
Retinoic Acid and POU Genes in Developing Amphioxus: A Focus on Neural Development

 ,
, 
 ,
,  ,
,  and
and 
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Vertebrates | Cephalochordates |
|---|---|---|
| I | Pou1f1 (Pit1; Ghf1) | Pit-1 (AmphiPOU1F1) [26,27,28] |
| II | Pou2f1 (Oct1; Otf1) | |
| Pou2f2 (Oct2; Otf2) | Pou2 [18] | |
| Pou2f3 (Oct11; Skn1a; Pla1; Epoc1) | ||
| III | Pou3f1 (Oct6; Otf6; Scip; Tst1; Xlpou1) | |
| Pou3f2 (Oct7; Otf7; Brn2; Xlpou3) | Brn1/2/4 (Pou3f) [29,30] | |
| Pou3f3 (Oct8; Otf8; Brn1) | Pou3l (Pou3fl) [18,30] | |
| Pou3f4 (Oct9; Otf9; Brn4; Dfn3; Xlpou2) | ||
| IV | Pou4f1 (Brn3a; Rdc-1; Oct-T1) | |
| Pou4f2 (Brn3b; Brn-3.2) | Pou4 (AmphiPOU-IV) [31] | |
| Pou4f3 (Brn3c; Brn-3.1; Dfna15) | ||
| V | Pou5f1 (Oct3; Oct4; Otf3; Otf4) | None [18] |
| Pou5f2 (Sprm1; Pou2/V; Pou5f3) | ||
| VI | Pou6f1 (Brn5; Mpou; Tcfb1)Pou6f2 (Emb; Rpf1) | Pou6 [18] |
2. Materials and Methods
2.1. Spawning, Embryo Collection, and Pharmacological Treatments
2.2. Gene Cloning and RNA Probe Synthesis
2.3. Embryo Fixation, Whole Mount In Situ Hybridization, Hybridization Chain Reaction, and Imaging
2.4. Statistical Analyses
3. Results
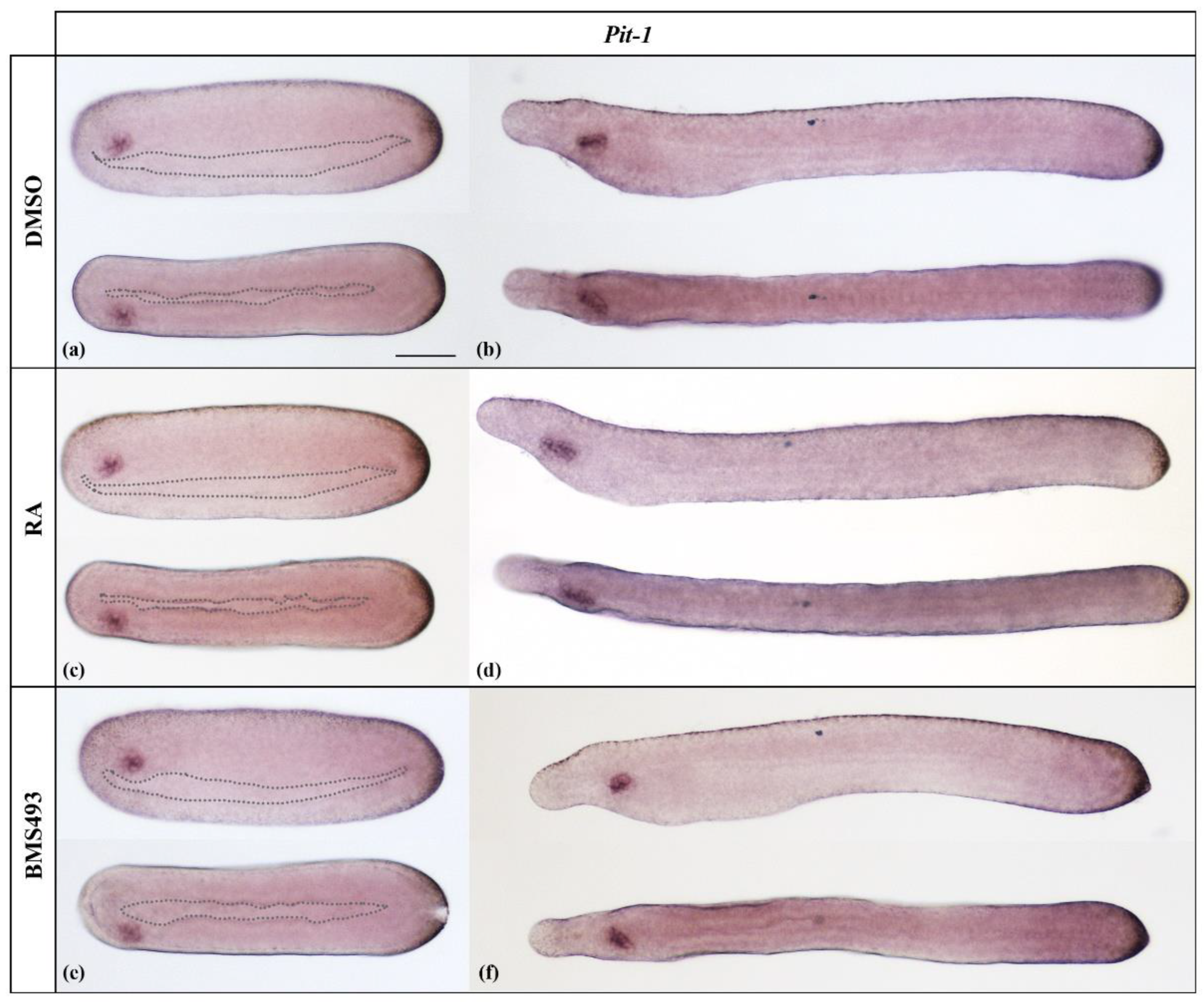
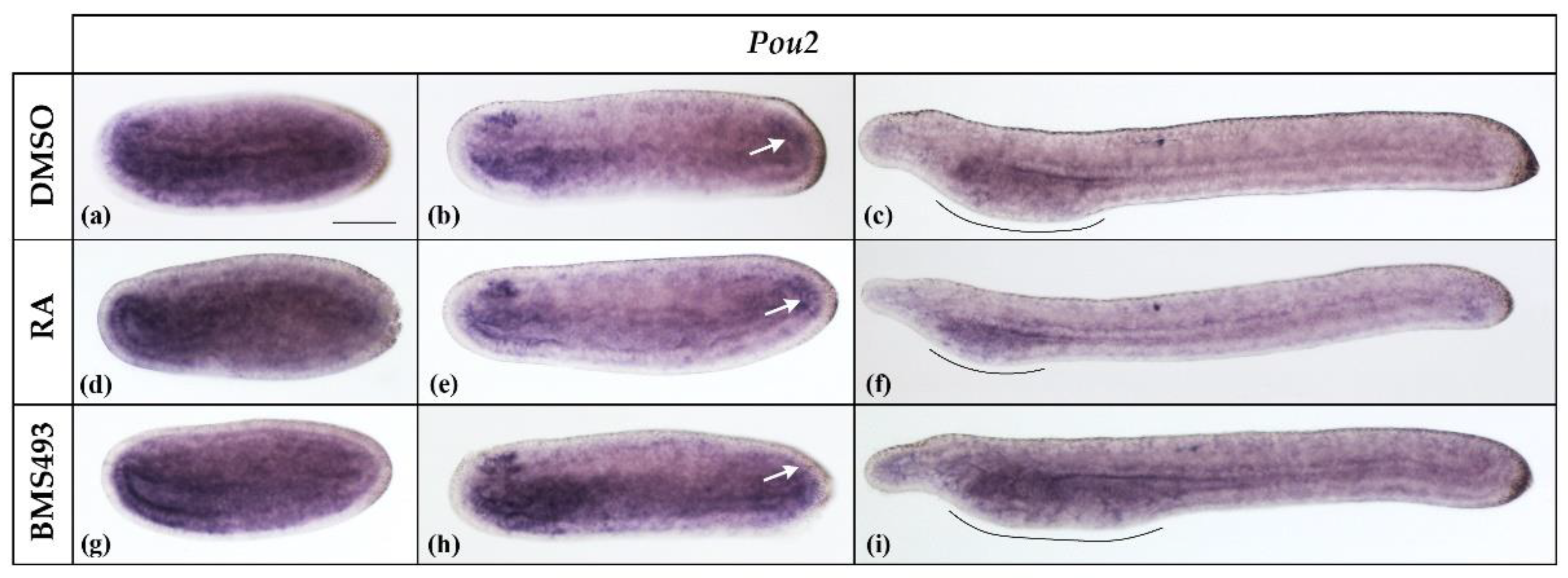
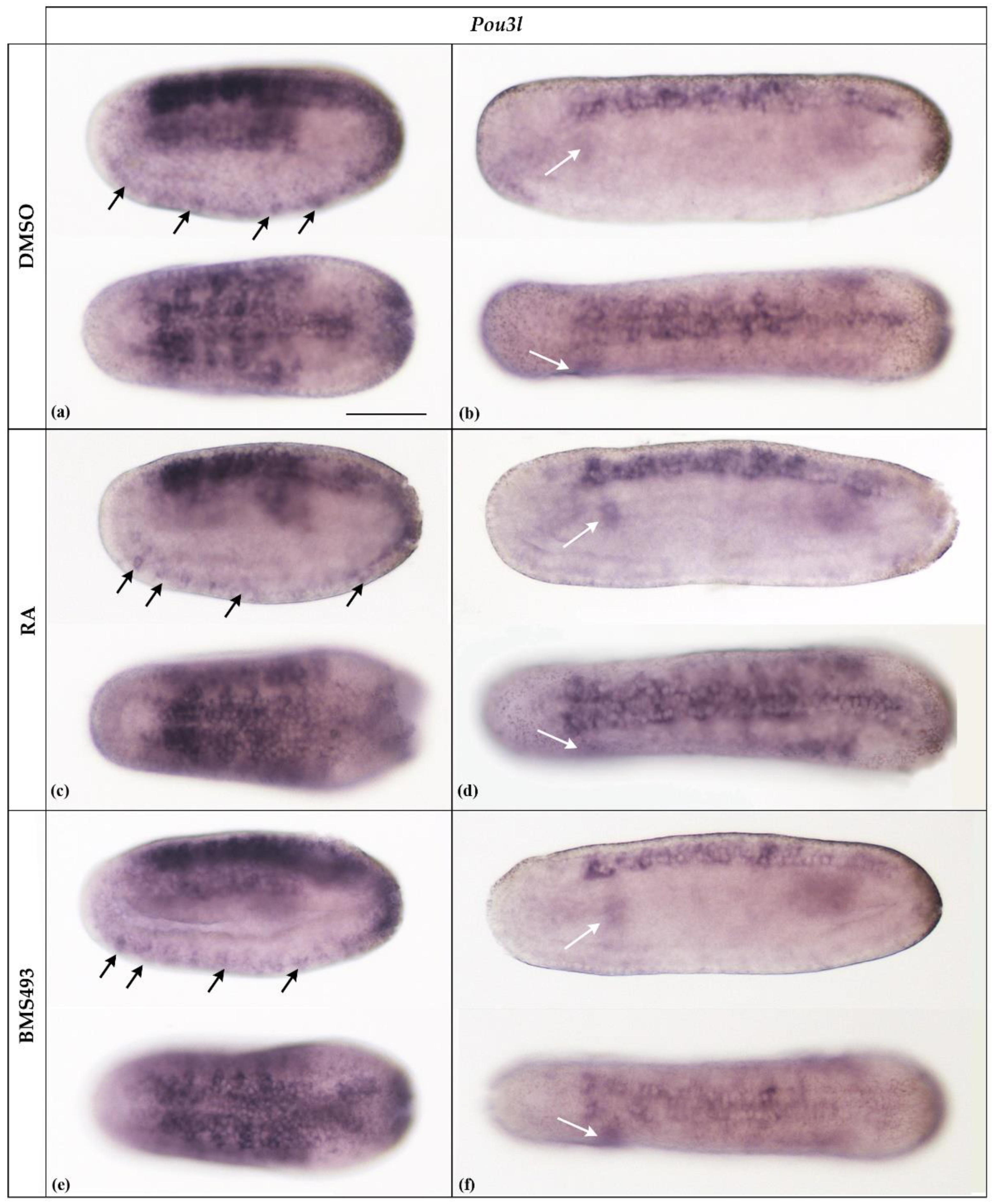
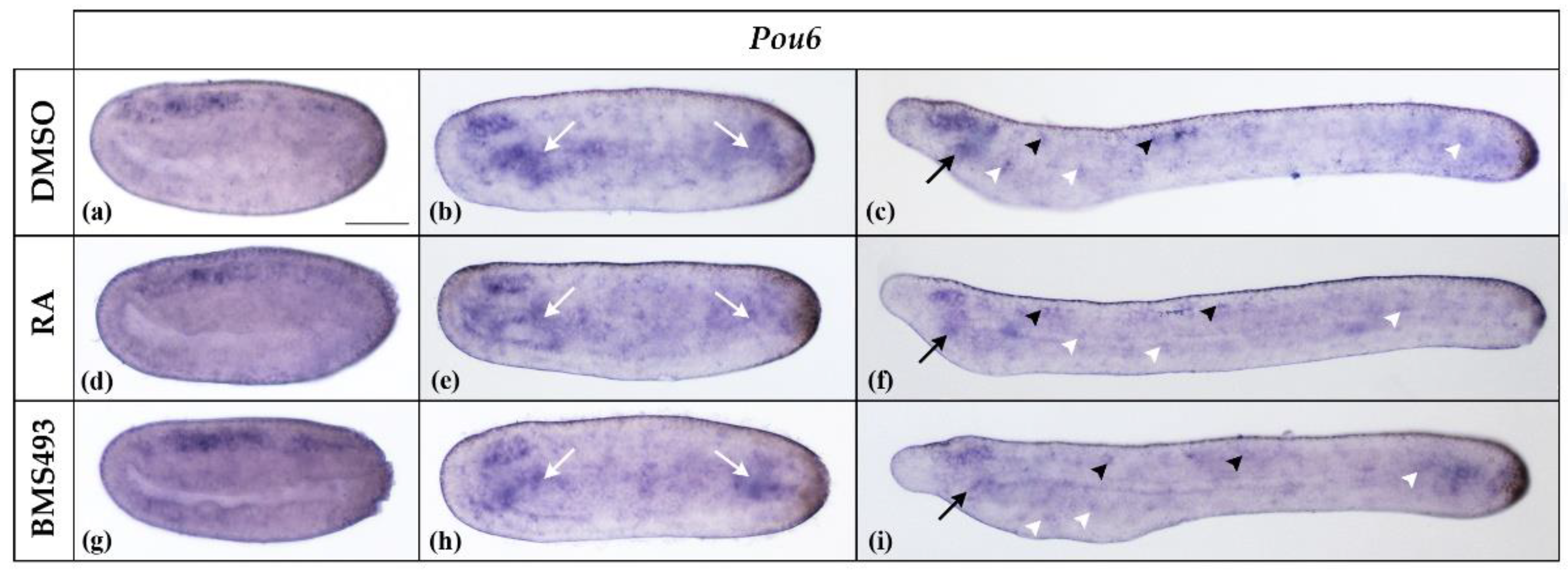
3.1. Retinoic Acid Signaling Does Not Affect Pit-1, Pou2, Pou3l, and Pou6 Expression in Developing Amphioxus
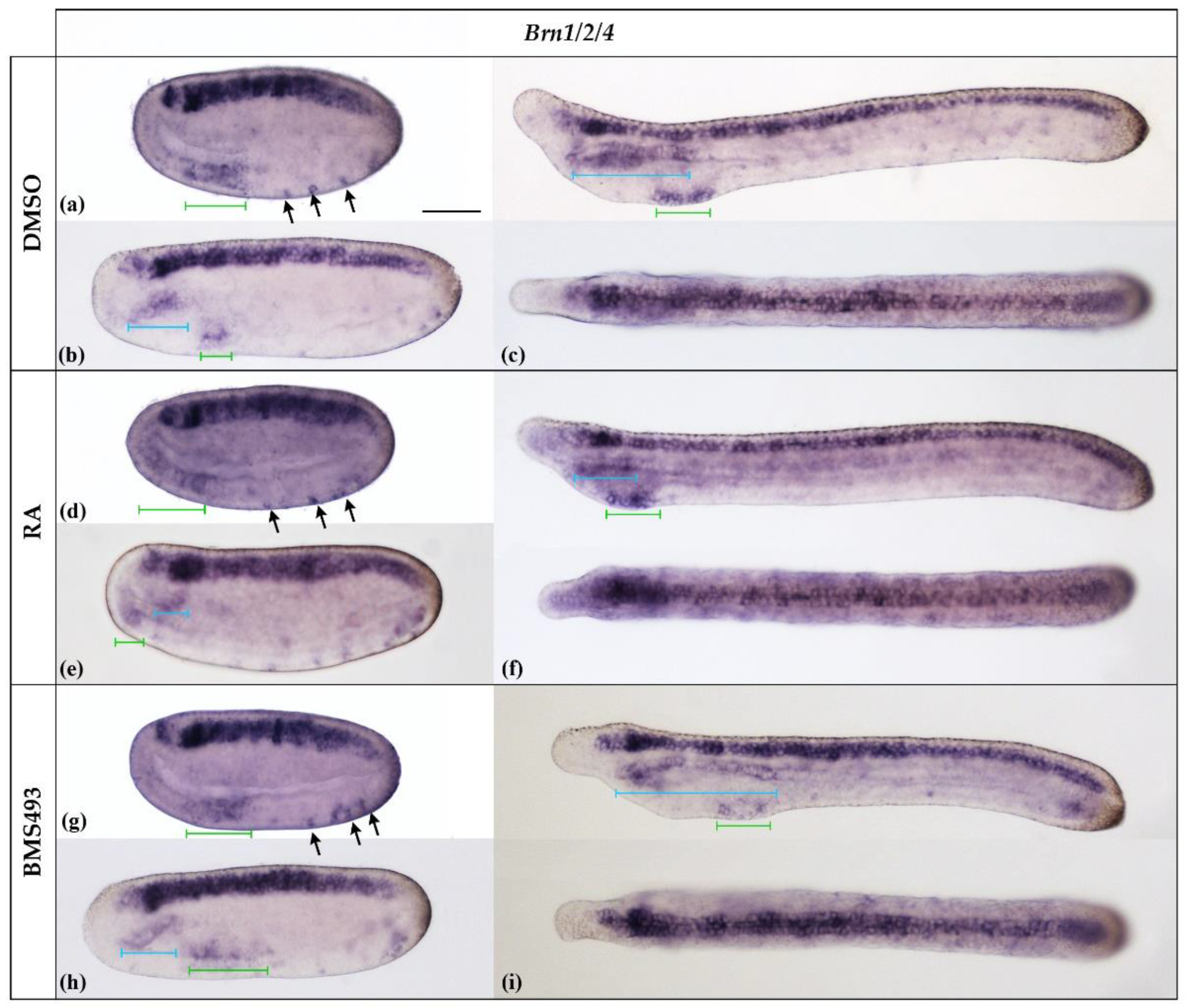
3.2. Pharmacological Disruption of Endogenous Retinoic Acid Signaling Differentially Affects Brn1/2/4 Expression in the Anterior Endoderm
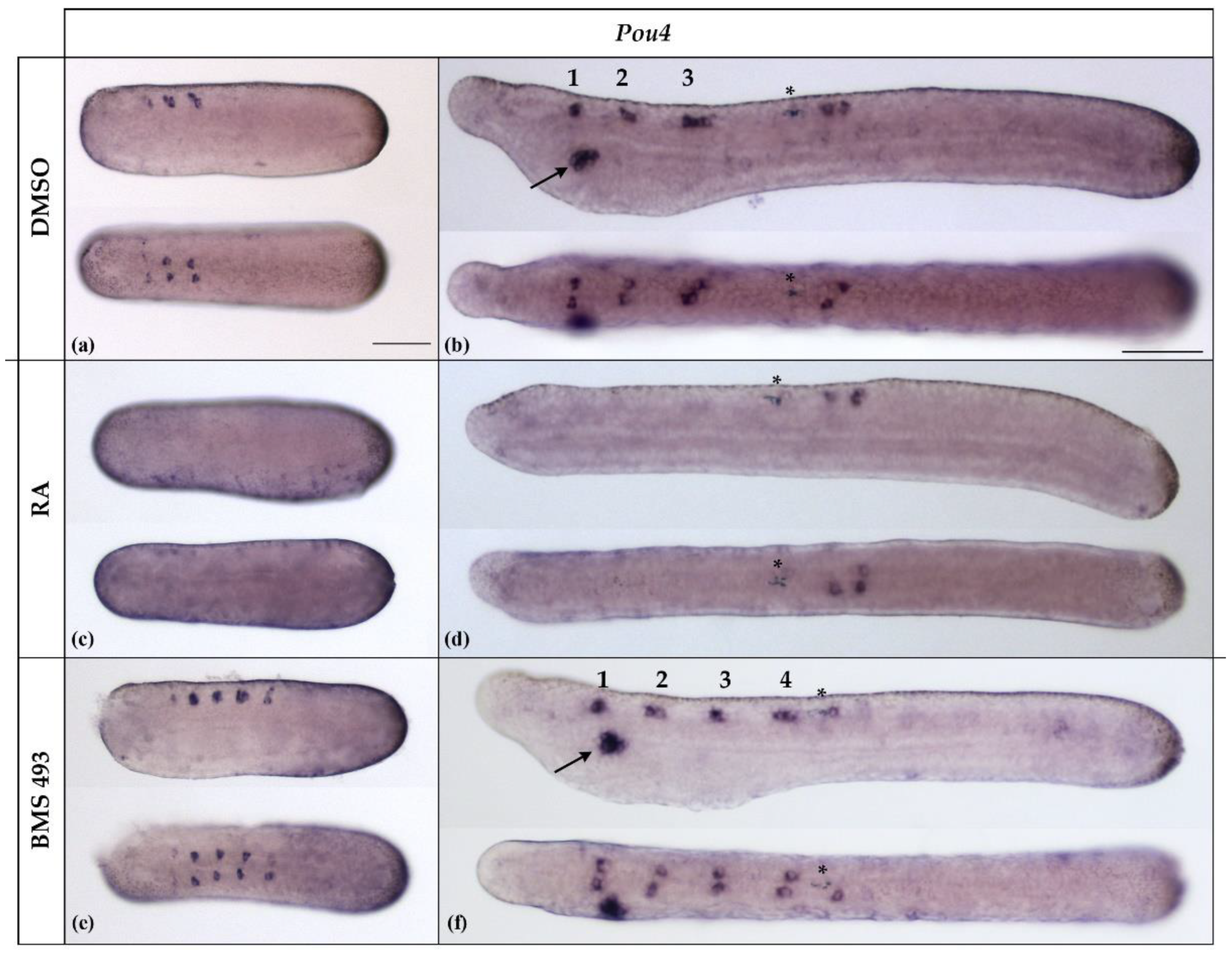
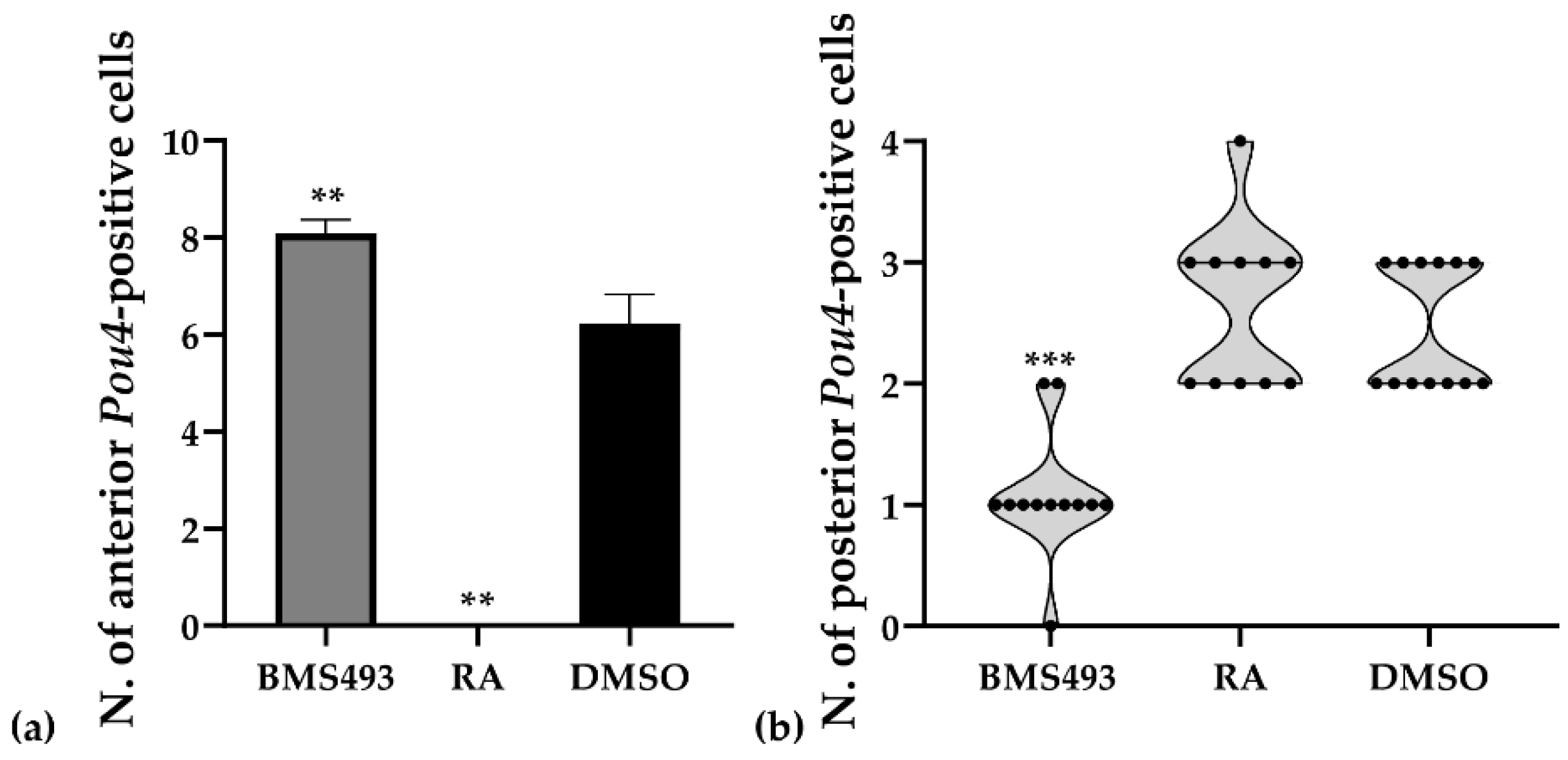
3.3. Interference with Endogenous Retinoic Acid Signaling Levels Differentially Affects Amphioxus Pou4 Expression along the Anteroposterior Axis of the Developing Nerve Cord
4. Discussion
4.1. Evolutionary Considerations on the Expression of POU Genes in Metazoans
4.2. Retinoic Acid-Dependent and -Independent Brn1/2/4 Expression in the Developing Pharynx
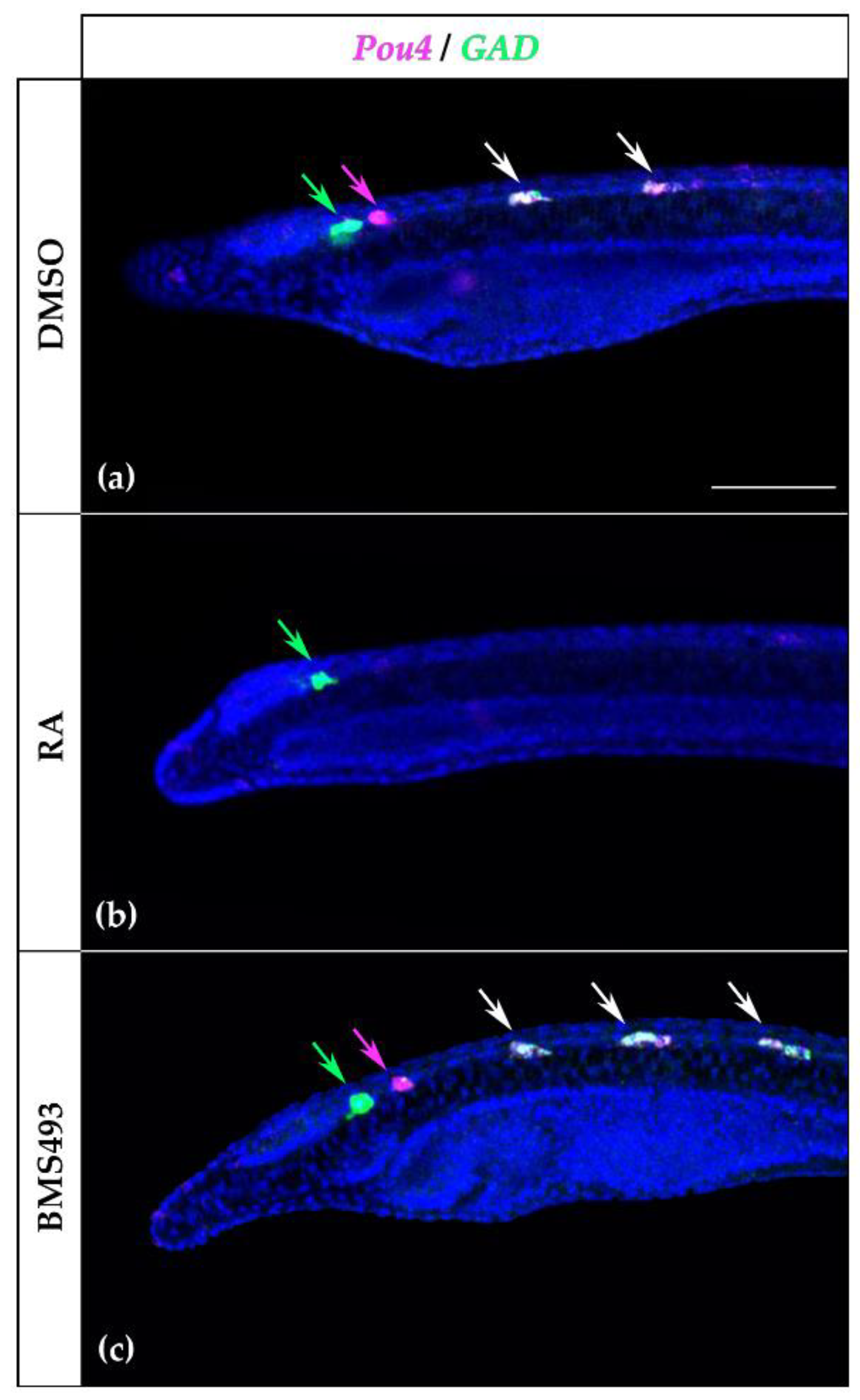
4.3. Evidence for Pou4 as a Terminal Selector of Neuron Types in Amphioxus
4.4. A Graded Response to Retinoic Acid Signaling Regionalizes the Developing Amphioxus Nerve Cord
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gold, D.A.; Gates, R.D.; Jacobs, D.K. The early expansion and evolutionary dynamics of POU class genes. Mol. Biol. Evol. 2014, 31, 3136–3147. [Google Scholar] [CrossRef]
- Phillips, K.; Luisi, B. The virtuoso of versatility: POU proteins that flex to fit. J. Mol. Biol. 2000, 302, 1023–1039. [Google Scholar] [CrossRef]
- Holland, P.W.H.; Booth, H.A.F.; Bruford, E.A. Classification and nomenclature of all human homeobox genes. BMC Biol. 2007, 5, 47. [Google Scholar] [CrossRef]
- Laudet, V.; Zieger, E.; Schubert, M. Evolution of the retinoic acid signaling pathway. In The Retinoids; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 75–90. ISBN 9781118628003. [Google Scholar]
- Carvalho, J.; Schubert, M. Retinoic acid: Metabolism, developmental functions, and evolution. In Vitamin-Binding Proteins—Functional Consequences; Dakshinamurti, K., Dakshinamurti, S., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 1–30. ISBN 9780429067563. [Google Scholar]
- Fonseca, E.S.S.; Hiromori, Y.; Kaite, Y.; Ruivo, R.; Franco, J.N.; Nakanishi, T.; Santos, M.M.; Castro, L.F.C. An orthologue of the retinoic acid receptor (RAR) is present in the ecdysozoa phylum Priapulida. Genes 2019, 10, 985. [Google Scholar] [CrossRef] [PubMed]
- Handberg-Thorsager, M.; Gutierrez-Mazariegos, J.; Arold, S.T.; Nadendla, E.K.; Bertucci, P.Y.; Germain, P.; Tomançak, P.; Pierzchalski, K.; Jones, J.W.; Albalat, R.; et al. The ancestral retinoic acid receptor was a low-affinity sensor triggering neuronal differentiation. Sci. Adv. 2018, 4, eaao1261. [Google Scholar] [CrossRef]
- Putnam, N.H.; Butts, T.; Ferrier, D.E.K.; Furlong, R.F.; Hellsten, U.; Kawashima, T.; Robinson-Rechavi, M.; Shoguchi, E.; Terry, A.; Yu, J.K.; et al. The amphioxus genome and the evolution of the chordate karyotype. Nature 2008, 453, 1064–1071. [Google Scholar] [CrossRef]
- Bertrand, S.; Escriva, H. Evolutionary crossroads in developmental biology: Amphioxus. Development 2011, 138, 4819–4830. [Google Scholar] [CrossRef]
- Albuixech-Crespo, B.; López-Blanch, L.; Burguera, D.; Maeso, I.; Sánchez-Arrones, L.; Moreno-Bravo, J.A.; Somorjai, I.; Pascual-Anaya, J.; Puelles, E.; Bovolenta, P.; et al. Molecular regionalization of the developing amphioxus neural tube challenges major partitions of the vertebrate brain. PLoS Biol. 2017, 15, e2001573. [Google Scholar] [CrossRef]
- Lacalli, T. An evolutionary perspective on chordate brain organization and function: Insights from amphioxus, and the problem of sentience. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2022, 377, 20200520. [Google Scholar] [CrossRef]
- Bozzo, M.; Macrì, S.; Calzia, D.; Sgarra, R.; Manfioletti, G.; Ramoino, P.; Lacalli, T.; Vignali, R.; Pestarino, M.; Candiani, S. The HMGA gene family in chordates: Evolutionary perspectives from amphioxus. Dev. Genes Evol. 2017, 227, 201–211. [Google Scholar] [CrossRef]
- Candiani, S.; Moronti, L.; Ramoino, P.; Schubert, M.; Pestarino, M. A neurochemical map of the developing amphioxus nervous system. BMC Neurosci. 2012, 13, 59. [Google Scholar] [CrossRef] [PubMed]
- Bozzo, M.; Costa, S.; Obino, V.; Bachetti, T.; Marcenaro, E.; Pestarino, M.; Schubert, M.; Candiani, S. Functional conservation and genetic divergence of chordate glycinergic neurotransmission: Insights from amphioxus glycine transporters. Cells 2021, 10, 3392. [Google Scholar] [CrossRef]
- Pergner, J.; Vavrova, A.; Kozmikova, I.; Kozmik, Z. Molecular fingerprint of amphioxus frontal eye illuminates the evolution of homologous cell types in the chordate retina. Front. Cell Dev. Biol. 2020, 8, 705. [Google Scholar] [CrossRef]
- Bozzo, M.; Pergner, J.; Kozmik, Z.; Kozmikova, I. Novel polyclonal antibodies as a useful tool for expression studies in amphioxus embryos. Int. J. Dev. Biol. 2017, 61, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Bozzo, M.; Lacalli, T.C.; Obino, V.; Caicci, F.; Marcenaro, E.; Bachetti, T.; Manni, L.; Pestarino, M.; Schubert, M.; Candiani, S. Amphioxus neuroglia: Molecular characterization and evidence for early compartmentalization of the developing nerve cord. Glia 2021, 69, 1654–1678. [Google Scholar] [CrossRef]
- Takatori, N.; Butts, T.; Candiani, S.; Pestarino, M.; Ferrier, D.E.K.; Saiga, H.; Holland, P.W.H. Comprehensive survey and classification of homeobox genes in the genome of amphioxus, Branchiostoma floridae. Dev. Genes Evol. 2008, 218, 579–590. [Google Scholar] [CrossRef]
- Schubert, M.; Holland, N.D.; Laudet, V.; Holland, L.Z. A retinoic acid-Hox hierarchy controls both anterior/posterior patterning and neuronal specification in the developing central nervous system of the cephalochordate amphioxus. Dev. Biol. 2006, 296, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Koop, D.; Holland, N.D.; Sémon, M.; Alvarez, S.; de Lera, A.R.; Laudet, V.; Holland, L.Z.; Schubert, M. Retinoic acid signaling targets Hox genes during the amphioxus gastrula stage: Insights into early anterior-posterior patterning of the chordate body plan. Dev. Biol. 2010, 338, 98–106. [Google Scholar] [CrossRef]
- Zieger, E.; Candiani, S.; Garbarino, G.; Croce, J.C.; Schubert, M. Roles of retinoic acid signaling in shaping the neuronal architecture of the developing amphioxus nervous system. Mol. Neurobiol. 2018, 55, 5210–5229. [Google Scholar] [CrossRef] [PubMed]
- Zieger, E.; Garbarino, G.; Robert, N.S.M.; Yu, J.-K.; Croce, J.C.; Candiani, S.; Schubert, M. Retinoic acid signaling and neurogenic niche regulation in the developing peripheral nervous system of the cephalochordate amphioxus. Cell. Mol. Life Sci. 2018, 75, 2407–2429. [Google Scholar] [CrossRef]
- Schubert, M.; Yu, K.; Holland, N.D.; Escriva, H.; Laudet, V.; Holland, L.Z. Retinoic acid signaling acts via Hox1 to establish the posterior limit of the pharynx in the chordate amphioxus. Development 2005, 132, 61–73. [Google Scholar] [CrossRef]
- Koop, D.; Chen, J.; Theodosiou, M.; Carvalho, J.E.; Alvarez, S.; de Lera, A.R.; Holland, L.Z.; Schubert, M. Roles of retinoic acid and Tbx1/10 in pharyngeal segmentation: Amphioxus and the ancestral chordate condition. EvoDevo 2014, 5, 36. [Google Scholar] [CrossRef]
- Schubert, M.; Holland, N.D.; Escriva, H.; Holland, L.Z.; Laudet, V. Retinoic acid influences, anteroposterior positioning of epidermal sensory neurons and their gene expression in a developing chordate (amphioxus). Proc. Natl. Acad. Sci. USA 2004, 101, 10320–10325. [Google Scholar] [CrossRef]
- Candiani, S.; Holland, N.D.; Oliveri, D.; Parodi, M.; Pestarino, M. Expression of the amphioxus Pit-1 gene (AmphiPOU1F1/Pit-1) exclusively in the developing preoral organ, a putative homolog of the vertebrate adenohypophysis. Brain Res. Bull. 2008, 75, 324–330. [Google Scholar] [CrossRef]
- Candiani, S.; Pestarino, M. Expression of the tissue-specific transcription factor Pit-1 in the lancelet, Branchiostoma lanceolatum. J. Comp. Neurol. 1998, 392, 343–351. [Google Scholar] [CrossRef]
- Candiani, S.; Pestarino, M. Evidence for the presence of the tissue-specific transcription factor Pit-1 in lancelet larvae. J. Comp. Neurol. 1998, 400, 310–316. [Google Scholar] [CrossRef]
- Candiani, S.; Castagnola, P.; Oliveri, D.; Pestarino, M. Cloning and developmental expression of AmphiBrn1/2/4, a POU III gene in amphioxus. Mech. Dev. 2002, 116, 231–234. [Google Scholar] [CrossRef]
- Leon, A.; Subirana, L.; Magre, K.; Cases, I.; Tena, J.J.; Irimia, M.; Gomez-Skarmeta, J.L.; Escriva, H.; Bertrand, S. Gene regulatory networks of epidermal and neural fate choice in a chordate. Mol. Biol. Evol. 2022, 39, msac055. [Google Scholar] [CrossRef]
- Candiani, S.; Oliveri, D.; Parodi, M.; Bertini, E.; Pestarino, M. Expression of AmphiPOU-IV in the developing neural tube and epidermal sensory neural precursors in amphioxus supports a conserved role of class IV POU genes in the sensory cells development. Dev. Genes Evol. 2006, 216, 623–633. [Google Scholar] [CrossRef]
- Turner, E.E.; Jenne, K.J.; Rosenfeld, M.G. Brn-3.2: A Brn-3-related transcription factor with distinctive central nervous system expression and regulation by retinoic acid. Neuron 1994, 12, 205–218. [Google Scholar] [CrossRef]
- Rosenfeld, M.G. POU-domain transcription factors: Pou-er-ful developmental regulators. Genes Dev. 1991, 5, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Urban, S.; Kobi, D.; Ennen, M.; Langer, D.; Le Gras, S.; Ye, T.; Davidson, I. A Brn2-Zic1 axis specifies the neuronal fate of retinoic-acidtreated embryonic stem cells. J. Cell Sci. 2015, 128, 2303–2318. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, M.; Schubert, M.; Dalfo, D.; Candiani, S.; Benito, E.; Gardenyes, J.; Godoy, L.; Moret, F.; Illas, M.; Patten, I.; et al. Preliminary observations on the spawning conditions of the European amphioxus (Branchiostoma lanceolatum) in captivity. J. Exp. Zool. Part B Mol. Dev. Evol. 2004, 302, 384–391. [Google Scholar] [CrossRef]
- Bozzo, M.; Candiani, S.; Schubert, M. Whole mount in situ hybridization and immunohistochemistry for studying retinoic acid signaling in developing amphioxus. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2020; Volume 637, pp. 419–452. ISBN 9780128201442. [Google Scholar]
- Germain, P.; Gaudon, C.; Pogenberg, V.; Sanglier, S.; Van Dorsselaer, A.; Royer, C.A.; Lazar, M.A.; Bourguet, W.; Gronemeyer, H. Differential action on coregulator interaction defines inverse retinoid agonists and neutral antagonists. Chem. Biol. 2009, 16, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.E.; Lahaye, F.; Yong, L.W.; Croce, J.C.; Escrivá, H.; Yu, J.-K.; Schubert, M. An updated staging system for cephalochordate development: One table suits them all. Front. Cell Dev. Biol. 2021, 9, 668006. [Google Scholar] [CrossRef]
- Brasó-Vives, M.; Marlétaz, F.; Echchiki, A.; Mantica, F.; Acemel, R.D.; Gómez-Skarmeta, J.L.; Hartasánchez, D.A.; Le Targa, L.; Pontarotti, P.; Tena, J.J.; et al. Parallel evolution of amphioxus and vertebrate small-scale gene duplications. Genome Biol. 2022, 23, 243. [Google Scholar] [CrossRef]
- Choi, H.M.T.; Schwarzkopf, M.; Fornace, M.E.; Acharya, A.; Artavanis, G.; Stegmaier, J.; Cunha, A.; Pierce, N.A. Third-generation in situ hybridization chain reaction: Multiplexed, quantitative, sensitive, versatile, robust. Development 2018, 145, dev165753. [Google Scholar] [CrossRef]
- Kuehn, E.; Clausen, D.S.; Null, R.W.; Metzger, B.M.; Willis, A.D.; Özpolat, B.D. Segment number threshold determines juvenile onset of germline cluster expansion in Platynereis dumerilii. J. Exp. Zool. Part B Mol. Dev. Evol. 2022, 338, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Bruce, H.S.; Jerz, G.; Kelly, S.R.; McCarthy, J.; Pomerantz, A.; Senevirathne, G.; Sherrard, A.; Sun, D.A.; Wolff, C.; Patel, N.H. Hybridization Chain Reaction (HCR) In Situ Protocol. 2021. Available online: https://dx.doi.org/10.17504/protocols.io.bunznvf6 (accessed on 9 August 2022).
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Escriva, H.; Holland, N.D.; Gronemeyer, H.; Laudet, V.; Holland, L.Z. The retinoic acid signaling pathway regulates anterior/posterior patterning in the nerve cord and pharynx of amphioxus, a chordate lacking neural crest. Development 2002, 129, 2905–2916. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, J.E.; Lahaye, F.; Croce, J.C.; Schubert, M. CYP26 function is required for the tissue-specific modulation of retinoic acid signaling during amphioxus development. Int. J. Dev. Biol. 2017, 61, 733–747. [Google Scholar] [CrossRef]
- Paps, J.; Xu, F.; Zhang, G.; Holland, P.W.H. Reinforcing the egg-timer: Recruitment of novel Lophotrochozoa homeobox genes to early and late development in the Pacific oyster. Genome Biol. Evol. 2015, 7, 677–688. [Google Scholar] [CrossRef]
- Ingraham, H.A.; Chen, R.; Mangalam, H.J.; Elsholtz, H.P.; Flynn, S.E.; Lin, C.R.; Simmons, D.M.; Swanson, L.; Rosenfeld, M.G. A tissue-specific transcription factor containing a homeodomain specifies a pituitary phenotype. Cell 1988, 55, 519–529. [Google Scholar] [CrossRef]
- Bodner, M.; Castriilo, J.L.; Theill, L.E.; Deerinck, T.; Ellisman, M.; Karin, M. The pituitary-specific transcription factor GHF-1 is a homeobox-containing protein. Cell 1988, 55, 505–518. [Google Scholar] [CrossRef]
- Malik, V.; Zimmer, D.; Jauch, R. Diversity among POU transcription factors in chromatin recognition and cell fate reprogramming. Cell. Mol. Life Sci. 2018, 75, 1587–1612. [Google Scholar] [CrossRef]
- Kozmik, Z.; Holland, N.D.; Kreslova, J.; Oliveri, D.; Schubert, M.; Jonasova, K.; Holland, L.Z.; Pestarino, M.; Benes, V.; Candiani, S. Pax–Six–Eya–Dach network during amphioxus development: Conservation in vitro but context specificity in vivo. Dev. Biol. 2007, 306, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, S.J.; Chen, R.; DiMattia, G.E.; Scully, K.M.; Kalla, K.A.; Lin, S.C.; Yu, V.C.; Rosenfeld, M.G. A tissue-specific enhancer confers Pit-1-dependent morphogen inducibility and autoregulation on the pit-1 gene. Genes Dev. 1993, 7, 913–932. [Google Scholar] [CrossRef] [PubMed]
- Maden, M.; Blentic, A.; Reijntjes, S.; Seguin, S.; Gale, E.; Graham, A. Retinoic acid is required for specification of the ventral eye field and for Rathke’s pouch in the avian embryo. Int. J. Dev. Biol. 2007, 51, 191–200. [Google Scholar] [CrossRef]
- Sturm, R.A.; Das, G.; Herr, W. The ubiquitous octamer-binding protein Oct-1 contains a POU domain with a homeo box subdomain. Genes Dev. 1988, 2, 1582–1599. [Google Scholar] [CrossRef]
- Andersen, B.; Weinberg, W.C.; Rennekampff, O.; McEvilly, R.J.; Bermingham, J.R.; Hooshmand, F.; Vasilyev, V.; Hansbrough, J.F.; Pittelkow, M.R.; Yuspa, S.H.; et al. Functions of the POU domain genes skn-1a/i and Tst-1/Oct-6/SCIP in epidermal differentiation. Genes Dev. 1997, 11, 1873–1884. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, I.; Ohmoto, M.; Narukawa, M.; Yoshihara, Y.; Abe, K. Skn-1a (Pou2f3) specifies taste receptor cell lineage. Nat. Neurosci. 2011, 14, 685–687. [Google Scholar] [CrossRef] [PubMed]
- Wollesen, T.; McDougall, C.; Degnan, B.M.; Wanninger, A. POU genes are expressed during the formation of individual ganglia of the cephalopod central nervous system. EvoDevo 2014, 5, 41. [Google Scholar] [CrossRef] [PubMed]
- Rieger, A.; Kemter, E.; Kumar, S.; Popper, B.; Aigner, B.; Wolf, E.; Wanke, R.; Blutke, A. Missense mutation of POU domain class 3 transcription factor 3 in Pou3f3L423P mice causes reduced nephron number and impaired development of the thick ascending limb of the loop of Henle. PLoS ONE 2016, 11, e0158977. [Google Scholar] [CrossRef] [PubMed]
- Gąsiorowski, L.; Andrikou, C.; Janssen, R.; Bump, P.; Budd, G.E.; Lowe, C.J.; Hejnol, A. Molecular evidence for a single origin of ultrafiltration-based excretory organs. Curr. Biol. 2021, 31, 3629–3638.e2. [Google Scholar] [CrossRef]
- Matsuo-Takasaki, M.; Lim, J.H.; Sato, S.M. The POU domain gene, XlPOU 2 is an essential downstream determinant of neural induction. Mech. Dev. 1999, 89, 75–85. [Google Scholar] [CrossRef]
- Zhu, Q.; Song, L.; Peng, G.; Sun, N.; Chen, J.; Zhang, T.; Sheng, N.; Tang, W.; Qian, C.; Qiao, Y.; et al. The transcription factor Pou3f1 promotes neural fate commitment via activation of neural lineage genes and inhibition of external signaling pathways. eLife 2014, 3, e02224. [Google Scholar] [CrossRef]
- Holland, N.D. Formation of the initial kidney and mouth opening in larval amphioxus studied with serial blockface scanning electron microscopy (SBSEM). EvoDevo 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Barske, L.; Fabian, P.; Hirschberger, C.; Jandzik, D.; Square, T.; Xu, P.; Nelson, N.; Vincent Yu, H.; Medeiros, D.M.; Gillis, J.A.; et al. Evolution of vertebrate gill covers via shifts in an ancient Pou3f3 enhancer. Proc. Natl. Acad. Sci. USA 2020, 117, 24876–24884. [Google Scholar] [CrossRef]
- Lankester, E.R.; Willey, A. Memoirs: The development of the atrial chamber of amphioxus. J. Cell Sci. 1890, 2, 445–466. [Google Scholar] [CrossRef]
- Clyne, P.J.; Certel, S.J.; De Bruyne, M.; Zaslavsky, L.; Johnson, W.A.; Carlson, J.R. The odor specificities of a subset of olfactory receptor neurons are governed by Acj6, a POU-domain transcription factor. Neuron 1999, 22, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Sze, J.Y.; Ruvkun, G. Activity of the Caenorhabditis elegans UNC-86 POU transcription factor modulates olfactory sensitivity. Proc. Natl. Acad. Sci. USA 2003, 100, 9560–9565. [Google Scholar] [CrossRef] [PubMed]
- Finney, M.; Ruvkun, G. The unc-86 gene product couples cell lineage and cell identity in C. elegans. Cell 1990, 63, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Tournière, O.; Dolan, D.; Richards, G.S.; Sunagar, K.; Columbus-Shenkar, Y.Y.; Moran, Y.; Rentzsch, F. NvPOU4/Brain3 functions as a terminal selector gene in the nervous system of the cnidarian Nematostella vectensis. Cell Rep. 2020, 30, 4473–4489.e5. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.K.; Rosenfeld, M.G. POU domain family values: Flexibility, partnerships, and developmental codes. Genes Dev. 1997, 11, 1207–1225. [Google Scholar] [CrossRef]
- Xiang, M.; Gan, L.; Li, D.; Chen, Z.Y.; Zhou, L.; O’Malley, B.W.; Klein, W.; Nathans, J. Essential role of POU-domain factor Brn-3c in auditory and vestibular hair cell development. Proc. Natl. Acad. Sci. USA 1997, 94, 9445–9450. [Google Scholar] [CrossRef]
- Erkman, L.; McEvilly, R.J.; Luo, L.; Ryan, A.K.; Hooshmand, F.; O’connell, S.M.; Keithley, E.M.; Rapaport, D.H.; Ryan, A.F.; Rosenfeld, M.G. Role of transcription factors Brn-3.1 and Brn-3.2 in auditory and visual system development. Nature 1996, 381, 603–606. [Google Scholar] [CrossRef]
- Gan, L.; Xiang, M.; Zhou, L.; Wagner, D.S.; Klein, W.H.; Nathans, J. POU domain factor Brn-3b is required for the development of a large set of retinal ganglion cells. Proc. Natl. Acad. Sci. USA 1996, 93, 3920–3925. [Google Scholar] [CrossRef] [PubMed]
- Candiani, S.; Pennati, R.; Oliveri, D.; Locascio, A.; Branno, M.; Castagnola, P.; Pestarino, M.; De Bernardi, F. Ci-POU-IV expression identifies PNS neurons in embryos and larvae of the ascidian Ciona intestinalis. Dev. Genes Evol. 2005, 215, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Ozment, E.; Tamvacakis, A.N.; Zhou, J.; Rosiles-Loeza, P.Y.; Escobar-Hernandez, E.E.; Fernandez-Valverde, S.L.; Nakanishi, N. Cnidarian hair cell development illuminates an ancient role for the class IV POU transcription factor in defining mechanoreceptor identity. eLife 2021, 10, e74336. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Morrison, G.M.; Brickman, J.M. Conserved roles for Oct4 homologues in maintaining multipotency during early vertebrate development. Development 2006, 133, 2011–2022. [Google Scholar] [CrossRef]
- Vanni, V.; Salonna, M.; Gasparini, F.; Martini, M.; Anselmi, C.; Gissi, C.; Manni, L. Yamanaka factors in the budding tunicate Botryllus schlosseri show a shared spatio-temporal expression pattern in chordates. Front. Cell Dev. Biol. 2022, 10, 782722. [Google Scholar] [CrossRef]
- Somorjai, I.M.L. Amphioxus regeneration: Evolutionary and biomedical implications. Int. J. Dev. Biol. 2017, 61, 689–696. [Google Scholar] [CrossRef]
- Sobreira, T.J.P.; Marlétaz, F.; Simões-Costa, M.; Schechtman, D.; Pereira, A.C.; Brunet, F.; Sweeney, S.; Pani, A.; Aronowicz, J.; Lowe, C.J.; et al. Structural shifts of aldehyde dehydrogenase enzymes were instrumental for the early evolution of retinoid-dependent axial patterning in metazoans. Proc. Natl. Acad. Sci. USA 2011, 108, 226–231. [Google Scholar] [CrossRef]
- Carvalho, J.E.; Theodosiou, M.; Chen, J.; Chevret, P.; Alvarez, S.; De Lera, A.R.; Laudet, V.; Croce, J.C.; Schubert, M. Lineage-specific duplication of amphioxus retinoic acid degrading enzymes (CYP26) resulted in sub-functionalization of patterning and homeostatic roles. BMC Evol. Biol. 2017, 17, 24. [Google Scholar] [CrossRef]
- Lee, M.H.; Salvaterra, P.M. Abnormal chemosensory jump 6 is a positive transcriptional regulator of the cholinergic gene locus in Drosophila olfactory neurons. J. Neurosci. 2002, 22, 5291–5299. [Google Scholar] [CrossRef]
- Leyva-Díaz, E.; Masoudi, N.; Serrano-Saiz, E.; Glenwinkel, L.; Hobert, O. Brn3/POU-IV-type POU homeobox genes—Paradigmatic regulators of neuronal identity across phylogeny. Wiley Interdiscip. Rev. Dev. Biol. 2020, 9, e374. [Google Scholar] [CrossRef]
- Serrano-Saiz, E.; Leyva-Díaz, E.; De La Cruz, E.; Hobert, O. BRN3-type POU homeobox genes maintain the identity of mature postmitotic neurons in nematodes and mice. Curr. Biol. 2018, 28, 2813–2823.e2. [Google Scholar] [CrossRef]
- Pennati, R.; Candiani, S.; Biggiogero, M.; Zega, G.; Groppelli, S.; Oliveri, D.; Parodi, M.; Di Bernardi, F.; Pestarino, M. Developmental expression of tryptophan hydroxylase gene in Ciona intestinalis. Dev. Genes Evol. 2007, 217, 307–313. [Google Scholar] [CrossRef]
- Lacalli, T.; Candiani, S. Locomotory control in amphioxus larvae: New insights from neurotransmitter data. EvoDevo 2017, 8, 4. [Google Scholar] [CrossRef] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bozzo, M.; Bellitto, D.; Amaroli, A.; Ferrando, S.; Schubert, M.; Candiani, S. Retinoic Acid and POU Genes in Developing Amphioxus: A Focus on Neural Development. Cells 2023, 12, 614. https://doi.org/10.3390/cells12040614
Bozzo M, Bellitto D, Amaroli A, Ferrando S, Schubert M, Candiani S. Retinoic Acid and POU Genes in Developing Amphioxus: A Focus on Neural Development. Cells. 2023; 12(4):614. https://doi.org/10.3390/cells12040614
Chicago/Turabian StyleBozzo, Matteo, Deianira Bellitto, Andrea Amaroli, Sara Ferrando, Michael Schubert, and Simona Candiani. 2023. "Retinoic Acid and POU Genes in Developing Amphioxus: A Focus on Neural Development" Cells 12, no. 4: 614. https://doi.org/10.3390/cells12040614
APA StyleBozzo, M., Bellitto, D., Amaroli, A., Ferrando, S., Schubert, M., & Candiani, S. (2023). Retinoic Acid and POU Genes in Developing Amphioxus: A Focus on Neural Development. Cells, 12(4), 614. https://doi.org/10.3390/cells12040614