–631. [Google Scholar] [CrossRef]
- Fish, E.W.; Wieczorek, L.A.; Rumple, A.; Suttie, M.; Moy, S.S.; Hammond, P.; Parnell, S.E. The enduring impact of neurulation stage alcohol exposure: A combined behavioral and structural neuroimaging study in adult male and female C57BL/6J mice. Behav. Brain Res. 2018, 338, 173–184. [Google Scholar] [CrossRef]
- Wang, R.; Shen, Y.L.; Hausknecht, K.A.; Chang, L.; Haj-Dahmane, S.; Vezina, P.; Shen, R.Y. Prenatal ethanol exposure increases risk of psychostimulant addiction. Behav. Brain Res. 2019, 356, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.C.; Spear, N.E.; Spear, L.P.; Mennella, J.A.; Lewis, M.J. The International society for developmental psychobiology 39th annual meeting symposium: Alcohol and development: Beyond fetal alcohol syndrome. Dev. Psychobiol. 2007, 49, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Hellemans, K.G.; Sliwowska, J.H.; Verma, P.; Weinberg, J. Prenatal alcohol exposure: Fetal programming and later life vulnerability to stress, depression and anxiety disorders. Neurosci. Biobehav. Rev. 2010, 34, 791–807. [Google Scholar] [CrossRef] [PubMed]
- Laucht, M.; Becker, K.; Blomeyer, D.; Schmidt, M.H. Novelty seeking involved in mediating the association between the dopamine D4 receptor gene exon III polymorphism and heavy drinking in male adolescents: Results from a high-risk community sample. Biol. Psychiatry 2007, 61, 87–92. [Google Scholar] [CrossRef]
- Masse, L.C.; Tremblay, R.E. Behavior of boys in kindergarten and the onset of substance use during adolescence. Arch. Gen. Psychiatry 1997, 54, 62–68. [Google Scholar] [CrossRef]
- Alati, R.; Al Mamun, A.; Williams, G.; O’Callaghan, M.; Najman, J.; Bor, W. In utero alcohol exposure and prediction of alcohol disorders in early adulthood: A birth cohort study. Arch. Gen. Psychiatry 2006, 63, 1009–1016. [Google Scholar] [CrossRef]
- Fernie, G.; Cole, J.C.; Goudie, A.J.; Field, M. Risk-taking but not response inhibition or delay discounting predict alcohol consumption in social drinkers. Drug Alcohol. Depend. 2010, 112, 54–61. [Google Scholar] [CrossRef]
- Petrelli, B.; Weinberg, J.; Hicks, G.G. Effects of prenatal alcohol exposure (PAE): Insights into FASD using mouse models of PAE. Biochem. Cell Biol. 2018, 96, 131–147. [Google Scholar] [CrossRef]
- Subbanna, S.; Basavarajappa, B.S. Binge-like Prenatal Ethanol Exposure Causes Impaired Cellular Differentiation in the Embryonic Forebrain and Synaptic and Behavioral Defects in Adult Mice. Brain Sci. 2022, 12, 793. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.; Chotro, M.G. Ethanol-induced preferences or aversions as a function of age in preweanling rats. Behav. Neurosci. 2006, 120, 710–718. [Google Scholar] [CrossRef]
- March, S.M.; Abate, P.; Spear, N.E.; Molina, J.C. Fetal exposure to moderate ethanol doses: Heightened operant responsiveness elicited by ethanol-related reinforcers. Alcohol. Clin. Exp. Res. 2009, 33, 1981–1993. [Google Scholar] [CrossRef]
- De Genna, N.M.; Goldschmidt, L.; Day, N.L.; Cornelius, M.D. Prenatal tobacco exposure, maternal postnatal nicotine dependence and adolescent risk for nicotine dependence: Birth cohort study. Neurotoxicol Teratol. 2017, 61, 128–132. [Google Scholar] [CrossRef]
- Duko, B.; Pereira, G.; Tait, R.J.; Nyadanu, S.D.; Betts, K.; Alati, R. Prenatal Tobacco Exposure and the Risk of Tobacco Smoking and Dependence in Offspring: A Systematic Review and Meta-Analysis. Drug Alcohol. Depend. 2021, 227, 108993. [Google Scholar] [CrossRef] [PubMed]
- Mantella, N.M.; Kent, P.F.; Youngentob, S.L. Fetal nicotine exposure increases preference for nicotine odor in early postnatal and adolescent, but not adult, rats. PLoS ONE 2013, 8, e84989. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.Q.; Karatayev, O.; Leibowitz, S.F. Prenatal exposure to nicotine stimulates neurogenesis of orexigenic peptide-expressing neurons in hypothalamus and amygdala. J. Neurosci. 2013, 33, 13600–13611. [Google Scholar] [CrossRef]
- LeSage, M.G.; Gustaf, E.; Dufek, M.B.; Pentel, P.R. Effects of maternal intravenous nicotine administration on locomotor behavior in pre-weanling rats. Pharmacol. Biochem. Behav. 2006, 85, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Chung, S.; Noh, J. Maternal Nicotine Exposure During Late Gestation and Lactation Increases Anxiety-Like and Impulsive Decision-Making Behavior in Adolescent Offspring of Rat. Toxicol. Res. 2016, 32, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Roncero, C.; Valriberas-Herrero, I.; Mezzatesta-Gava, M.; Villegas, J.L.; Aguilar, L.; Grau-Lopez, L. Cannabis use during pregnancy and its relationship with fetal developmental outcomes and psychiatric disorders. A systematic review. Reprod. Health 2020, 17, 25. [Google Scholar] [CrossRef]
- Tirado-Munoz, J.; Lopez-Rodriguez, A.B.; Fonseca, F.; Farre, M.; Torrens, M.; Viveros, M.P. Effects of cannabis exposure in the prenatal and adolescent periods: Preclinical and clinical studies in both sexes. Front. Neuroendocr. 2020, 57, 100841. [Google Scholar] [CrossRef]
- Lallai, V.; Manca, L.; Sherafat, Y.; Fowler, C.D. Effects of Prenatal Nicotine, THC, or Co-Exposure on Cognitive Behaviors in Adolescent Male and Female Rats. Nicotine Tob. Res. 2022, 24, 1150–1160. [Google Scholar] [CrossRef] [PubMed]
- Frau, R.; Miczan, V.; Traccis, F.; Aroni, S.; Pongor, C.I.; Saba, P.; Serra, V.; Sagheddu, C.; Fanni, S.; Congiu, M.; et al. Prenatal THC exposure produces a hyperdopaminergic phenotype rescued by pregnenolone. Nat. Neurosci. 2019, 22, 1975–1985. [Google Scholar] [CrossRef]
- Del Ciampo, L.A.; Del Ciampo, I.R.L. Cannabis use in adolescence: Physical and functional repercussions. Int. J. Med. Rev. Case Rep. 2020, 3, 761. [Google Scholar] [CrossRef]
- Spano, M.S.; Ellgren, M.; Wang, X.; Hurd, Y.L. Prenatal cannabis exposure increases heroin seeking with allostatic changes in limbic enkephalin systems in adulthood. Biol. Psychiatry 2007, 61, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Neale, M.C.; Prescott, C.A.; Kendler, K.S. A twin study of early cannabis use and subsequent use and abuse/dependence of other illicit drugs. Psychol. Med. 2004, 34, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Lynskey, M.T.; Heath, A.C.; Bucholz, K.K.; Slutske, W.S.; Madden, P.A.; Nelson, E.C.; Statham, D.J.; Martin, N.G. Escalation of drug use in early-onset cannabis users vs co-twin controls. JAMA 2003, 289, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Day, N.L.; Goldschmidt, L.; Thomas, C.A. Prenatal marijuana exposure contributes to the prediction of marijuana use at age 14. Addiction 2006, 101, 1313–1322. [Google Scholar] [CrossRef]
- Alhowail, A. Mechanisms Underlying Cognitive Impairment Induced by Prenatal Alcohol Exposure. Brain Sci. 2022, 12, 1667. [Google Scholar] [CrossRef]
- Chung, D.D.; Pinson, M.R.; Bhenderu, L.S.; Lai, M.S.; Patel, R.A.; Miranda, R.C. Toxic and teratogenic effects of prenatal alcohol exposure on fetal development, adolescence, and adulthood. Int. J. Mol. Sci. 2021, 22, 8785. [Google Scholar] [CrossRef]
- Chang, G.Q.; Karatayev, O.; Liang, S.C.; Barson, J.R.; Leibowitz, S.F. Prenatal ethanol exposure stimulates neurogenesis in hypothalamic and limbic peptide systems: Possible mechanism for offspring ethanol overconsumption. Neuroscience 2012, 222, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Mooney, S.M.; Miller, M.W. Prenatal exposure to ethanol affects postnatal neurogenesis in thalamus. Exp. Neurol. 2010, 223, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Gil-Mohapel, J.; Boehme, F.; Patten, A.; Cox, A.; Kainer, L.; Giles, E.; Brocardo, P.S.; Christie, B.R. Altered adult hippocampal neuronal maturation in a rat model of fetal alcohol syndrome. Brain Res. 2011, 1384, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.Q.; Yasmin, N.; Collier, A.D.; Karatayev, O.; Khalizova, N.; Onoichenco, A.; Fam, M.; Albeg, A.S.; Campbell, S.; Leibowitz, S.F. Fibroblast growth factor 2: Role in prenatal alcohol-induced stimulation of hypothalamic peptide neurons. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 116, 110536. [Google Scholar] [CrossRef]
- Abate, P.; Hernandez-Fonseca, K.; Reyes-Guzman, A.C.; Barbosa-Luna, I.G.; Mendez, M. Prenatal ethanol exposure alters met-enkephalin expression in brain regions related with reinforcement: Possible mechanism for ethanol consumption in offspring. Behav. Brain Res. 2014, 274, 194–204. [Google Scholar] [CrossRef]
- Bordner, K.; Deak, T. Endogenous opioids as substrates for ethanol intake in the neonatal rat: The impact of prenatal ethanol exposure on the opioid family in the early postnatal period. Physiol. Behav. 2015, 148, 100–110. [Google Scholar] [CrossRef]
- Jacobson, L.H.; Hoyer, D.; de Lecea, L. Hypocretins (orexins): The ultimate translational neuropeptides. J. Intern. Med. 2022, 291, 533–556. [Google Scholar] [CrossRef]
- Moorman, D.E. The hypocretin/orexin system as a target for excessive motivation in alcohol use disorders. Psychopharmacology 2018, 235, 1663–1680. [Google Scholar] [CrossRef]
- Wee, S.; Koob, G.F. The role of the dynorphin–κ opioid system in the reinforcing effects of drugs of abuse. Psychopharmacology 2010, 210, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Louth, E.L.; Luctkar, H.D.; Heney, K.A.; Bailey, C.D. Developmental ethanol exposure alters the morphology of mouse prefrontal neurons in a layer-specific manner. Brain Res. 2018, 1678, 94–105. [Google Scholar] [CrossRef]
- Boychuk, C.R.; Fuller, D.D.; Hayward, L.F. Sex differences in heart rate variability during sleep following prenatal nicotine exposure in rat pups. Behav. Brain Res. 2011, 219, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Barson, J.R.; Poon, K.; Ho, H.T.; Alam, M.I.; Sanzalone, L.; Leibowitz, S.F. Substance P in the anterior thalamic paraventricular nucleus: Promotion of ethanol drinking in response to orexin from the hypothalamus. Addict. Biol. 2017, 22, 58–69. [Google Scholar] [CrossRef]
- Morgan, A.J.; Harrod, S.B.; Lacy, R.T.; Stanley, E.M.; Fadel, J.R. Intravenous prenatal nicotine exposure increases orexin expression in the lateral hypothalamus and orexin innervation of the ventral tegmental area in adult male rats. Drug Alcohol. Depend. 2013, 132, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Reid, H.M.O.; Lysenko-Martin, M.R.; Snowden, T.M.; Thomas, J.D.; Christie, B.R. A Systematic Review of the Effects of Perinatal Alcohol Exposure and Perinatal Marijuana Exposure on Adult Neurogenesis in the Dentate Gyrus. Alcohol. Clin. Exp. Res. 2020, 44, 1164–1174. [Google Scholar] [CrossRef] [PubMed]
- Rubino, T.; Vigano, D.; Realini, N.; Guidali, C.; Braida, D.; Capurro, V.; Castiglioni, C.; Cherubino, F.; Romualdi, P.; Candeletti, S.; et al. Chronic delta 9-tetrahydrocannabinol during adolescence provokes sex-dependent changes in the emotional profile in adult rats: Behavioral and biochemical correlates. Neuropsychopharmacology 2008, 33, 2760–2771. [Google Scholar] [CrossRef] [PubMed]
- Manikandan, P.; Sarmah, S.; Marrs, J.A. Ethanol Effects on Early Developmental Stages Studied Using the Zebrafish. Biomedicines 2022, 10, 2555. [Google Scholar] [CrossRef]
- Schaidhauer, F.G.; Caetano, H.A.; da Silva, G.P.; da Silva, R.S. Contributions of Zebrafish Studies to the Behavioural Consequences of Early Alcohol Exposure: A Systematic Review. Curr. Neuropharmacol. 2022, 20, 579–593. [Google Scholar] [CrossRef]
- Facciol, A.; Tsang, B.; Gerlai, R. Alcohol exposure during embryonic development: An opportunity to conduct systematic developmental time course analyses in zebrafish. Neurosci. Biobehav. Rev. 2019, 98, 185–193. [Google Scholar] [CrossRef]
- Wronikowska, O.; Michalak, A.; Skalicka-Wozniak, K.; Crawford, A.D.; Budzynska, B. Fishing for a deeper understanding of nicotine effects using zebrafish behavioural models. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 98, 109826. [Google Scholar] [CrossRef]
- Licitra, R.; Marchese, M.; Naef, V.; Ogi, A.; Martinelli, M.; Kiferle, C.; Fronte, B.; Santorelli, F.M. A Review on the Bioactivity of Cannabinoids on Zebrafish Models: Emphasis on Neurodevelopment. Biomedicines 2022, 10, 1820. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Klee, E.W.; Schneider, H.; Clark, K.J.; Cousin, M.A.; Ebbert, J.O.; Hooten, W.M.; Karpyak, V.M.; Warner, D.O.; Ekker, S.C. Zebrafish: A model for the study of addiction genetics. Hum. Genet. 2012, 131, 977–1008. [Google Scholar] [CrossRef] [PubMed]
- Weaver, I.C.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Abozaid, A.; Hung, J.; Tsang, B.; Motlana, K.; Al-Ani, R.; Gerlai, R. Behavioral effects of acute ethanol in larval zebrafish (D. rerio) depend on genotype and volume of experimental well. Prog. Neuropsychopharmacol. Biol. Psychiatry 2022, 112, 110411. [Google Scholar] [CrossRef] [PubMed]
- Facciol, A.; Bailleul, C.; Nguyen, S.; Chatterjee, D.; Gerlai, R. Developmental stage-dependent deficits induced by embryonic ethanol exposure in zebrafish: A neurochemical analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 99, 109859. [Google Scholar] [CrossRef]
- Agues-Barbosa, T.; de Souza, A.M.; Gomes-de-Lima, J.N.; Luchiari, A.C. Long-term behavioral alterations following embryonic alcohol exposure in three zebrafish populations. Neurotoxicology 2023, 96, 174–183. [Google Scholar] [CrossRef]
- Facciol, A.; Marawi, T.; Syed, E.; Gerlai, R. Age-dependent effects of embryonic ethanol exposure on anxiety-like behaviours in young zebrafish: A genotype comparison study. Pharmacol. Biochem. Behav. 2022, 214, 173342. [Google Scholar] [CrossRef]
- Mahabir, S.; Chatterjee, D.; Gerlai, R. Strain dependent neurochemical changes induced by embryonic alcohol exposure in zebrafish. Neurotoxicol Teratol. 2014, 41, 1–7. [Google Scholar] [CrossRef]
- Fernandes, Y.; Gerlai, R. Long-term behavioral changes in response to early developmental exposure to ethanol in zebrafish. Alcohol. Clin. Exp. Res. 2009, 33, 601–609. [Google Scholar] [CrossRef]
- Ellertsdottir, E.; Lenard, A.; Blum, Y.; Krudewig, A.; Herwig, L.; Affolter, M.; Belting, H.G. Vascular morphogenesis in the zebrafish embryo. Dev. Biol. 2010, 341, 56–65. [Google Scholar] [CrossRef]
- Lovely, C.B.; Nobles, R.D.; Eberhart, J.K. Developmental age strengthens barriers to ethanol accumulation in zebrafish. Alcohol. 2014, 48, 595–602. [Google Scholar] [CrossRef]
- Lovely, C.B. Quantification of Ethanol Levels in Zebrafish Embryos Using Head Space Gas Chromatography. J. Vis. Exp. 2020, 156, e60766. [Google Scholar] [CrossRef]
- Pinheiro-da-Silva, J.; Agues-Barbosa, T.; Luchiari, A.C. Embryonic Exposure to Ethanol Increases Anxiety-Like Behavior in Fry Zebrafish. Alcohol. Alcohol. 2020, 55, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Boa-Amponsem, O.; Zhang, C.; Mukhopadhyay, S.; Ardrey, I.; Cole, G.J. Ethanol and cannabinoids interact to alter behavior in a zebrafish fetal alcohol spectrum disorder model. Birth Defects Res. 2019, 111, 775–788. [Google Scholar] [CrossRef]
- Burd, L.; Blair, J.; Dropps, K. Prenatal alcohol exposure, blood alcohol concentrations and alcohol elimination rates for the mother, fetus and newborn. J. Perinatol. 2012, 32, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Klee, E.W.; Ebbert, J.O.; Schneider, H.; Hurt, R.D.; Ekker, S.C. Zebrafish for the study of the biological effects of nicotine. Nicotine Tob. Res. 2011, 13, 301–312. [Google Scholar] [CrossRef]
- Demin, K.A.; Meshalkina, D.A.; Kysil, E.V.; Antonova, K.A.; Volgin, A.D.; Yakovlev, O.A.; Alekseeva, P.A.; Firuleva, M.M.; Lakstygal, A.M.; de Abreu, M.S.; et al. Zebrafish models relevant to studying central opioid and endocannabinoid systems. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 86, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Borrego-Soto, G.; Eberhart, J.K. Embryonic Nicotine Exposure Disrupts Adult Social Behavior and Craniofacial Development in Zebrafish. Toxics 2022, 10, 612. [Google Scholar] [CrossRef]
- Albadri, S.; Del Bene, F.; Revenu, C. Genome editing using CRISPR/Cas9-based knock-in approaches in zebrafish. Methods 2017, 121–122, 77–85. [Google Scholar] [CrossRef]
- Klem, J.R.; Gray, R.; Lovely, C.B. The Zebrafish Tol2 System: A Modular and Flexible Gateway-Based Transgenesis Approach. J. Vis. Exp. 2022, 189, e64679. [Google Scholar] [CrossRef]
- Kemmler, C.L.; Moran, H.R.; Murray, B.F.; Scoresby, A.; Klem, J.R.; Eckert, R.L.; Lepovsky, E.; Bertho, S.; Nieuwenhuize, S.; Burger, S.; et al. Next-generation plasmids for transgenesis in zebrafish and beyond. Development 2023, 150, dev201531. [Google Scholar] [CrossRef]
- Rieger, S.; Wang, F.; Sagasti, A. Time-lapse imaging of neural development: Zebrafish lead the way into the fourth dimension. Genesis 2011, 49, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Cararo, J.H.; Rico, E.P. Long-lasting implications of embryonic exposure to alcohol: Insights from zebrafish research. Dev. Neurobiol. 2022, 82, 29–40. [Google Scholar] [CrossRef]
- Guo, S. Linking genes to brain, behavior and neurological diseases: What can we learn from zebrafish? Genes. Brain Behav. 2004, 3, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, Y.; Lovely, C.B. Zebrafish models of fetal alcohol spectrum disorders. Genesis 2021, 59, e23460. [Google Scholar] [CrossRef]
- Facciol, A.; Gerlai, R. Zebrafish Shoaling, Its Behavioral and Neurobiological Mechanisms, and Its Alteration by Embryonic Alcohol Exposure: A Review. Front. Behav. Neurosci. 2020, 14, 572175. [Google Scholar] [CrossRef] [PubMed]
- Seguin, D.; Gerlai, R. Fetal alcohol spectrum disorders: Zebrafish in the analysis of the milder and more prevalent form of the disease. Behav. Brain Res. 2018, 352, 125–132. [Google Scholar] [CrossRef]
- Tran, S. Acute and chronic alcohol effects in zebrafish. In Behavioral and Neural Genetics of Zebrafish; Elsevier: Amsterdam, The Netherlands, 2020; pp. 325–341. [Google Scholar]
- Lockwood, B.; Bjerke, S.; Kobayashi, K.; Guo, S. Acute effects of alcohol on larval zebrafish: A genetic system for large-scale screening. Pharmacol. Biochem. Behav. 2004, 77, 647–654. [Google Scholar] [CrossRef]
- Cousin, M.A.; Ebbert, J.O.; Wiinamaki, A.R.; Urban, M.D.; Argue, D.P.; Ekker, S.C.; Klee, E.W. Larval zebrafish model for FDA-approved drug repositioning for tobacco dependence treatment. PLoS ONE 2014, 9, e90467. [Google Scholar] [CrossRef]
- Akhtar, M.T.; Ali, S.; Rashidi, H.; van der Kooy, F.; Verpoorte, R.; Richardson, M.K. Developmental effects of cannabinoids on zebrafish larvae. Zebrafish 2013, 10, 283–293. [Google Scholar] [CrossRef]
- Schneider, H.; Pearson, A.; Harris, D.; Krause, S.; Tucker, A.; Gardner, K.; Chinyanya, K. Identification of nicotine-seeking and avoiding larval zebrafish using a new three-choice behavioral assay. Front. Mol. Neurosci. 2023, 16, 1112927. [Google Scholar] [CrossRef]
- Mech, A.M.; Merteroglu, M.; Sealy, I.M.; Teh, M.T.; White, R.J.; Havelange, W.; Brennan, C.H.; Busch-Nentwich, E.M. Behavioral and Gene Regulatory Responses to Developmental Drug Exposures in Zebrafish. Front. Psychiatry 2021, 12, 795175. [Google Scholar] [CrossRef]
- Chen, B. Effects of acute nicotine on larval zebrafish exploratory behavior in a complex environment. Spring 2015, 125. [Google Scholar] [CrossRef]
- Razmara, P.; Zaveri, D.; Thannhauser, M.; Ali, D.W. Acute effect of Delta-9-tetrahydrocannabinol on neuromuscular transmission and locomotive behaviors in larval zebrafish. J. Neurophysiol. 2023, 129, 833–842. [Google Scholar] [CrossRef]
- Hayton, S.J.; Mahoney, M.K.; Olmstead, M.C. Behavioral traits predicting alcohol drinking in outbred rats: An investigation of anxiety, novelty seeking, and cognitive flexibility. Alcohol. Clin. Exp. Res. 2012, 36, 594–603. [Google Scholar] [CrossRef]
- Pena-Oliver, Y.; Giuliano, C.; Economidou, D.; Goodlett, C.R.; Robbins, T.W.; Dalley, J.W.; Everitt, B.J. Alcohol-Preferring Rats Show Goal Oriented Behaviour to Food Incentives but Are Neither Sign-Trackers Nor Impulsive. PLoS ONE 2015, 10, e0131016. [Google Scholar] [CrossRef]
- Karatayev, O.; Barson, J.R.; Carr, A.J.; Baylan, J.; Chen, Y.W.; Leibowitz, S.F. Predictors of ethanol consumption in adult Sprague-Dawley rats: Relation to hypothalamic peptides that stimulate ethanol intake. Alcohol 2010, 44, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Lees, B.; Mewton, L.; Stapinski, L.A.; Teesson, M.; Squeglia, L.M. Association of prenatal alcohol exposure with preadolescent alcohol sipping in the ABCD study(R). Drug Alcohol. Depend. 2020, 214, 108187. [Google Scholar] [CrossRef] [PubMed]
- Collier, A.D.; Min, S.S.; Campbell, S.D.; Roberts, M.Y.; Camidge, K.; Leibowitz, S.F. Maternal ethanol consumption before paternal fertilization: Stimulation of hypocretin neurogenesis and ethanol intake in zebrafish offspring. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 96, 109728. [Google Scholar] [CrossRef]
- Fernandes, P.; Monteiro, S.M.; Venancio, C.; Felix, L. 24-Epibrassinolide protects against ethanol-induced behavioural teratogenesis in zebrafish embryo. Chem. Biol. Interact. 2020, 328, 109193. [Google Scholar] [CrossRef]
- Leech, S.L.; Larkby, C.A.; Day, R.; Day, N.L. Predictors and correlates of high levels of depression and anxiety symptoms among children at age 10. J. Am. Acad. Child. Adolesc. Psychiatry 2006, 45, 223–230. [Google Scholar] [CrossRef]
- Bailey, J.M.; Oliveri, A.N.; Zhang, C.; Frazier, J.M.; Mackinnon, S.; Cole, G.J.; Levin, E.D. Long-term behavioral impairment following acute embryonic ethanol exposure in zebrafish. Neurotoxicol Teratol. 2015, 48, 1–8. [Google Scholar] [CrossRef]
- Wang, R.; Martin, C.D.; Lei, A.L.; Hausknecht, K.A.; Ishiwari, K.; Richards, J.B.; Haj-Dahmane, S.; Shen, R.Y. Prenatal Ethanol Exposure Leads to Attention Deficits in Both Male and Female Rats. Front. Neurosci. 2020, 14, 12. [Google Scholar] [CrossRef]
- Allan, A.M.; Chynoweth, J.; Tyler, L.A.; Caldwell, K.K. A mouse model of prenatal ethanol exposure using a voluntary drinking paradigm. Alcohol. Clin. Exp. Res. 2003, 27, 2009–2016. [Google Scholar] [CrossRef]
- Sterling, M.E.; Chang, G.Q.; Karatayev, O.; Chang, S.Y.; Leibowitz, S.F. Effects of embryonic ethanol exposure at low doses on neuronal development, voluntary ethanol consumption and related behaviors in larval and adult zebrafish: Role of hypothalamic orexigenic peptides. Behav. Brain Res. 2016, 304, 125–138. [Google Scholar] [CrossRef]
- Dodge, N.C.; Jacobson, J.L.; Lundahl, L.H.; Jacobson, S.W. Prenatal alcohol exposure and attention-deficit/hyperactivity disorder independently predict greater substance use in young adulthood. Alcohol 2023, 47, 1143–1155. [Google Scholar] [CrossRef]
- Jansone, K.; Eichler, A.; Fasching, P.A.; Kornhuber, J.; Kaiser, A.; Millenet, S.; Banaschewski, T.; Nees, F.; On Behalf of The Imac-Mind, C. Association of Maternal Smoking during Pregnancy with Neurophysiological and ADHD-Related Outcomes in School-Aged Children. Int. J. Env. Res. Public. Health 2023, 20, 4716. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhang, X.; Xu, Y.; Spencer, T.J.; Biederman, J.; Bhide, P.G. Prenatal nicotine exposure mouse model showing hyperactivity, reduced cingulate cortex volume, reduced dopamine turnover, and responsiveness to oral methylphenidate treatment. J. Neurosci. 2012, 32, 9410–9418. [Google Scholar] [CrossRef] [PubMed]
- Moylan, S.; Gustavson, K.; Overland, S.; Karevold, E.B.; Jacka, F.N.; Pasco, J.A.; Berk, M. The impact of maternal smoking during pregnancy on depressive and anxiety behaviors in children: The Norwegian Mother and Child Cohort Study. BMC Med. 2015, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Ashford, J.; van Lier, P.A.; Timmermans, M.; Cuijpers, P.; Koot, H.M. Prenatal smoking and internalizing and externalizing problems in children studied from childhood to late adolescence. J. Am. Acad. Child. Adolesc. Psychiatry 2008, 47, 779–787. [Google Scholar] [CrossRef]
- Santiago, S.E.; Huffman, K.J. Prenatal nicotine exposure increases anxiety and modifies sensorimotor integration behaviors in adult female mice. Neurosci. Res. 2014, 79, 41–51. [Google Scholar] [CrossRef]
- Polli, F.S.; Scharff, M.B.; Ipsen, T.H.; Aznar, S.; Kohlmeier, K.A.; Andreasen, J.T. Prenatal nicotine exposure in mice induces sex-dependent anxiety-like behavior, cognitive deficits, hyperactivity, and changes in the expression of glutamate receptor associated-genes in the prefrontal cortex. Pharmacol. Biochem. Behav. 2020, 195, 172951. [Google Scholar] [CrossRef]
- Bryden, D.W.; Burton, A.C.; Barnett, B.R.; Cohen, V.J.; Hearn, T.N.; Jones, E.A.; Kariyil, R.J.; Kunin, A.; Kwak, S.I.; Lee, J.; et al. Prenatal Nicotine Exposure Impairs Executive Control Signals in Medial Prefrontal Cortex. Neuropsychopharmacology 2016, 41, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.M.; McCarthy, D.M.; Schatschneider, C.; Trupiano, M.X.; Jones, S.K.; Kalluri, A.; Bhide, P.G. Effects of Developmental Nicotine Exposure on Frontal Cortical GABA-to-Non-GABA Neuron Ratio and Novelty-Seeking Behavior. Cereb. Cortex 2020, 30, 1830–1842. [Google Scholar] [CrossRef] [PubMed]
- Vaglenova, J.; Birru, S.; Pandiella, N.M.; Breese, C.R. An assessment of the long-term developmental and behavioral teratogenicity of prenatal nicotine exposure. Behav. Brain Res. 2004, 150, 159–170. [Google Scholar] [CrossRef]
- Goldschmidt, L.; Day, N.L.; Richardson, G.A. Effects of prenatal marijuana exposure on child behavior problems at age 10. Neurotoxicol Teratol. 2000, 22, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Mereu, G.; Fa, M.; Ferraro, L.; Cagiano, R.; Antonelli, T.; Tattoli, M.; Ghiglieri, V.; Tanganelli, S.; Gessa, G.L.; Cuomo, V. Prenatal exposure to a cannabinoid agonist produces memory deficits linked to dysfunction in hippocampal long-term potentiation and glutamate release. Proc. Natl. Acad. Sci. USA 2003, 100, 4915–4920. [Google Scholar] [CrossRef] [PubMed]
- Rompala, G.; Nomura, Y.; Hurd, Y.L. Maternal cannabis use is associated with suppression of immune gene networks in placenta and increased anxiety phenotypes in offspring. Proc. Natl. Acad. Sci. USA 2021, 118, e2106115118. [Google Scholar] [CrossRef]
- Weimar, H.V.; Wright, H.R.; Warrick, C.R.; Brown, A.M.; Lugo, J.M.; Freels, T.G.; McLaughlin, R.J. Long-term effects of maternal cannabis vapor exposure on emotional reactivity, social behavior, and behavioral flexibility in offspring. Neuropharmacology 2020, 179, 108288. [Google Scholar] [CrossRef]
- Goldschmidt, L.; Richardson, G.A.; Willford, J.; Day, N.L. Prenatal marijuana exposure and intelligence test performance at age 6. J. Am. Acad. Child. Adolesc. Psychiatry 2008, 47, 254–263. [Google Scholar] [CrossRef]
- Day, N.L.; Leech, S.L.; Goldschmidt, L. The effects of prenatal marijuana exposure on delinquent behaviors are mediated by measures of neurocognitive functioning. Neurotoxicol Teratol. 2011, 33, 129–136. [Google Scholar] [CrossRef]
- Scheyer, A.F.; Borsoi, M.; Pelissier-Alicot, A.L.; Manzoni, O.J.J. Perinatal THC exposure via lactation induces lasting alterations to social behavior and prefrontal cortex function in rats at adulthood. Neuropsychopharmacology 2020, 45, 1826–1833. [Google Scholar] [CrossRef] [PubMed]
- Boa-Amponsem, O.; Zhang, C.; Burton, D.; Williams, K.P.; Cole, G.J. Ethanol and Cannabinoids Regulate Zebrafish GABAergic Neuron Development and Behavior in a Sonic Hedgehog and Fibroblast Growth Factor-Dependent Mechanism. Alcohol. Clin. Exp. Res. 2020, 44, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.F.; Boa-Amponsem, O.M.; Dixon, M.S.; Hopkins, M.J.; Herbin, T.A.; Toney, S.; Tarpley, M.; Rodriguez, B.V.; Fish, E.W.; Parnell, S.E.; et al. Pharmacological activation of the Sonic hedgehog pathway with a Smoothened small molecule agonist ameliorates the severity of alcohol-induced morphological and behavioral birth defects in a zebrafish model of fetal alcohol spectrum disorder. J. Neurosci. Res. 2022, 100, 1585–1601. [Google Scholar] [CrossRef]
- Eddins, D.; Cerutti, D.; Williams, P.; Linney, E.; Levin, E.D. Zebrafish provide a sensitive model of persisting neurobehavioral effects of developmental chlorpyrifos exposure: Comparison with nicotine and pilocarpine effects and relationship to dopamine deficits. Neurotoxicol Teratol. 2010, 32, 99–108. [Google Scholar] [CrossRef]
- Sterling, M.E.; Karatayev, O.; Chang, G.Q.; Algava, D.B.; Leibowitz, S.F. Model of voluntary ethanol intake in zebrafish: Effect on behavior and hypothalamic orexigenic peptides. Behav. Brain Res. 2015, 278, 29–39. [Google Scholar] [CrossRef]
- Bailone, R.L.; Fukushima, H.C.S.; de Aguiar, L.K.; Borra, R.C. The endocannabinoid system in zebrafish and its potential to study the effects of Cannabis in humans. Lab. Anim. Res. 2022, 38, 5. [Google Scholar] [CrossRef]
- Ali, S.; Champagne, D.L.; Alia, A.; Richardson, M.K. Large-scale analysis of acute ethanol exposure in zebrafish development: A critical time window and resilience. PLoS ONE 2011, 6, e20037. [Google Scholar] [CrossRef] [PubMed]
- Joya, X.; Garcia-Algar, O.; Vall, O.; Pujades, C. Transient exposure to ethanol during zebrafish embryogenesis results in defects in neuronal differentiation: An alternative model system to study FASD. PLoS ONE 2014, 9, e112851. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Yao, F.; Chen, X.; Wang, N.; Wang, H.; Chang, H.E.; Yuan, Z.; Wu, B. Ethanol reduces neural precursor cells and inhibits neuronal and glial differentiation in zebrafish embryos. Nan Fang Yi Ke Da Xue Xue Bao 2014, 34, 1555–1561. [Google Scholar]
- Collier, A.D.; Khalizova, N.; Chang, G.Q.; Min, S.; Campbell, S.; Gulati, G.; Leibowitz, S.F. Involvement of Cxcl12a/Cxcr4b Chemokine System in Mediating the Stimulatory Effect of Embryonic Ethanol Exposure on Neuronal Density in Zebrafish Hypothalamus. Alcohol. Clin. Exp. Res. 2020, 44, 2519–2535. [Google Scholar] [CrossRef]
- Collier, A.D.; Yasmin, N.; Khalizova, N.; Campbell, S.; Onoichenco, A.; Fam, M.; Albeg, A.S.; Leibowitz, S.F. Sexually dimorphic and asymmetric effects of embryonic ethanol exposure on hypocretin/orexin neurons as related to behavioral changes in zebrafish. Sci. Rep. 2021, 11, 16078. [Google Scholar] [CrossRef]
- Du, W.; Chen, X.; Shi, M.; Bian, F.; Zhao, Z. Ethanol affects behavior and HPA axis activity during development in zebrafish larvae. Sci. Rep. 2020, 10, 21402. [Google Scholar] [CrossRef] [PubMed]
- Buske, C.; Gerlai, R. Early embryonic ethanol exposure impairs shoaling and the dopaminergic and serotoninergic systems in adult zebrafish. Neurotoxicology Teratol. 2011, 33, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.Q.; Collier, A.D.; Karatayev, O.; Gulati, G.; Boorgu, D.; Leibowitz, S.F. Moderate Prenatal Ethanol Exposure Stimulates CXCL12/CXCR4 Chemokine System in Radial Glia Progenitor Cells in Hypothalamic Neuroepithelium and Peptide Neurons in Lateral Hypothalamus of the Embryo and Postnatal Offspring. Alcohol. Clin. Exp. Res. 2020, 44, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.Q.; Karatayev, O.; Leibowitz, S.F. Prenatal exposure to ethanol stimulates hypothalamic CCR2 chemokine receptor system: Possible relation to increased density of orexigenic peptide neurons and ethanol drinking in adolescent offspring. Neuroscience 2015, 310, 163–175. [Google Scholar] [CrossRef]
- Bodnar, T.S.; Raineki, C.; Wertelecki, W.; Yevtushok, L.; Plotka, L.; Granovska, I.; Zymak-Zakutnya, N.; Pashtepa, A.; Wells, A.; Honerkamp-Smith, G.; et al. Immune network dysregulation associated with child neurodevelopmental delay: Modulatory role of prenatal alcohol exposure. J. Neuroinflammation 2020, 17, 39. [Google Scholar] [CrossRef]
- Menelaou, E.; Svoboda, K.R. Secondary motoneurons in juvenile and adult zebrafish: Axonal pathfinding errors caused by embryonic nicotine exposure. J. Comp. Neurol. 2009, 512, 305–322. [Google Scholar] [CrossRef]
- Cerpa, V.J.; Aylwin, M.d.l.L.O.; Beltrán-Castillo, S.; Bravo, E.U.; Llona, I.R.; Richerson, G.B.; Eugenín, J.L. The alteration of neonatal raphe neurons by prenatal–perinatal nicotine. Meaning for sudden infant death syndrome. Am. J. Respir. Cell Mol. Biol. 2015, 53, 489–499. [Google Scholar] [CrossRef]
- He, X.; Lu, J.; Dong, W.; Jiao, Z.; Zhang, C.; Yu, Y.; Zhang, Z.; Wang, H.; Xu, D. Prenatal nicotine exposure induces HPA axis-hypersensitivity in offspring rats via the intrauterine programming of up-regulation of hippocampal GAD67. Arch. Toxicol. 2017, 91, 3927–3943. [Google Scholar] [CrossRef]
- Wang, X.; Dow-Edwards, D.; Anderson, V.; Minkoff, H.; Hurd, Y.L. In utero marijuana exposure associated with abnormal amygdala dopamine D2 gene expression in the human fetus. Biol. Psychiatry 2004, 56, 909–915. [Google Scholar] [CrossRef]
- de Salas-Quiroga, A.; Diaz-Alonso, J.; Garcia-Rincon, D.; Remmers, F.; Vega, D.; Gomez-Canas, M.; Lutz, B.; Guzman, M.; Galve-Roperh, I. Prenatal exposure to cannabinoids evokes long-lasting functional alterations by targeting CB1 receptors on developing cortical neurons. Proc. Natl. Acad. Sci. USA 2015, 112, 13693–13698. [Google Scholar] [CrossRef]
- Amin, M.R.; Ahmed, K.T.; Ali, D.W. Cannabinoid receptor 2 (Cb2r) mediates cannabinol (CBN) induced developmental defects in zebrafish. Sci. Rep. 2022, 12, 20251. [Google Scholar] [CrossRef]
- Fried, P.A.; Watkinson, B. Visuoperceptual functioning differs in 9- to 12-year olds prenatally exposed to cigarettes and marihuana. Neurotoxicol Teratol. 2000, 22, 11–20. [Google Scholar] [CrossRef]
- Rivkin, M.J.; Davis, P.E.; Lemaster, J.L.; Cabral, H.J.; Warfield, S.K.; Mulkern, R.V.; Robson, C.D.; Rose-Jacobs, R.; Frank, D.A. Volumetric MRI study of brain in children with intrauterine exposure to cocaine, alcohol, tobacco, and marijuana. Pediatrics 2008, 121, 741–750. [Google Scholar] [CrossRef]
- Kanyo, R.; Amin, M.R.; Locskai, L.F.; Bouvier, D.D.; Olthuis, A.M.; Allison, W.T.; Ali, D.W. Medium-throughput zebrafish optogenetic platform identifies deficits in subsequent neural activity following brief early exposure to cannabidiol and Delta(9)-tetrahydrocannabinol. Sci. Rep. 2021, 11, 11515. [Google Scholar] [CrossRef]
- Holman, P.J.; Raineki, C.; Chao, A.; Grewal, R.; Haghighat, S.; Fung, C.; Morgan, E.; Ellis, L.; Yu, W.; Weinberg, J. Altered social recognition memory and hypothalamic neuropeptide expression in adolescent male and female rats following prenatal alcohol exposure and/or early-life adversity. Psychoneuroendocrinology 2021, 126, 105146. [Google Scholar] [CrossRef] [PubMed]
- Morganstern, I.; Gulati, G.; Leibowitz, S.F. Role of melanin-concentrating hormone in drug use disorders. Brain Res. 2020, 1741, 146872. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.I.; Moorman, D.E.; Becker, H.C. Contribution of Dynorphin and Orexin Neuropeptide Systems to the Motivational Effects of Alcohol. Handb. Exp. Pharmacol. 2018, 248, 473–503. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, A.J.; Cowen, M.S.; Yang, H.J.; Chen, F.; Oldfield, B. The orexin system regulates alcohol-seeking in rats. Br. J. Pharmacol. 2006, 148, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Van Camp, K.A.; Baggerman, G.; Blust, R.; Husson, S.J. Peptidomics of the zebrafish Danio rerio: In search for neuropeptides. J. Proteom. 2017, 150, 290–296. [Google Scholar] [CrossRef]
- Akinrinade, I.; Kareklas, K.; Teles, M.C.; Reis, T.K.; Gliksberg, M.; Petri, G.; Levkowitz, G.; Oliveira, R.F. Evolutionarily conserved role of oxytocin in social fear contagion in zebrafish. Science 2023, 379, 1232–1237. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.; Rihel, J.; Prober, D.A. Neuropeptide Y Regulates Sleep by Modulating Noradrenergic Signaling. Curr. Biol. 2017, 27, 3796–3811.e5. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, I.; Levitas-Djerbi, T.; Appelbaum, L. The Hypocretin/Orexin Neuronal Networks in Zebrafish. Curr. Top. Behav. Neurosci. 2017, 33, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Berman, J.R.; Skariah, G.; Maro, G.S.; Mignot, E.; Mourrain, P. Characterization of two melanin-concentrating hormone genes in zebrafish reveals evolutionary and physiological links with the mammalian MCH system. J. Comp. Neurol. 2009, 517, 695–710. [Google Scholar] [CrossRef]
- Corradi, L.; Bruzzone, M.; Maschio, M.D.; Sawamiphak, S.; Filosa, A. Hypothalamic Galanin-producing neurons regulate stress in zebrafish through a peptidergic, self-inhibitory loop. Curr. Biol. 2022, 32, 1497–1510.e5. [Google Scholar] [CrossRef]
- Podlasz, P.; Jakimiuk, A.; Kasica-Jarosz, N.; Czaja, K.; Wasowicz, K. Neuroanatomical Localization of Galanin in Zebrafish Telencephalon and Anticonvulsant Effect of Galanin Overexpression. ACS Chem. Neurosci. 2018, 9, 3049–3059. [Google Scholar] [CrossRef]
- Nunez, V.G.; Sarmiento, R.G.; Rodrıguez, R. Characterization of zebrafish proenkephalin reveals novel opioid sequences. Mol. Brain Res. 2003, 114, 31–39. [Google Scholar] [CrossRef]
- Gonzalez-Nunez, V.; de Velasco, E.M.F.; Arsequell, G.; Valencia, G.; Rodríguez, R. Identification of dynorphin a from zebrafish: A comparative study with mammalian dynorphin A. Neuroscience 2007, 144, 675–684. [Google Scholar] [CrossRef]
- Cui, C.; Wang, L.F.; Huang, S.B.; Zhao, P.; Chen, Y.Q.; Wu, Y.B.; Qiao, C.M.; Zhao, W.J.; Shen, Y.Q. Adequate expression of neuropeptide Y is essential for the recovery of zebrafish motor function following spinal cord injury. Exp. Neurol. 2021, 345, 113831. [Google Scholar] [CrossRef]
- Prober, D.A.; Rihel, J.; Onah, A.A.; Sung, R.J.; Schier, A.F. Hypocretin/orexin overexpression induces an insomnia-like phenotype in zebrafish. J. Neurosci. 2006, 26, 13400–13410. [Google Scholar] [CrossRef]
- Shiozaki, K.; Kawabe, M.; Karasuyama, K.; Kurachi, T.; Hayashi, A.; Ataka, K.; Iwai, H.; Takeno, H.; Hayasaka, O.; Kotani, T.; et al. Neuropeptide Y deficiency induces anxiety-like behaviours in zebrafish (Danio rerio). Sci. Rep. 2020, 10, 5913. [Google Scholar] [CrossRef] [PubMed]
- Kawabe, M.; Hayashi, A.; Komatsu, M.; Inui, A.; Shiozaki, K. Ninjinyoeito improves anxiety behavior in neuropeptide Y deficient zebrafish. Neuropeptides 2021, 87, 102136. [Google Scholar] [CrossRef]
- Appelbaum, L.; Wang, G.X.; Maro, G.S.; Mori, R.; Tovin, A.; Marin, W.; Yokogawa, T.; Kawakami, K.; Smith, S.J.; Gothilf, Y.; et al. Sleep-wake regulation and hypocretin-melatonin interaction in zebrafish. Proc. Natl. Acad. Sci. USA 2009, 106, 21942–21947. [Google Scholar] [CrossRef]
- Chen, A.; Singh, C.; Oikonomou, G.; Prober, D.A. Genetic analysis of histamine signaling in larval zebrafish sleep. Eneuro 2017, 4. [Google Scholar] [CrossRef]
- Yokobori, E.; Azuma, M.; Nishiguchi, R.; Kang, K.S.; Kamijo, M.; Uchiyama, M.; Matsuda, K. Neuropeptide Y stimulates food intake in the Zebrafish, Danio rerio. J. Neuroendocr. 2012, 24, 766–773. [Google Scholar] [CrossRef]
- Khor, B.S.; Jamil, M.F.; Adenan, M.I.; Shu-Chien, A.C. Mitragynine attenuates withdrawal syndrome in morphine-withdrawn zebrafish. PLoS ONE 2011, 6, e28340. [Google Scholar] [CrossRef] [PubMed]
- Skofitsch, G.; Jacobowitz, D.M.; Zamir, N. Immunohistochemical localization of a melanin concentrating hormone-like peptide in the rat brain. Brain Res. Bull. 1985, 15, 635–649. [Google Scholar] [CrossRef] [PubMed]
- de Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.; Gautvik, V.T.; Bartlett, F.S., 2nd; et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. USA 1998, 95, 322–327. [Google Scholar] [CrossRef]
- Coffey, C.M.; Solleveld, P.A.; Fang, J.; Roberts, A.K.; Hong, S.K.; Dawid, I.B.; Laverriere, C.E.; Glasgow, E. Novel oxytocin gene expression in the hindbrain is induced by alcohol exposure: Transgenic zebrafish enable visualization of sensitive neurons. PLoS ONE 2013, 8, e53991. [Google Scholar] [CrossRef] [PubMed]
- Buckley, D.M.; Sidik, A.; Kar, R.D.; Eberhart, J.K. Differentially sensitive neuronal subpopulations in the central nervous system and the formation of hindbrain heterotopias in ethanol-exposed zebrafish. Birth Defects Res. 2019, 111, 700–713. [Google Scholar] [CrossRef]
- Boric, K.; Orio, P.; Viéville, T.; Whitlock, K. Quantitative analysis of cell migration using optical flow. PLoS ONE 2013, 8, e69574. [Google Scholar] [CrossRef]
- Marjonen, H.; Sierra, A.; Nyman, A.; Rogojin, V.; Grohn, O.; Linden, A.M.; Hautaniemi, S.; Kaminen-Ahola, N. Early maternal alcohol consumption alters hippocampal DNA methylation, gene expression and volume in a mouse model. PLoS ONE 2015, 10, e0124931. [Google Scholar] [CrossRef] [PubMed]
- Gordon, H.W. Laterality of Brain Activation for Risk Factors of Addiction. Curr. Drug Abus. Rev. 2016, 9, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Murphy, F.C.; Nimmo-Smith, I.; Lawrence, A.D. Functional neuroanatomy of emotions: A meta-analysis. Cogn. Affect. Behav. Neurosci. 2003, 3, 207–233. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Bellido, R.; Barreto-Valer, K.; Rodriguez, R.E. Substance P mRNA expression during zebrafish development: Influence of mu opioid receptor and cocaine. Neuroscience 2013, 242, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Sagi, D.; de Lecea, L.; Appelbaum, L. Heterogeneity of Hypocretin/Orexin Neurons. Front. Neurol. Neurosci. 2021, 45, 61–74. [Google Scholar] [CrossRef]
- Liu, J.; Merkle, F.T.; Gandhi, A.V.; Gagnon, J.A.; Woods, I.G.; Chiu, C.N.; Shimogori, T.; Schier, A.F.; Prober, D.A. Evolutionarily conserved regulation of hypocretin neuron specification by Lhx9. Development 2015, 142, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Muschamp, J.W.; Hollander, J.A.; Thompson, J.L.; Voren, G.; Hassinger, L.C.; Onvani, S.; Kamenecka, T.M.; Borgland, S.L.; Kenny, P.J.; Carlezon, W.A., Jr. Hypocretin (orexin) facilitates reward by attenuating the antireward effects of its cotransmitter dynorphin in ventral tegmental area. Proc. Natl. Acad. Sci. USA 2014, 111, E1648–E1655. [Google Scholar] [CrossRef]
- Li, Y.; van den Pol, A.N. Differential target-dependent actions of coexpressed inhibitory dynorphin and excitatory hypocretin/orexin neuropeptides. J. Neurosci. 2006, 26, 13037–13047. [Google Scholar] [CrossRef]
- Bruchas, M.; Land, B.; Chavkin, C. The dynorphin/kappa opioid system as a modulator of stress-induced and pro-addictive behaviors. Brain Res. 2010, 1314, 44–55. [Google Scholar] [CrossRef]
- Carroll, F.I.; Carlezon, W.A., Jr. Development of kappa opioid receptor antagonists. J. Med. Chem. 2013, 56, 2178–2195. [Google Scholar] [CrossRef] [PubMed]
- Olateju, O.I.; Bhagwandin, A.; Ihunwo, A.O.; Manger, P.R. Changes in the Cholinergic, Catecholaminergic, Orexinergic and Serotonergic Structures Forming Part of the Sleep Systems of Adult Mice Exposed to Intrauterine Alcohol. Front. Neuroanat. 2017, 11, 110. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Naik, V.; Orzabal, M.; Lunde-Young, R.; Ramadoss, J. Morphological alteration in rat hippocampal neuronal dendrites following chronic binge prenatal alcohol exposure. Brain Res. 2021, 1768, 147587. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, G.; Altermatt, M.; Zhang, R.W.; Coughlin, G.M.; Montz, C.; Gradinaru, V.; Prober, D.A. The Serotonergic Raphe Promote Sleep in Zebrafish and Mice. Neuron 2019, 103, 686–701.e8. [Google Scholar] [CrossRef]
- Imperatore, R.; D’Angelo, L.; Safari, O.; Motlagh, H.A.; Piscitelli, F.; de Girolamo, P.; Cristino, L.; Varricchio, E.; di Marzo, V.; Paolucci, M. Overlapping Distribution of Orexin and Endocannabinoid Receptors and Their Functional Interaction in the Brain of Adult Zebrafish. Front. Neuroanat. 2018, 12, 62. [Google Scholar] [CrossRef]
- von Trotha, J.W.; Vernier, P.; Bally-Cuif, L. Emotions and motivated behavior converge on an amygdala-like structure in the zebrafish. Eur. J. Neurosci. 2014, 40, 3302–3315. [Google Scholar] [CrossRef]
- Goeke, C.M.; Roberts, M.L.; Hashimoto, J.G.; Finn, D.A.; Guizzetti, M. Neonatal Ethanol and Choline Treatments Alter the Morphology of Developing Rat Hippocampal Pyramidal Neurons in Opposite Directions. Neuroscience 2018, 374, 13–24. [Google Scholar] [CrossRef]
- Svoboda, K.R.; Vijayaraghavan, S.; Tanguay, R.L. Nicotinic receptors mediate changes in spinal motoneuron development and axonal pathfinding in embryonic zebrafish exposed to nicotine. J. Neurosci. 2002, 22, 10731–10741. [Google Scholar] [CrossRef]
- Amin, M.R.; Ahmed, K.T.; Ali, D.W. Early Exposure to THC Alters M-Cell Development in Zebrafish Embryos. Biomedicines 2020, 8, 5. [Google Scholar] [CrossRef]
- Ahmed, K.T.; Amin, M.R.; Shah, P.; Ali, D.W. Motor neuron development in zebrafish is altered by brief (5-hr) exposures to THC (∆(9)-tetrahydrocannabinol) or CBD (cannabidiol) during gastrulation. Sci. Rep. 2018, 8, 10518. [Google Scholar] [CrossRef]
- Memi, F.; Abe, P.; Cariboni, A.; MacKay, F.; Parnavelas, J.G.; Stumm, R. CXC Chemokine Receptor 7 (CXCR7) Affects the Migration of GnRH Neurons by Regulating CXCL12 Availability. J. Neurosci. 2013, 33, 17527–17537. [Google Scholar] [CrossRef]
- Palevitch, O.; Abraham, E.; Borodovsky, N.; Levkowitz, G.; Zohar, Y.; Gothilf, Y. Cxcl12a–Cxcr4b signaling is important for proper development of the forebrain GnRH system in zebrafish. Gen. Comp. Endocrinol. 2010, 165, 262–268. [Google Scholar] [CrossRef]
- Wu, Y.; Peng, H.; Cui, M.; Whitney, N.P.; Huang, Y.; Zheng, J.C. CXCL12 increases human neural progenitor cell proliferation through Akt-1/FOXO3a signaling pathway. J. Neurochem. 2009, 109, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Teicher, B.A.; Fricker, S.P. CXCL12 (SDF-1)/CXCR4 pathway in cancer. Clin. Cancer Res. 2010, 16, 2927–2931. [Google Scholar] [CrossRef]
- Watson, A.E.S.; Goodkey, K.; Footz, T.; Voronova, A. Regulation of CNS precursor function by neuronal chemokines. Neurosci. Lett. 2020, 715, 134533. [Google Scholar] [CrossRef]
- Chédotal, A. Roles of axon guidance molecules in neuronal wiring in the developing spinal cord. Nat. Rev. Neurosci. 2019, 20, 380–396. [Google Scholar] [CrossRef]
- Festa, L.K.; Irollo, E.; Platt, B.J.; Tian, Y.; Floresco, S.; Meucci, O. CXCL12-induced rescue of cortical dendritic spines and cognitive flexibility. Elife 2020, 9, e49717. [Google Scholar] [CrossRef]
- Li, J.; Wu, Y.; Chen, P.; Huang, X.; Liu, Y.; Peng, M.; Wu, R.-Q. CXCL12 promotes spinal nerve regeneration and functional recovery after spinal cord injury. Neuroreport 2021, 32, 450–457. [Google Scholar] [CrossRef]
- Even-Chen, O.; Barak, S. The role of fibroblast growth factor 2 in drug addiction. Eur. J. Neurosci. 2019, 50, 2552–2561. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, M.E.; Xu, B.; Lu, B.; Hempstead, B.L. New insights in the biology of BDNF synthesis and release: Implications in CNS function. J. Neurosci. 2009, 29, 12764–12767. [Google Scholar] [CrossRef] [PubMed]
- Traiffort, E.; Angot, E.; Ruat, M. Sonic Hedgehog signaling in the mammalian brain. J. Neurochem. 2010, 113, 576–590. [Google Scholar] [CrossRef]
- Chang, G.-Q.; Karatayev, O.; Kavya, B.D.S.S.; Leibowitz, S.F. CCL2/CCR2 Chemokine System in Embryonic Hypothalamus: Involvement in Sexually Dimorphic Stimulatory Effects of Prenatal Ethanol Exposure on Peptide-Expressing Neurons. Neuroscience 2020, 424, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Valenta, J.P.; Gonzales, R.A. Chronic Intracerebroventricular Infusion of Monocyte Chemoattractant Protein-1 Leads to a Persistent Increase in Sweetened Ethanol Consumption During Operant Self-Administration But Does Not Influence Sucrose Consumption in Long-Evans Rats. Alcohol. Clin. Exp. Res. 2016, 40, 187–195. [Google Scholar] [CrossRef]
- Noor, S.; Milligan, E.D. Lifelong Impacts of Moderate Prenatal Alcohol Exposure on Neuroimmune Function. Front. Immunol. 2018, 9, 1107. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Connelly, K.L.; Unterwald, E.M.; Rawls, S.M. Chemokines and cocaine: CXCR4 receptor antagonist AMD3100 attenuates cocaine place preference and locomotor stimulation in rats. Brain Behav. Immun. 2017, 62, 30–34. [Google Scholar] [CrossRef]
- Burton, D.F.; Zhang, C.; Boa-Amponsem, O.; Mackinnon, S.; Cole, G.J. Long-term behavioral change as a result of acute ethanol exposure in zebrafish: Evidence for a role for sonic hedgehog but not retinoic acid signaling. Neurotoxicol Teratol. 2017, 61, 66–73. [Google Scholar] [CrossRef]
- Mahabir, S.; Chatterjee, D.; Misquitta, K.; Chatterjee, D.; Gerlai, R. Lasting changes induced by mild alcohol exposure during embryonic development in BDNF, NCAM and synaptophysin-positive neurons quantified in adult zebrafish. Eur. J. Neurosci. 2018, 47, 1457–1473. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chen, J.; Sheng, Z.; Lian, W.; Wu, Y.; Liu, M. Embryonic exposure to fentanyl induces behavioral changes and neurotoxicity in zebrafish larvae. PeerJ 2022, 10, e14524. [Google Scholar] [CrossRef]
- Aoto, K.; Shikata, Y.; Higashiyama, D.; Shiota, K.; Motoyama, J. Fetal ethanol exposure activates protein kinase A and impairs Shh expression in prechordal mesendoderm cells in the pathogenesis of holoprosencephaly. Birth Defects Res. A Clin. Mol. Teratol. 2008, 82, 224–231. [Google Scholar] [CrossRef]
- Even-Chen, O.; Herburg, L.; Kefalakes, E.; Urshansky, N.; Grothe, C.; Barak, S. FGF2 is an endogenous regulator of alcohol reward and consumption. Addict. Biol. 2022, 27, e13115. [Google Scholar] [CrossRef]
- Eberhart, J.K.; Parnell, S.E. The Genetics of Fetal Alcohol Spectrum Disorders. Alcohol. Clin. Exp. Res. 2016, 40, 1154–1165. [Google Scholar] [CrossRef] [PubMed]
- Boschen, K.E.; Fish, E.W.; Parnell, S.E. Prenatal alcohol exposure disrupts Sonic hedgehog pathway and primary cilia genes in the mouse neural tube. Reprod. Toxicol. 2021, 105, 136–147. [Google Scholar] [CrossRef]
- Chang, G.Q.; Karatayev, O.; Boorgu, D.; Leibowitz, S.F. CCL2/CCR2 system in neuroepithelial radial glia progenitor cells: Involvement in stimulatory, sexually dimorphic effects of maternal ethanol on embryonic development of hypothalamic peptide neurons. J. Neuroinflammation 2020, 17, 207. [Google Scholar] [CrossRef]
- Roberson, S.; Halpern, M.E. Convergence of signaling pathways underlying habenular formation and axonal outgrowth in zebrafish. Development 2017, 144, 2652–2662. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ojiaku, P.; Cole, G.J. Forebrain and hindbrain development in zebrafish is sensitive to ethanol exposure involving agrin, Fgf, and sonic hedgehog function. Birth Defects Res. A Clin. Mol. Teratol. 2013, 97, 8–27. [Google Scholar] [CrossRef]
- Liran, M.; Rahamim, N.; Ron, D.; Barak, S. Growth Factors and Alcohol Use Disorder. Cold Spring Harb. Perspect. Med. 2020, 10, a039271. [Google Scholar] [CrossRef] [PubMed]
- Arrenberg, A.B.; Del Bene, F.; Baier, H. Optical control of zebrafish behavior with halorhodopsin. Proc. Natl. Acad. Sci. USA 2009, 106, 17968–17973. [Google Scholar] [CrossRef] [PubMed]
- Biechele-Speziale, D.; Camarillo, M.; Martin, N.R.; Biechele-Speziale, J.; Lein, P.J.; Plavicki, J.S. Assessing CaMPARI as new approach methodology for evaluating neurotoxicity. Neurotoxicology 2023, 97, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Wang, Z.; Chai, Y.; Hang, W.; Shang, C.; Yang, W.; Bai, L.; Du, J.; Wang, K.; Wen, Q. Rapid whole brain imaging of neural activity in freely behaving larval zebrafish (Danio rerio). eLife 2017, 6, e28158. [Google Scholar] [CrossRef] [PubMed]
- Volpe, B.A.; Fotino, T.H.; Steiner, A.B. Confocal Microscope-Based Laser Ablation and Regeneration Assay in Zebrafish Interneuromast Cells. J. Vis. Exp. 2020, 159. [Google Scholar] [CrossRef]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.R.; Joung, J.K. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhao, L.; Page-McCaw, P.S.; Chen, W. Zebrafish Genome Engineering Using the CRISPR-Cas9 System. Trends Genet. 2016, 32, 815–827. [Google Scholar] [CrossRef]
- Stainier, D.Y.R.; Raz, E.; Lawson, N.D.; Ekker, S.C.; Burdine, R.D.; Eisen, J.S.; Ingham, P.W.; Schulte-Merker, S.; Yelon, D.; Weinstein, B.M.; et al. Guidelines for morpholino use in zebrafish. PLoS Genet. 2017, 13, e1007000. [Google Scholar] [CrossRef] [PubMed]
- Bedell, V.M.; Westcot, S.E.; Ekker, S.C. Lessons from morpholino-based screening in zebrafish. Brief. Funct. Genom. 2011, 10, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.; Oikonomou, G.; Prober, D.A. Norepinephrine is required to promote wakefulness and for hypocretin-induced arousal in zebrafish. Elife 2015, 4, e07000. [Google Scholar] [CrossRef]
- Heydendael, W.; Sengupta, A.; Beck, S.; Bhatnagar, S. Optogenetic examination identifies a context-specific role for orexins/hypocretins in anxiety-related behavior. Physiol. Behav. 2014, 130, 182–190. [Google Scholar] [CrossRef]
- Eacret, D.; Grafe, L.A.; Dobkin, J.; Gotter, A.L.; Renger, J.J.; Winrow, C.J.; Bhatnagar, S. Orexin signaling during social defeat stress influences subsequent social interaction behaviour and recognition memory. Behav. Brain Res. 2019, 356, 444–452. [Google Scholar] [CrossRef]
- Tyree, S.M.; Jennings, K.J.; Gonzalez, O.C.; Li, S.B.; Nicholson, J.R.; von Heimendahl, M.; de Lecea, L. Optogenetic and pharmacological interventions link hypocretin neurons to impulsivity in mice. Commun. Biol. 2023, 6, 74. [Google Scholar] [CrossRef]
- Fosque, B.F.; Sun, Y.; Dana, H.; Yang, C.T.; Ohyama, T.; Tadross, M.R.; Patel, R.; Zlatic, M.; Kim, D.S.; Ahrens, M.B.; et al. Neural circuits. Labeling of active neural circuits in vivo with designed calcium integrators. Science 2015, 347, 755–760. [Google Scholar] [CrossRef]
- Menelaou, E.; Udvadia, A.J.; Tanguay, R.L.; Svoboda, K.R. Activation of alpha2A-containing nicotinic acetylcholine receptors mediates nicotine-induced motor output in embryonic zebrafish. Eur. J. Neurosci. 2014, 40, 2225–2240. [Google Scholar] [CrossRef]
- Faillace, M.P.; Bernabeu, R.O. Epigenetic mechanisms mediate nicotine-induced reward and behaviour in zebrafish. Curr. Neuropharmacol. 2022, 20, 510. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Drug Intake-Related Behaviors | ||||
|---|---|---|---|---|
| Drug | Behaviors | Human | Rodent | Zebrafish |
| Alcohol | Hyperactivity/Locomotor Activity | ↑ [14] | ↑ [51] ↓ [49] | ↑ [140] ↓ [141] |
| Anxiety/Startle Response | ↑ [142] | ↑ [50] ↑ [61] | ↑ [140] ↓ [143] | |
| Impulsivity | ↑ [11] | ↑ [144] | ||
| Novelty Seeking/Risk Taking/Exploration | - | ↑ [145] ↑ [53] | ↑ [146] | |
| Drug Seeking/Intake | ↑ [147] ↑ [139] | ↑ [49] | ↑ [140] | |
| Nicotine | Hyperactivity/Locomotor Activity | ↑ [148] | ↑ [149] | ↑ [118] |
| Anxiety/Startle Response | ↑ [150] ↑ [151] | ↑ [152] ↑ [153] ↓ [72] | - | |
| Impulsivity | - | ↑ [154] | - | |
| Novelty Seeking/Risk Taking/Exploration | - | ↑ [155] ↓ [156] | - | |
| Drug Seeking/Intake | - | - | - | |
| Cannabis | Hyperactivity/Locomotor Activity | ↑ [157] | ↑ [158] | ↑ [131] |
| Anxiety/Startle Response | ↑ [159] | ↑ [160] | - | |
| Impulsivity | ↑ [161] ↑ [162] | - | - | |
| Novelty Seeking/Risk Taking/Exploration | - | ↑ [163] | ↑ [164] | |
| Drug Seeking/Intake | ↑ [78] | ↑ [75] | - | |
| Neuronal Effects | ||||
|---|---|---|---|---|
| Drug | Neuronal Population | Human | Rodent | Zebrafish |
| Alcohol | Hcrt | - | ↑ Expression [176] ↑ Density [81] ↑ Proliferation [81] ↑ Ectopic Location [5] ↑ Projections [5] ↑ Morphology [5] | ↑ Number [3,173] ↑ Proliferation [7] ↑ Asymmetry [3,173] ↑ Migration [3] ↑ Ectopic Location [3,5] ↑ Subpopulations [6,7] ↑ Projections [5] ↑ Morphology [5] |
| Mch | - | ↑ Expression [176] ↑ Density [177] ↑ Proliferation [177] ↑ Ectopic Location [5] ↑ Subpopulations [84] ↑ Projections [5] ↑ Morphology [5] | - | |
| Dyn | 0 effect [31] | ↑ Expression [86] | ↑ Colocalization w/ Hcrt [7] | |
| KOR | ↓ Expression [31] | - | - | |
| Enk | - | ↑ Density [81] ↑ Proliferation [81] | - | |
| Gal | - | ↑ Density [81] ↑ Proliferation [81] | ↑ Density [146] | |
| Cxcl12 | - | ↑ Expression [176] | ↑ Expression [172] ↑ Density [6,172] ↑ Colocalization w/ Hcrt [6] | |
| Cxcr4 | - | ↑ Expression [176] ↑ Density [176] ↑ Colocalization w/ Hcrt [176] ↑ Colocalization w/ Hcrt [176] ↑ Colocalization w/ Radial Glia [176] | ↑ Expression [172] ↑ Density [6,172] ↑ Colocalization w/ Hcrt [6] | |
| Fgf | ↑ Levels [178] | ↑ Expression ↑ Colocalization w/ Mch [176] | - | |
| Gad 1 | - | - | ↓ Expression [114] | |
| Nicotine | Hcrt | - | ↑ Expression [67] ↑ Density [67] ↑ Proliferation [67] | - |
| Mch | - | ↑ Expression [67] ↑ Density [67] ↑ Proliferation [67] | - | |
| Enk | - | ↑ Expression [67] ↑ Density [67] ↑ Proliferation [67] | - | |
| Motor Neurons | - | - | ↑ Morphology [179] ↑ Axon Pathfinding Errors [179] | |
| Immediate Early Genes | - | ↓ Expression [180] | ↓ Expression [133] | |
| Gad 67 | - | ↑ Expression [181] | - | |
| Human Induced Pluripotent Stem Cells | ↑ Differentiation [37] | - | - | |
| Cannabis | Dyn | 0 Effect [31] | - | - |
| Enk | ↓ Expression [31] | ↑ ↓ Expression [75] | - | |
| KOR | ↓ Expression [31] | ↑ Expression [95] | - | |
| Mu Receptor | ↑ Expression [31] | - | - | |
| CBR1 | 0 Effect [182] | ↓ Expression [183] | ||
| Motor Neurons | - | ↓Development [183] | ↓Branching [184] | |
| Whole Brain | ↓ Brain Size [185] ↓ Brain Volume [186] | - | ↓ Neural Activity [187] | |
| Mauthner Cells | - | - | ↑ Morphology [187] ↓ Axon Diameter [187] | |
| D2 Receptor | ↓ Expression [182] | - | - | |
| Gad 1 | - | - | ↓ Expression [114] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collier, A.D.; Abdulai, A.R.; Leibowitz, S.F. Utility of the Zebrafish Model for Studying Neuronal and Behavioral Disturbances Induced by Embryonic Exposure to Alcohol, Nicotine, and Cannabis. Cells 2023, 12, 2505. https://doi.org/10.3390/cells12202505
Collier AD, Abdulai AR, Leibowitz SF. Utility of the Zebrafish Model for Studying Neuronal and Behavioral Disturbances Induced by Embryonic Exposure to Alcohol, Nicotine, and Cannabis. Cells. 2023; 12(20):2505. https://doi.org/10.3390/cells12202505
Chicago/Turabian StyleCollier, Adam D., Abdul R. Abdulai, and Sarah F. Leibowitz. 2023. "Utility of the Zebrafish Model for Studying Neuronal and Behavioral Disturbances Induced by Embryonic Exposure to Alcohol, Nicotine, and Cannabis" Cells 12, no. 20: 2505. https://doi.org/10.3390/cells12202505
APA StyleCollier, A. D., Abdulai, A. R., & Leibowitz, S. F. (2023). Utility of the Zebrafish Model for Studying Neuronal and Behavioral Disturbances Induced by Embryonic Exposure to Alcohol, Nicotine, and Cannabis. Cells, 12(20), 2505. https://doi.org/10.3390/cells12202505