
Separable Roles for Neur and Ubiquitin in Delta Signalling in the Drosophila CNS Lineages


Abstract
:1. Introduction
2. Materials and Methods
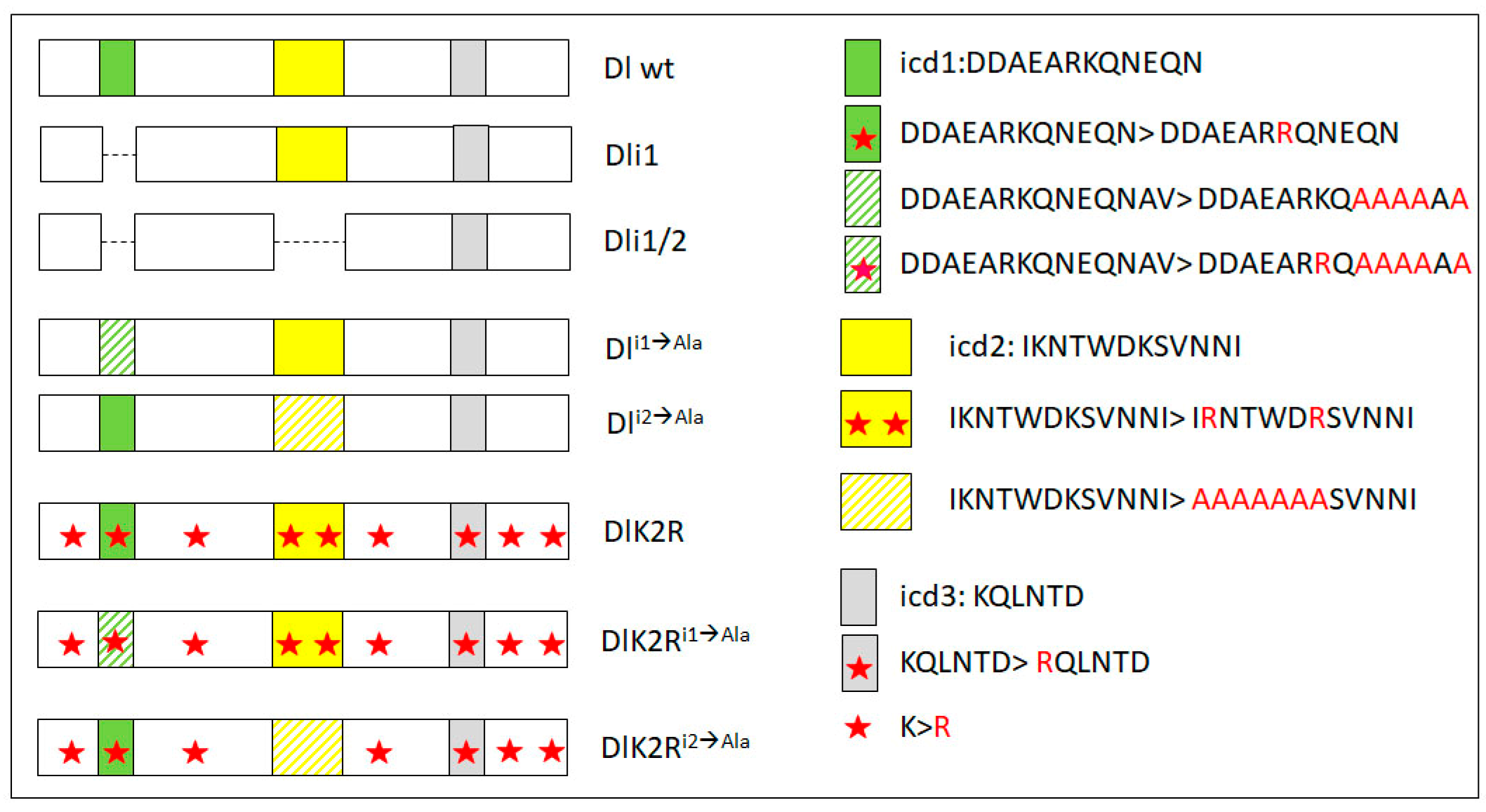
2.1. Generation of Constructs
2.2. Drosophila Stocks and Genetics
2.3. Crosses for Mosaic Analysis
2.4. Crosses for Lateral Inhibition Assay in the Embryonic Neuroectoderm
2.5. Immunohistochemistry—Antibodies
2.6. Confocal Microscopy—Ganglion Mother Cells’ Asymmetric Cell Division (GMC-ACD) Assay Scoring and Image Analysis
2.7. Lateral Inhibition Assay in the Embryonic Neuroectoderm
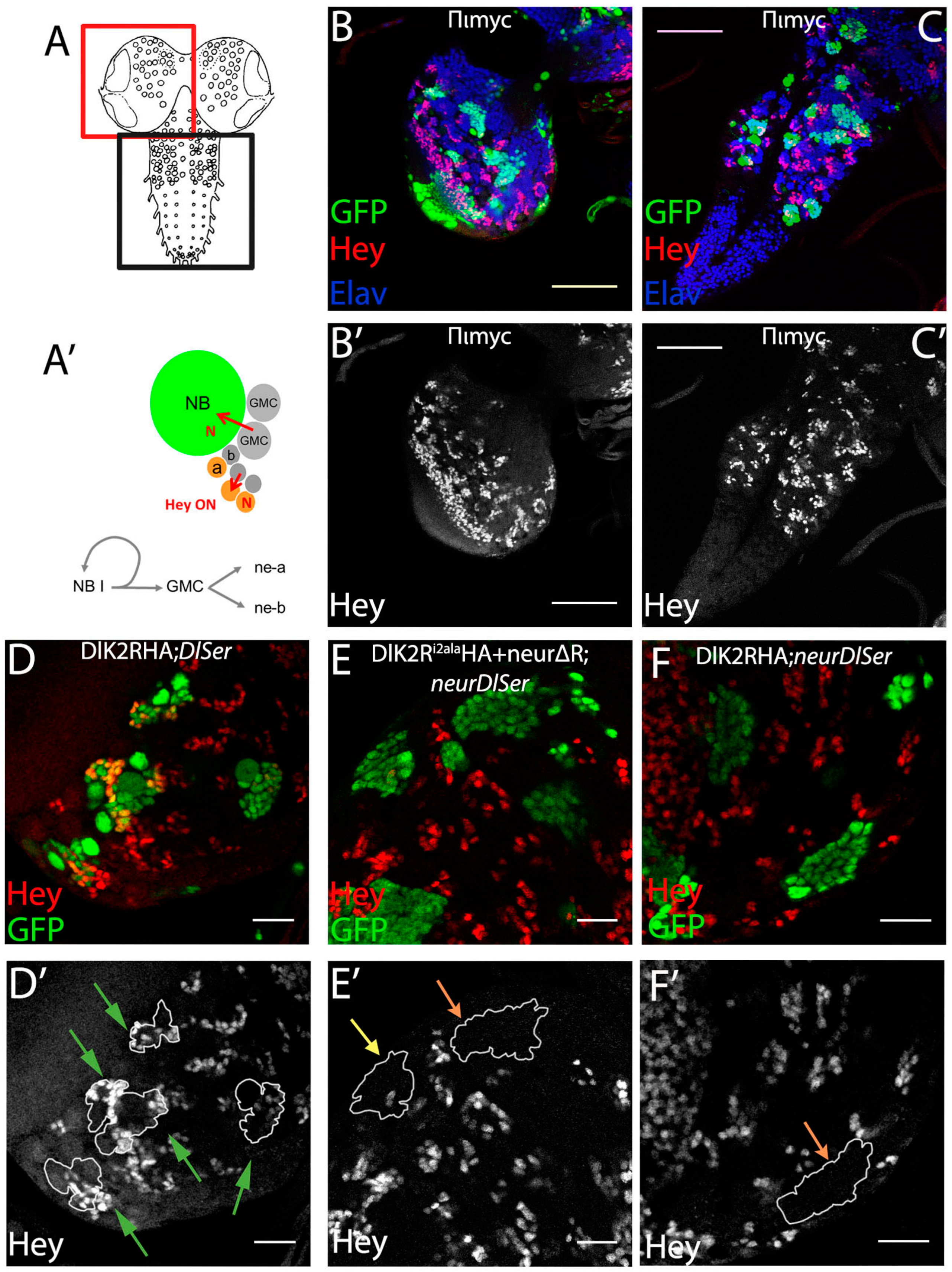
3. Results
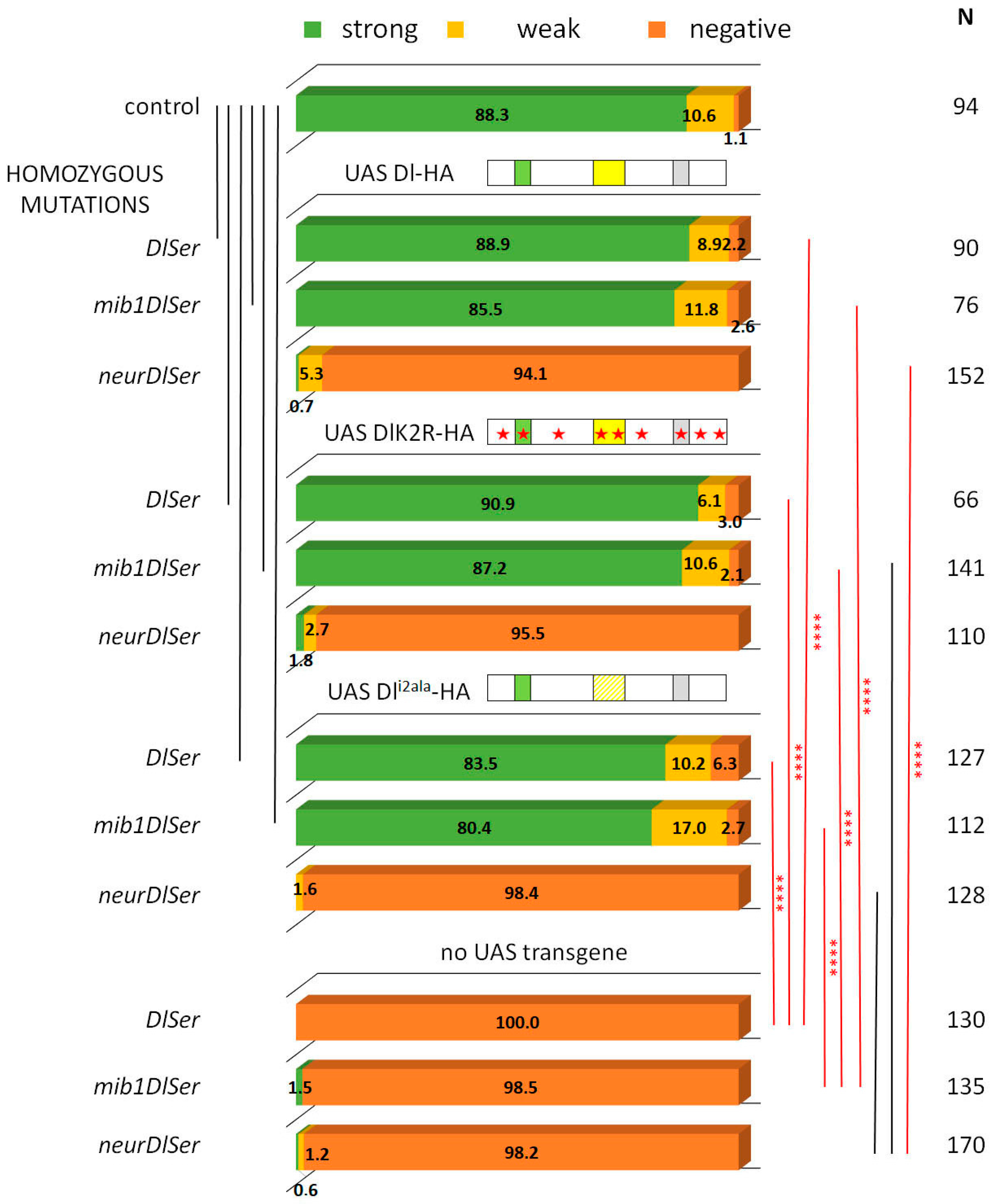
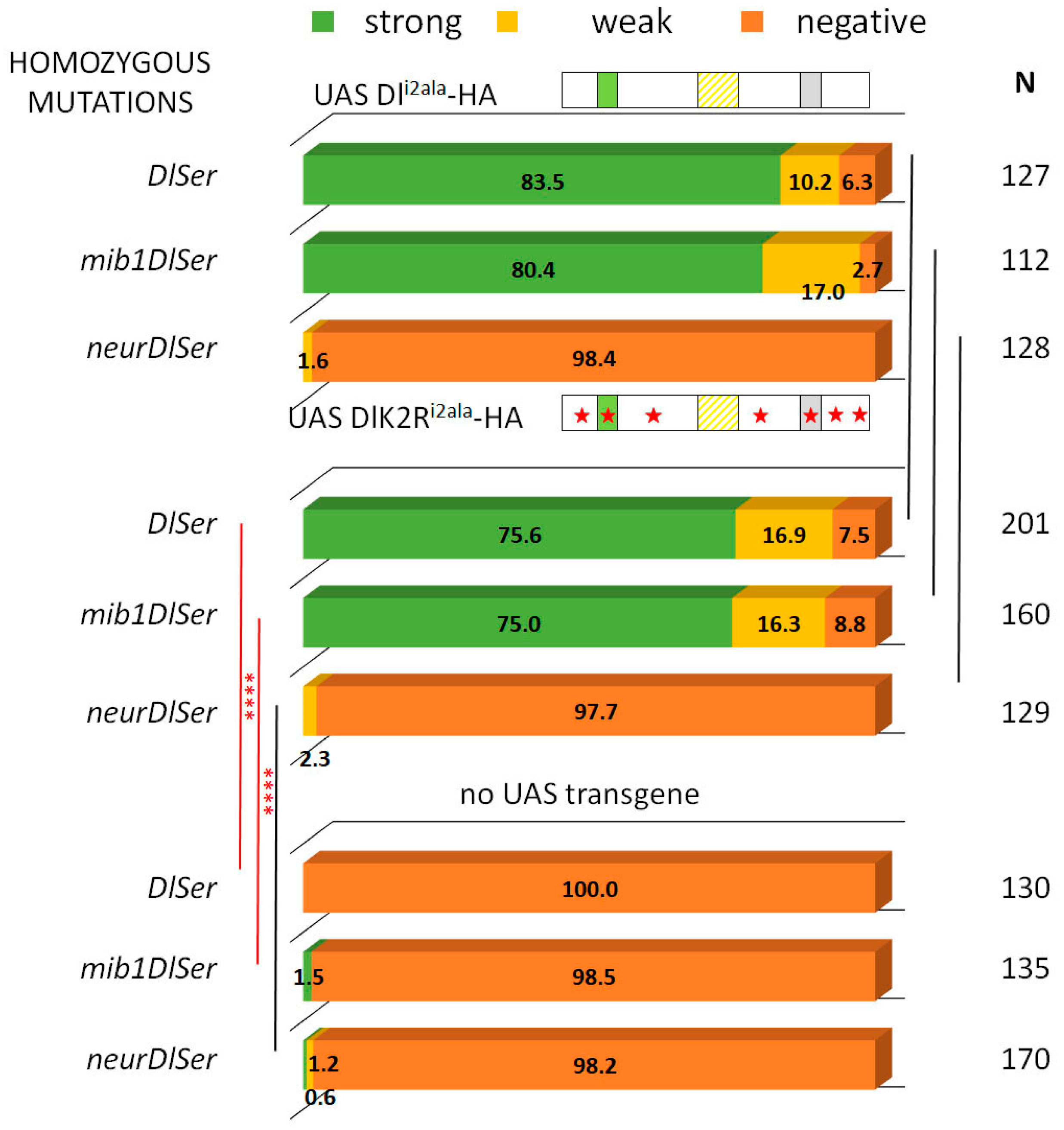
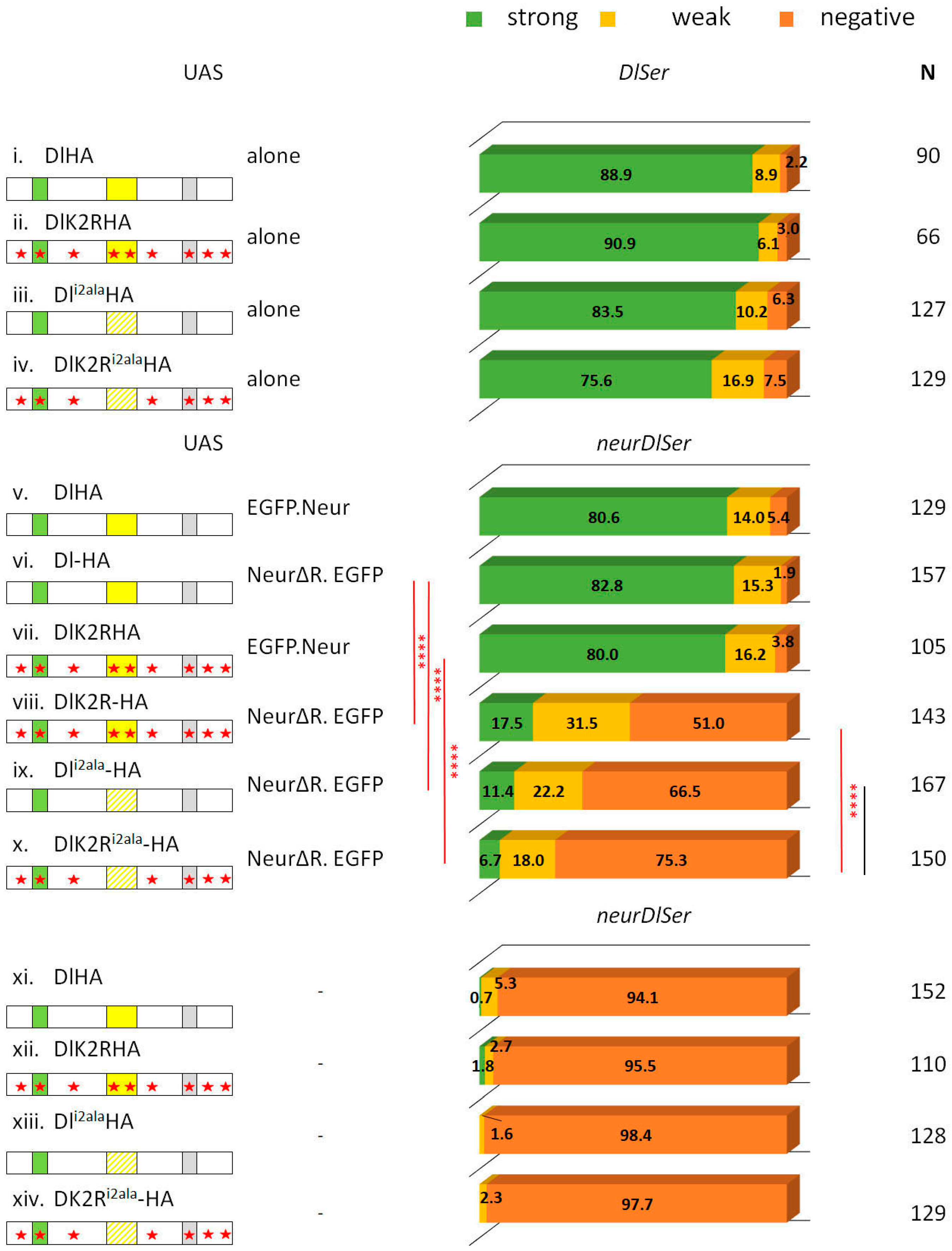
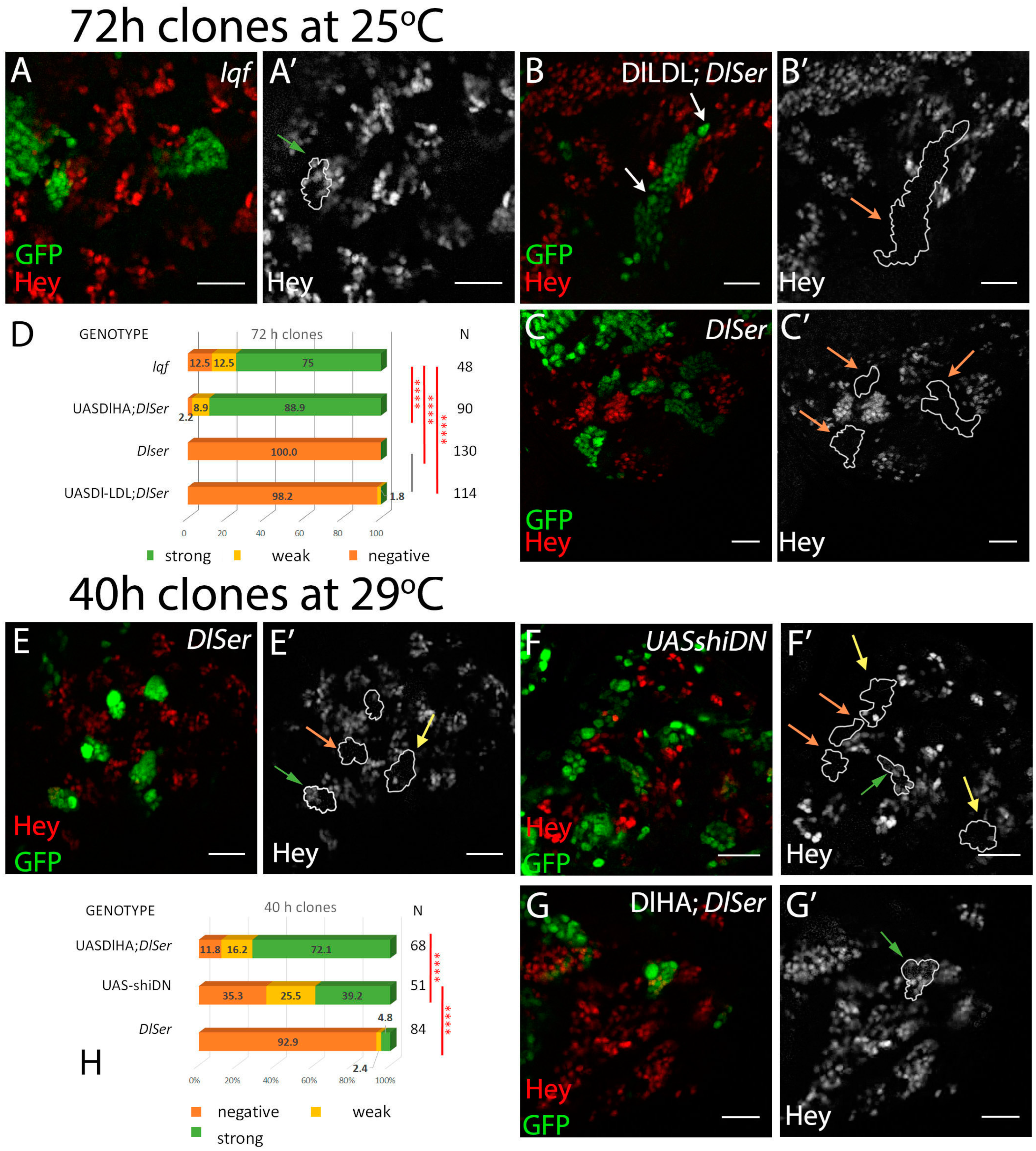
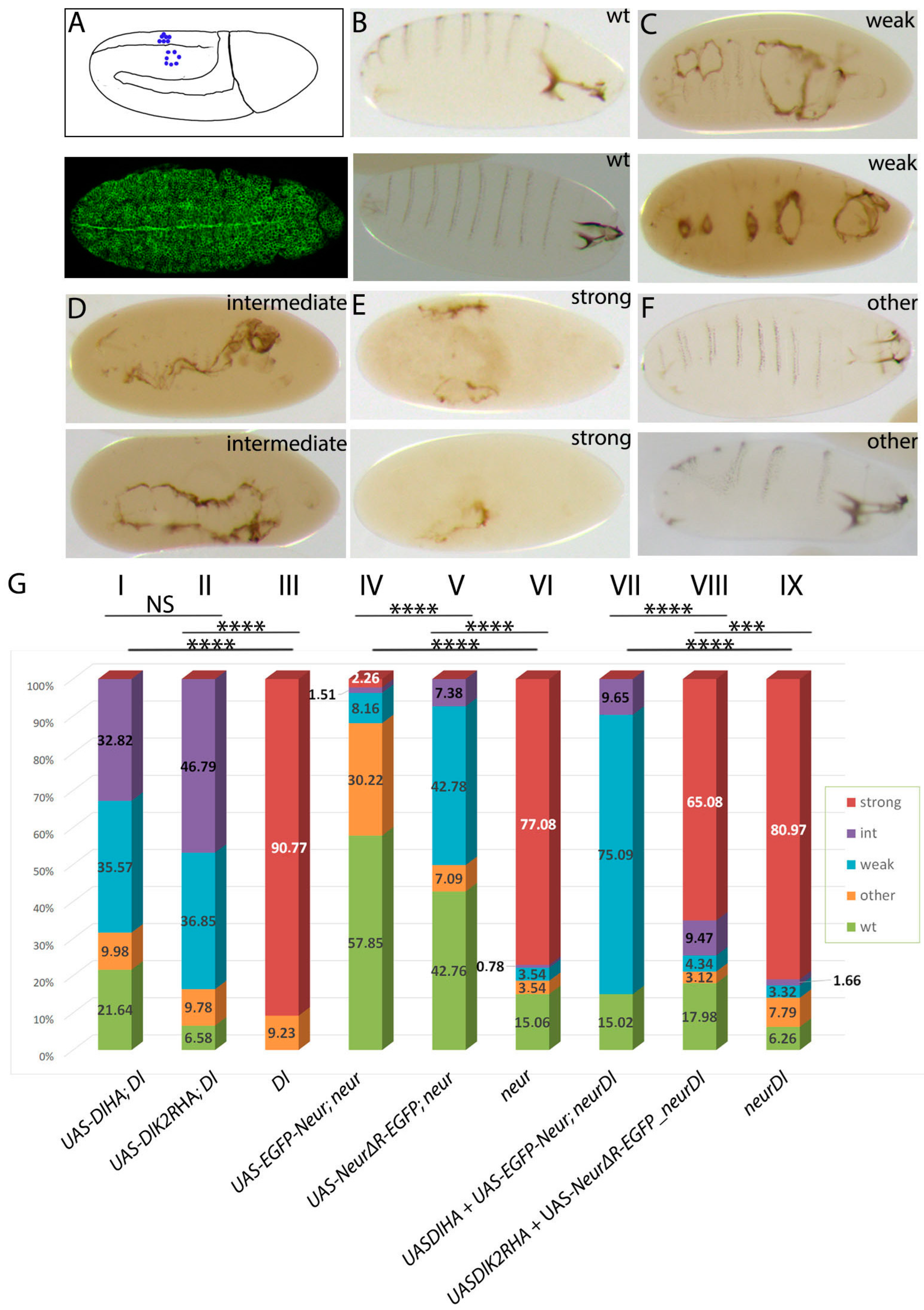
3.1. Delta–Neur Signaling in Newborn Neurons Can Function in the Absence of the Delta Lysines and Mib1
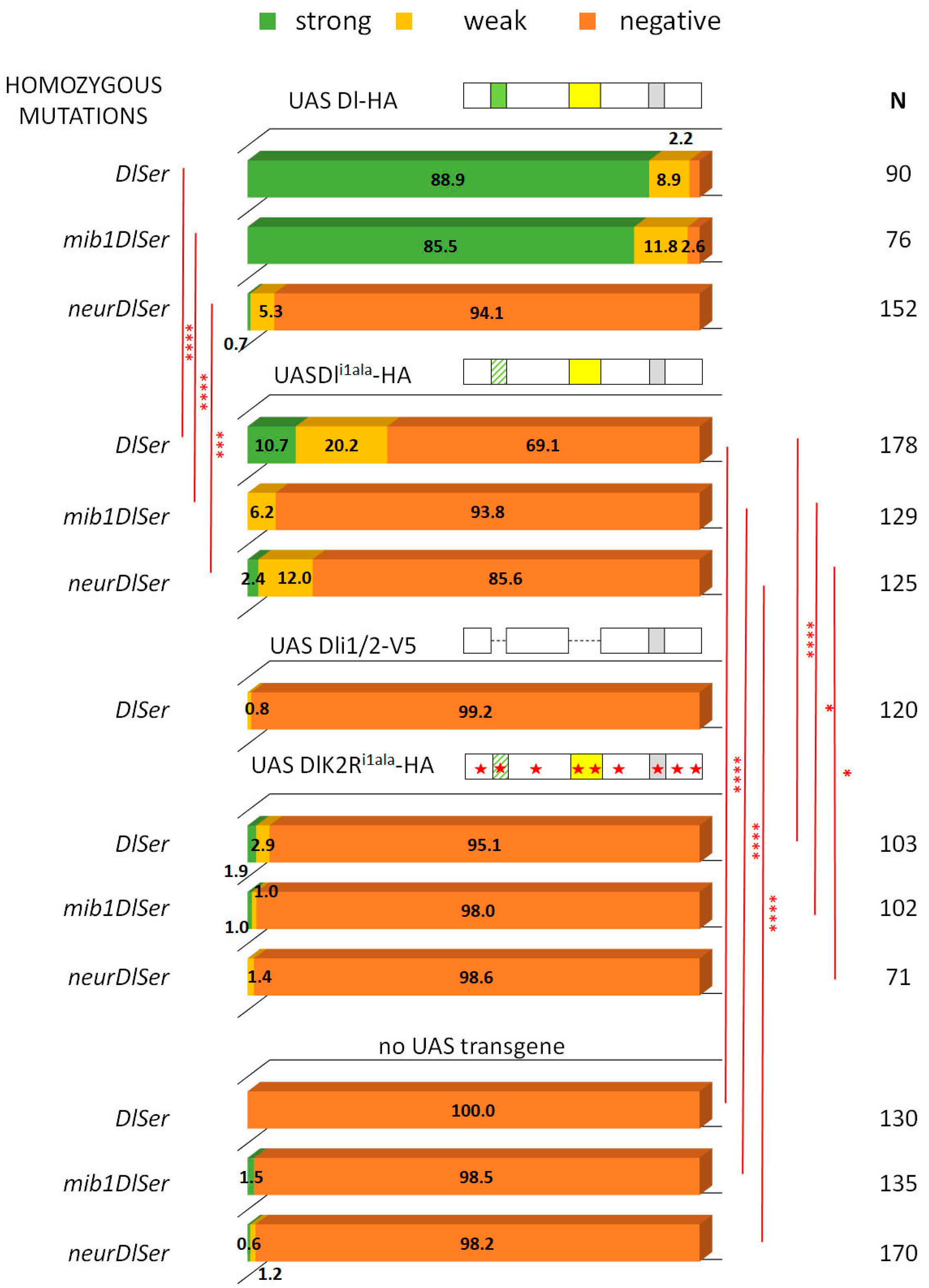
3.2. A Role for Delta Lysines and Mib1 Is Revealed When Delta–Neur Interaction Is Compromised
3.3. The Neur RING Domain Is Important but Not Indispensable for Delta Signaling
3.4. The Role of Endocytosis in Delta Activation in CNS Lineages
3.5. The Dual Role of Neuralized Is Also Evident during Embryonic Neurogenesis
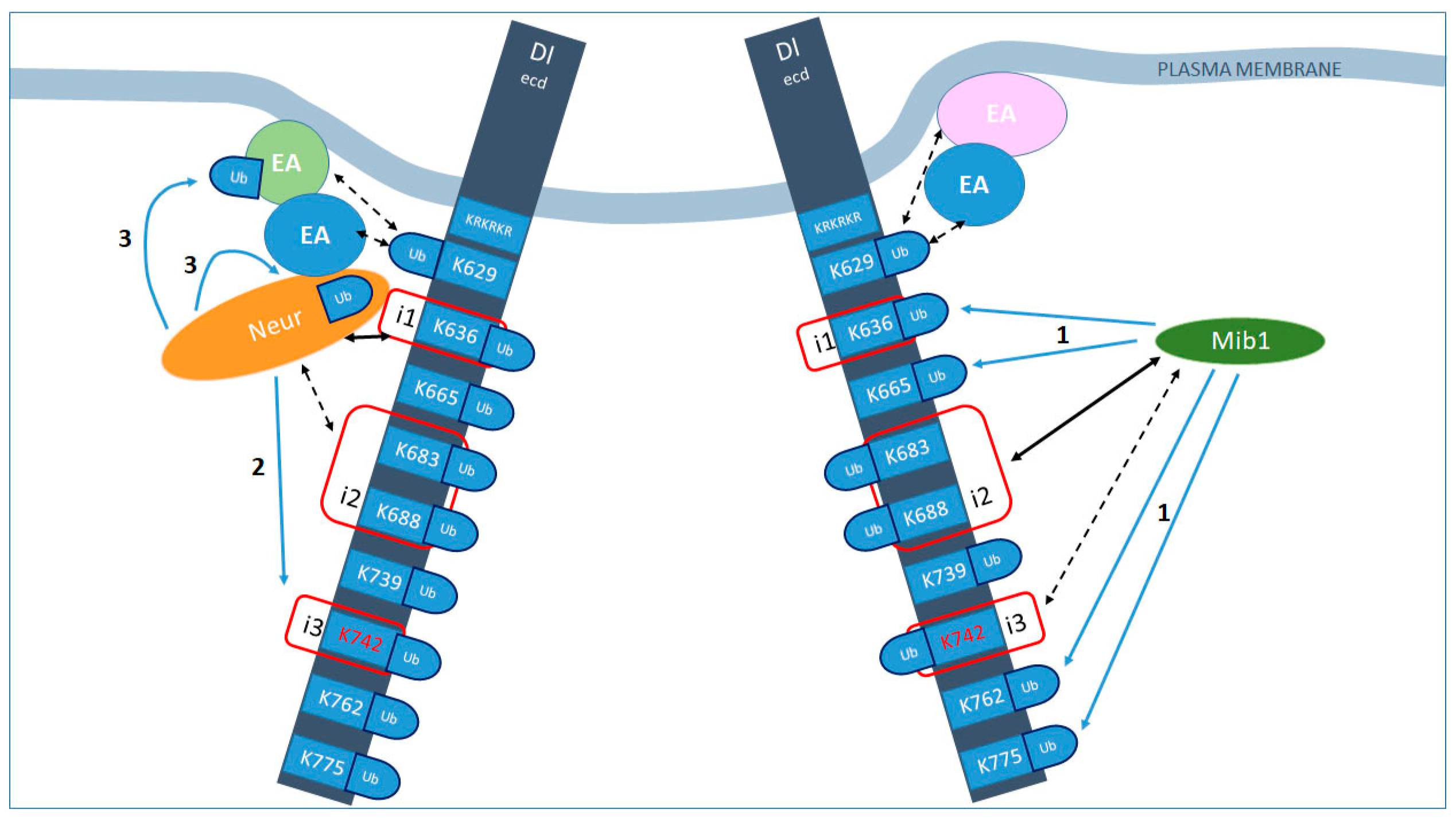
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Artavanis-Tsakonas, S.; Matthew, D.; Rand, R.J.L. Notch Signaling: Cell Fate Control and Signal Integration in Development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef]
- Kovall, R.A.; Gebelein, B.; Sprinzak, D.; Kopan, R. The Canonical Notch Signaling Pathway: Structural and Biochemical Insights into Shape, Sugar, and Force. Dev. Cell 2017, 41, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Bray, S.J. Notch signalling: A simple pathway becomes complex. Nat. Rev. Mol. Cell Biol. 2006, 7, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Gordon, W.R.; Roy, M.; Vardar-Ulu, D.; Garfinkel, M.; Mansour, M.R.; Aster, J.C.; Blacklow, S.C. Structure of the Notch1-negative regulatory region: Implications for normal activation and pathogenic signaling in T-ALL. Blood 2009, 113, 4381–4390. [Google Scholar] [CrossRef]
- Gordon, W.R.; Vardar-Ulu, D.; Histen, G.; Sanchez-Irizarry, C.; Aster, J.C.; Blacklow, S.C. Structural basis for autoinhibition of Notch. Nat. Struct. Mol. Biol. 2007, 14, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Gordon, W.R.; Vardar-Ulu, D.; L’Heureux, S.; Ashworth, T.; Malecki, M.J.; Sanchez-Irizarry, C.; McArthur, D.G.; Histen, G.; Mitchell, J.L.; Aster, J.C.; et al. Effects of S1 cleavage on the structure, surface export, and signaling activity of human Notch1 and Notch2. PLoS ONE 2009, 4, e6613. [Google Scholar] [CrossRef]
- Gordon, W.R.; Zimmerman, B.; He, L.; Miles, L.J.; Huang, J.; Tiyanont, K.; McArthur, D.G.; Aster, J.C.; Perrimon, N.; Loparo, J.J.; et al. Mechanical Allostery: Evidence for a Force Requirement in the Proteolytic Activation of Notch. Dev. Cell 2015, 33, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Pavlopoulos, E.; Pitsouli, C.; Klueg, K.M.; Muskavitch, M.A.T.; Moschonas, N.K.; Delidakis, C. neuralized Encodes a Peripheral Membrane Protein Involved in Delta Signaling and Endocytosis. Dev. Cell 2001, 1, 807–816. [Google Scholar] [CrossRef]
- Daskalaki, A.; Shalaby, N.A.; Kux, K.; Tsoumpekos, G.; Tsibidis, G.D.; Muskavitch, M.A.; Delidakis, C. Distinct intracellular motifs of Delta mediate its ubiquitylation and activation by Mindbomb1 and Neuralized. J. Cell Biol. 2011, 195, 1017–1031. [Google Scholar] [CrossRef]
- Wang, W.; Struhl, G. Distinct roles for Mind bomb, Neuralized and Epsin in mediating DSL endocytosis and signaling in Drosophila. Development 2005, 132, 2883–2894. [Google Scholar] [CrossRef]
- Lai, E.C.; Roegiers, F.; Qin, X.; Jan, Y.N.; Rubin, G.M. The ubiquitin ligase Drosophila mind bomb promotes notch signaling by regulating the localization and activity of Serrate and Delta. Development 2005, 132, 2319–2332. [Google Scholar] [CrossRef]
- Pitsouli, C.; Delidakis, C. The interplay between DSL proteins and ubiquitin ligases in Notch signaling. Development 2005, 132, 4041–4050. [Google Scholar] [CrossRef]
- Le Borgne, R.; Remaud, S.; Hamel, S.; Schweisguth, F. Two distinct E3 ubiquitin ligases have complementary functions in the regulation of delta and serrate signaling in Drosophila. PLoS Biol. 2005, 3, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Piper, R.C.; Dikic, I.; Lukacs, G.L. Ubiquitin-dependent sorting in endocytosis. Cold Spring Harb. Perspect. Biol. 2014, 6, a016808. [Google Scholar] [CrossRef] [PubMed]
- Berlin, I.; Sapmaz, A.; Stévenin, V.; Neefjes, J. Ubiquitin and its relatives as wizards of the endolysosomal system. J. Cell Sci. 2023, 136, jcs260101. [Google Scholar] [CrossRef] [PubMed]
- Meloty-Kapella, L.; Shergill, B.; Kuon, J.; Botvinick, E.; Weinmaster, G. Notch Ligand Endocytosis Generates Mechanical Pulling Force Dependent on Dynamin, Epsins, and Actin. Dev. Cell 2012, 22, 1299–1312. [Google Scholar] [CrossRef] [PubMed]
- McMillan, B.J.; Schnute, B.; Ohlenhard, N.; Zimmerman, B.; Miles, L.; Beglova, N.; Klein, T.; Blacklow, S.C. A Tail of Two Sites: A Bipartite Mechanism for Recognition of Notch Ligands by Mind Bomb E3 Ligases. Mol. Cell 2015, 57, 912–924. [Google Scholar] [CrossRef] [PubMed]
- Deblandre, G.A.; Lai, E.C.; Kintner, C. Xenopus Neuralized Is a Ubiquitin Ligase that Interacts with XDelta1 and Regulates Notch Signaling. Dev. Cell 2001, 1, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Koutelou, E.; Sato, S.; Tomomori-Sato, C.; Florens, L.; Swanson, S.K.; Washburn, M.P.; Kokkinaki, M.; Conaway, R.C.; Conaway, J.W.; Moschonas, N.K. Neuralized-like 1 (Neurl1) targeted to the plasma membrane by N-myristoylation regulates the notch ligand jagged1. J. Biol. Chem. 2008, 283, 3846–3853. [Google Scholar] [CrossRef]
- Berndt, N.; Seib, E.; Kim, S.; Troost, T.; Lyga, M.; Langenbach, J.; Haensch, S.; Kalodimou, K.; Delidakis, C.; Klein, T. Ubiquitylation-independent activation of Notch signalling by Delta. eLife 2017, 6, e27346. [Google Scholar] [CrossRef]
- Khamaisi, B.; Luca, V.C.; Blacklow, S.C.; Sprinzak, D. Functional Comparison between Endogenous and Synthetic Notch Systems. ACS Synth. Biol. 2022, 11, 3343–3353. [Google Scholar] [CrossRef] [PubMed]
- Langridge, P.D.; Struhl, G. Epsin-Dependent Ligand Endocytosis Activates Notch by Force. Cell 2017, 171, 1383–1396.e12. [Google Scholar] [CrossRef] [PubMed]
- Morsut, L.; Roybal, K.T.; Xiong, X.; Gordley, R.M.; Coyle, S.M.; Thomson, M.; Lim, W.A. Engineering Customized Cell Sensing and Response Behaviors Using Synthetic Notch Receptors. Cell 2016, 164, 780–791. [Google Scholar] [CrossRef] [PubMed]
- Seib, E.; Klein, T. The role of ligand endocytosis in notch signalling. Biol. Cell 2021, 113, 401–418. [Google Scholar] [CrossRef] [PubMed]
- Windler, S.L.; Bilder, D. Endocytic Internalization Routes Required for Delta/Notch Signaling. Curr. Biol. 2010, 20, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Lai, E.C.; Rubin, G.M. Neuralized functions cell-autonomously to regulate a subset of notch-dependent processes during adult Drosophila development. Dev. Biol. 2001, 231, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, R.; Jiménez, F.; Dietrich, U.; Campos-Ortega, J.A. On the phenotype and development of mutants of early neurogenesis in Drosophila melanogaster. Wilhelm Roux’s Arch. Dev. Biol. 1983, 192, 62–74. [Google Scholar] [CrossRef]
- Monastirioti, M.; Giagtzoglou, N.; Koumbanakis, K.A.; Zacharioudaki, E.; Deligiannaki, M.; Wech, I.; Almeida, M.; Preiss, A.; Bray, S.; Delidakis, C. Drosophila Hey is a target of Notch in asymmetric divisions during embryonic and larval neurogenesis. Development 2010, 137, 191–201. [Google Scholar] [CrossRef]
- Bischof, J.; Maeda, R.K.; Hediger, M.; Karch, F.; Basler, K. An optimized transgenesis system for Drosophila using germ-line-specific φC31 integrases. Proc. Natl. Acad. Sci. USA 2007, 104, 3312–3317. [Google Scholar] [CrossRef]
- Wang, W.; Struhl, G. Drosophila Epsin mediates a select endocytic pathway that DSL ligands must enter to activate Notch. Development 2004, 131, 5367–5380. [Google Scholar] [CrossRef]
- Wodarz, A.; Hinz, U.; Engelbert, M.; Knust, E. Expression of crumbs confers apical character on plasma membrane domains of ectodermal epithelia of Drosophila. Cell 1995, 82, 67–76. [Google Scholar] [CrossRef]
- Nagarkar-Jaiswal, S.; Lee, P.T.; Campbell, M.E.; Chen, K.; Anguiano-Zarate, S.; Gutierrez, M.C.; Busby, T.; Lin, W.-W.; He, Y.; Schulze, K.L.; et al. A library of MiMICs allows tagging of genes and reversible, spatial and temporal knockdown of proteins in Drosophila. eLife 2015, 2015, e05338. [Google Scholar] [CrossRef]
- Magadi, S.S.; Voutyraki, C.; Anagnostopoulos, G.; Zacharioudaki, E.; Poutakidou, I.K.; Efraimoglou, C.; Stapountzi, M.; Theodorou, V.; Nikolaou, C.; Koumbanakis, K.A.; et al. Dissecting Hes-centred transcriptional networks in neural stem cell maintenance and tumorigenesis in Drosophila. Development 2020, 147, dev191544. [Google Scholar] [CrossRef] [PubMed]
- Wieschaus, E.; Nόsslein-Volhard, C. Looking at Embryos in Drosophila, a Practical Approach; Roberts, D.B., Ed.; IRL Press: Oxford, UK, 1986. [Google Scholar]
- Fontana, J.R.; Posakony, J.W. Both inhibition and activation of Notch signaling rely on a conserved Neuralized-binding motif in Bearded proteins and the Notch ligand Delta. Dev. Biol. 2009, 333, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Bardin, A.J.; Schweisguth, F. Bearded family members inhibit neuralized-mediated endocytosis and signaling activity of Delta in Drosophila. Dev. Cell 2006, 10, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Overstreet, E.; Fitch, E.; Fischer, J.A. Fat facets and Liquid facets promote Delta endocytosis and Delta signaling in the signaling cells. Development 2004, 131, 5355–5366. [Google Scholar] [CrossRef] [PubMed]
- Speicher, S.A.; Thomas, U.; Hinz, U.; Knust, E. The Serrate locus of Drosophila and its role in morphogenesis of the wing imaginal discs: Control of cell proliferation. Development 1994, 120, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Delidakis, C.; Preiss, A.; Hartley, D.A.; Artavanis-Tsakonas, S. Two genetically and molecularly distinct functions involved in early neurogenesis reside within the Enhancer of split locus of Drosophila melanogaster. Genetics 1991, 129, 803–823. [Google Scholar] [CrossRef] [PubMed]
- Sprinzak, D.; Blacklow, S.C. Biophysics of Notch Signaling. Annu. Rev. Biophys. 2021, 50, 157–189. [Google Scholar] [CrossRef]
- Troost, T.; Seib, E.; Airich, A.; Vüllings, N.; Necakov, A.; De Renzis, S.; Klein, T. The meaning of ubiquitylation of the DSL ligand Delta for the development of Drosophila. BMC Biol. 2023, 21, 260. [Google Scholar] [CrossRef]
- Liu, S.; Boulianne, G.L. The NHR domains of Neuralized and related proteins: Beyond Notch signalling. Cell. Signal. 2017, 29, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Gollapudi, S.; Day, K.; Ashby, G.; Sangani, A.; Malady, B.; Wang, L.; Lafer, E.M.; Huibregtse, J.; Stachowiak, J. Ubiquitin-driven protein condensation promotes clathrin-mediated endocytosis. bioRxiv 2023. [Google Scholar] [CrossRef]
- Clarke, D.N.; Martin, A.C. Actin-based force generation and cell adhesion in tissue morphogenesis. Curr. Biol. 2021, 31, R667–R680. [Google Scholar] [CrossRef] [PubMed]
- Varnum-Finney, B.; Wu, L.; Yu, M.; Brashem-Stein, C.; Staats, S.; Flowers, D.; Griffin, J.D.; Bernstein, I.D. Immobilization of Notch ligand, Delta-1, is required for induction of Notch signaling. J. Cell Sci. 2000, 113, 4313–4318. [Google Scholar] [CrossRef] [PubMed]
- Perez-Mockus, G.; Roca, V.; Mazouni, K.; Schweisguth, F. Neuralized regulates Crumbs endocytosis and epithelium morphogenesis via specific Stardust isoforms. J. Cell Biol. 2017, 216, 1405–1420. [Google Scholar] [CrossRef] [PubMed]
- Shard, C.; Luna-Escalante, J.; Schweisguth, F. Tissue-wide coordination of epithelium-to-neural stem cell transition in the Drosophila optic lobe requires Neuralized. J. Cell Biol. 2020, 219, e202005035. [Google Scholar] [CrossRef]
- Contreras, E.G.; Egger, B.; Gold, K.S.; Brand, A.H. Dynamic Notch signalling regulates neural stem cell state progression in the Drosophila optic lobe. Neural Dev. 2018, 13, 25. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutation | Primer Name | Primer Sequence |
|---|---|---|
| NEQNAV to AAAAAA (Dli1ala) | Dl_sdm_NxxN2A_fwd | G GCC AGG AAG CAG GCC GCA GCG GCT GCG GCG GCC ACA ATG CAT C |
| Dl_sdm_NxxN2A_rev | G ATG CAT TGT GGC CGC CGC AGC CGC TGC GGC CTG CTT CCT GGC C | |
| IRNTWDR to AAAAAAA (DlK2Ri2ala) | DlK2R_sdm_NBox2A_fwd | GGC GGC AAC CCG AAT ATC GCC GCT GCC GCC GCG GCC GCA TCG GTC AAC AAC ATT TGT GCC |
| DlK2R_sdm_NBox2A_rev | GGC ACA AAT GTT GTT GAC CGA TGC GGC CGC GGC GGC AGC GGC GAT ATT CGG GTT GCC GCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalodimou, K.; Stapountzi, M.; Vüllings, N.; Seib, E.; Klein, T.; Delidakis, C. Separable Roles for Neur and Ubiquitin in Delta Signalling in the Drosophila CNS Lineages. Cells 2023, 12, 2833. https://doi.org/10.3390/cells12242833
Kalodimou K, Stapountzi M, Vüllings N, Seib E, Klein T, Delidakis C. Separable Roles for Neur and Ubiquitin in Delta Signalling in the Drosophila CNS Lineages. Cells. 2023; 12(24):2833. https://doi.org/10.3390/cells12242833
Chicago/Turabian StyleKalodimou, Konstantina, Margarita Stapountzi, Nicole Vüllings, Ekaterina Seib, Thomas Klein, and Christos Delidakis. 2023. "Separable Roles for Neur and Ubiquitin in Delta Signalling in the Drosophila CNS Lineages" Cells 12, no. 24: 2833. https://doi.org/10.3390/cells12242833
APA StyleKalodimou, K., Stapountzi, M., Vüllings, N., Seib, E., Klein, T., & Delidakis, C. (2023). Separable Roles for Neur and Ubiquitin in Delta Signalling in the Drosophila CNS Lineages. Cells, 12(24), 2833. https://doi.org/10.3390/cells12242833