

Genetic and Chemical Controls of Sperm Fate and Spermatocyte Dedifferentiation via PUF-8 and MPK-1 in Caenorhabditis elegans
 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Worm Maintenance and Strains
2.2. RNA Interference (RNAi)
2.3. Generation of puf-8(q725)/mln1[mIs14 dpy-10(e128)]; fem-3(q20gf) Mutants
2.4. Generation of lin-41(tn1541[GFP::tev::s::lin-41]); puf-8(q725)/mln1[mIs14 dpy-10(e128)]; fem-3(q20gf) Mutant
2.5. Germline Antibody Staining
2.6. 5-Ethynyl-2′-deoxyuridine (EdU) Labeling
2.7. Resveratrol (RSV) Treatment
2.8. Cell Culture and RSV Treatment
2.9. Western Blot Analysis
3. Results
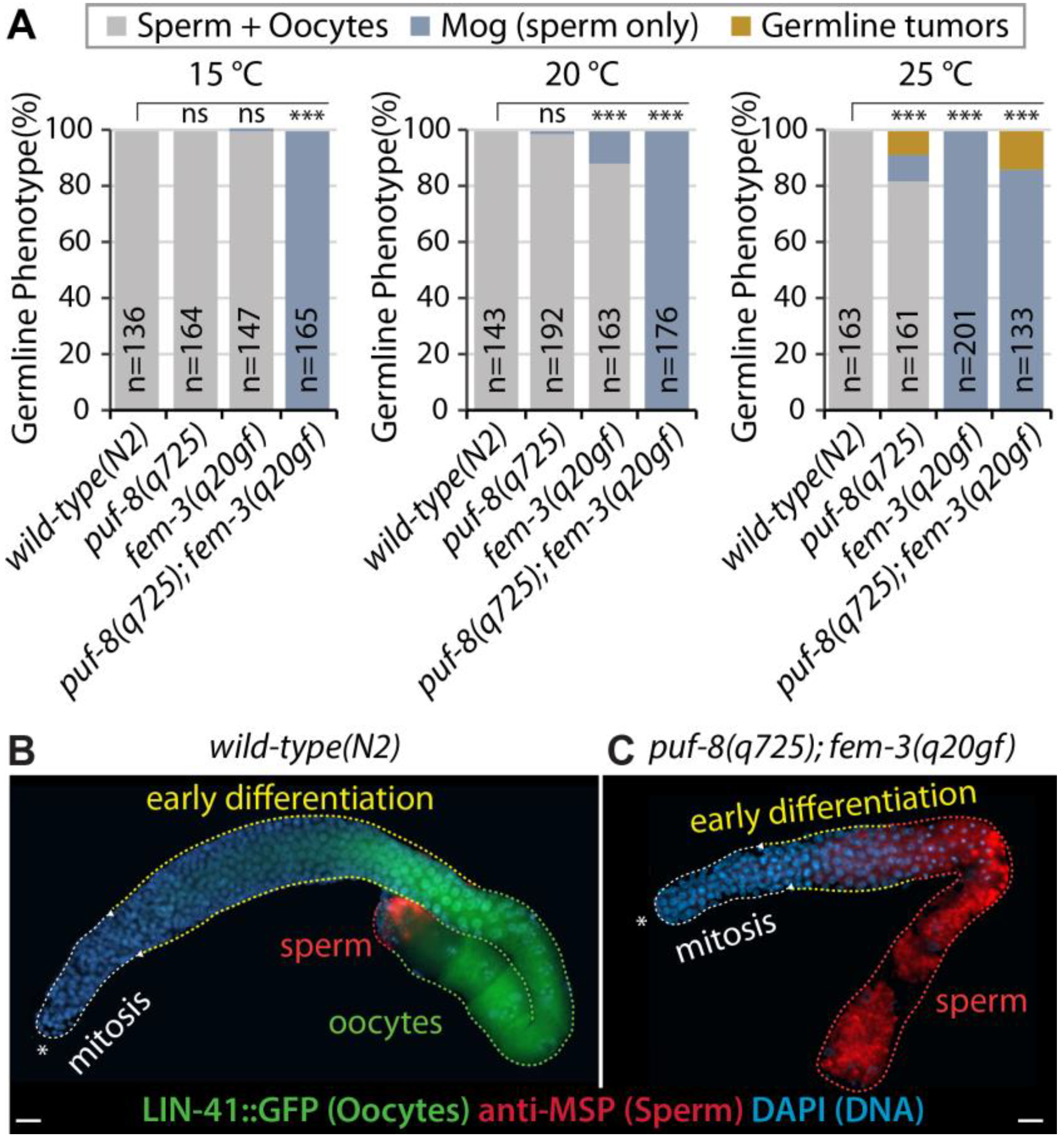
3.1. puf-8(q725) Mutation Enhances fem-3(q20gf) Mog Phenotype
3.2. RNAi of fog-1, fog-2, or fog-3 Rescues puf-8(q725); fem-3(q20gf) Mog Phenotype
3.3. MPK-1 Dependence of puf-8(q725); lip-1(zh15) and puf-8(q725); fem-3(q20gf) Mog Phenotype
3.4. Competence for Spermatocyte Dedifferentiation in the Absence of PUF-8
3.5. Resveratrol Induces Germline Tumors by Activating MPK-1 in the Absence of PUF-8
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Van Pelt, K.M.; Truttmann, M.C. Caenorhabditis elegans as a model system for studying aging-associated neurodegenerative diseases. Transl. Med. Aging 2020, 4, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Markaki, M.; Tavernarakis, N. Caenorhabditis elegans as a model system for human diseases. Curr. Opin. Biotechnol. 2020, 63, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Kobet, R.A.; Pan, X.; Zhang, B.; Pak, S.C.; Asch, A.S.; Lee, M.H. Caenorhabditis elegans: A Model System for Anti-Cancer Drug Discovery and Therapeutic Target Identification. Biomol. Ther. 2014, 22, 371–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.; Sternberg, P.W. C. elegans vulval development as a model system to study the cancer biology of EGFR signaling. Cancer Metastasis Rev. 1999, 18, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, E.J.; Greenstein, D. Introduction to the germ line. WormBook Online Rev. C. Elegans Biol. 2005, 1–4. [Google Scholar] [CrossRef]
- Wickens, M.; Bernstein, D.S.; Kimble, J.; Parker, R. A PUF family portrait: 3’UTR regulation as a way of life. Trends Genet. TIG 2002, 18, 150–157. [Google Scholar] [CrossRef]
- Kimble, J.; Crittenden, S.L. Germline proliferation and its control. WormBook Online Rev. C. Elegans Biol. 2005, 1–14. [Google Scholar] [CrossRef]
- Kimble, J.; Crittenden, S.L. Controls of germline stem cells, entry into meiosis, and the sperm/oocyte decision in Caenorhabditis elegans. Annu. Rev. Cell Dev. Biol. 2007, 23, 405–433. [Google Scholar] [CrossRef] [Green Version]
- Greenwald, I. LIN-12/Notch signaling in C. elegans. WormBook Online Rev. C. Elegans Biol. 2005, 1–16. [Google Scholar] [CrossRef]
- Shin, H.; Haupt, K.A.; Kershner, A.M.; Kroll-Conner, P.; Wickens, M.; Kimble, J. SYGL-1 and LST-1 link niche signaling to PUF RNA repression for stem cell maintenance in Caenorhabditis elegans. PLoS Genet. 2017, 13, e1007121. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Wu, X.; Zhu, Y. RNA-binding protein PUM2 regulates mesenchymal stem cell fate via repression of JAK2 and RUNX2 mRNAs. J. Cell. Physiol. 2020, 235, 3874–3885. [Google Scholar] [CrossRef] [PubMed]
- Uyhazi, K.E.; Yang, Y.; Liu, N.; Qi, H.; Huang, X.A.; Mak, W.; Weatherbee, S.D.; de Prisco, N.; Gennarino, V.A.; Song, X.; et al. Pumilio proteins utilize distinct regulatory mechanisms to achieve complementary functions required for pluripotency and embryogenesis. Proc. Natl. Acad. Sci. USA 2020, 117, 7851–7862. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.C.; Randolph, L.K.; Iascone, D.M.; Pernice, H.F.; Polleux, F.; Hengst, U. Pum2 Shapes the Transcriptome in Developing Axons through Retention of Target mRNAs in the Cell Body. Neuron 2019, 104, 931–946.e935. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Chen, D.; Xia, J.; Han, W.; Cui, X.; Neuenkirchen, N.; Hermes, G.; Sestan, N.; Lin, H. Post-transcriptional regulation of mouse neurogenesis by Pumilio proteins. Genes Dev. 2017, 31, 1354–1369. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zheng, W.; Lin, A.; Uyhazi, K.; Zhao, H.; Lin, H. Pumilio 1 suppresses multiple activators of p53 to safeguard spermatogenesis. Curr. Biol. CB 2012, 22, 420–425. [Google Scholar] [CrossRef] [Green Version]
- Mak, W.; Fang, C.; Holden, T.; Dratver, M.B.; Lin, H. An Important Role of Pumilio 1 in Regulating the Development of the Mammalian Female Germline. Biol. Reprod. 2016, 94, 134. [Google Scholar] [CrossRef]
- D’Amico, D.; Mottis, A.; Potenza, F.; Sorrentino, V.; Li, H.; Romani, M.; Lemos, V.; Schoonjans, K.; Zamboni, N.; Knott, G.; et al. The RNA-Binding Protein PUM2 Impairs Mitochondrial Dynamics and Mitophagy During Aging. Mol. Cell 2019, 73, 775–787 e710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Voronina, E. Diverse Roles of PUF Proteins in Germline Stem and Progenitor Cell Development in C. elegans. Front. Cell Dev. Biol. 2020, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Datla, U.S.; Scovill, N.C.; Brokamp, A.J.; Kim, E.; Asch, A.S.; Lee, M.H. Role of PUF-8/PUF protein in stem cell control, sperm-oocyte decision and cell fate reprogramming. J. Cell. Physiol. 2014, 229, 1306–1311. [Google Scholar] [CrossRef]
- Park, Y.; O’Rourke, S.; Taki, F.A.; Alfhili, M.A.; Lee, M.H. Dose-Dependent Effects of GLD-2 and GLD-1 on Germline Differentiation and Dedifferentiation in the Absence of PUF-8. Front. Cell Dev. Biol. 2020, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Ariz, M.; Mainpal, R.; Subramaniam, K.C. elegans RNA-binding proteins PUF-8 and MEX-3 function redundantly to promote germline stem cell mitosis. Dev. Biol. 2009, 326, 295–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, D.S.; Datla, U.S.; Hollis, S.E.; Kimble, J.; Lee, M.H. The Ras-ERK MAPK regulatory network controls dedifferentiation in Caenorhabditis elegans germline. Biochim. Biophys. Acta 2012, 1823, 1847–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, K.; Seydoux, G. Dedifferentiation of primary spermatocytes into germ cell tumors in C. elegans lacking the pumilio-like protein PUF-8. Curr. Biol. CB 2003, 13, 134–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, C.T.; Lee, M.H.; Kimble, J. Chemical reprogramming of Caenorhabditis elegans germ cell fate. Nat. Chem. Biol. 2010, 6, 102–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, J.A.; Cummings, E.E.; O’Reilly, L.P.; Lee, M.H.; Pak, S.C. A high-content assay for identifying small molecules that reprogram C. elegans germ cell fate. Methods 2014, 68, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.X.; Hu, D.H.; Jia, C.Y.; Fu, X.B. Progress of cellular dedifferentiation research. Chin. J. Traumatol. Zhonghua Chuang Shang Za Zhi 2006, 9, 308–315. [Google Scholar] [PubMed]
- Tiong, Y.L.; Ng, K.Y.; Koh, R.Y.; Ponnudurai, G.; Chye, S.M. Melatonin promotes Schwann cell dedifferentiation and proliferation through the Ras/Raf/ERK and MAPK pathways, and glial cell-derived neurotrophic factor expression. Exp. Ther. Med. 2020, 20, 16. [Google Scholar] [CrossRef]
- Almairac, F.; Turchi, L.; Sakakini, N.; Debruyne, D.N.; Elkeurti, S.; Gjernes, E.; Polo, B.; Bianchini, L.; Fontaine, D.; Paquis, P.; et al. ERK-Mediated Loss of miR-199a-3p and Induction of EGR1 Act as a “Toggle Switch” of GBM Cell Dedifferentiation into NANOG- and OCT4-Positive Cells. Cancer Res. 2020, 80, 3236–3250. [Google Scholar] [CrossRef]
- Boerboom, A.; Reusch, C.; Pieltain, A.; Chariot, A.; Franzen, R. KIAA1199: A novel regulator of MEK/ERK-induced Schwann cell dedifferentiation. Glia 2017, 65, 1682–1696. [Google Scholar] [CrossRef]
- Chen, R.; Lee, W.Y.; Zhang, X.H.; Zhang, J.T.; Lin, S.; Xu, L.L.; Huang, B.; Yang, F.Y.; Liu, H.L.; Wang, B.; et al. Epigenetic Modification of the CCL5/CCR1/ERK Axis Enhances Glioma Targeting in Dedifferentiation-Reprogrammed BMSCs. Stem Cell Rep. 2017, 8, 743–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrisingh, M.C.; Perez-Nadales, E.; Parkinson, D.B.; Malcolm, D.S.; Mudge, A.W.; Lloyd, A.C. The Ras/Raf/ERK signalling pathway drives Schwann cell dedifferentiation. EMBO J. 2004, 23, 3061–3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamath, R.S.; Martinez-Campos, M.; Zipperlen, P.; Fraser, A.G.; Ahringer, J. Effectiveness of specific RNA-mediated interference through ingested double-stranded RNA in Caenorhabditis elegans. Genome Biol. 2001, 2, research0002.1. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Hook, B.; Pan, G.; Kershner, A.M.; Merritt, C.; Seydoux, G.; Thomson, J.A.; Wickens, M.; Kimble, J. Conserved regulation of MAP kinase expression by PUF RNA-binding proteins. PLoS Genet. 2007, 3, e233. [Google Scholar] [CrossRef] [Green Version]
- Yoon, D.S.; Pendergrass, D.L.; Lee, M.H. A simple and rapid method for combining fluorescent in situ RNA hybridization (FISH) and immunofluorescence in the C. elegans germline. MethodsX 2016, 3, 378–385. [Google Scholar] [CrossRef]
- Meier, J.A.; Hyun, M.; Cantwell, M.; Raza, A.; Mertens, C.; Raje, V.; Sisler, J.; Tracy, E.; Torres-Odio, S.; Gispert, S.; et al. Stress-induced dynamic regulation of mitochondrial STAT3 and its association with cyclophilin D reduce mitochondrial ROS production. Sci. Signal. 2017, 10, eaag2588. [Google Scholar] [CrossRef] [Green Version]
- Bachorik, J.L.; Kimble, J. Redundant control of the Caenorhabditis elegans sperm/oocyte switch by PUF-8 and FBF-1, two distinct PUF RNA-binding proteins. Proc. Natl. Acad. Sci. USA 2005, 102, 10893–10897. [Google Scholar] [CrossRef] [Green Version]
- Ahringer, J.; Rosenquist, T.A.; Lawson, D.N.; Kimble, J. The Caenorhabditis elegans sex determining gene fem-3 is regulated post-transcriptionally. EMBO J. 1992, 11, 2303–2310. [Google Scholar] [CrossRef]
- Miller, M.A.; Nguyen, V.Q.; Lee, M.H.; Kosinski, M.; Schedl, T.; Caprioli, R.M.; Greenstein, D. A sperm cytoskeletal protein that signals oocyte meiotic maturation and ovulation. Science 2001, 291, 2144–2147. [Google Scholar] [CrossRef] [Green Version]
- Spike, C.A.; Tsukamoto, T.; Greenstein, D. Ubiquitin ligases and a processive proteasome facilitate protein clearance during the oocyte-to-embryo transition in Caenorhabditis elegans. Genetics 2022, 221, iyac051. [Google Scholar] [CrossRef]
- Ellis, R.E. Sex determination in the Caenorhabditis elegans germ line. Curr. Top. Dev. Biol. 2008, 83, 41–64. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, S.; Puoti, A. Sex determination in the Caenorhabditis elegans germline. Adv. Exp. Med. Biol. 2013, 757, 41–69. [Google Scholar] [CrossRef]
- Ellis, R.; Schedl, T. Sex determination in the germ line. WormBook Online Rev. C. Elegans Biol. 2007, 1–13. [Google Scholar] [CrossRef]
- Lee, M.H.; Kim, K.W.; Morgan, C.T.; Morgan, D.E.; Kimble, J. Phosphorylation state of a Tob/BTG protein, FOG-3, regulates initiation and maintenance of the Caenorhabditis elegans sperm fate program. Proc. Natl. Acad. Sci. USA 2011, 108, 9125–9130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crittenden, S.L.; Bernstein, D.S.; Bachorik, J.L.; Thompson, B.E.; Gallegos, M.; Petcherski, A.G.; Moulder, G.; Barstead, R.; Wickens, M.; Kimble, J. A conserved RNA-binding protein controls germline stem cells in Caenorhabditis elegans. Nature 2002, 417, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Ohmachi, M.; Arur, S.; Nayak, S.; Francis, R.; Church, D.; Lambie, E.; Schedl, T. Multiple functions and dynamic activation of MPK-1 extracellular signal-regulated kinase signaling in Caenorhabditis elegans germline development. Genetics 2007, 177, 2039–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson-Thiewes, S.; Dufour, B.; Martel, P.O.; Lechasseur, X.; Brou, A.A.D.; Roy, V.; Chen, Y.; Kimble, J.; Narbonne, P. Non-autonomous regulation of germline stem cell proliferation by somatic MPK-1/MAPK activity in C. elegans. Cell Rep. 2021, 35, 109162. [Google Scholar] [CrossRef]
- Vaid, S.; Ariz, M.; Chaturbedi, A.; Kumar, G.A.; Subramaniam, K. PUF-8 negatively regulates RAS/MAPK signalling to promote differentiation of C. elegans germ cells. Development 2013, 140, 1645–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, D.S.; Cha, D.S.; Choi, Y.; Lee, J.W.; Lee, M.H. MPK-1/ERK is required for the full activity of resveratrol in extended lifespan and reproduction. Aging Cell 2019, 18, e12867. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Chen, L.; Zhu, F.; Han, X.; Sun, L.; Chen, K. The Cytotoxicity Effect of Resveratrol: Cell Cycle Arrest and Induced Apoptosis of Breast Cancer 4T1 Cells. Toxins 2019, 11, 731. [Google Scholar] [CrossRef]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential Adverse Effects of Resveratrol: A Literature Review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashina, M.; Inoue, S.; Tomihara, K.; Tomita, K.; Hattori, K.; Zhao, Q.L.; Suzuki, T.; Noguchi, M.; Ohashi, W.; Hattori, Y. Different effect of resveratrol to induction of apoptosis depending on the type of human cancer cells. Int. J. Oncol. 2017, 50, 787–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hentze, M.W.; Castello, A.; Schwarzl, T.; Preiss, T. A brave new world of RNA-binding proteins. Nat. Rev. Mol. Cell Biol. 2018, 19, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Shigunov, P.; Sotelo-Silveira, J.; Kuligovski, C.; de Aguiar, A.M.; Rebelatto, C.K.; Moutinho, J.A.; Brofman, P.S.; Krieger, M.A.; Goldenberg, S.; Munroe, D.; et al. PUMILIO-2 is involved in the positive regulation of cellular proliferation in human adipose-derived stem cells. Stem Cells Dev. 2012, 21, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Xu, E.Y.; Chang, R.; Salmon, N.A.; Reijo Pera, R.A. A gene trap mutation of a murine homolog of the Drosophila stem cell factor Pumilio results in smaller testes but does not affect litter size or fertility. Mol. Reprod. Dev. 2007, 74, 912–921. [Google Scholar] [CrossRef]
- Moore, F.L.; Jaruzelska, J.; Fox, M.S.; Urano, J.; Firpo, M.T.; Turek, P.J.; Dorfman, D.M.; Pera, R.A. Human Pumilio-2 is expressed in embryonic stem cells and germ cells and interacts with DAZ (Deleted in AZoospermia) and DAZ-like proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Naudin, C.; Hattabi, A.; Michelet, F.; Miri-Nezhad, A.; Benyoucef, A.; Pflumio, F.; Guillonneau, F.; Fichelson, S.; Vigon, I.; Dusanter-Fourt, I.; et al. PUMILIO/FOXP1 signaling drives expansion of hematopoietic stem/progenitor and leukemia cells. Blood 2017, 129, 2493–2506. [Google Scholar] [CrossRef] [Green Version]
- Murata, Y.; Wharton, R.P. Binding of pumilio to maternal hunchback mRNA is required for posterior patterning in Drosophila embryos. Cell 1995, 80, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Spradling, A.C. A novel group of pumilio mutations affects the asymmetric division of germline stem cells in the Drosophila ovary. Development 1997, 124, 2463–2476. [Google Scholar] [CrossRef]
- Porter, D.F.; Koh, Y.Y.; VanVeller, B.; Raines, R.T.; Wickens, M. Target selection by natural and redesigned PUF proteins. Proc. Natl. Acad. Sci. USA 2015, 112, 15868–15873. [Google Scholar] [CrossRef]
- Ohkuni, K.; Kikuchi, Y.; Hara, K.; Taneda, T.; Hayashi, N.; Kikuchi, A. Suppressor analysis of the mpt5/htr1/uth4/puf5 deletion in Saccharomyces cerevisiae. Mol. Genet. Genom. MGG 2006, 275, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M.; Guarente, L. Saccharomyces cerevisiae MPT5 and SSD1 function in parallel pathways to promote cell wall integrity. Genetics 2002, 160, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Kim, J.Y.; Malik, S.; Son, W.; Kwon, K.S.; Kim, C. Negative regulation of EGFR/MAPK pathway by Pumilio in Drosophila melanogaster. PloS ONE 2012, 7, e34016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takasu, N.; Yamada, T.; Takasu, M.; Komiya, I.; Nagasawa, Y.; Asawa, T.; Shinoda, T.; Aizawa, T.; Koizumi, Y. Disappearance of thyrotropin-blocking antibodies and spontaneous recovery from hypothyroidism in autoimmune thyroiditis. New Engl. J. Med. 1992, 326, 513–518. [Google Scholar] [CrossRef]
- Bansal, N.; Banerjee, D. Tumor initiating cells. Curr. Pharm. Biotechnol. 2009, 10, 192–196. [Google Scholar] [CrossRef]
- Daley, G.Q. Common themes of dedifferentiation in somatic cell reprogramming and cancer. Cold Spring Harb. Symp. Quant. Biol. 2008, 73, 171–174. [Google Scholar] [CrossRef] [Green Version]
- Jopling, C.; Sleep, E.; Raya, M.; Marti, M.; Raya, A.; Izpisua Belmonte, J.C. Zebrafish heart regeneration occurs by cardiomyocyte dedifferentiation and proliferation. Nature 2010, 464, 606–609. [Google Scholar] [CrossRef] [Green Version]
- Kai, T.; Spradling, A. Differentiating germ cells can revert into functional stem cells in Drosophila melanogaster ovaries. Nature 2004, 428, 564–569. [Google Scholar] [CrossRef]
- Brawley, C.; Matunis, E. Regeneration of male germline stem cells by spermatogonial dedifferentiation in vivo. Science 2004, 304, 1331–1334. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Turkel, N.; Hemati, N.; Fuller, M.T.; Hunt, A.J.; Yamashita, Y.M. Centrosome misorientation reduces stem cell division during ageing. Nature 2008, 456, 599–604. [Google Scholar] [CrossRef]
- Tata, P.R.; Mou, H.; Pardo-Saganta, A.; Zhao, R.; Prabhu, M.; Law, B.M.; Vinarsky, V.; Cho, J.L.; Breton, S.; Sahay, A.; et al. Dedifferentiation of committed epithelial cells into stem cells in vivo. Nature 2013, 503, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwitalla, S.; Fingerle, A.A.; Cammareri, P.; Nebelsiek, T.; Goktuna, S.I.; Ziegler, P.K.; Canli, O.; Heijmans, J.; Huels, D.J.; Moreaux, G.; et al. Intestinal tumorigenesis initiated by dedifferentiation and acquisition of stem-cell-like properties. Cell 2013, 152, 25–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.R.; Schedl, T. Mutations in gld-1, a female germ cell-specific tumor suppressor gene in Caenorhabditis elegans, affect a conserved domain also found in Src-associated protein Sam68. Genes Dev. 1995, 9, 1491–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akay, A.; Craig, A.; Lehrbach, N.; Larance, M.; Pourkarimi, E.; Wright, J.E.; Lamond, A.; Miska, E.; Gartner, A. RNA-binding protein GLD-1/quaking genetically interacts with the mir-35 and the let-7 miRNA pathways in Caenorhabditis elegans. Open Biol. 2013, 3, 130151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berset, T.; Hoier, E.F.; Battu, G.; Canevascini, S.; Hajnal, A. Notch inhibition of RAS signaling through MAP kinase phosphatase LIP-1 during C. elegans vulval development. Science 2001, 291, 1055–1058. [Google Scholar] [CrossRef] [PubMed]
- Hajnal, A.; Berset, T. The C.elegans MAPK phosphatase LIP-1 is required for the G(2)/M meiotic arrest of developing oocytes. EMBO J. 2002, 21, 4317–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.H.; Hook, B.; Lamont, L.B.; Wickens, M.; Kimble, J. LIP-1 phosphatase controls the extent of germline proliferation in Caenorhabditis elegans. EMBO J. 2006, 25, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.S.; Zhang, Z.H.; Jin, X.; Wei, P.; Hu, X.Q.; Chen, M.; Lu, C.L.; Lue, Y.H.; Hu, Z.Y.; Sinha Hikim, A.P.; et al. Dedifferentiation of adult monkey Sertoli cells through activation of extracellularly regulated kinase 1/2 induced by heat treatment. Endocrinology 2006, 147, 1237–1245. [Google Scholar] [CrossRef]
- Chen, X.; Mao, Z.; Liu, S.; Liu, H.; Wang, X.; Wu, H.; Wu, Y.; Zhao, T.; Fan, W.; Li, Y.; et al. Dedifferentiation of adult human myoblasts induced by ciliary neurotrophic factor in vitro. Mol. Biol. Cell 2005, 16, 3140–3151. [Google Scholar] [CrossRef] [Green Version]
- Hanley, S.C.; Assouline-Thomas, B.; Makhlin, J.; Rosenberg, L. Epidermal growth factor induces adult human islet cell dedifferentiation. J. Endocrinol. 2011, 211, 231–239. [Google Scholar] [CrossRef]
- Asencio, C.; Rodriguez-Aguilera, J.C.; Ruiz-Ferrer, M.; Vela, J.; Navas, P. Silencing of ubiquinone biosynthesis genes extends life span in Caenorhabditis elegans. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2003, 17, 1135–1137. [Google Scholar] [CrossRef]
- Reinke, S.N.; Hu, X.; Sykes, B.D.; Lemire, B.D. Caenorhabditis elegans diet significantly affects metabolic profile, mitochondrial DNA levels, lifespan and brood size. Mol. Genet. Metab. 2010, 100, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Ronai, D.; Than, M.T.; Walker, C.J.; Sewell, A.K.; Han, M. Nucleotide levels regulate germline proliferation through modulating GLP-1/Notch signaling in C. elegans. Genes Dev. 2016, 30, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Angelo, G.; Van Gilst, M.R. Starvation protects germline stem cells and extends reproductive longevity in C. elegans. Science 2009, 326, 954–958. [Google Scholar] [CrossRef] [PubMed]
- Seidel, H.S.; Kimble, J. Cell-cycle quiescence maintains Caenorhabditis elegans germline stem cells independent of GLP-1/Notch. eLife 2015, 4, e10832. [Google Scholar] [CrossRef] [PubMed]
- Chou, W.Y.; Lin, Y.C.; Lee, Y.H. Short-term starvation stress at young adult stages enhances meiotic activity of germ cells to maintain spermatogenesis in aged male Caenorhabditis elegans. Aging Cell 2019, 18, e12930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiamehr, M.; Heiskanen, L.; Laufer, T.; Dusterloh, A.; Kahraman, M.; Kakela, R.; Laaksonen, R.; Aalto-Setala, K. Dedifferentiation of Primary Hepatocytes is Accompanied with Reorganization of Lipid Metabolism Indicated by Altered Molecular Lipid and miRNA Profiles. Int. J. Mol. Sci. 2019, 20, 2910. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.; Gaddy, M.; Hyun, M.; Jones, M.E.; Aslam, H.M.; Lee, M.H. Genetic and Chemical Controls of Sperm Fate and Spermatocyte Dedifferentiation via PUF-8 and MPK-1 in Caenorhabditis elegans. Cells 2023, 12, 434. https://doi.org/10.3390/cells12030434
Park Y, Gaddy M, Hyun M, Jones ME, Aslam HM, Lee MH. Genetic and Chemical Controls of Sperm Fate and Spermatocyte Dedifferentiation via PUF-8 and MPK-1 in Caenorhabditis elegans. Cells. 2023; 12(3):434. https://doi.org/10.3390/cells12030434
Chicago/Turabian StylePark, Youngyong, Matthew Gaddy, Moonjung Hyun, Mariah E. Jones, Hafiz M. Aslam, and Myon Hee Lee. 2023. "Genetic and Chemical Controls of Sperm Fate and Spermatocyte Dedifferentiation via PUF-8 and MPK-1 in Caenorhabditis elegans" Cells 12, no. 3: 434. https://doi.org/10.3390/cells12030434
APA StylePark, Y., Gaddy, M., Hyun, M., Jones, M. E., Aslam, H. M., & Lee, M. H. (2023). Genetic and Chemical Controls of Sperm Fate and Spermatocyte Dedifferentiation via PUF-8 and MPK-1 in Caenorhabditis elegans. Cells, 12(3), 434. https://doi.org/10.3390/cells12030434