



NFATc1 Regulation of Dexamethasone-Induced TGFB2 Expression Is Cell Cycle Dependent in Trabecular Meshwork Cells

 and
and
Abstract
:1. Introduction
2. Materials and Methods
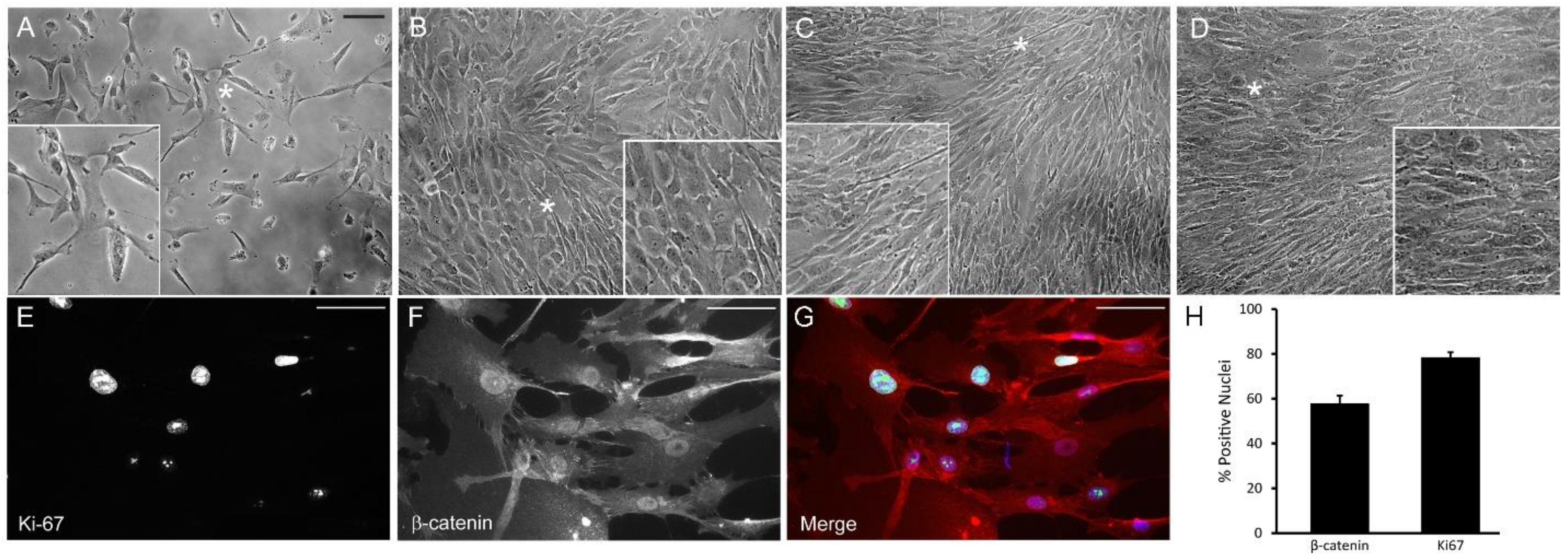
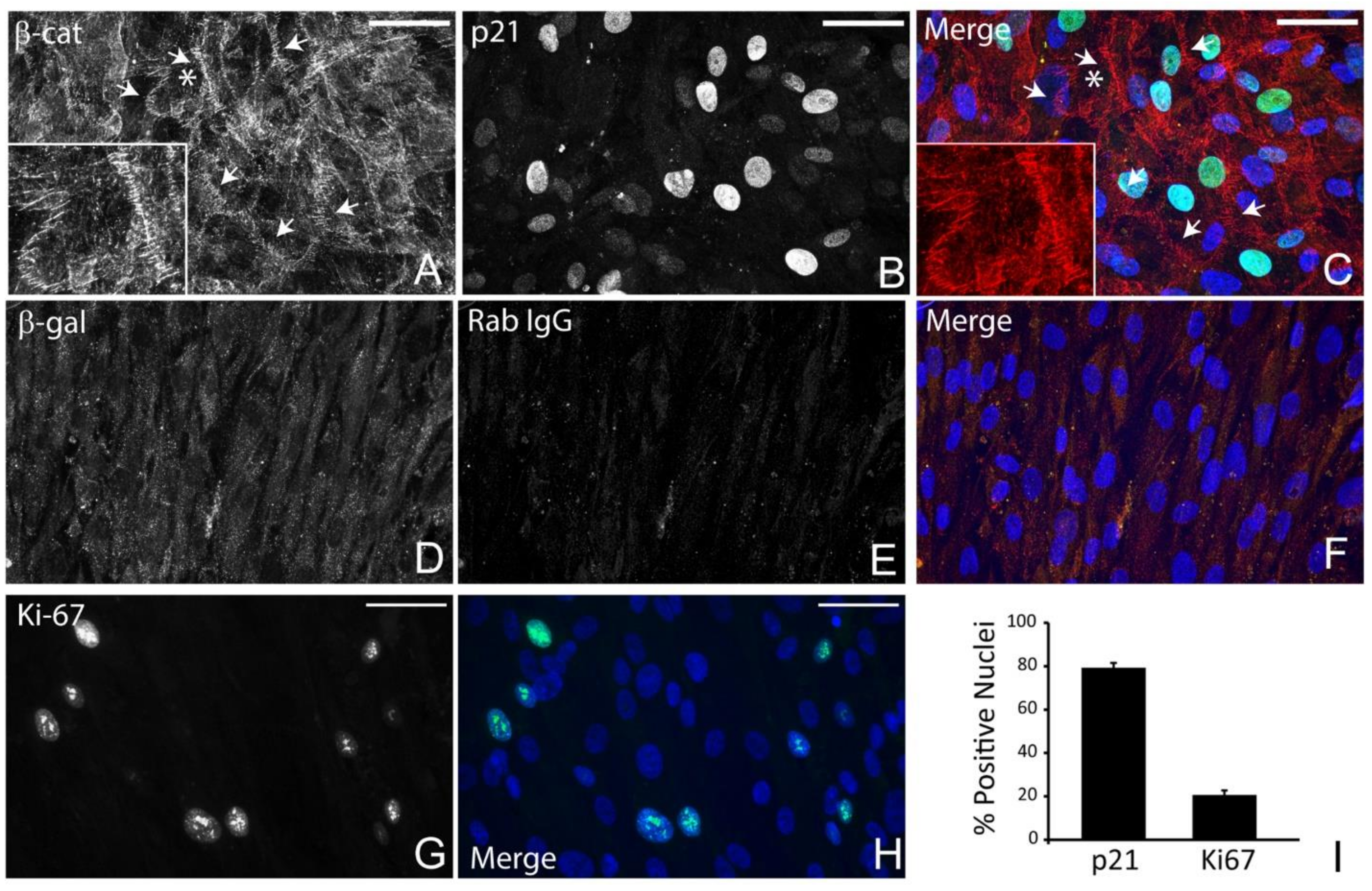
2.1. Cell Culture
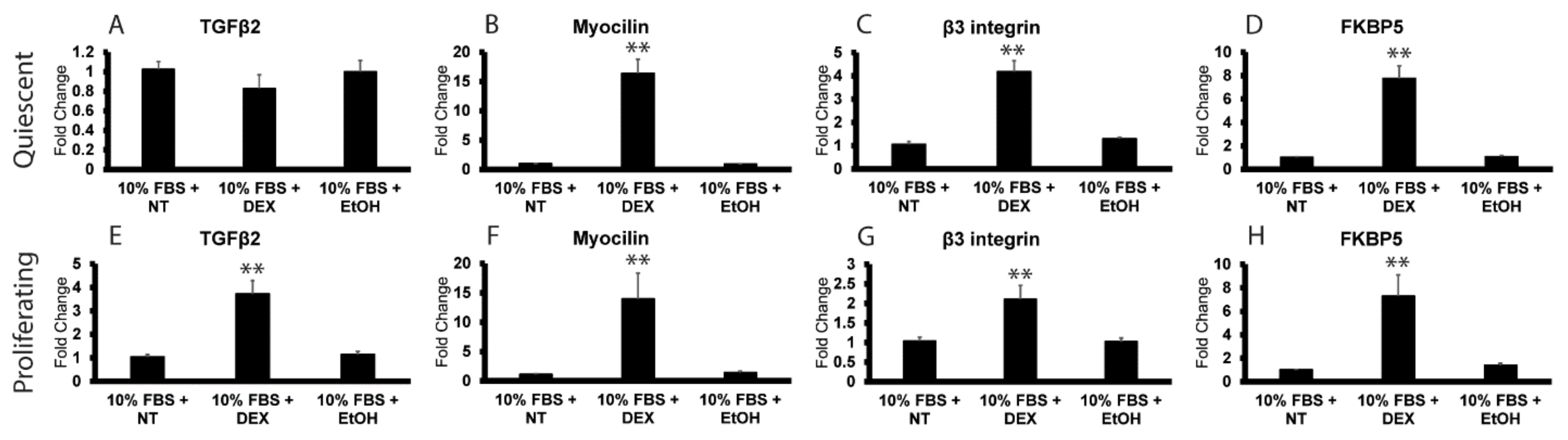
2.2. Induction of TGFβ2 with Dexamethasone and Effects of NFATc1 Inhibitors
2.3. RNA Isolation and RT-qPCR
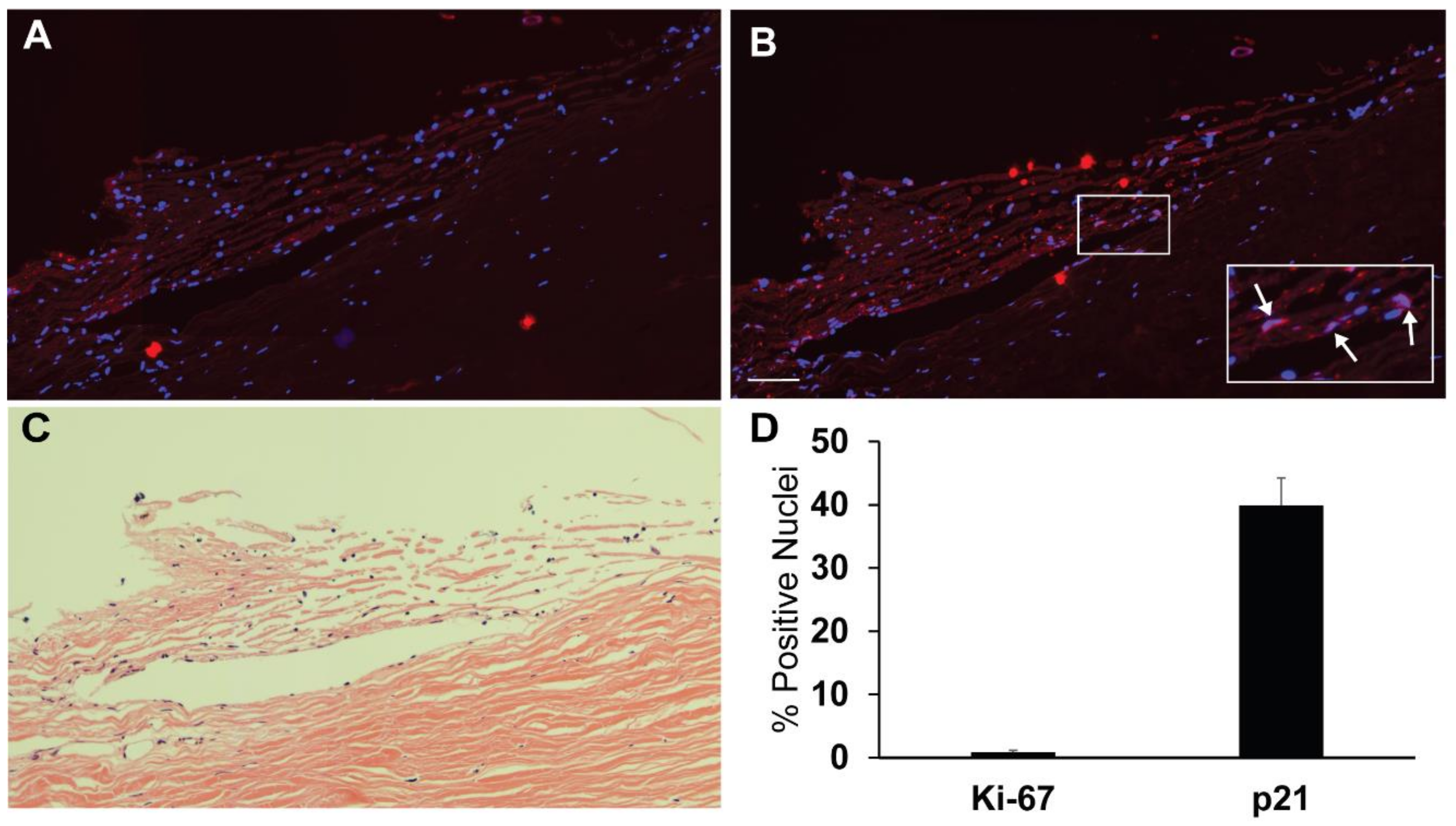
2.4. Immunolabeling and Quantification of Cell Cycle Protein-Positive Cells
2.5. Immunohistochemistry and Quantification of Cells in Tissue
2.6. Data Analysis
3. Results
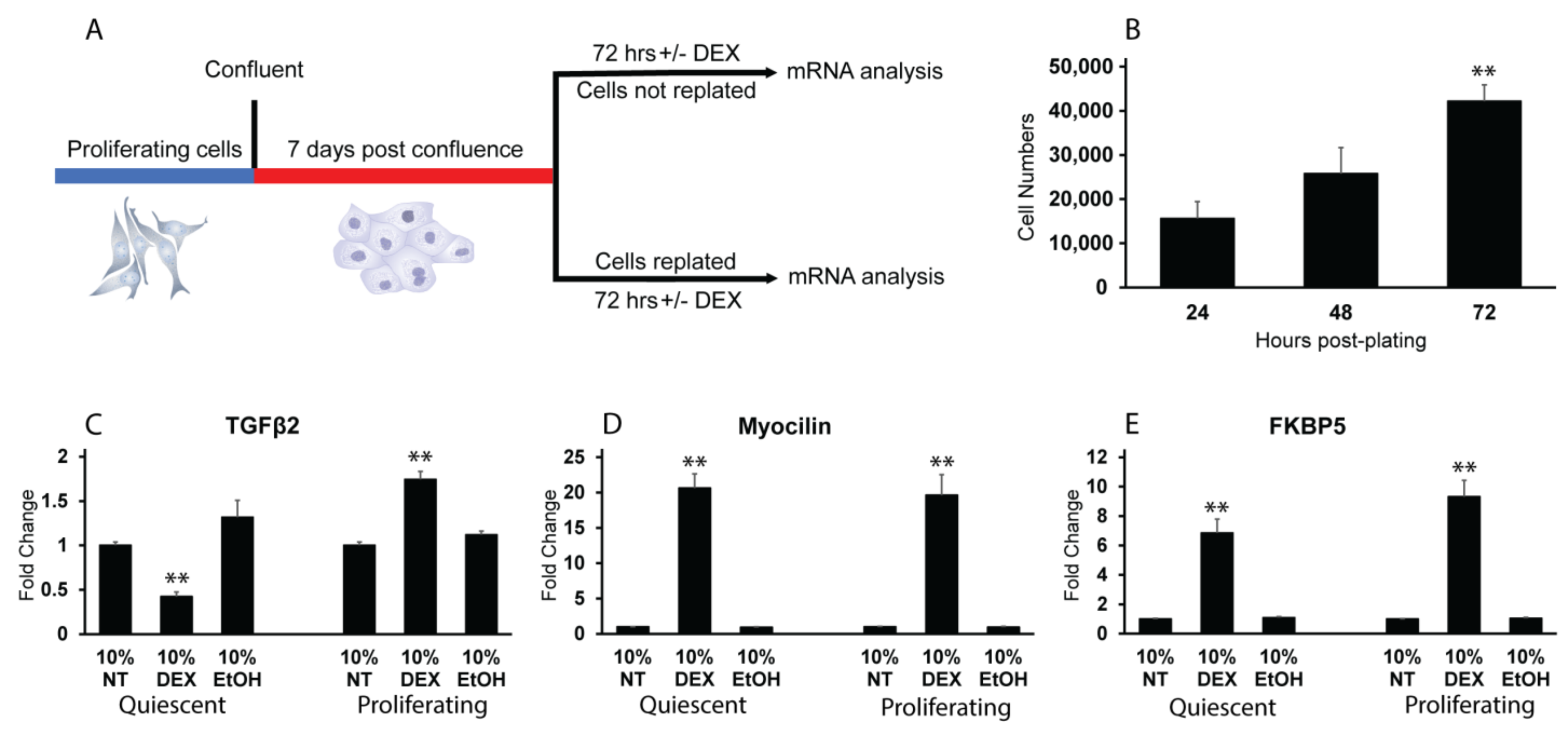
3.1. Detection of Quiescent and Proliferating Cells In Vivo and In Vitro
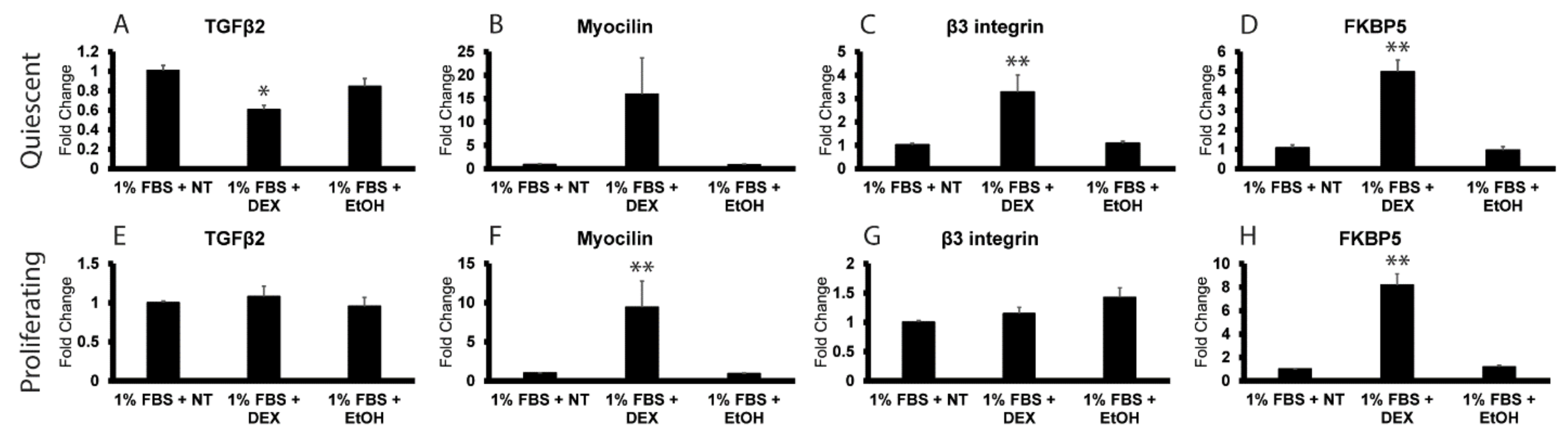
3.2. DEX Induces TGFβ2 mRNA in Proliferating HTM Cells but Not Quiescent Cells
3.3. Reactivation of the Cell Cycle in Quiescent HTM Cells Leads to an Upregulation in TGFβ2 mRNA by DEX
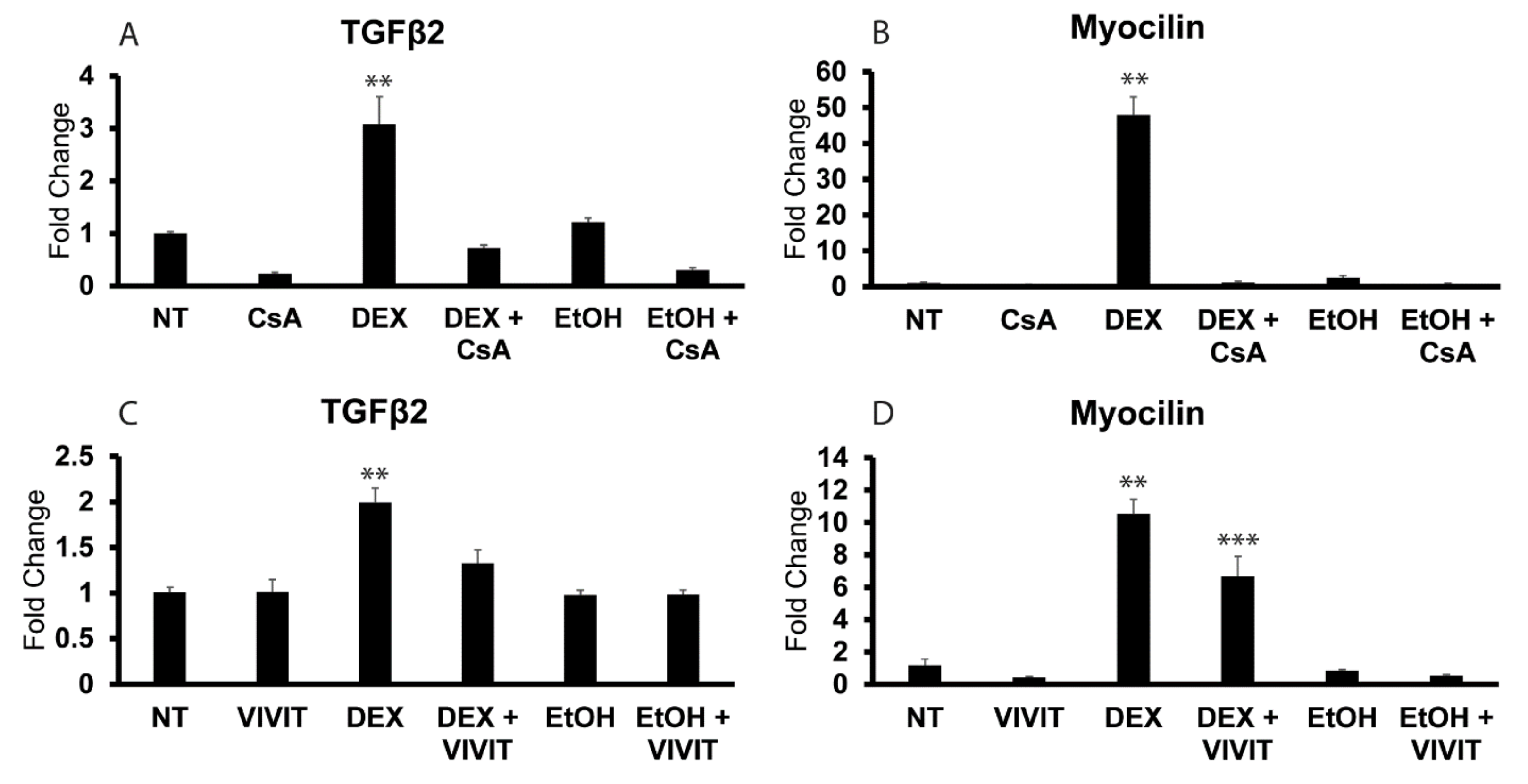
3.4. NFATc1 Regulates Expression of TGFβ2 mRNA in Proliferating HTM Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Grierson, I.; Hogg, P. The proliferative and migratory activities of trabecular meshwork cells. Prog. Retin. Eye Res. 1995, 15, 33–67. [Google Scholar] [CrossRef]
- Polansky, J.R.; Weinreb, R.N.; Baxter, J.D.; Alvarado, J. Human trabecular cells. I. Establishment in tissue culture and growth characteristics. Investig. Ophthalmol. Vis. Sci. 1979, 18, 1043–1049. [Google Scholar]
- Alvarado, J.A.; Wood, I.; Polansky, J. Human Trabecular cells II. Growth pattern and ultrastructural characteristics. Investig. Ophthalmol. Vis. Sci. 1982, 23, 464–478. [Google Scholar]
- Tominaga, K.; Suzuki, H.I. TGF-β Signaling in Cellular Senescence and Aging-Related Pathology. Int. J. Mol. Sci. 2019, 20, 5002. [Google Scholar] [CrossRef]
- Chen, K.-H.; Harris, D.L.; Joyce, N.C. TGF-β2 in aqueous humor suppresses S-Phase entry in cultured corneal endothelial cells. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2513–2519. [Google Scholar]
- Joyce, N.C.; Harris, D.L.; Mello, D.M. Mechanisms of mitotic inhibition in corneal endothelium: Contact inhibition and TGF-β2. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2152–2159. [Google Scholar]
- Tripathi, R.C.; Li, J.; Chan, W.F.; Tripathi, B.J. Aqueous humor in glaucomatous eyes contains an increased level of TGF-β2. Exp. Eye Res. 1994, 59, 723–727. [Google Scholar] [CrossRef]
- Inatani, M.; Tanihara, H.; Katsuta, M.; Honjo, M.; Kido, N.; Honda, Y. Transforming growth factor-beta 2 levels in aqueous humor of glaucomatous eyes. Graefes Arch. Clin. Exp. Ophthalmol. 2001, 239, 109–113. [Google Scholar] [CrossRef]
- Picht, G.; Welge-Lussen, U.; Grehn, F.; Lutjen-Drecoll, E. Transforming growth factor beta 2 levels in the aqueous humor in different types of glaucoma and the relation to filtering bleb development. Graefes Arch. Clin. Exp. Ophthalmol. 2001, 239, 199–207. [Google Scholar] [CrossRef]
- Gottanka, J.; Chan, D.; Eichhorn, M.; Lutjen-Drecoll, E.; Ethier, C.R. Effects of TGF-beta2 in Perfused Eyes. Investig. Ophthalmol. Vis. Sci. 2004, 45, 153–158. [Google Scholar] [CrossRef]
- Fleenor, D.L.; Shepard, A.R.; Hellberg, P.E.; Jacobson, N.; Pang, I.-H.; Clark, A.F. TGFbeta2-induced changes in trabecular meshwork: Implications for intraocular pressure. Investig. Ophthalmol. Vis. Sci. 2006, 47, 226–234. [Google Scholar] [CrossRef]
- Bhattacharya, S.K.; Gabelt, B.T.; Ruiz, J.; Picciani, R.; Kaufman, P.L. Cochlin expression in anterior organ culture models after TGF-β2 treatment. Investig. Ophthalmol. Vis. Sci. 2009, 50, 551–559. [Google Scholar] [CrossRef]
- Shepard, A.R.; Millar, J.C.; Pang, I.-H.; Jacobson, N.; Wang, W.H.; Clark, A.F. Adenoviral gene transfer of active human transforming growth factor-β2 elevates intraocular pressure and reduces outflow facility in rodent eyes. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2067–2076. [Google Scholar] [CrossRef]
- Welge-Lussen, U.; May, C.A.; Lutjen-Drecoll, E. Induction of tissue transglutaminase in the trabecular meshwork by TGF-β1 and TGF-β2. Investig. Ophthalmol. Vis. Sci. 2000, 2000, 2229–2238. [Google Scholar]
- Last, J.A.; Pan, T.; Ding, Y.; Reilly, C.M.; Keller, K.; Acott, T.S.; Fautsch, M.P.; Murphy, C.J.; Russell, P. Elastic modulus determination of normal and glaucomatous human trabecular meshwork. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2147–2152. [Google Scholar] [CrossRef] [Green Version]
- Camras, L.J.; Stamer, W.D.; Epstein, D.L.; Gonzalez, P.; Yuan, F. Circumferential Tensile Stiffness of Glaucomatous Trabecular Meshwork. Investig. Ophthalmol. Vis. Sci. 2014, 55, 814–823. [Google Scholar] [CrossRef]
- Yu, A.L.; Birke, K.; Moriniere, J.; Welge-Lussen, U. TGF-β2 Induces Senescence-Associated Changes in Human Trabecular Meshwork Cells. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5718–5723. [Google Scholar] [CrossRef]
- Fuchshofer, R.; Tamm, E.R. The role of TGF-β in the pathogenesis of primary open-angle glaucoma. Cell Tissue Res. 2012, 347, 279–290. [Google Scholar] [CrossRef]
- Glick, A.B.; Flanders, K.C.; Danielpour, D.; Yuspa, S.H.; Sporn, M.B. Retinoic acid induces transforming growth factor-beta 2 in cultured keratinocytes and mouse epidermis. Cell Regul. 1989, 1, 87–97. [Google Scholar] [CrossRef]
- Ghatpande, S.K.; Zhou, H.R.; Cakstina, I.; Carlson, C.; Rondini, E.A.; Romeih, M.; Zile, M.H. Transforming growth factor beta2 is negatively regulated by endogenous retinoic acid during early heart morphogenesis. Dev. Growth Differ. 2010, 52, 433–455. [Google Scholar] [CrossRef]
- Pemmari, A.; Leppanen, T.; Hamalanen, M.; Moilanen, T.; Vuoltennaho, K.; Moilanen, E. Widespread regulation of gene expression by glucocorticoids in chondrocytes from patients with osteoarthritis as determined by RNA-Seq. Arthritis Res. Ther. 2020, 22, 271. [Google Scholar] [CrossRef] [PubMed]
- Kasetti, R.B.; Maddineni, P.; Patel, P.; Searby, C.; Sheffield, V.C.; Zode, G.S. Transforming growth factor β2 (TGFβ2) signaling plays a key role in glucocorticoid-induced ocular hypertension. J. Biol. Chem. 2018, 293, 9854–9868. [Google Scholar] [CrossRef] [PubMed]
- Filla, M.S.; Meyers, K.A.; Faralli, J.A.; Peters, D.M. Overexpression and activation of avb3 integrin differentially affects TGFβ2 signaling in human trabecular meshwork cells. Cells 2021, 10, 1923. [Google Scholar] [CrossRef] [PubMed]
- Faralli, J.A.; Gagen, D.; Filla, M.S.; Crotti, T.N.; Peters, D.M. Dexamethasone increases αvβ3 integrin expression and affinity through a calcineurin/NFAT pathway. BBA Mol. Cell Res. 2013, 1833, 3306–3313. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-U.; Kim, L.-K.; Choi, J.-M. Revisiting the Concept of Targeting NFAT to Control T Cell Immunity and Autoimmune Diseases. Front. Immunol. 2018, 9, 2747. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Singh, S.K.; Konig, A.; Reutlinger, K.; Nye, M.D.; Adhikary, T.; Ellers, M.; Gress, T.M.; Fernandez-Zapico, M.E.; Ellenrieder, V. Sequential Activation of NFAT and c-Myc Transcription Factors Mediates the TGF-β Switch from a Suppressor to a Promoter of Cancer Cell Proliferation. J. Biol. Chem. 2010, 285, 27241–27250. [Google Scholar] [CrossRef]
- Tong, D.C.; Buck, S.M.; Roberts, B.R.; Klein, J.D.; Tumlin, J.A. Calcineurin phosphatase activity: Activation by glucocorticoids and role of intracellular calcium. Transplantation 2004, 77, 259–267. [Google Scholar] [CrossRef]
- Panaettieri, R.A.; Schaafsma, D.; Amrani, Y.; Koziol-White, C.; Ostrom, R.S. Non-genomic Effects of Glucocorticoids: An Updated View. Trends Pharmacol. Sci. 2019, 40, 38–49. [Google Scholar] [CrossRef]
- Grierson, I.; Lee, W.R. The fine structure of the trabecular meshwork at graded levels of intraocular pressure. (1) Pressure effects within the near-physiological range (8–30 mmHg). Exp. Eye Res. 1975, 20, 505–521. [Google Scholar] [CrossRef]
- Grierson, I.; Lee, W.R. Light microscopic quantitation of the endothelial vacuoles in Schlemm’s canal. Am. J. Ophthalmol. 1977, 84, 234–246. [Google Scholar] [CrossRef]
- Filla, M.; Woods, A.; Kaufman, P.L.; Peters, D.M. β1 and β3 integrins cooperate to induce syndecan-4-containing cross-linked actin networks in human trabecular meshwork cells. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1956–1967. [Google Scholar] [CrossRef]
- Filla, M.S.; Lui, X.; Nguyen, T.D.; Polansky, J.R.; Brandt, B.R.; Kaufman, P.L.; Peters, D.M. In vitro localization of TIGR/MYOC in trabecular meshwork extracellular matrix and binding to fibronectin. Investig. Ophthalmol. Vis. Sci. 2002, 43, 151–161. [Google Scholar]
- Keller, K.E.; Bhattacharya, S.K.; Borrás, T.; Brunner, T.M.; Chansangpetch, S.; Clark, A.F.; Dismuke, W.M.; Du, Y.; Elliott, M.H.; Ethier, C.R.; et al. Consensus recommendations for trabecular meshwork cell isolation, characterization and culture. Exp. Eye Res. 2018, 171, 164–173. [Google Scholar] [CrossRef]
- Clark, R.; Nosie, A.; Walker, T.M.; Faralli, J.A.; Filla, M.S.; Barrett-Wilt, G.; Peters, D.M. Comparative genomic and proteomic analysis of cytoskeletal changes in dexamethasone-treated trabecular meshwork cells. Mol. Cell. Proteom. 2013, 12, 194–206. [Google Scholar] [CrossRef]
- Miller, I.; Min, M.; Yang, C.; Tian, C.; Gookin, S.; Carter, D.; Spencer, S.L. Ki67 is a Graded Rather than a Binary Marker of Proliferation versus Quiescence. Cell Rep. 2018, 24, 1105–1112. [Google Scholar] [CrossRef]
- Harper, J.W.; Adami, G.R.; Wei, N.; Keyomarsi, K.; Elledge, S.J. The p21 Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell 1993, 75, 805–816. [Google Scholar] [CrossRef]
- Abercrombie, M. Contact inhibition in tissue culture. Vitr. Cell. Dev. Biol.–Plant 1970, 6, 128–142. [Google Scholar] [CrossRef]
- Polansky, J.R.; Wood, I.S.; Maglio, M.T.; Alvarado, J.A. Trabecular meshwork cell culture in glaucoma research: Evaluation of biological activity and structural properties of human trabecular cells in vitro. Ophthalmology 1984, 91, 580–595. [Google Scholar] [CrossRef]
- Olmeda, D.; Castel, S.; Vilaro, S.; Cano, A. Beta-catenin regulation during the cell cycle: Implications in G2/M and apoptosis. Mol. Biol. Cell 2003, 14, 2844–2860. [Google Scholar] [CrossRef]
- Nehme, A.; Lobenhofer, G.J.; Stamer, W.D.; Edelman, J.L. Glucocorticoids with different chemical structures but similar glucocorticoid receptor potency regulate subsets of common and unique genes in human trabecular meshwork cells. BMC Med. Genom. 2009, 2, 58–91. [Google Scholar] [CrossRef]
- Faralli, J.A.; Clark, R.W.; Filla, M.S.; Peters, D.M. NFATc1 activity regulates the expression of myocilin induced by dexamethasone. Exp. Eye Res. 2015, 130, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Coller, H.A.; Sang, L.; Roberts, J.M. A new description of cellular quiescence. PLoS Biol. 2006, 4, e83. [Google Scholar] [CrossRef] [PubMed]
- Gooch, J.L.; Gorin, Y.; Zhang, B.-X.; Abbound, H.E. Involvement of Calcineurin in Transforming Growth Factor-β-mediated Regulation of Extracellular Matrix Accumulation. J. Biol. Chem. 2004, 279, 15561–15570. [Google Scholar] [CrossRef] [PubMed]
- Cobbs, S.L.; Gooch, J.L. NFATc is required for TGFbeta-mediated transcriptional regulation of fibronectin. Biochem. Biophys. Res. Commun. 2007, 362, 288–294. [Google Scholar] [CrossRef]
- Stallaert, W.; Taylor, R.R.; Kedziora, K.M.; Taylor, C.D.; Sobon, H.K.; Young, C.L.; Limas, J.C.; Holloway, J.V.; Johnson, M.S.; Cook, J.G.; et al. The molecular architecture of cell cycle arrest. Mol. Syst. Biol. 2022, 18, e11087. [Google Scholar] [CrossRef]
- Roth, G.S.; Adelman, R.C. Age related changes in hormone binding by target cells and tissues; Possible role in altered adaptive responsiveness. Exp. Gerontol. 1975, 10, 1–11. [Google Scholar] [CrossRef]
- Brown, J.A.; Yonekubo, Y.; Hanson, N.; Sastre-Perona, A.; Basin, A.; Rytlewski, J.A.; Dolgalev, I.; Meehan, S.; Tsirigos, A.; Beronja, S.; et al. TGF-β-Induced Quiescence Mediates Chemoresistance of Tumor-Propagating Cells in Squamous Cell Carcinoma. Cell Stem Cell 2017, 21, 650–664. [Google Scholar] [CrossRef]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-β and the TGF-β Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef]
- Sureshbabu, A.; Muhsin, S.A.; Choi, M.E. TGF-β signaling in the kidney: Profibrotic and protective effects. Am. J. Physiol. Renal. Physiol. 2016, 310, F596–F606. [Google Scholar] [CrossRef]
- Fujimaki, K.; Yao, G. Cell dormancy plasticity: Quiescence deepens into senescence through a dimmer switch. Physiol. Genom. 2020, 52, 558–562. [Google Scholar] [CrossRef]
- Rathi, S.; Danford, I.; Gudiseva, H.V.; Verkuil, L.; Pistilli, M.; Vishwakarma, S.; Kaur, I.; Dave, T.V.; O′Brien, J.M.; Chavali, V.R.M. Molecular genetics and functional analysis implicate CDKN2BAS1-CDKN2B involvement in POAG pathogenesis. Cells 2020, 9, 1934. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, S.; Li, K.; Tan, K.; Liang, K.; Wang, J.; Shen, J.; Zou, W.; Hu, L.; Cai, D.; et al. mTORC1 inhibits NF-κB/NFATc1 signaling and prevents osteoclast precursor differentiation, in vitro and in mice. J. Bone Miner. Res. 2017, 32, 1829–1840. [Google Scholar] [CrossRef]
- Sharma, S.; Findlay, G.M.; Bandukwala, H.S.; Oberdoerffer, S.; Baust, B.; Li, Z.; Schmidt, V.; Hogan, P.G.; Sacks, D.B.; Rao, A. Dephosphorylation of the nuclear factor of activated T cells (NFAT) transcription factor is regulated by an RNA-protein scaffold complex. Proc. Natl. Acad. Sci. USA 2011, 108, 11381–11386. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Reverse Sequence | Forward Sequence |
|---|---|---|
| HPRT1 | GGTCCTTTTCACCAGCAAGCT | TGACACTGGCAAAACAATGCA |
| SDHA | CACCACTGCATCAAATTCATG | TGGGAACAAGAGGGCATCTG |
| ITGB3 | TTCTTCGAATCATCTGGCC | GTGACCTGAAGGAGAATCTGC |
| TGFB2 | CCTCGGGCTCAGGATAGTCT | CAGCACACTCGATATGGACCA |
| MYOC | CTCAGCGTGAGAGGCTCTCC | GCCCATCTGGCTATCTCAGG |
| FKBP5 | CCCTCTCCTTTCCGTTTGGTT | CTCCCTAAAATTCCCTCGAATGC |
| Target | Monoclonal Antibody | Host | Company | Product # | Concentration (μg/mL) |
|---|---|---|---|---|---|
| Ki-67 | SP6 | Rb | Abcam | ab16667 | 4 (cells) 5 (sections) |
| β-catenin | 15B8 | Ms | Millipore-Sigma | C7207 | 1 |
| P21 | R.229.6 | Rb | Invitrogen | MA5-14949 | 1 (cells) 5 (sections) |
| β-galactosidase | GAL-13 | Ms | Millipore-Sigma | G8021 | 1 |
| KLH-conjugated small molecule (nonimmune) | [EPR25A] | Rb | Abcam | ab172730 | 1 (cells) 4 (cells) 5 (sections) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filla, M.S.; Faralli, J.A.; Dunn, C.R.; Khan, H.; Peters, D.M. NFATc1 Regulation of Dexamethasone-Induced TGFB2 Expression Is Cell Cycle Dependent in Trabecular Meshwork Cells. Cells 2023, 12, 504. https://doi.org/10.3390/cells12030504
Filla MS, Faralli JA, Dunn CR, Khan H, Peters DM. NFATc1 Regulation of Dexamethasone-Induced TGFB2 Expression Is Cell Cycle Dependent in Trabecular Meshwork Cells. Cells. 2023; 12(3):504. https://doi.org/10.3390/cells12030504
Chicago/Turabian StyleFilla, Mark S., Jennifer A. Faralli, Caleigh R. Dunn, Haania Khan, and Donna M. Peters. 2023. "NFATc1 Regulation of Dexamethasone-Induced TGFB2 Expression Is Cell Cycle Dependent in Trabecular Meshwork Cells" Cells 12, no. 3: 504. https://doi.org/10.3390/cells12030504
APA StyleFilla, M. S., Faralli, J. A., Dunn, C. R., Khan, H., & Peters, D. M. (2023). NFATc1 Regulation of Dexamethasone-Induced TGFB2 Expression Is Cell Cycle Dependent in Trabecular Meshwork Cells. Cells, 12(3), 504. https://doi.org/10.3390/cells12030504