
Profiling of G-Protein Coupled Receptors in Adipose Tissue and Differentiating Adipocytes Offers a Translational Resource for Obesity/Metabolic Research

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Culture and Adipogenic Differentiation of 3T3-L1 Adipocytes
2.3. Isolation of Mouse Adipose Tissue Depots
2.4. RNA Isolation and RT-PCR by Taqman Arrays
2.5. Western Blotting
2.6. RNA Sequencing Re-Analysis
2.7. Statistical Analyses
3. Results
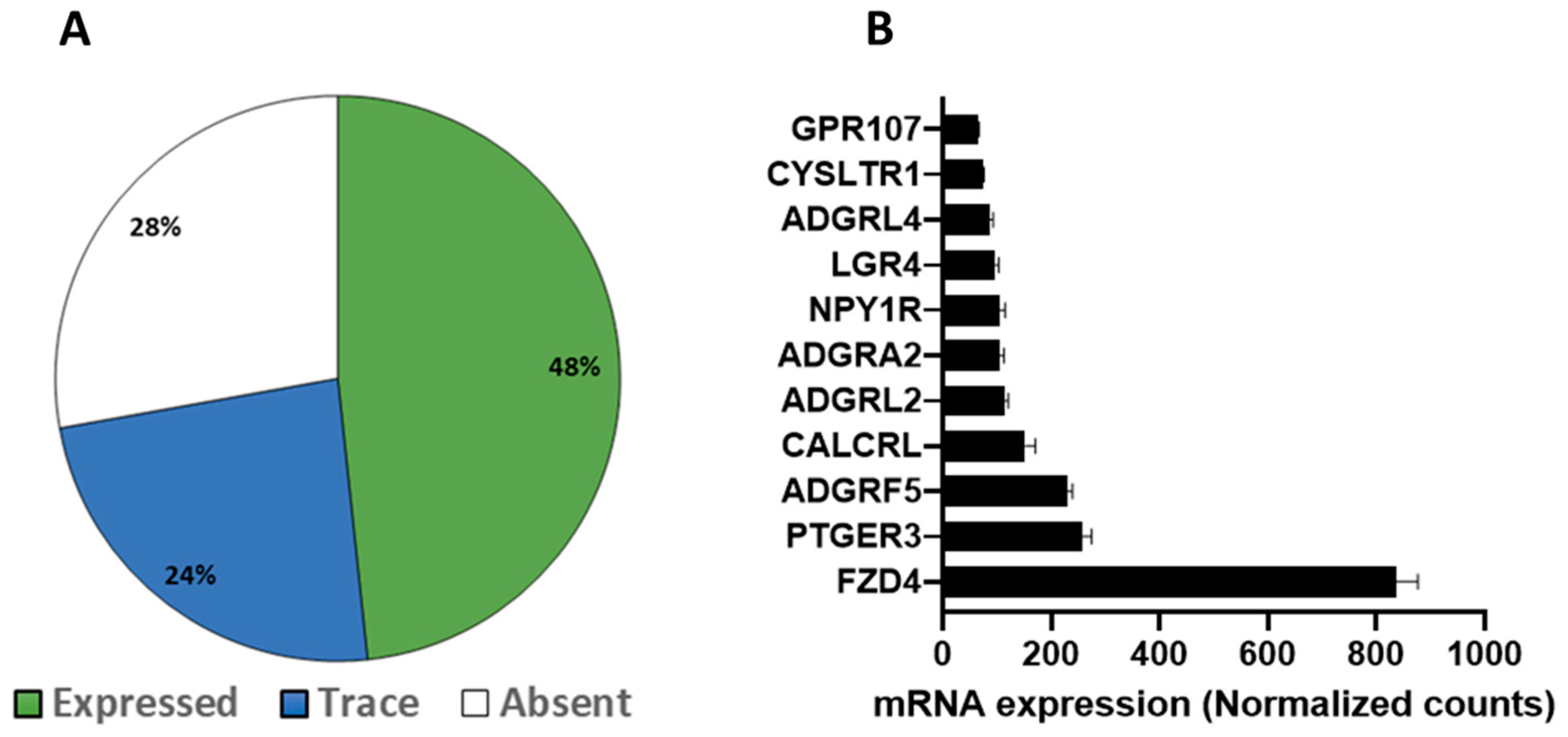
3.1. GPCR Expression Profile in Mouse White and Brown Adipose Tissue
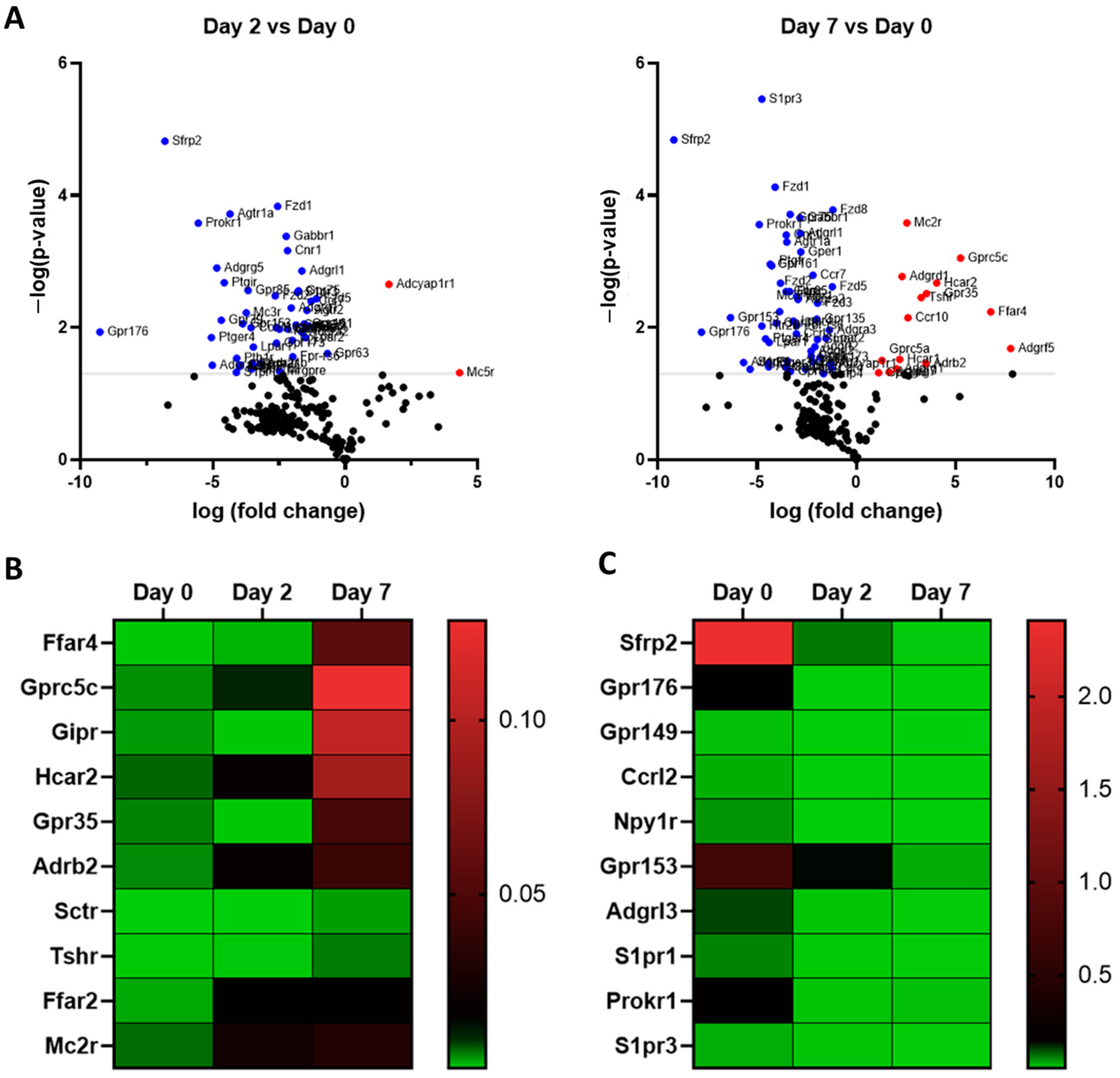
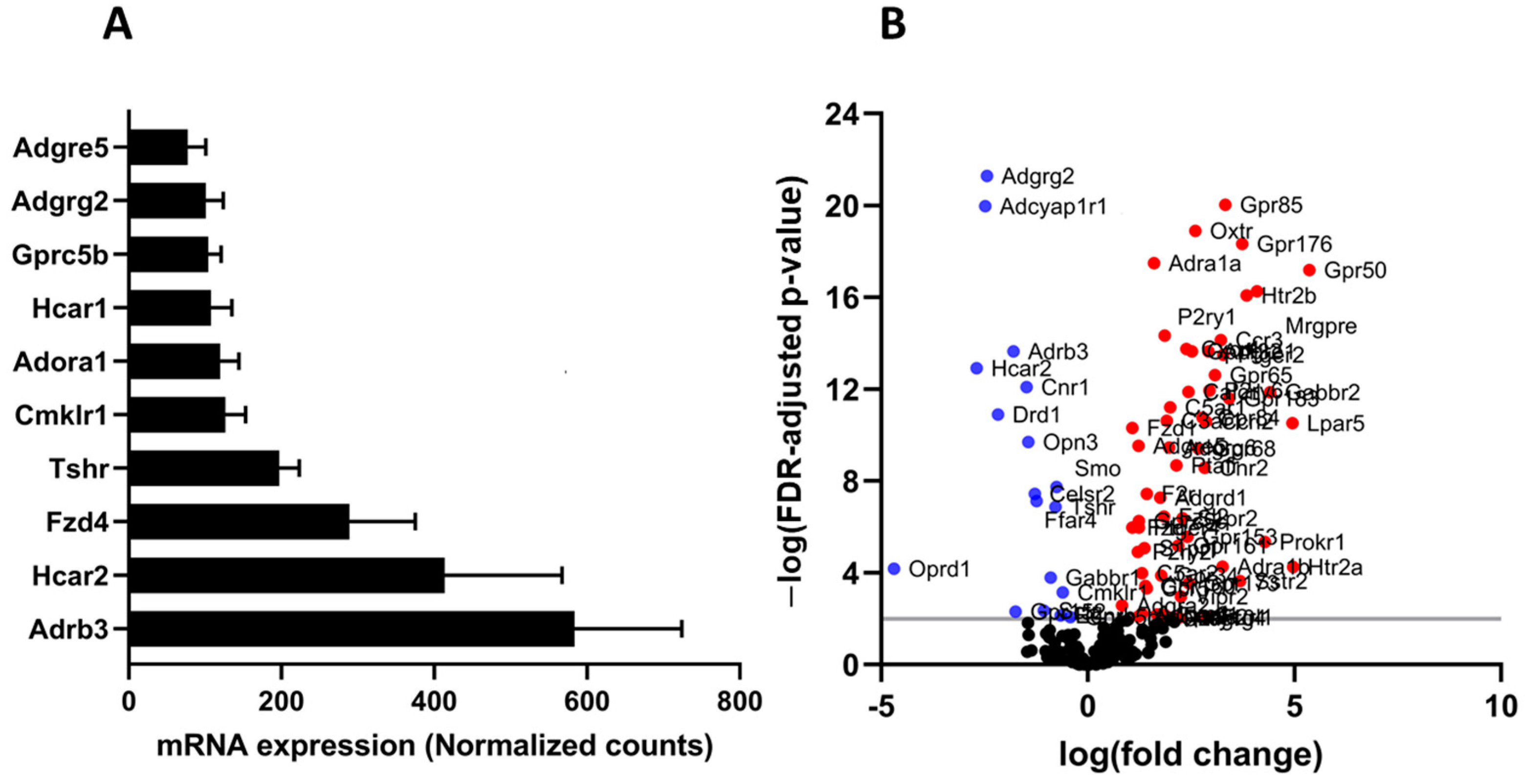
3.2. Comparative Analysis of GPCR mRNA Expression in Undifferentiated and Differentiated 3T3-L1 Adipocytes
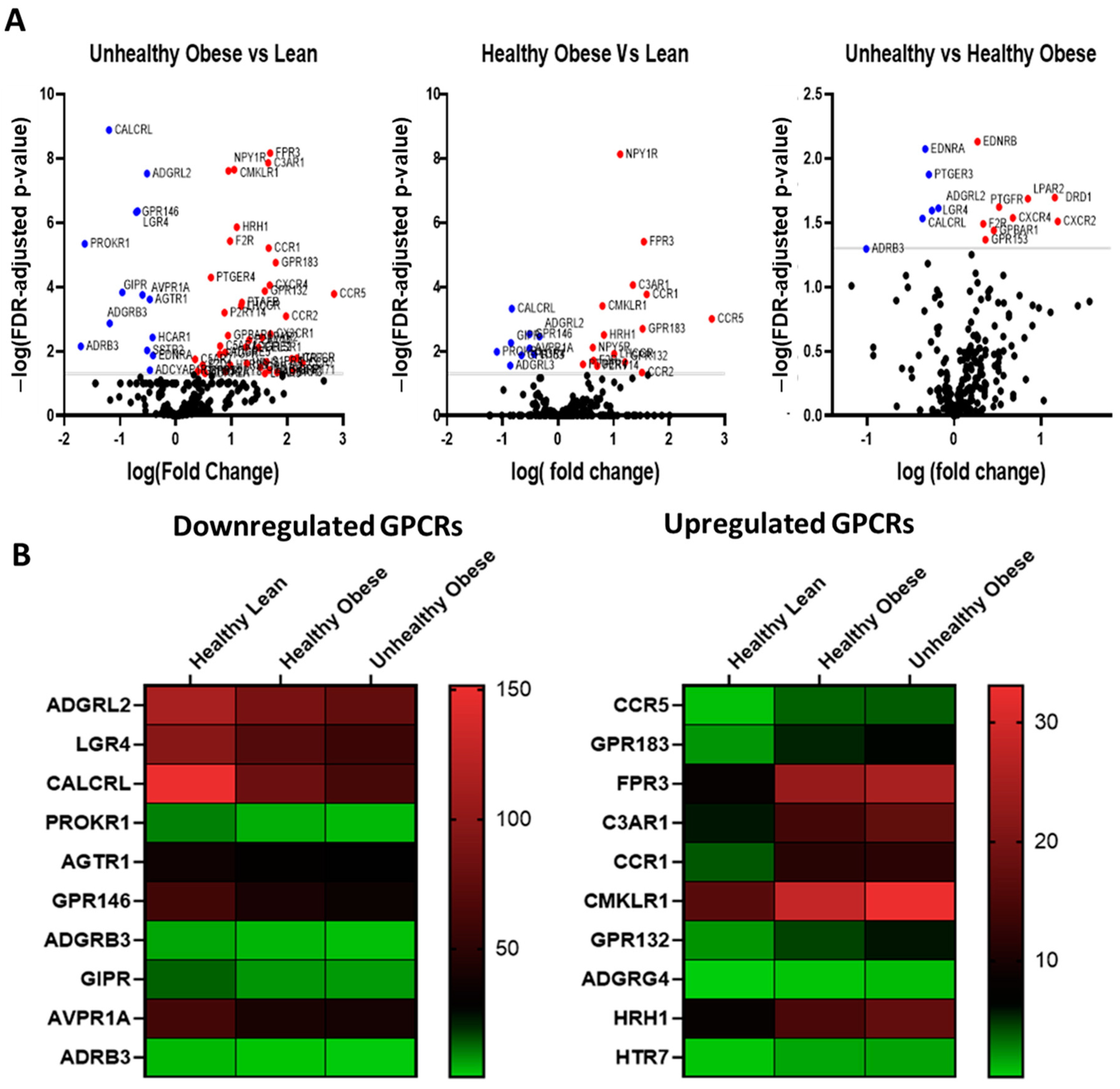
3.3. GPCR Expression in Adipocytes Isolated from Normal and High-Fat Fed Mice
3.4. GPCRs of Interest
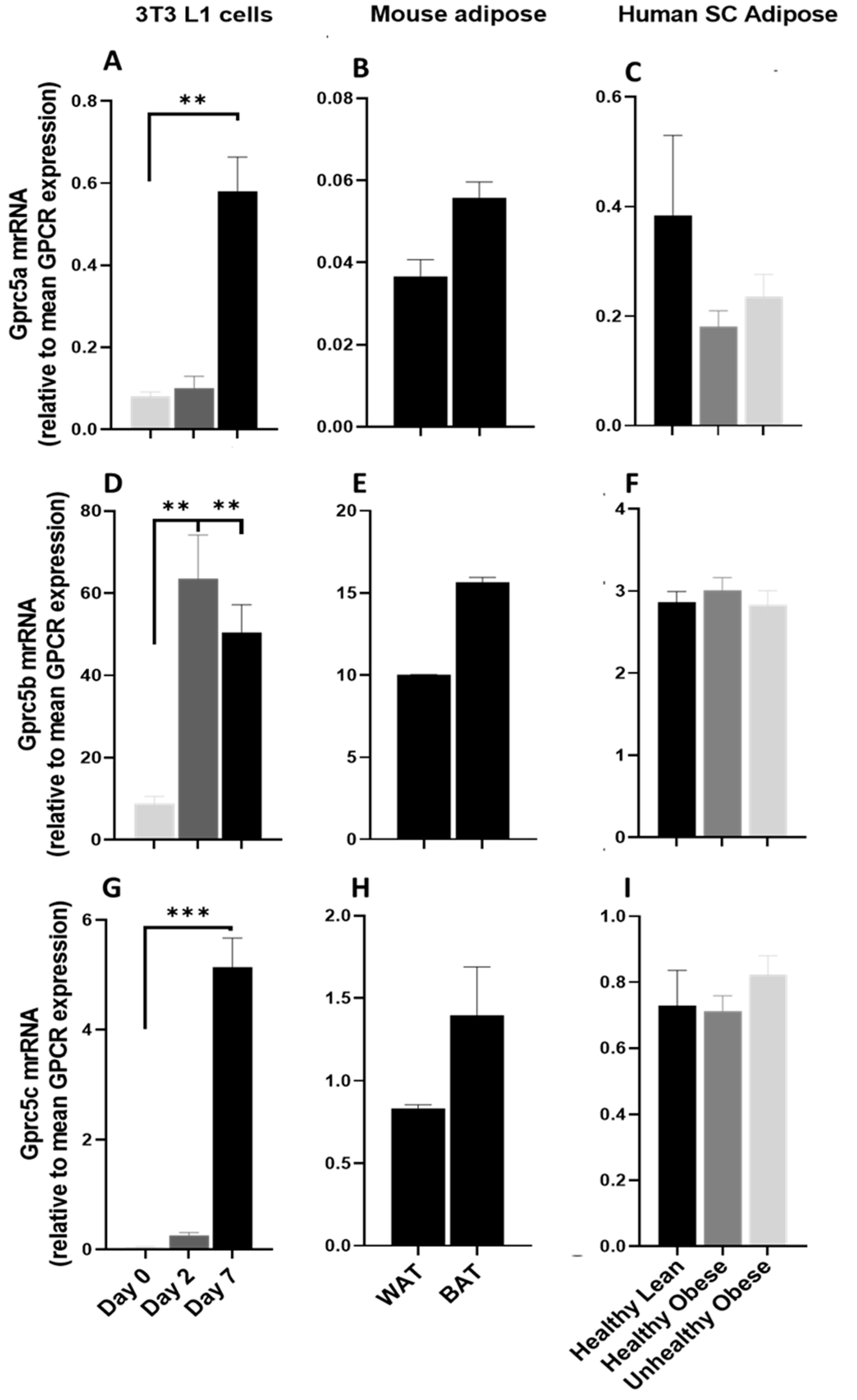
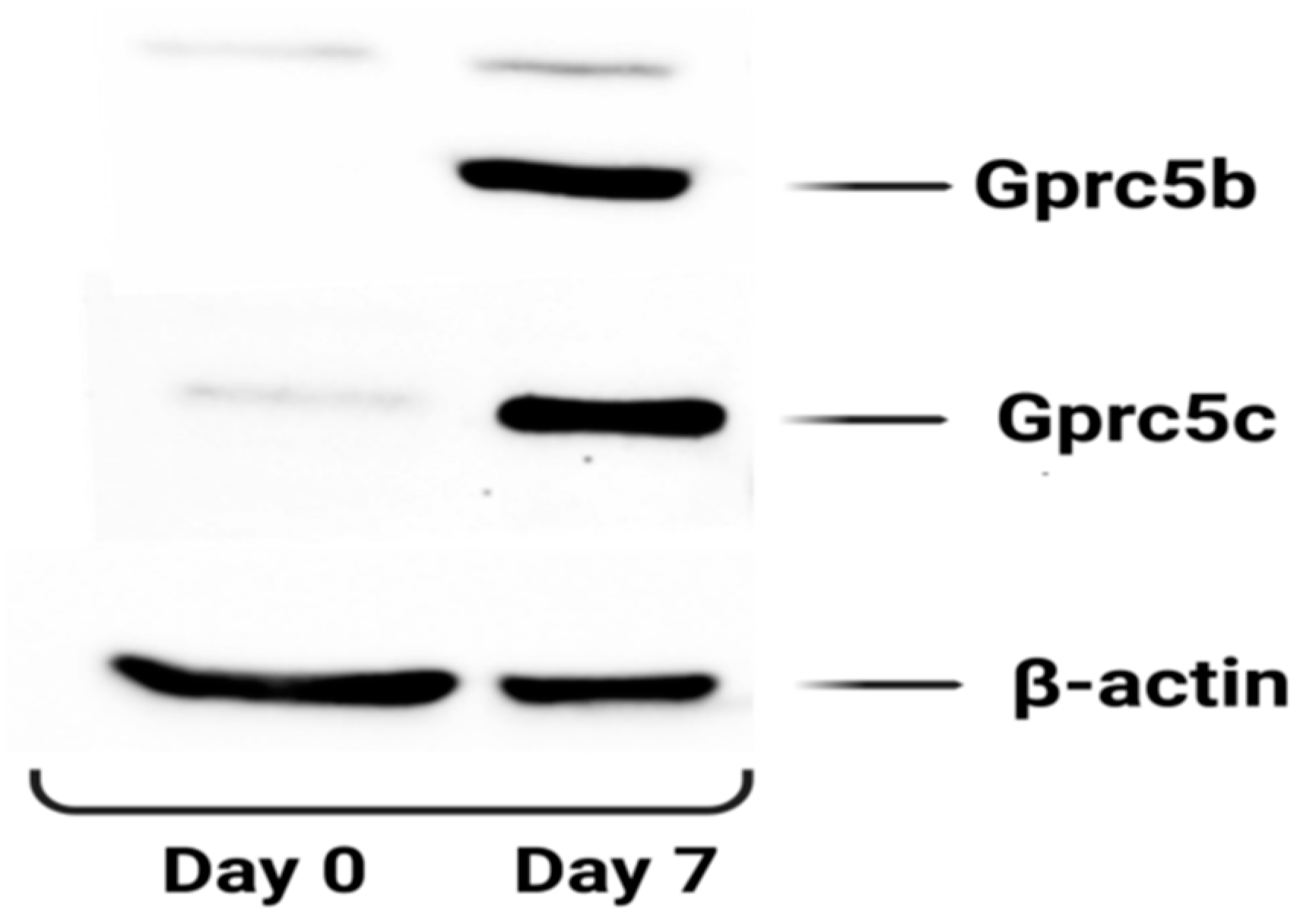
3.4.1. GPRC5 Family of Receptors
3.4.2. G Protein-Coupled Receptor 146 (GPR146)
3.4.3. G Protein-Coupled Receptor 137 (GPR137)
3.4.4. Arginine Vasopressin Receptor 1A (AVPR1A)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierce, K.L.; Premont, R.; Lefkowitz, R.J. Seven-transmembrane receptors. Nat. Rev. Mol. Cell Biol. 2002, 3, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Strange, P.G. Signaling Mechanisms of GPCR Ligands. Curr. Opin. Drug Discov. Dev. 2008, 11, 196–202. [Google Scholar]
- Pavlos, N.J.; Friedman, P.A. GPCR Signaling and Trafficking: The Long and Short of It. Trends Endocrinol. Metab. 2017, 28, 213–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhu, L.; Meister, J.; Bone, D.B.; Pydi, S.P.; Rossi, M.; Wess, J. Use of DREADD Technology to Identify Novel Targets for Antidiabetic Drugs. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 421–440. [Google Scholar] [CrossRef]
- Sriram, K.; Insel, P.A. G Protein-Coupled Receptors as Targets for Approved Drugs: How Many Targets and How Many Drugs? Mol. Pharmacol. 2018, 93, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef]
- Al Mahri, S.; Malik, S.S.; Al Ibrahim, M.; Haji, E.; Dairi, G.; Mohammad, S. Free Fatty Acid Receptors (FFARs) in Adipose: Physiological Role and Therapeutic Outlook. Cells 2022, 11, 750. [Google Scholar] [CrossRef]
- Amisten, S.; Neville, M.; Hawkes, R.; Persaud, S.J.; Karpe, F.; Salehi, A. An atlas of G-protein coupled receptor expression and function in human subcutaneous adipose tissue. Pharmacol. Ther. 2015, 146, 61–93. [Google Scholar] [CrossRef]
- Im, H.; Park, J.-H.; Im, S.; Han, J.; Kim, K.; Lee, Y.-H. Regulatory roles of G-protein coupled receptors in adipose tissue metabolism and their therapeutic potential. Arch. Pharmacal Res. 2021, 44, 133–145. [Google Scholar] [CrossRef]
- Suchý, T.; Zieschang, C.; Popkova, Y.; Kaczmarek, I.; Weiner, J.; Liebing, A.-D.; Çakir, M.V.; Landgraf, K.; Gericke, M.; Pospisilik, J.A.; et al. The repertoire of Adhesion G protein-coupled receptors in adipocytes and their functional relevance. Int. J. Obes. 2020, 44, 2124–2136. [Google Scholar] [CrossRef] [Green Version]
- Collins, S.; Surwit, R.S. The beta-adrenergic receptors and the control of adipose tissue metabolism and thermogenesis. Recent Prog. Horm. Res. 2001, 56, 309–328. [Google Scholar] [CrossRef]
- London, E.; Stratakis, C.A. The regulation of PKA signaling in obesity and in the maintenance of metabolic health. Pharmacol. Ther. 2022, 237, 108113. [Google Scholar] [CrossRef]
- Collins, S. β-adrenoceptor signaling networks in adipocytes for recruiting stored fat and energy expenditure. Front. Endocrinol. 2011, 2, 102. [Google Scholar] [CrossRef] [Green Version]
- Langin, D.; Portillo, M.P.; Saulnier-Blache, J.-S.; Lafontan, M. Coexistence of three β-adrenoceptor subtypes in white fat cells of various mammalian species. Eur. J. Pharmacol. 1991, 199, 291–301. [Google Scholar] [CrossRef]
- Atgié, C.; D’Allaire, F.; Bukowiecki, L.J. Role of β1- and β3-adrenoceptors in the regulation of lipolysis and thermogenesis in rat brown adipocytes. Am. J. Physiol. 1997, 273 (4 Pt 1), C1136–C1142. [Google Scholar] [CrossRef]
- Evans, B.A.; Merlin, J.; Bengtsson, T.; Hutchinson, D.S. Adrenoceptors in white, brown, and brite adipocytes. Br. J. Pharmacol. 2019, 176, 2416–2432. [Google Scholar] [CrossRef]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef] [Green Version]
- Gozal, D.; Qiao, Z.; Almendros, I.; Zheng, J.; Khalyfa, A.; Shimpukade, B.; Ulven, T. Treatment with TUG891, a free fatty acid receptor 4 agonist, restores adipose tissue metabolic dysfunction following chronic sleep fragmentation in mice. Int. J. Obes. 2016, 40, 1143–1149. [Google Scholar] [CrossRef]
- Meriño, M.; Briones, L.; Palma, V.; Herlitz, K.; Escudero, C. Rol de los receptores de adenosina en la interacción adipocito-macrófago durante la obesidad. Endocrinología, Diabetes y Nutrición 2017, 64, 317–327. [Google Scholar] [CrossRef]
- Olaniru, O.E.; Persaud, S.J. Adhesion G-protein coupled receptors: Implications for metabolic function. Pharmacol. Ther. 2019, 198, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Georgiadi, A.; Lopez-Salazar, V.; Merahbi, R.E.; Karikari, R.A.; Ma, X.; Mourão, A.; Klepac, K.; Bühler, L.; Alfaro, A.J.; Kaczmarek, I.; et al. Orphan GPR116 mediates the insulin sensitizing effects of the hepatokine FNDC4 in adipose tissue. Nat. Commun. 2021, 12, 2999. [Google Scholar] [CrossRef] [PubMed]
- Wanders, D.; Graff, E.C.; Judd, R.L. Effects of high fat diet on GPR109A and GPR81 gene expression. Biochem. Biophys. Res. Commun. 2012, 425, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.-Q.; Ren, N.; Jin, L.; Cheng, K.; Kash, S.; Chen, R.; Wright, S.D.; Taggart, A.K.; Waters, M.G. Role of GPR81 in lactate-mediated reduction of adipose lipolysis. Biochem. Biophys. Res. Commun. 2008, 377, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Cifarelli, V.; Beeman, S.C.; Smith, G.I.; Yoshino, J.; Morozov, D.; Beals, J.W.; Kayser, B.D.; Watrous, J.D.; Jain, M.; Patterson, B.W.; et al. Decreased adipose tissue oxygenation associates with insulin resistance in individuals with obesity. J. Clin. Investig. 2020, 130, 6688–6699. [Google Scholar] [CrossRef]
- Jones, J.E.C.; Rabhi, N.; Orofino, J.; Gamini, R.; Perissi, V.; Vernochet, C.; Farmer, S.R. The Adipocyte Acquires a Fibroblast-Like Transcriptional Signature in Response to a High Fat Diet. Sci. Rep. 2020, 10, 2380. [Google Scholar] [CrossRef] [Green Version]
- Atanes, P.; Ashik, T.; Persaud, S.J. Obesity-induced changes in human islet G protein-coupled receptor expression: Implications for metabolic regulation. Pharmacol. Ther. 2021, 228, 107928. [Google Scholar] [CrossRef]
- Mahi, N.A.; Najafabadi, M.F.; Pilarczyk, M.; Kouril, M.; Medvedovic, M. GREIN: An Interactive Web Platform for Re-analyzing GEO RNA-seq Data. Sci. Rep. 2019, 9, 7580. [Google Scholar] [CrossRef] [Green Version]
- Sriram, K.; Wiley, S.Z.; Moyung, K.; Gorr, M.W.; Salmerón, C.; Marucut, J.; French, R.P.; Lowy, A.M.; Insel, P.A. Detection and Quantification of GPCR mRNA: An Assessment and Implications of Data from High-Content Methods. ACS Omega 2019, 4, 17048–17059. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, S.; Patel, R.T.; Bruno, J.; Panhwar, M.S.; Wen, J.; McGraw, T.E. A Naturally Occurring GIP Receptor Variant Undergoes Enhanced Agonist-Induced Desensitization, Which Impairs GIP Control of Adipose Insulin Sensitivity. Mol. Cell. Biol. 2014, 34, 3618–3629. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, S.; Ramos, L.S.; Buck, J.; Levin, L.R.; Rubino, F.; McGraw, T.E. Gastric Inhibitory Peptide Controls Adipose Insulin Sensitivity via Activation of cAMP-response Element-binding Protein and p110β Isoform of Phosphatidylinositol 3-Kinase. J. Biol. Chem. 2011, 286, 43062–43070. [Google Scholar] [CrossRef] [Green Version]
- Ceperuelo-Mallafré, V.; Duran, X.; Pachón, G.; Roche, K.; Garrido-Sánchez, L.; Vilarrasa, N.; Tinahones, F.J.; Vicente, V.; Pujol, J.; Vendrell, J.; et al. Disruption of GIP/GIPR Axis in Human Adipose Tissue Is Linked to Obesity and Insulin Resistance. J. Clin. Endocrinol. Metab. 2014, 99, E908–E919. [Google Scholar] [CrossRef]
- Agudelo, L.Z.; Ferreira, D.M.S.; Cervenka, I.; Bryzgalova, G.; Dadvar, S.; Jannig, P.R.; Pettersson-Klein, A.T.; Lakshmikanth, T.; Sustarsic, E.G.; Porsmyr-Palmertz, M.; et al. Kynurenic Acid and Gpr35 Regulate Adipose Tissue Energy Homeostasis and Inflammation. Cell Metab. 2018, 27, 378–392. [Google Scholar] [CrossRef] [Green Version]
- Sekar, R.; Chow, B.K.C. Lipolytic actions of secretin in mouse adipocytes. J. Lipid Res. 2014, 55, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Lundbäck, V.; Kulyté, A.; Dahlman, I.; Marcus, C. Adipose-specific inactivation of thyroid stimulating hormone receptors in mice modifies body weight, temperature and gene expression in adipocytes. Physiol. Rep. 2020, 8, e14538. [Google Scholar] [CrossRef]
- Mohammad, S. Role of Free Fatty Acid Receptor 2 (FFAR2) in the Regulation of Metabolic Homeostasis. Curr. Drug Targets 2015, 16, 771–775. [Google Scholar] [CrossRef]
- Gan, L.; Liu, Z.; Chen, Y.; Luo, D.; Feng, F.; Liu, G.; Sun, C. α-MSH and Foxc2 promote fatty acid oxidation through C/EBPβ negative transcription in mice adipose tissue. Sci. Rep. 2016, 6, 36661. [Google Scholar] [CrossRef] [Green Version]
- Hudson, A.D.; Kauffman, A.S. Metabolic actions of kisspeptin signaling: Effects on body weight, energy expenditure, and feeding. Pharmacol. Ther. 2021, 231, 107974. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Sano, T.; Nabetani, T.; Asano, Y.; Hirabayashi, Y. GPRC5B Activates Obesity-Associated Inflammatory Signaling in Adipocytes. Sci. Signal. 2012, 5, ra85. [Google Scholar] [CrossRef] [Green Version]
- Xue, W.; Fan, Z.; Li, L.; Lu, J.; Zhai, Y.; Zhao, J. The chemokine system and its role in obesity. J. Cell. Physiol. 2018, 234, 3336–3346. [Google Scholar] [CrossRef]
- Xu, L.; Kitade, H.; Ni, Y.; Ota, T. Roles of Chemokines and Chemokine Receptors in Obesity-Associated Insulin Resistance and Nonalcoholic Fatty Liver Disease. Biomolecules 2015, 5, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Gericke, M.T.; Schröder, T.; Kosacka, J.; Nowicki, M.; Klöting, N.; Spanel-Borowski, K. Neuropeptide Y impairs insulin-stimulated translocation of glucose transporter 4 in 3T3-L1 adipocytes through the Y1 receptor. Mol. Cell. Endocrinol. 2012, 348, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-H.; Ann-Onda, D.; Lin, X.; Fynch, S.; Nadarajah, S.; Pappas, E.G.; Liu, X.; Scott, J.W.; Oakhill, J.S.; Galic, S.; et al. Neuropeptide Y1 receptor antagonism protects β-cells and improves glycemic control in type 2 diabetes. Mol. Metab. 2022, 55, 101413. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Bonillas, A.C.; Wong, J.; Padilla, S.L.; Brooks, V.L. Neuropeptide Y suppresses thermogenic and cardiovascular sympathetic nerve activity via Y1 receptors in the paraventricular nucleus and dorsomedial hypothalamus. J. Neuroendocr. 2021, 33, e13006. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Zeng, T.; Lee, K.; Nobis, M.; Loh, K.; Gou, L.; Xia, Z.; Gao, Z.; Bensellam, M.; Hughes, W.; et al. Peripheral-specific Y1 receptor antagonism increases thermogenesis and protects against diet-induced obesity. Nat. Commun. 2021, 12, 2622. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.-Y.; Liu, Z.; Guo, F.; Yu, J.; Li, H.; Yin, X. Adipocyte ADRB3 Down-Regulated in Chinese Overweight Individuals Adipocyte ADRB3 in Overweight. Obes. Facts 2018, 11, 524–533. [Google Scholar] [CrossRef]
- Valentine, J.M.; Ahmadian, M.; Keinan, O.; Abu-Odeh, M.; Zhao, P.; Zhou, X.; Keller, M.P.; Gao, H.; Yu, R.T.; Liddle, C.; et al. β3-Adrenergic receptor downregulation leads to adipocyte catecholamine resistance in obesity. J. Clin. Investig. 2022, 132, e153357. [Google Scholar] [CrossRef]
- Tekola-Ayele, F.; Lee, A.; Workalemahu, T.; Sanchez-Pozos, K. Shared genetic underpinnings of childhood obesity and adult cardiometabolic diseases. Hum. Genom. 2019, 13, e153357. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhao, S.; Zhu, T.; Zhang, J. GPRC5A Is a Negative Regulator of the Pro-Survival PI3K/Akt Signaling Pathway in Triple-Negative Breast Cancer. Front. Oncol. 2021, 10, 624493. [Google Scholar] [CrossRef]
- Yosten, G.L.C.; Kolar, G.R.; Redlinger, L.J.; Samson, W.K. Evidence for an interaction between proinsulin C-peptide and GPR. J. Endocrinol. 2013, 218, B1–B8. [Google Scholar] [CrossRef]
- Yu, H.; Rimbert, A.; Palmer, A.E.; Toyohara, T.; Xia, Y.; Xia, F.; Ferreira, L.M.; Chen, Z.; Chen, T.; Loaiza, N.; et al. GPR146 Deficiency Protects against Hypercholesterolemia and Atherosclerosis. Cell 2019, 179, 1276–1288. [Google Scholar] [CrossRef]
- Fernández-Ruiz, I. GPR146 is a potential new therapeutic target for lipid lowering. Nat. Rev. Cardiol. 2019, 17, 132–133. [Google Scholar] [CrossRef]
- Han, F.; Liu, X.; Chen, C.; Liu, Y.; Du, M.; Zhou, Y.; Liu, Y.; Song, B.-L.; He, H.H.; Wang, Y. Hypercholesterolemia risk-associated GPR146 is an orphan G-protein coupled receptor that regulates blood cholesterol levels in humans and mice. Cell Res. 2020, 30, 363–365. [Google Scholar] [CrossRef]
- Ren, J.; Pan, X.; Li, L.; Huang, Y.; Huang, H.; Gao, Y.; Xu, H.; Qu, F.; Chen, L.; Wang, L.; et al. Knockdown ofGPR137,G Protein-coupled receptor 137, Inhibits the Proliferation and Migration of Human Prostate Cancer Cells. Chem. Biol. Drug Des. 2016, 87, 704–713. [Google Scholar] [CrossRef]
- Lu, J.; Zhong, F.; Sun, B.; Wang, C. GPR137 is a promising novel bio-marker for the prognosis of bladder cancer patients. Medicine 2019, 98, e16576. [Google Scholar] [CrossRef]
- Yang, S.Y.; Kim, S.A.; Hur, G.M.; Park, M.; Park, J.-E.; Yoo, H.J. Replicative genetic association study between functional polymorphisms in AVPR1A and social behavior scales of autism spectrum disorder in the Korean population. Mol. Autism 2017, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Avinun, R.; Israel, S.; Shalev, I.; Gritsenko, I.; Bornstein, G.; Ebstein, R.P.; Knafo, A. AVPR1A Variant Associated with Preschoolers’ Lower Altruistic Behavior. PLoS ONE 2011, 6, e25274. [Google Scholar] [CrossRef]
- Fenner, A. AVPR1A: A target in CRPC? Nat. Rev. Urol. 2019, 16, 508. [Google Scholar] [CrossRef]
- Urbach, J.; Goldsmith, S.R. Vasopressin antagonism in heart failure: A review of the hemodynamic studies and major clinical trials. Ther. Adv. Cardiovasc. Dis. 2021, 15, 1753944720977741. [Google Scholar] [CrossRef]
- Mok, J.; Park, T.S.; Kim, S.; Kim, D.; Choi, C.S.; Park, J. Prokineticin receptor 1 ameliorates insulin resistance in skeletal muscle. FASEB J. 2020, 35, e21179. [Google Scholar] [CrossRef]
- Zou, Y.; Ning, T.; Shi, J.; Chen, M.; Ding, L.; Huang, Y.; Kauderer, S.; Xu, M.; Cui, B.; Bi, Y.; et al. Association of a gain-of-function variant in LGR4 with central obesity. Obesity 2017, 25, 252–260. [Google Scholar] [CrossRef]
- Wang, J.; Liu, R.; Wang, F.; Hong, J.; Li, X.; Chen, M.; Ke, Y.; Zhang, X.; Ma, Q.; Wang, R.; et al. Ablation of LGR4 promotes energy expenditure by driving white-to-brown fat switch. Nature 2013, 15, 1455–1463. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Only in WAT | Only in BAT |
|---|---|
| Mrgprb3 | Lpar1 |
| Mrgpra1 | Tacr2 |
| Ffar3 | Adgrf3 |
| Mrgpra3 | Nmbr |
| Gpr12 | Adgrb3 |
| Ltb4r2 | Mchr1 |
| Gpr119 | Drd2 |
| Npy2r | |
| Gpr6 | |
| Tacr1 | |
| Npy5r | |
| Npbwr1 | |
| Oxgr1 | |
| Rgr | |
| Grm4 | |
| Gpr83 | |
| Hrh4 | |
| Hcrtr2 | |
| Ptgdr | |
| Gpr139 | |
| Glp2r | |
| Npsr1 | |
| Cckar | |
| Tacr3 | |
| Glp1r | |
| Galr3 | |
| Grm6 | |
| Nmur1 |
| Name | Log Fold Change | Role in Adipose | References |
|---|---|---|---|
| Adora1 | 7.865386 | Induces lipolysis and increases energy expenditure | [19] |
| Adgrf5 | 7.774302 | promotes insulin signaling and insulin-mediated glucose uptake | [20] |
| Ffar4 | 6.765778 | Enhances insulin sensitivity, increases glucose uptake and thermogenesis. | [7] |
| Gprc5c | 5.247853 | Unknown | |
| Gipr | 5.197849 | Enhances insulin sensitivity, increases glucose uptake and lipoprotein lipase activity. | [29,30,31] |
| Hcar2 | 4.04649 | Unknown | |
| Gpr35 | 3.526624 | Regulates lipid metabolism, thermogenic, and anti-inflammatory gene expression in adipose tissue | [32] |
| Adrb2 | 3.493813 | Regulates lipolysis and thermogenesis | [11,12,13,14] |
| Sctr | 3.402992 | Induces lipolysis | [33] |
| Tshr | 3.266825 | Regulates BAT adipogenesis | [34] |
| Ffar2 | 2.625755 | Regulated adipogenesis | [7,35] |
| Ccr10 | 2.592673 | Unknown | |
| Mc2r | 2.541011 | Unknown | |
| Mc5r | 2.435111 | Promotes lipolysis and impairs re-esterification in adipocytes | [36] |
| Adgrd1 | 2.292871 | Unknown | |
| Hcar1 | 2.172346 | Inhibits lipolysis and stimulates glucose uptake | [22,23] |
| Adgrg1 | 2.029279 | Regulated adipogenesis | [20] |
| Adgrg7 | 1.64874 | Unknown | |
| Gprc5a | 1.279647 | Unknown | |
| Gpr146 | 1.114973 | Unknown | |
| Kiss1r | 1.020995 | Promotes adipocyte differentiation and fat accumulation | [37] |
| Gprc5b | 0.974769 | Activates obesity-associated inflammatory signaling in adipocytes | [38] |
| Gpr17 | 0.948663 | Unknown | |
| F2r | 0.738862 | Unknown | |
| Gpr137 | 0.717915 | Unknown |
| GPCR Name | Log Fold Change (Human Adipose) | Log Fold Hange 3T3-L1 | Expression in Mouse Adipose | Previously Studied in Adipose | ||
|---|---|---|---|---|---|---|
| Healthy Obese vs. Lean | Unhealthy Obese vs. Lean | Day 7 vs. Day 0 | WAT | BAT | ||
| GPR50 | −0.680 | −2.517 | Not detected | No | No | Yes |
| GPR179 | −0.657 | −2.153 | Not detected | No | No | No |
| ADRB3 | −0.759 | −1.694 | 2.36 | Yes | Yes | Yes |
| PROKR1 | −1.054 | −1.666 | −4.89 | Yes | Yes | yes |
| SSTR1 | −0.387 | −1.322 | −1.81 | Yes | Yes | Yes |
| ADGRB3 | −0.635 | −1.237 | Not detected | No | Yes | No |
| CALCRL | −0.835 | −1.206 | −3.56 | Yes | Yes | Yes |
| GIPR | −0.838 | −0.955 | 5.19 | Yes | Yes | Yes |
| GPR146 | −0.520 | −0.725 | 1.11 | Yes | Yes | No |
| CHRM3 | −0.441 | −0.709 | −2.7 | Yes | Yes | No |
| LGR4 | −0.428 | −0.695 | −1.26 | Yes | Yes | Yes |
| GPR63 | −0.590 | −0.653 | −0.05 | Yes | Yes | No |
| AVPR1A | −0.521 | −0.608 | ND | Yes | Yes | No |
| ADGRL2 | −0.341 | −0.528 | −2.28 | Yes | Yes | No |
| SSTR2 | −0.314 | −0.519 | 0.67 | Yes | Yes | No |
| AGTR1 | −0.309 | −0.489 | −3.50 | Yes | Yes | NO |
| ADCYAP1R1 | −0.441 | −0.473 | −1.30 | Yes | Yes | Yes |
| HCAR1 | −0.341 | −0.426 | 2.17 | Yes | Yes | Yes |
| EDNRA | −0.080 | −0.418 | −0.57 | Yes | Yes | Yes |
| GPCR Name | Log Fold Change Human Adipose | Log Fold Change 3T3-L1 | Expression in Mouse Adipose | Previously Studied in Adipose | ||
|---|---|---|---|---|---|---|
| Healthy Obese vs. Lean | Unhealthy Obese vs. Lean | Day 7 vs. Day 0 | WAT | BAT | ||
| C3AR1 | 1.306 | 3.748 | −0.67 | Yes | Yes | Yes |
| CCR5 | 2.391 | 2.494 | ND | Yes | Yes | Yes |
| CCR7 | 1.51 | 2.377 | −2.20 | Yes | Yes | Yes |
| ADGRG3 | 0.876 | 2.258 | −2.72 | Yes | Yes | Yes |
| FPR1 | 0.93 | 2.117 | −1.83 | Yes | Yes | Yes |
| CXCR2 | 0.956 | 1.989 | −2.16 | Yes | Yes | Yes |
| CCR2 | 1.485 | 1.959 | −2.10 | Yes | Yes | Yes |
| GPR183 | 1.463 | 1.725 | −1.52 | Yes | Yes | Yes |
| CXCR4 | 1.053 | 1.702 | ND | Yes | Yes | Yes |
| CCR1 | 1.596 | 1.67 | −1.66 | Yes | Yes | Yes |
| FPR3 | 1.524 | 1.666 | ND | No | No | No |
| HTR7 | 1.611 | 1.655 | ND | Yes | Yes | No |
| S1PR4 | 0.864 | 1.633 | −2.53 | Yes | Yes | No |
| ADGRE1 | 0.919 | 1.623 | ND | Yes | Yes | No |
| CX3CR1 | 1.285 | 1.622 | −1.97 | Yes | Yes | Yes |
| GPR132 | 1.141 | 1.534 | −2.08 | Yes | Yes | Yes |
| ADORA3 | 1.057 | 1.44 | ND | Trace | Trace | No |
| LPAR2 | 0.611 | 1.432 | −1.53 | Trace | Trace | Yes |
| CELSR1 | 1.058 | 1.379 | ND | Yes | Yes | No |
| HRH2 | 0.806 | 1.354 | −2.10 | Yes | Yes | Yes |
| LPAR5 | 0.841 | 1.334 | −1.91 | Yes | Yes | No |
| P2RY13 | 0.754 | 1.314 | −2.04 | Yes | Yes | No |
| FPR2 | 0.713 | 1.295 | −2.27 | Yes | Trace | Yes |
| GPR85 | 1.038 | 1.253 | −3.55 | Yes | Yes | No |
| ADGRE2 | 0.74 | 1.227 | ND | No | No | No |
| DRD1 | −0.073 | 1.211 | −1.53 | Yes | trace | Yes |
| GPR162 | 0.923 | 1.2 | −1.40 | Yes | Yes | No |
| LHCGR | 1.004 | 1.151 | ND | Yes | Yes | No |
| PTAFR | 0.77 | 1.148 | ND | Yes | Yes | Yes |
| HRH1 | 0.833 | 1.084 | −1.63 | Yes | Yes | No |
| CMKLR1 | 0.809 | 1.047 | −2.51 | Yes | Yes | Yes |
| OPRK1 | 1.717 | 1.014 | ND | No | No | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Mahri, S.; Okla, M.; Rashid, M.; Malik, S.S.; Iqbal, J.; Al Ibrahim, M.; Dairi, G.; Mahmood, A.; Muthurangan, M.; Yaqinuddin, A.; et al. Profiling of G-Protein Coupled Receptors in Adipose Tissue and Differentiating Adipocytes Offers a Translational Resource for Obesity/Metabolic Research. Cells 2023, 12, 377. https://doi.org/10.3390/cells12030377
Al Mahri S, Okla M, Rashid M, Malik SS, Iqbal J, Al Ibrahim M, Dairi G, Mahmood A, Muthurangan M, Yaqinuddin A, et al. Profiling of G-Protein Coupled Receptors in Adipose Tissue and Differentiating Adipocytes Offers a Translational Resource for Obesity/Metabolic Research. Cells. 2023; 12(3):377. https://doi.org/10.3390/cells12030377
Chicago/Turabian StyleAl Mahri, Saeed, Meshail Okla, Mamoon Rashid, Shuja Shafi Malik, Jahangir Iqbal, Maria Al Ibrahim, Ghida Dairi, Amer Mahmood, Manikandan Muthurangan, Ahmed Yaqinuddin, and et al. 2023. "Profiling of G-Protein Coupled Receptors in Adipose Tissue and Differentiating Adipocytes Offers a Translational Resource for Obesity/Metabolic Research" Cells 12, no. 3: 377. https://doi.org/10.3390/cells12030377
APA StyleAl Mahri, S., Okla, M., Rashid, M., Malik, S. S., Iqbal, J., Al Ibrahim, M., Dairi, G., Mahmood, A., Muthurangan, M., Yaqinuddin, A., & Mohammad, S. (2023). Profiling of G-Protein Coupled Receptors in Adipose Tissue and Differentiating Adipocytes Offers a Translational Resource for Obesity/Metabolic Research. Cells, 12(3), 377. https://doi.org/10.3390/cells12030377