
Lens Epithelial Explants Treated with Vitreous Humor Undergo Alterations in Chromatin Landscape with Concurrent Activation of Genes Associated with Fiber Cell Differentiation and Innate Immune Response

 ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
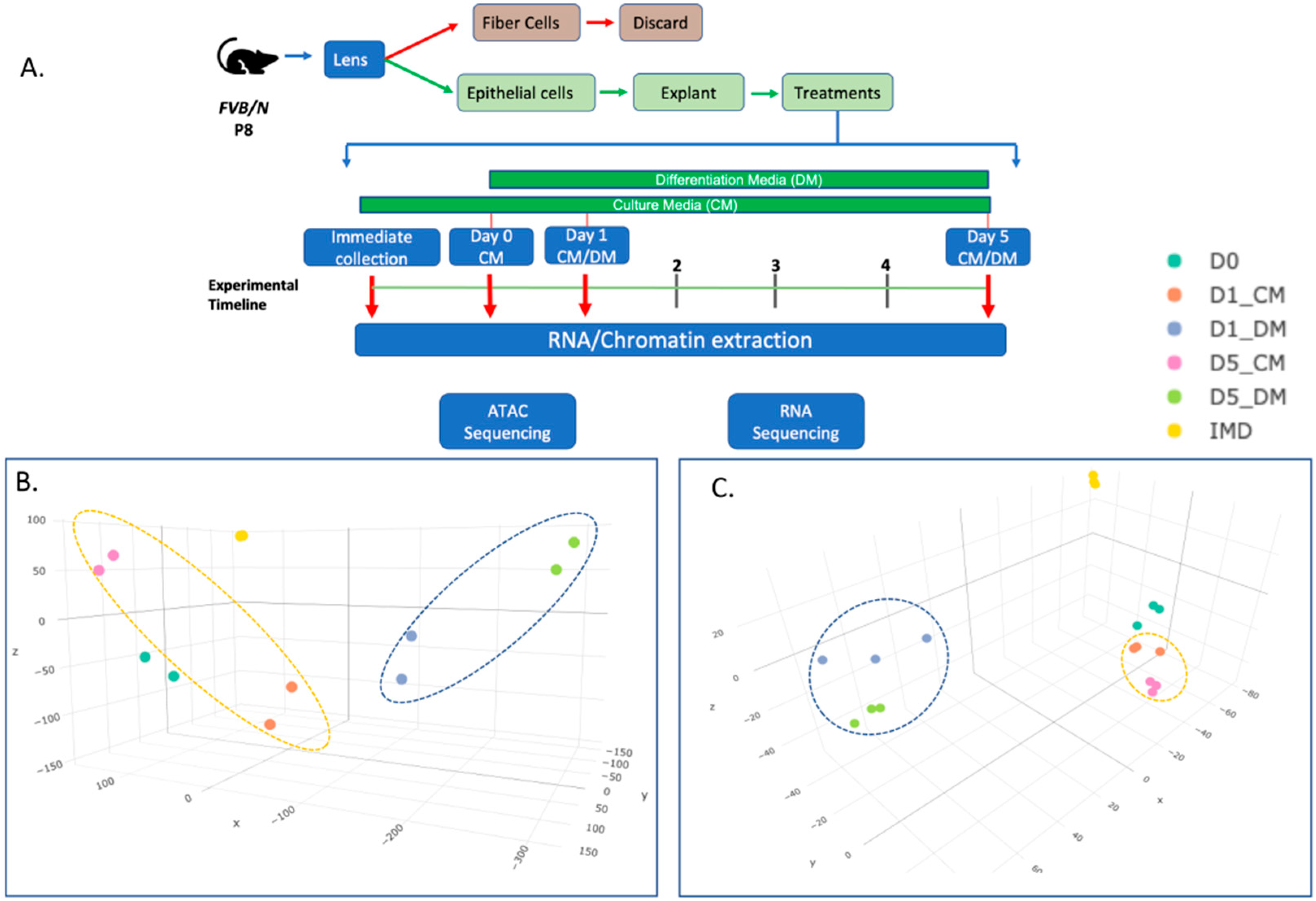
2.1. Animals
2.2. Lens Epithelial Explant Generation and Culture
2.3. RNA-Seq Library Preparation and Sequencing
2.4. RNA Seq Data Analysis
2.5. RT-qPCR Validation of RNA-Seq Data
2.6. ATAC-Seq Library Preparation and Sequencing
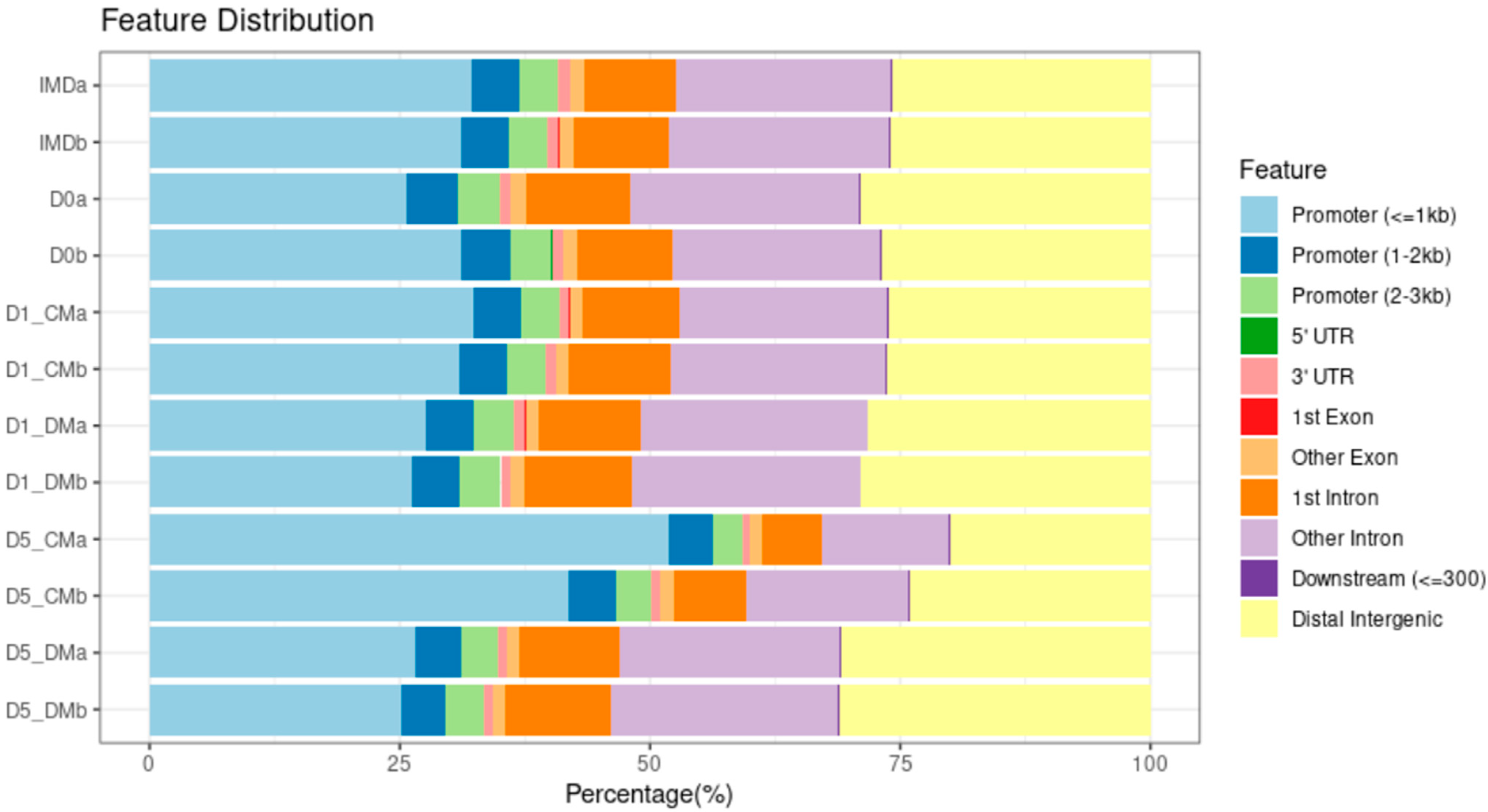
2.7. ATAC-Seq Data Analysis
2.8. Genomic Annotation and Motif Prediction
2.9. Single Cell Data Analysis
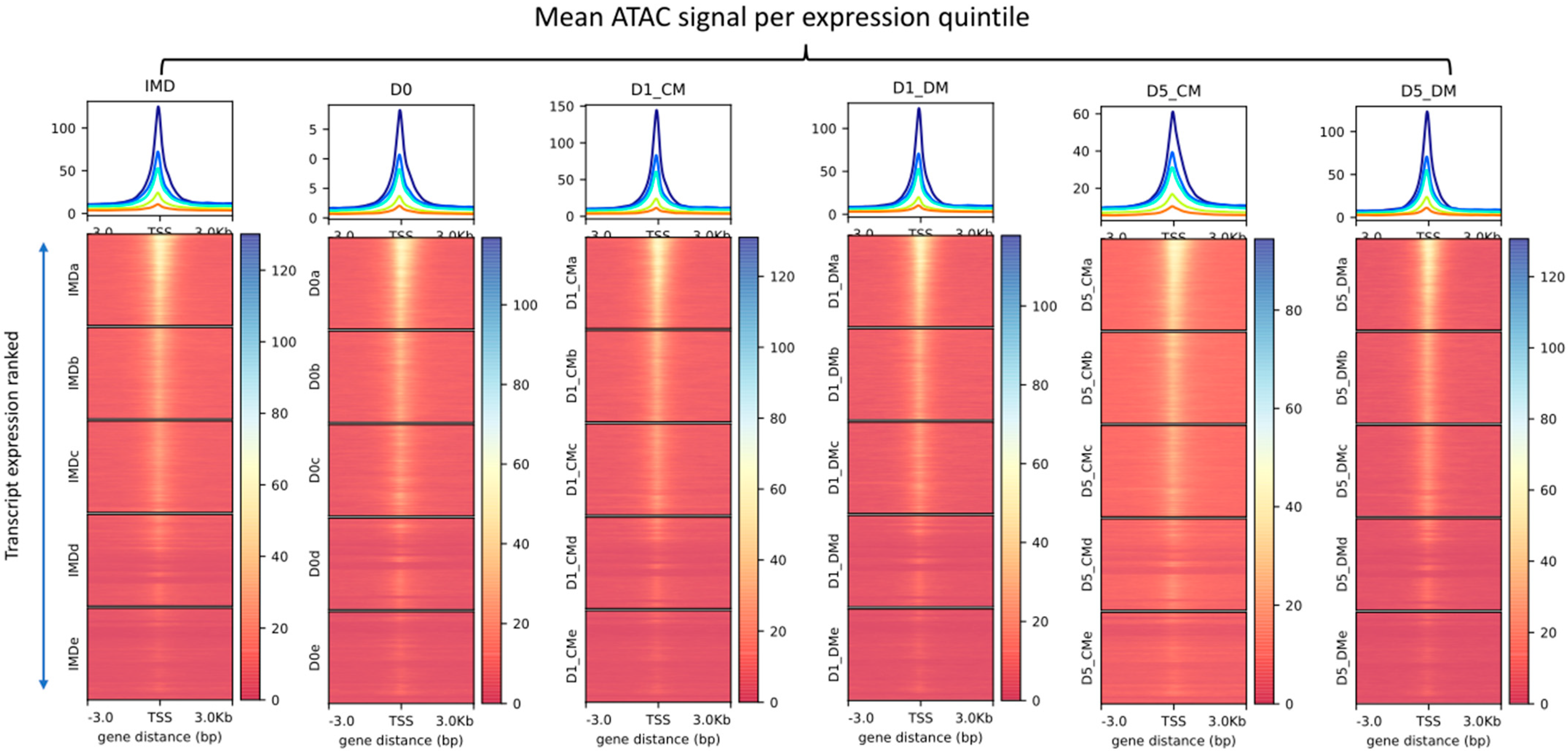
2.10. Relationship between ATAC-Seq and RNA Seq
3. Results
3.1. Chromatin Accessibility and Gene Expression Changes in Lens Epithelial Explants Resulting from Explanting, the Addition of Vitreous, and Time in Culture
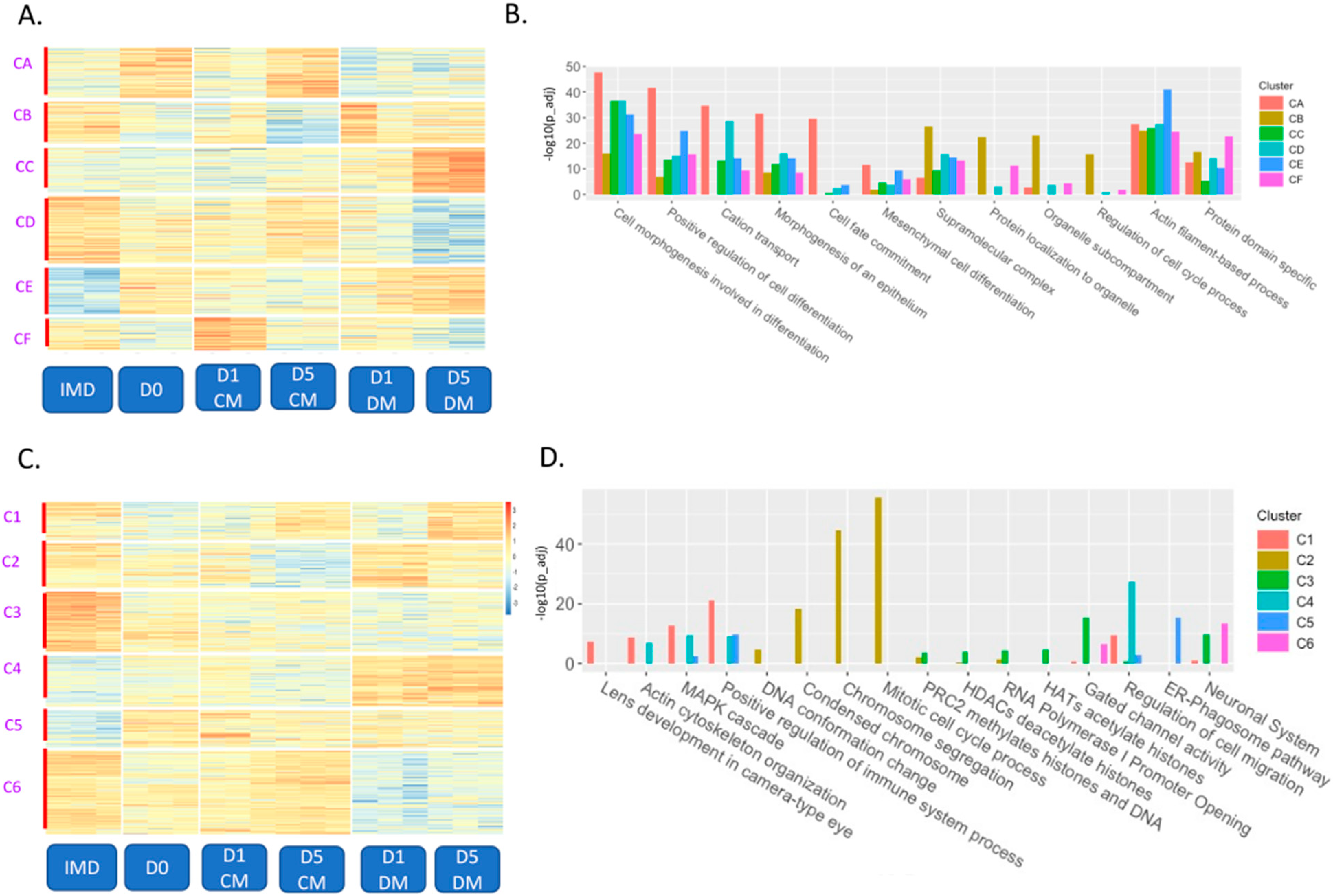
3.2. Clustering of ATAC-Seq and RNA-Seq Data to Evaluate Effects of In Vitro Culture, Time in Culture, and the Addition of Vitreous Humor on Chromatin Organization and Gene Expression
3.3. Assessment of Genome-Wide Dynamic Changes in Chromatin Accessibility with the Addition of Vitreous Humor
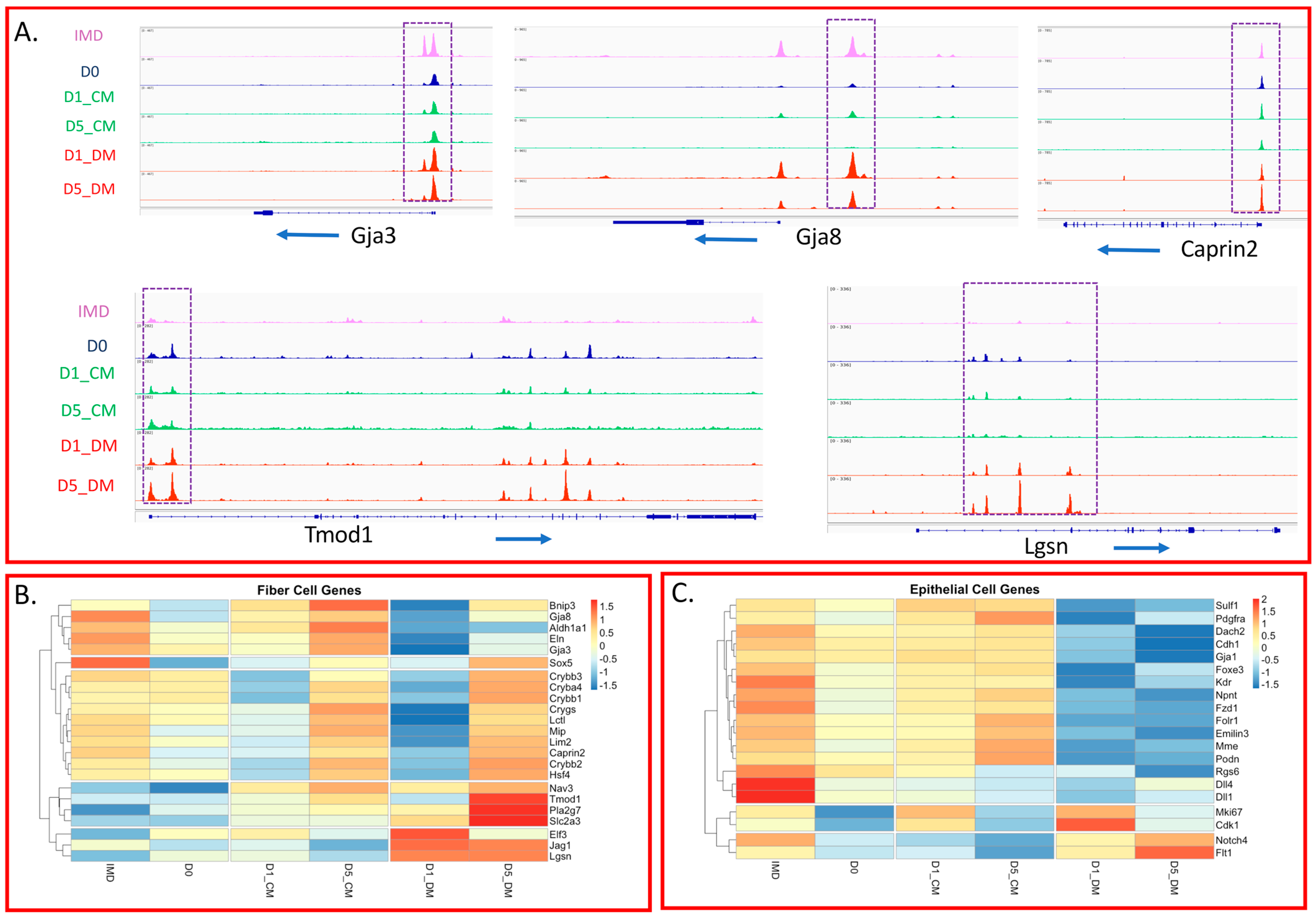
3.4. Specific Changes in Chromatin Accessibility and Gene Expression in Vitreous-Treated Lens Epithelial Explants with Respect to Fiber Cell Differentiation
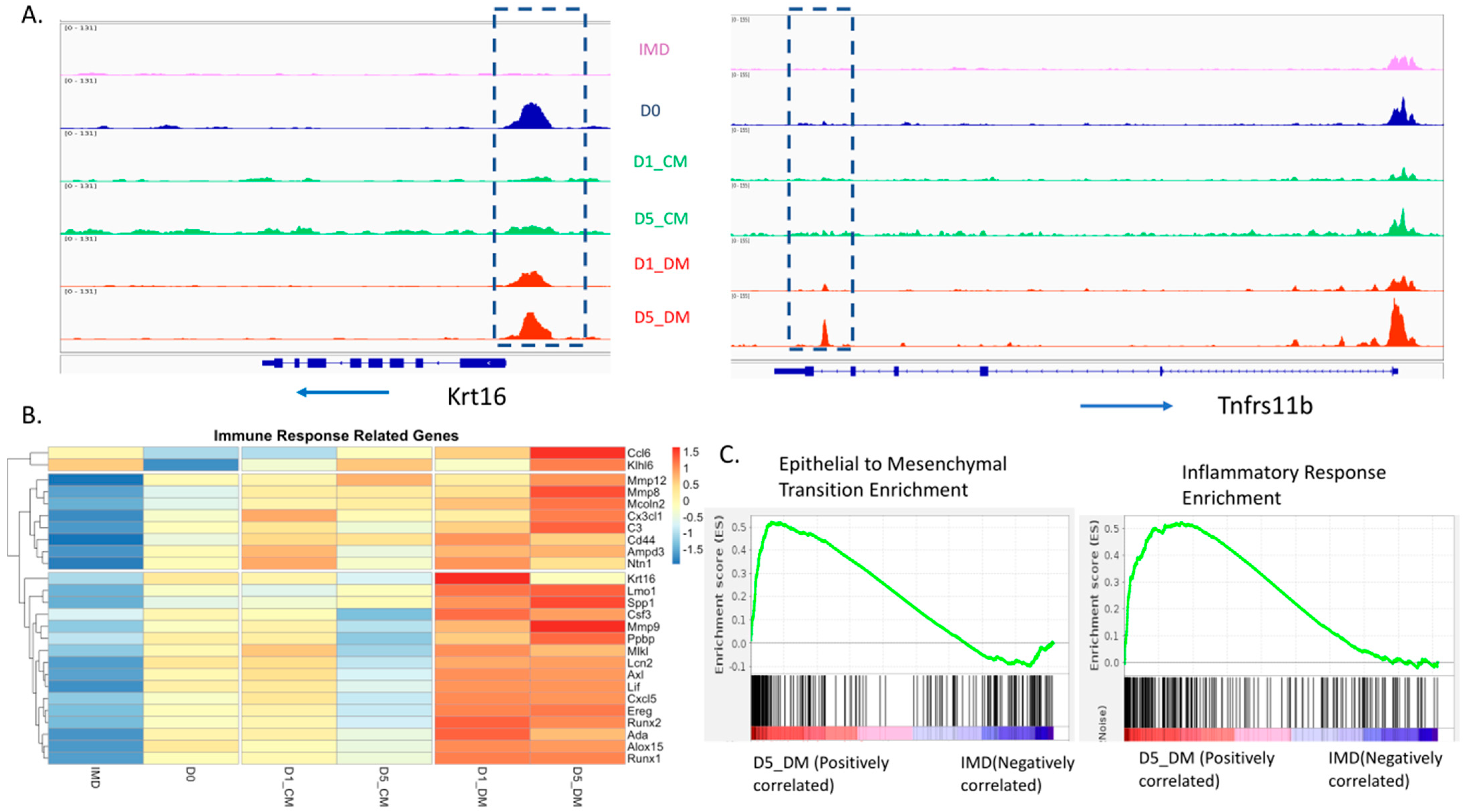
3.5. Increases in Both Promoter Accessibility and Gene Expression for Epithelial to Mesenchymal Transition and Inflammatory Response Genes Induced by Vitreous Humor
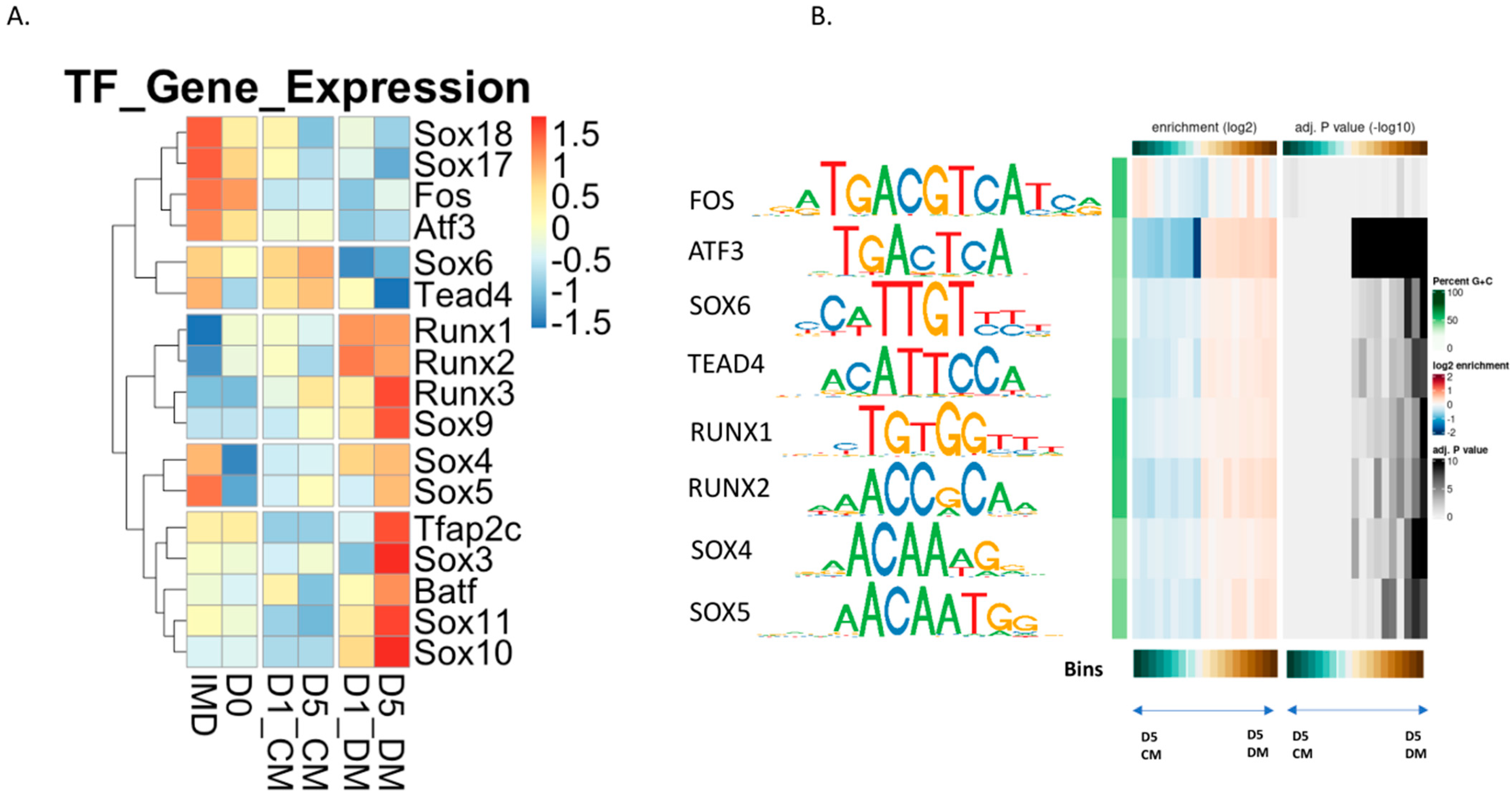
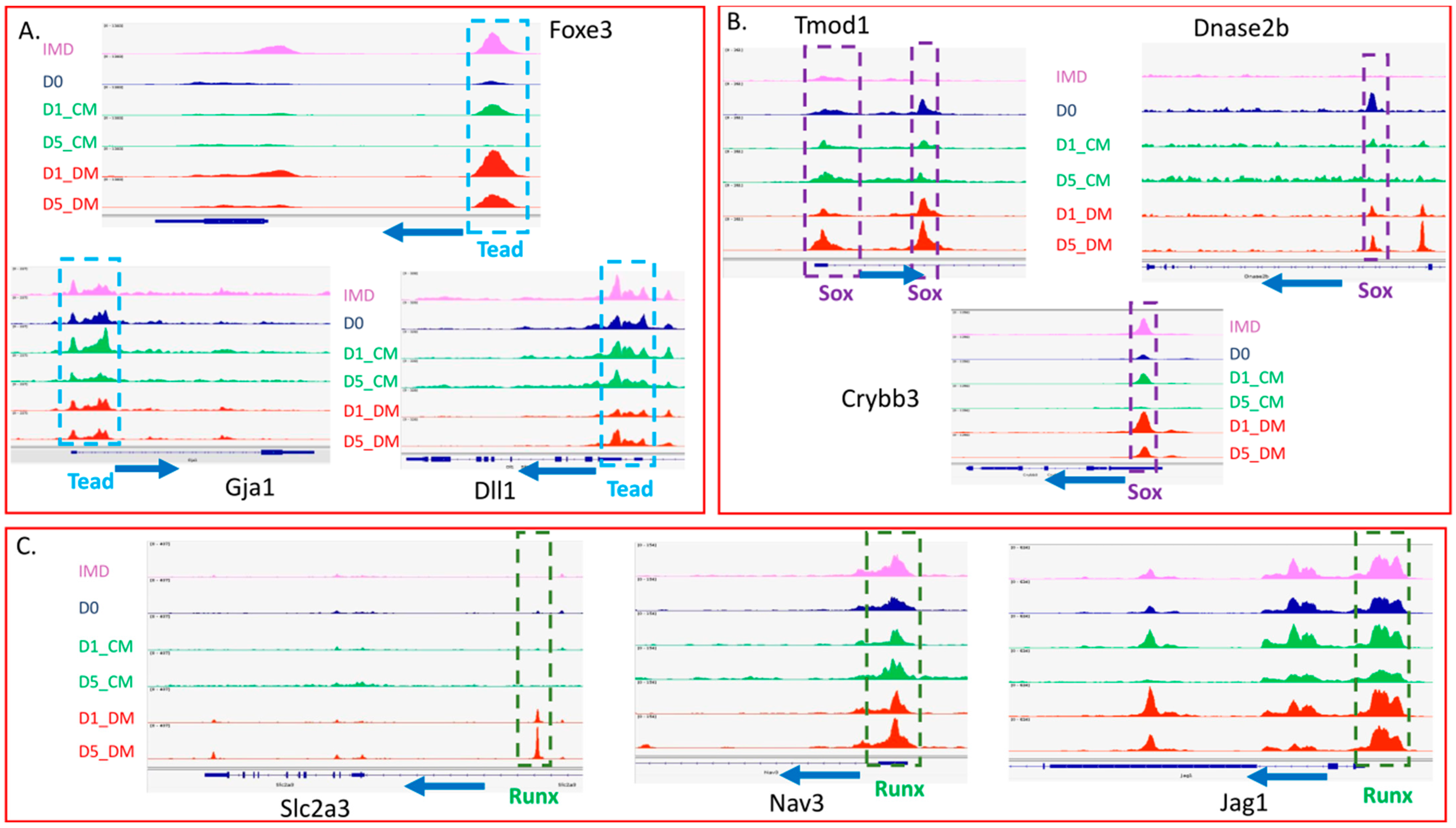
3.6. Evaluation of Transcription Factor Binding Motifs in DARs Induced in Lens Epithelial Explants by the Addition of Vitreous Humor
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Audette, D.S.; Scheiblin, D.A.; Duncan, M.K. The Molecular Mechanisms Underlying Lens Fiber Elongation. Exp. Eye Res. 2017, 156, 41–49. [Google Scholar] [CrossRef]
- Brennan, L.; Disatham, J.; Kantorow, M. Mechanisms of Organelle Elimination for Lens Development and Differentiation. Exp. Eye Res. 2021, 209, 108682. [Google Scholar] [CrossRef] [PubMed]
- Cvekl, A.; Eliscovich, C. Crystallin Gene Expression: Insights from Studies of Transcriptional Bursting. Exp. Eye Res. 2021, 207, 108564. [Google Scholar] [CrossRef] [PubMed]
- Coulombre, J.L.; Coulombre, A.J. Lens development: Fiber elongation and lens orientation. Science 1963, 142, 1489–1490. [Google Scholar] [CrossRef]
- Yamamoto, Y. Growth of Lens and Ocular Environment: Role of Neural Retina in the Growth of Mouse Lens as Revealed by an Implantation Experiment. Dev. Growth Differ. 1976, 18, 273–278. [Google Scholar] [CrossRef]
- Spemann, H. Uber Korrelationen in Der Entwicklung Des Auges. Verh. Anat. Ges. 1901, 15, 61–79. [Google Scholar]
- Henry, J.J.; Tsonis, P.A. Molecular and Cellular Aspects of Amphibian Lens Regeneration. Prog. Retin. Eye Res. 2010, 29, 543–555. [Google Scholar] [CrossRef]
- Maki, N.; Tsonis, P.A.; Agata, K. Changes in Global Histone Modifications during Dedifferentiation in Newt Lens Regeneration. Mol. Vis. 2010, 16, 1893–1897. [Google Scholar]
- Overbeek, P.A.; Chepelinsky, A.B.; Khillan, J.S.; Piatigorsky, J.; Westphal, H. Lens-Specific Expression and Developmental Regulation of the Bacterial Chloramphenicol Acetyltransferase Gene Driven by the Murine Alpha A-Crystallin Promoter in Transgenic Mice. Proc. Natl. Acad. Sci. USA 1985, 82, 7815–7819. [Google Scholar] [CrossRef] [PubMed]
- Lakso, M.; Sauer, B.; Mosinger, B., Jr.; Lee, E.J.; Manning, R.W.; Yu, S.H.; Mulder, K.L.; Westphal, H. Targeted Oncogene Activation by Site-Specific Recombination in Transgenic Mice. Proc. Natl. Acad. Sci. USA 1992, 89, 6232–6236. [Google Scholar] [CrossRef]
- Pan, H.; Griep, A.E. Altered Cell Cycle Regulation in the Lens of HPV-16 E6 or E7 Transgenic Mice: Implications for Tumor Suppressor Gene Function in Development. Genes Dev. 1994, 8, 1285–1299. [Google Scholar] [CrossRef]
- Fromm, L.; Shawlot, W.; Gunning, K.; Butel, J.S.; Overbeek, P.A. The Retinoblastoma Protein-Binding Region of Simian Virus 40 Large T Antigen Alters Cell Cycle Regulation in Lenses of Transgenic Mice. Mol. Cell. Biol. 1994, 14, 6743–6754. [Google Scholar] [CrossRef] [PubMed]
- Griep, A.E. Cell Cycle Regulation in the Developing Lens. Semin. Cell Dev. Biol. 2006, 17, 686–697. [Google Scholar] [CrossRef]
- Taiyab, A.; West-Mays, J. Lens Fibrosis: Understanding the Dynamics of Cell Adhesion Signaling in Lens Epithelial-Mesenchymal Transition. Front. Cell Dev. Biol. 2022, 10, 886053. [Google Scholar] [CrossRef]
- Wormstone, I.M.; Wormstone, Y.M.; Smith, A.J.O.; Eldred, J.A. Posterior Capsule Opacification: What’s in the Bag? Prog. Retin. Eye Res. 2021, 82, 100905. [Google Scholar] [CrossRef]
- Li, M.; Luan, F.; Zhao, Y.; Hao, H.; Zhou, Y.; Han, W.; Fu, X. Epithelial-Mesenchymal Transition: An Emerging Target in Tissue Fibrosis. Exp. Biol. Med. 2016, 241, 1–13. [Google Scholar] [CrossRef]
- Shirai, K.; Tanaka, S.-I.; Lovicu, F.J.; Saika, S. The Murine Lens: A Model to Investigate in Vivo Epithelial-Mesenchymal Transition. Dev. Dyn. 2018, 247, 340–345. [Google Scholar] [CrossRef] [PubMed]
- Philpott, G.W.; Coulombre, A.J. Lens development. II. The Differentiation of Embryonic Chick Lens Epithelial Cells In Vitro and In Vivo. Exp. Cell Res. 1965, 38, 635–644. [Google Scholar] [CrossRef] [PubMed]
- West-Mays, J.A.; Pino, G.; Lovicu, F.J. Development and Use of the Lens Epithelial Explant System to Study Lens Differentiation and Cataractogenesis. Prog. Retin. Eye Res. 2010, 29, 135–143. [Google Scholar] [CrossRef]
- Liu, J.; Hales, A.M.; Chamberlain, C.G.; McAvoy, J.W. Induction of Cataract-like Changes in Rat Lens Epithelial Explants by Transforming Growth Factor Beta. Investig. Ophthalmol. Vis. Sci. 1994, 35, 388–401. [Google Scholar]
- Hales, A.M.; Schulz, M.W.; Chamberlain, C.G.; McAvoy, J.W. TGF-Beta 1 Induces Lens Cells to Accumulate Alpha-Smooth Muscle Actin, a Marker for Subcapsular Cataracts. Curr. Eye Res. 1994, 13, 885–890. [Google Scholar] [CrossRef]
- Davidson, M.G.; Wormstone, M.; Morgan, D.; Malakof, R.; Allen, J.; McGahan, M.C. Ex Vivo Canine Lens Capsular Sac Explants. Arbeitsphysiologie 2000, 238, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Park, S.Y.; Joo, C.-K. Hepatocyte Growth Factor Induces Proliferation of Lens Epithelial Cells through Activation of ERK1/2 and JNK/SAPK. Investig. Ophthalmol. Vis. Sci. 2004, 45, 2696–2704. [Google Scholar] [CrossRef] [Green Version]
- Taliana, L.; Evans, M.D.M.; Ang, S.; McAvoy, J.W. Vitronectin Is Present in Epithelial Cells of the Intact Lens and Promotes Epithelial Mesenchymal Transition in Lens Epithelial Explants. Mol. Vis. 2006, 12, 1233–1242. [Google Scholar]
- Jun, J.H.; Sohn, W.-J.; Lee, Y.; Chang, S.D.; Kim, J.-Y. Experimental Lens Capsular Bag Model for Posterior Capsule Opacification. Cell Tissue Res. 2014, 357, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Santhoshkumar, P.; Reneker, L.W.; Sharma, K.K. Histone Deacetylase Inhibitors Trichostatin A and Vorinostat Inhibit TGFβ2-Induced Lens Epithelial-to-Mesenchymal Cell Transition. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4731–4740. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.T.; McAvoy, J.W. Onset of Fibre Differentiation in Cultured Rat Lens Epithelium under the Influence of Neural Retina-Conditioned Medium. Exp. Eye Res. 1984, 39, 83–94. [Google Scholar] [CrossRef]
- Schulz, M.W.; Chamberlain, C.G.; de Iongh, R.U.; McAvoy, J.W. Acidic and Basic FGF in Ocular Media and Lens: Implications for Lens Polarity and Growth Patterns. Development 1993, 118, 117–126. [Google Scholar] [CrossRef]
- Chamberlain, C.G.; McAvoy, J.W. Induction of Lens Fibre Differentiation by Acidic and Basic Fibroblast Growth Factor (FGF). Growth Factors 1989, 1, 125–134. [Google Scholar] [CrossRef]
- Wang, Q.; McAvoy, J.W.; Lovicu, F.J. Growth Factor Signaling in Vitreous Humor-Induced Lens Fiber Differentiation. Invest. Ophthalmol. Vis. Sci. 2010, 51, 3599–3610. [Google Scholar] [CrossRef]
- Huang, J.-X.; Feldmeier, M.; Shui, Y.-B.; Beebe, D.C. Evaluation of Fibroblast Growth Factor Signaling during Lens Fiber Cell Differentiation. Investig. Ophthalmol. Vis. Sci. 2003, 44, 680–690. [Google Scholar] [CrossRef]
- Padula, S.L.; Sidler, E.P.; Wagner, B.D.; Manz, C.J.; Lovicu, F.J.; Robinson, M.L. Lens Fiber Cell Differentiation Occurs Independently of Fibroblast Growth Factor Receptor Signaling in the Absence of Pten. Dev. Biol. 2020, 467, 1–13. [Google Scholar] [CrossRef]
- Murthy, K.R.; Goel, R.; Subbannayya, Y.; Jacob, H.K.; Murthy, P.R.; Manda, S.S.; Patil, A.H.; Sharma, R.; Sahasrabuddhe, N.A.; Parashar, A.; et al. Proteomic Analysis of Human Vitreous Humor. Clin. Proteom. 2014, 11, 29. [Google Scholar] [CrossRef]
- Beebe, D.C.; Silver, M.H.; Belcher, K.S.; Van Wyk, J.J.; Svoboda, M.E.; Zelenka, P.S. Lentropin, a Protein That Controls Lens Fiber Formation, Is Related Functionally and Immunologically to the Insulin-like Growth Factors. Proc. Natl. Acad. Sci. USA 1987, 84, 2327–2330. [Google Scholar] [CrossRef] [Green Version]
- Cassidy, L.; Barry, P.; Shaw, C.; Duffy, J.; Kennedy, S. Platelet Derived Growth Factor and Fibroblast Growth Factor Basic Levels in the Vitreous of Patients with Vitreoretinal Disorders. Br. J. Ophthalmol. 1998, 82, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Majima, K. Presence of Growth Factor in Human Vitreous. Ophthalmologica 1997, 211, 226–228. [Google Scholar] [CrossRef] [PubMed]
- Katsura, Y.; Okano, T.; Noritake, M.; Kosano, H.; Nishigori, H.; Kado, S.; Matsuoka, T. Hepatocyte Growth Factor in Vitreous Fluid of Patients with Proliferative Diabetic Retinopathy and Other Retinal Disorders. Diabetes Care 1998, 21, 1759–1763. [Google Scholar] [CrossRef]
- Shui, Y.-B.; Wang, X.; Hu, J.S.; Wang, S.-P.; Garcia, C.M.; Potts, J.D.; Sharma, Y.; Beebe, D.C. Vascular Endothelial Growth Factor Expression and Signaling in the Lens. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3911–3919. [Google Scholar] [CrossRef] [PubMed]
- Brooks, H.L., Jr.; Caballero, S., Jr.; Newell, C.K.; Steinmetz, R.L.; Watson, D.; Segal, M.S.; Harrison, J.K.; Scott, E.W.; Grant, M.B. Vitreous Levels of Vascular Endothelial Growth Factor and Stromal-Derived Factor 1 in Patients with Diabetic Retinopathy and Cystoid Macular Edema before and after Intraocular Injection of Triamcinolone. Arch. Ophthalmol. 2004, 122, 1801–1807. [Google Scholar] [CrossRef]
- Boswell, B.A.; Lein, P.J.; Musil, L.S. Cross-Talk between Fibroblast Growth Factor and Bone Morphogenetic Proteins Regulates Gap Junction-Mediated Intercellular Communication in Lens Cells. Mol. Biol. Cell 2008, 19, 2631–2641. [Google Scholar] [CrossRef]
- Lovicu, F.J.; McAvoy, J.W. FGF-Induced Lens Cell Proliferation and Differentiation Is Dependent on MAPK (ERK1/2) Signalling. Development 2001, 128, 5075–5084. [Google Scholar] [CrossRef] [PubMed]
- Boswell, B.A.; Musil, L.S. Synergistic Interaction between the Fibroblast Growth Factor and Bone Morphogenetic Protein Signaling Pathways in Lens Cells. Mol. Biol. Cell 2015, 26, 2561–2572. [Google Scholar] [CrossRef] [PubMed]
- Shu, D.Y.; Lovicu, F.J. Insights into Bone Morphogenetic Protein-(BMP-) Signaling in Ocular Lens Biology and Pathology. Cells 2021, 10, 2604. [Google Scholar] [CrossRef]
- Boswell, B.A.; Korol, A.; West-Mays, J.A.; Musil, L.S. Dual Function of TGFβ in Lens Epithelial Cell Fate: Implications for Secondary Cataract. Mol. Biol. Cell 2017, 28, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.V.; Kumar, P.K.R.; Sutharzan, S.; Tsonis, P.A.; Liang, C.; Robinson, M.L. Comparative Transcriptome Analysis of Epithelial and Fiber Cells in Newborn Mouse Lenses with RNA Sequencing. Mol. Vis. 2014, 20, 1491–1517. [Google Scholar]
- Zhao, Y.; Zheng, D.; Cvekl, A. A Comprehensive Spatial-Temporal Transcriptomic Analysis of Differentiating Nascent Mouse Lens Epithelial and Fiber Cells. Exp. Eye Res. 2018, 175, 56–72. [Google Scholar] [CrossRef]
- Chauss, D.; Basu, S.; Rajakaruna, S.; Ma, Z.; Gau, V.; Anastas, S.; Brennan, L.A.; Hejtmancik, J.F.; Menko, A.S.; Kantorow, M. Differentiation State-Specific Mitochondrial Dynamic Regulatory Networks Are Revealed by Global Transcriptional Analysis of the Developing Chicken Lens. G3 2014, 4, 1515–1527. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, D.; Cvekl, A. Profiling of Chromatin Accessibility and Identification of General Cis-Regulatory Mechanisms That Control Two Ocular Lens Differentiation Pathways. Epigenetics Chromatin 2019, 12, 27. [Google Scholar] [CrossRef]
- Disatham, J.; Chauss, D.; Gheyas, R.; Brennan, L.; Blanco, D.; Daley, L.; Menko, A.S.; Kantorow, M. Lens Differentiation Is Characterized by Stage-Specific Changes in Chromatin Accessibility Correlating with Differentiation State-Specific Gene Expression. Dev. Biol. 2019, 453, 86–104. [Google Scholar] [CrossRef]
- Farnsworth, D.R.; Posner, M.; Miller, A.C. Single Cell Transcriptomics of the Developing Zebrafish Lens and Identification of Putative Controllers of Lens Development. Exp. Eye Res. 2021, 206, 108535. [Google Scholar] [CrossRef]
- van Zyl, T.; Yan, W.; McAdams, A.M.; Monavarfeshani, A.; Hageman, G.S.; Sanes, J.R. Cell Atlas of the Human Ocular Anterior Segment: Tissue-Specific and Shared Cell Types. Proc. Natl. Acad. Sci. USA 2022, 119, e2200914119. [Google Scholar] [CrossRef] [PubMed]
- FastQC. A Quality Control Tool for High Throughput Sequence Data; Babraham Bioinformatics Web Site. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 November 2021).
- Babraham Bioinformatics-FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 7 July 2022).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize Analysis Results for Multiple Tools and Samples in a Single Report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet.journal 2011, 17, 10. [Google Scholar] [CrossRef]
- Babraham Bioinformatics-Trim Galore! Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 7 July 2022).
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Frankish, A.; Diekhans, M.; Jungreis, I.; Lagarde, J.; Loveland, J.E.; Mudge, J.M.; Sisu, C.; Wright, J.C.; Armstrong, J.; Barnes, I.; et al. GENCODE 2021. Nucleic Acids Res. 2021, 49, D916–D923. [Google Scholar] [CrossRef] [PubMed]
- Skovaka/Stringtie2. Available online: https://github.com/skovaka/stringtie2 (accessed on 7 July 2022).
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Han, J. K-Means clustering. In Encyclopedia of Machine Learning and Data Mining; Springer: Boston, MA, USA, 2017; pp. 695–697. ISBN 9781489976857. [Google Scholar]
- Hartigan, J.A.; Wong, M.A. Algorithm AS 136: A K-Means Clustering Algorithm. J. R. Stat. Soc. Ser. C. Appl. Stat. 1979, 28, 100. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Envirronment for Statistical Computing. Available online: https//www.R-project.org/ (accessed on 1 October 2021).
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G:Profiler: A Web Server for Functional Enrichment Analysis and Conversions of Gene Lists (2019 Update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.-F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1α-Responsive Genes Involved in Oxidative Phosphorylation Are Coordinately Downregulated in Human Diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene Set Enrichment Analysis: A Knowledge-Based Approach for Interpreting Genome-Wide Expression Profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SAGE Publications: Thousand Oaks, CA, USA, 2018; ISBN 9781544336473. [Google Scholar]
- Lenth, R.V. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 27 January 2023).
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-Based Analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-Performance Genomics Data Visualization and Exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, F.; Ryan, D.P.; Grüning, B.; Bhardwaj, V.; Kilpert, F.; Richter, A.S.; Heyne, S.; Dündar, F.; Manke, T. DeepTools2: A next Generation Web Server for Deep-Sequencing Data Analysis. Nucleic Acids Res. 2016, 44, W160-5. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.-G.; He, Q.-Y. ChIPseeker: An R/Bioconductor Package for ChIP Peak Annotation, Comparison and Visualization. Bioinformatics 2015, 31, 2382–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machlab, D.; Burger, L.; Soneson, C.; Rijli, F.M.; Schübeler, D.; Stadler, M.B. MonaLisa: An R/Bioconductor Package for Identifying Regulatory Motifs. Bioinformatics 2022, 38, 2624–2625. [Google Scholar] [CrossRef]
- Castro-Mondragon, J.A.; Riudavets-Puig, R.; Rauluseviciute, I.; Lemma, R.B.; Turchi, L.; Blanc-Mathieu, R.; Lucas, J.; Boddie, P.; Khan, A.; Manosalva Pérez, N.; et al. JASPAR 2022: The 9th Release of the Open-Access Database of Transcription Factor Binding Profiles. Nucleic Acids Res. 2022, 50, D165–D173. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, J.C. Venny; An Interactive Tool for Comparing Lists with Venn’s Diagrams. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 10 November 2022).
- Wang, X.; Garcia, C.M.; Shui, Y.-B.; Beebe, D.C. Expression and Regulation of Alpha-, Beta-, and Gamma-Crystallins in Mammalian Lens Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3608–3619. [Google Scholar] [CrossRef]
- Iyengar, L.; Wang, Q.; Rasko, J.E.J.; McAvoy, J.W.; Lovicu, F.J. Duration of ERK1/2 Phosphorylation Induced by FGF or Ocular Media Determines Lens Cell Fate. Differentiation 2007, 75, 662–668. [Google Scholar] [CrossRef]
- Hu, H.; Miao, Y.-R.; Jia, L.-H.; Yu, Q.-Y.; Zhang, Q.; Guo, A.-Y. AnimalTFDB 3.0: A Comprehensive Resource for Annotation and Prediction of Animal Transcription Factors. Nucleic Acids Res. 2019, 47, D33–D38. [Google Scholar] [CrossRef]
- Menko, A.S.; Klukas, K.A.; Johnson, R.G. Chicken Embryo Lens Cultures Mimic Differentiation in the Lens. Dev. Biol. 1984, 103, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Musil, L.S. Primary Cultures of Embryonic Chick Lens Cells as a Model System to Study Lens Gap Junctions and Fiber Cell Differentiation. J. Membr. Biol. 2012, 245, 357–368. [Google Scholar] [CrossRef]
- Chamberlain, C.G.; McAvoy, J.W. Evidence That Fibroblast Growth Factor Promotes Lens Fibre Differentiation. Curr. Eye Res. 1987, 6, 1165–1169. [Google Scholar] [CrossRef]
- McAvoy, J.W.; Chamberlain, C.G. Fibroblast Growth Factor (FGF) Induces Different Responses in Lens Epithelial Cells Depending on Its Concentration. Development 1989, 107, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.M.; Yu, K.; Zhao, H.; Ashery-Padan, R.; Ornitz, D.M.; Robinson, M.L.; Beebe, D.C. Signaling through FGF Receptor-2 Is Required for Lens Cell Survival and for Withdrawal from the Cell Cycle during Lens Fiber Cell Differentiation. Dev. Dyn. 2005, 233, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yang, T.; Madakashira, B.P.; Thiels, C.A.; Bechtle, C.A.; Garcia, C.M.; Zhang, H.; Yu, K.; Ornitz, D.M.; Beebe, D.C.; et al. Fibroblast Growth Factor Receptor Signaling Is Essential for Lens Fiber Cell Differentiation. Dev. Biol. 2008, 318, 276–288. [Google Scholar] [CrossRef]
- Garcia, C.M.; Huang, J.; Madakashira, B.P.; Liu, Y.; Rajagopal, R.; Dattilo, L.; Robinson, M.L.; Beebe, D.C. The Function of FGF Signaling in the Lens Placode. Dev. Biol. 2011, 351, 176–185. [Google Scholar] [CrossRef]
- Boswell, B.A.; Overbeek, P.A.; Musil, L.S. Essential Role of BMPs in FGF-Induced Secondary Lens Fiber Differentiation. Dev. Biol. 2008, 324, 202–212. [Google Scholar] [CrossRef]
- Belecky-Adams, T.L.; Adler, R.; Beebe, D.C. Bone Morphogenetic Protein Signaling and the Initiation of Lens Fiber Cell Differentiation. Development 2002, 129, 3795–3802. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Z.G.; Taiyab, A.; West-Mays, J.A. MMP9 Differentially Regulates Proteins Involved in Actin Polymerization and Cell Migration during TGF-β-Induced EMT in the Lens. Int. J. Mol. Sci. 2021, 22, 11988. [Google Scholar] [CrossRef]
- Symonds, J.G.; Lovicu, F.J.; Chamberlain, C.G. Posterior Capsule Opacification-like Changes in Rat Lens Explants Cultured with TGFbeta and FGF: Effects of Cell Coverage and Regional Differences. Exp. Eye Res. 2006, 82, 693–699. [Google Scholar] [CrossRef]
- Taiyab, A.; Holms, J.; West-Mays, J.A. β-Catenin/Smad3 Interaction Regulates Transforming Growth Factor-β-Induced Epithelial to Mesenchymal Transition in the Lens. Int. J. Mol. Sci. 2019, 20, 2078. [Google Scholar] [CrossRef]
- Nahomi, R.B.; Pantcheva, M.B.; Nagaraj, R.H. AB-Crystallin Is Essential for the TGF-Β2-Mediated Epithelial to Mesenchymal Transition of Lens Epithelial Cells. Biochem. J. 2016, 473, 1455–1469. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, Y.; Uchikawa, M.; Collignon, J.; Lovell-Badge, R.; Kondoh, H. Involvement of Sox1, 2 and 3 in the Early and Subsequent Molecular Events of Lens Induction. Development 1998, 125, 2521–2532. [Google Scholar] [CrossRef] [PubMed]
- Dvorakova, M.; Macova, I.; Bohuslavova, R.; Anderova, M.; Fritzsch, B.; Pavlinkova, G. Early Ear Neuronal Development, but Not Olfactory or Lens Development, Can Proceed without SOX2. Dev. Biol. 2020, 457, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, H.; Uchikawa, M.; Kamachi, Y. Interplay of Pax6 and SOX2 in Lens Development as a Paradigm of Genetic Switch Mechanisms for Cell Differentiation. Int. J. Dev. Biol. 2004, 48, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Nishiguchi, S.; Wood, H.; Kondoh, H.; Lovell-Badge, R.; Episkopou, V. Sox1 Directly Regulates the Gamma-Crystallin Genes and Is Essential for Lens Development in Mice. Genes Dev. 1998, 12, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Donner, A.L.; Episkopou, V.; Maas, R.L. Sox2 and Pou2f1 Interact to Control Lens and Olfactory Placode Development. Dev. Biol. 2007, 303, 784–799. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Pirity, M.K.; Wang, W.-L.; Wolf, L.; Chauhan, B.K.; Cveklova, K.; Tamm, E.R.; Ashery-Padan, R.; Metzger, D.; Nakai, A.; et al. Chromatin Remodeling Enzyme Brg1 Is Required for Mouse Lens Fiber Cell Terminal Differentiation and Its Denucleation. Epigenetics Chromatin 2010, 3, 21. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; van Deursen, J.; Hiebert, S.W.; Grosveld, G.; Downing, J.R. AML1, the Target of Multiple Chromosomal Translocations in Human Leukemia, Is Essential for Normal Fetal Liver Hematopoiesis. Cell 1996, 84, 321–330. [Google Scholar] [CrossRef]
- Wang, Q.; Stacy, T.; Binder, M.; Marin-Padilla, M.; Sharpe, A.H.; Speck, N.A. Disruption of the Cbfa2 Gene Causes Necrosis and Hemorrhaging in the Central Nervous System and Blocks Definitive Hematopoiesis. Proc. Natl. Acad. Sci. USA 1996, 93, 3444–3449. [Google Scholar] [CrossRef] [PubMed]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.H.; Inada, M.; et al. Targeted Disruption of Cbfa1 Results in a Complete Lack of Bone Formation Owing to Maturational Arrest of Osteoblasts. Cell 1997, 89, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Otto, F.; Thornell, A.P.; Crompton, T.; Denzel, A.; Gilmour, K.C.; Rosewell, I.R.; Stamp, G.W.; Beddington, R.S.; Mundlos, S.; Olsen, B.R.; et al. Cbfa1, a Candidate Gene for Cleidocranial Dysplasia Syndrome, Is Essential for Osteoblast Differentiation and Bone Development. Cell 1997, 89, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.-I.; Ozaki, S.; Shiga, T.; Ito, K.; Masuda, T.; Okado, N.; Iseda, T.; Kawaguchi, S.; Ogawa, M.; Bae, S.-C.; et al. Runx3 Controls the Axonal Projection of Proprioceptive Dorsal Root Ganglion Neurons. Nat. Neurosci. 2002, 5, 946–954. [Google Scholar] [CrossRef]
- Levanon, D.; Bettoun, D.; Harris-Cerruti, C.; Woolf, E.; Negreanu, V.; Eilam, R.; Bernstein, Y.; Goldenberg, D.; Xiao, C.; Fliegauf, M.; et al. The Runx3 Transcription Factor Regulates Development and Survival of TrkC Dorsal Root Ganglia Neurons. EMBO J. 2002, 21, 3454–3463. [Google Scholar] [CrossRef]
- Li, Q.L.; Ito, K.; Sakakura, C.; Fukamachi, H.; Inoue, K.I.; Chi, X.Z.; Lee, K.Y.; Nomura, S.; Lee, C.W.; Han, S.B.; et al. Causal Relationship between the Loss of RUNX3 Expression and Gastric Cancer. Cell 2002, 109, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Taniuchi, I.; Osato, M.; Egawa, T.; Sunshine, M.J.; Bae, S.C.; Komori, T.; Ito, Y.; Littman, D.R. Differential Requirements for Runx Proteins in CD4 Repression and Epigenetic Silencing during T Lymphocyte Development. Cell 2002, 111, 621–633. [Google Scholar] [CrossRef] [PubMed]
- Fainaru, O.; Shseyov, D.; Hantisteanu, S.; Groner, Y. Accelerated Chemokine Receptor 7-Mediated Dendritic Cell Migration in Runx3 Knockout Mice and the Spontaneous Development of Asthma-like Disease. Proc. Natl. Acad. Sci. USA 2005, 102, 10598–10603. [Google Scholar] [CrossRef] [PubMed]
- Voronov, D.; Gromova, A.; Liu, D.; Zoukhri, D.; Medvinsky, A.; Meech, R.; Makarenkova, H.P. Transcription Factors Runx1 to 3 Are Expressed in the Lacrimal Gland Epithelium and Are Involved in Regulation of Gland Morphogenesis and Regeneration. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3115–3125. [Google Scholar] [CrossRef] [PubMed]
- Gazon, H.; Barbeau, B.; Mesnard, J.-M.; Peloponese, J.-M., Jr. Hijacking of the AP-1 Signaling Pathway during Development of ATL. Front. Microbiol. 2017, 8, 2686. [Google Scholar] [CrossRef]
- Chen, M.; Liu, Y.; Yang, Y.; Qiu, Y.; Wang, Z.; Li, X.; Zhang, W. Emerging Roles of Activating Transcription Factor (ATF) Family Members in Tumourigenesis and Immunity: Implications in Cancer Immunotherapy. Genes Dis. 2022, 9, 981–999. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Tsujimura, T.; Takeda, K.; Sugihara, A.; Maekawa, A.; Terada, N.; Yoshida, N.; Akira, S. Targeted Disruption of ATF4 Discloses Its Essential Role in the Formation of Eye Lens Fibres. Genes Cells 1998, 3, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Fatma, N.; Singh, P.; Chhunchha, B.; Kubo, E.; Shinohara, T.; Bhargavan, B.; Singh, D.P. Deficiency of Prdx6 in Lens Epithelial Cells Induces ER Stress Response-Mediated Impaired Homeostasis and Apoptosis. Am. J. Physiol. Cell Physiol. 2011, 301, C954–C967. [Google Scholar] [CrossRef] [PubMed]
- Mulhern, M.L.; Madson, C.J.; Danford, A.; Ikesugi, K.; Kador, P.F.; Shinohara, T. The Unfolded Protein Response in Lens Epithelial Cells from Galactosemic Rat Lenses. Investig. Ophthalmol. Vis. Sci. 2006, 47, 3951–3959. [Google Scholar] [CrossRef]
- Zhou, Y.; Bennett, T.M.; Shiels, A. Lens ER-Stress Response during Cataract Development in Mip-Mutant Mice. Biochim. Biophys. Acta 2016, 1862, 1433–1442. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhou, S.; Gu, J.; Wang, Y.; Guo, M.; Liu, Y. Differences in Unfolded Protein Response Pathway Activation in the Lenses of Three Types of Cataracts. PLoS ONE 2015, 10, e0130705. [Google Scholar] [CrossRef]
- Berthoud, V.M.; Minogue, P.J.; Lambert, P.A.; Snabb, J.I.; Beyer, E.C. The Cataract-Linked Mutant Connexin50D47A Causes Endoplasmic Reticulum Stress in Mouse Lenses. J. Biol. Chem. 2016, 291, 17569–17578. [Google Scholar] [CrossRef]
- Lovicu, F.J.; Chamberlain, C.G.; McAvoy, J.W. Differential Effects of Aqueous and Vitreous on Fiber Differentiation and Extracellular Matrix Accumulation in Lens Epithelial Explants. Investig. Ophthalmol. Vis. Sci. 1995, 36, 1459–1469. [Google Scholar]
- Klok, E.j.; Lubsen, N.H.; Chamberlain, C.G.; McAvoy, J.W. Induction and Maintenance of Differentiation of Rat Lens Epithelium by FGF-2, Insulin and IGF-1. Exp. Eye Res. 1998, 67, 425–431. [Google Scholar] [CrossRef]
- McAvoy, J.W.; Chamberlain, C.G.; de Iongh, R.U.; Richardson, N.A.; Lovicu, F.J. The Role of Fibroblast Growth Factor in Eye Lens Development. Ann. N. Y. Acad. Sci. 1991, 638, 256–274. [Google Scholar] [CrossRef]
- Dawes, L.J.; Sugiyama, Y.; Tanedo, A.S.; Lovicu, F.J.; McAvoy, J.W. Wnt-Frizzled Signaling Is Part of an FGF-Induced Cascade That Promotes Lens Fiber Differentiation. Investig. Ophthalmol. Vis. Sci. 2013, 54, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Golestaneh, N.; Fan, J.; Fariss, R.N.; Lo, W.-K.; Zelenka, P.S.; Chepelinsky, A.B. Lens Major Intrinsic Protein (MIP)/Aquaporin 0 Expression in Rat Lens Epithelia Explants Requires Fibroblast Growth Factor-Induced ERK and JNK Signaling. J. Biol. Chem. 2004, 279, 31813–31822. [Google Scholar] [CrossRef] [PubMed]
- Saravanamuthu, S.S.; Gao, C.Y.; Zelenka, P.S. Notch Signaling Is Required for Lateral Induction of Jagged1 during FGF-Induced Lens Fiber Differentiation. Dev. Biol. 2009, 332, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Lovicu, F.J.; McAvoy, J.W. Structural Analysis of Lens Epithelial Explants Induced to Differentiate into Fibres by Fibroblast Growth Factor (FGF). Exp. Eye Res. 1989, 49, 479–494. [Google Scholar] [CrossRef]
- Wang, Q.; Stump, R.; McAvoy, J.W.; Lovicu, F.J. MAPK/ERK1/2 and PI3-Kinase Signalling Pathways Are Required for Vitreous-Induced Lens Fibre Cell Differentiation. Exp. Eye Res. 2009, 88, 293–306. [Google Scholar] [CrossRef] [PubMed]
- Madakashira, B.P.; Kobrinski, D.A.; Hancher, A.D.; Arneman, E.C.; Wagner, B.D.; Wang, F.; Shin, H.; Lovicu, F.J.; Reneker, L.W.; Robinson, M.L. Frs2α Enhances Fibroblast Growth Factor-Mediated Survival and Differentiation in Lens Development. Development 2012, 139, 4601–4612. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CA | CB | CC | CD | CE | CF | |
|---|---|---|---|---|---|---|
| C1 | 267 | 157 | 153 | 227 | 67 | 37 |
| C2 | 300 | 163 | 129 | 198 | 107 | 66 |
| C3 | 433 | 198 | 152 | 417 | 78 | 91 |
| C4 | 395 | 141 | 236 | 166 | 269 | 46 |
| C5 | 296 | 96 | 106 | 124 | 152 | 36 |
| C6 | 487 | 434 | 217 | 514 | 119 | 119 |
| ATAC-Seq CM_D1 Up | ATAC-Seq No Difference | ATAC-Seq DM_D1_Up | |
|---|---|---|---|
| RNA seq CM_1_up | 12 | 3382 | 193 |
| RNA seq no difference | 61 | 9180 | 405 |
| RNA seq DM_1_up | 26 | 4221 | 145 |
| ATAC-Seq CM_D5_Up | ATAC-Seq no Difference | ATAC-Seq DM_D5_Up | |
| RNA seq CM_D5_up | 636 | 2434 | 915 |
| RNA seq no difference | 1848 | 5881 | 2580 |
| RNA seq DM_D5_up | 593 | 2544 | 1131 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Upreti, A.; Padula, S.L.; Tangeman, J.A.; Wagner, B.D.; O’Connell, M.J.; Jaquish, T.J.; Palko, R.K.; Mantz, C.J.; Anand, D.; Lovicu, F.J.; et al. Lens Epithelial Explants Treated with Vitreous Humor Undergo Alterations in Chromatin Landscape with Concurrent Activation of Genes Associated with Fiber Cell Differentiation and Innate Immune Response. Cells 2023, 12, 501. https://doi.org/10.3390/cells12030501
Upreti A, Padula SL, Tangeman JA, Wagner BD, O’Connell MJ, Jaquish TJ, Palko RK, Mantz CJ, Anand D, Lovicu FJ, et al. Lens Epithelial Explants Treated with Vitreous Humor Undergo Alterations in Chromatin Landscape with Concurrent Activation of Genes Associated with Fiber Cell Differentiation and Innate Immune Response. Cells. 2023; 12(3):501. https://doi.org/10.3390/cells12030501
Chicago/Turabian StyleUpreti, Anil, Stephanie L. Padula, Jared A. Tangeman, Brad D. Wagner, Michael J. O’Connell, Tycho J. Jaquish, Raye K. Palko, Courtney J. Mantz, Deepti Anand, Frank J. Lovicu, and et al. 2023. "Lens Epithelial Explants Treated with Vitreous Humor Undergo Alterations in Chromatin Landscape with Concurrent Activation of Genes Associated with Fiber Cell Differentiation and Innate Immune Response" Cells 12, no. 3: 501. https://doi.org/10.3390/cells12030501
APA StyleUpreti, A., Padula, S. L., Tangeman, J. A., Wagner, B. D., O’Connell, M. J., Jaquish, T. J., Palko, R. K., Mantz, C. J., Anand, D., Lovicu, F. J., Lachke, S. A., & Robinson, M. L. (2023). Lens Epithelial Explants Treated with Vitreous Humor Undergo Alterations in Chromatin Landscape with Concurrent Activation of Genes Associated with Fiber Cell Differentiation and Innate Immune Response. Cells, 12(3), 501. https://doi.org/10.3390/cells12030501