
Analysis of the Anticipatory Behavior Formation Mechanism Induced by Methamphetamine Using a Single Hair
 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Methamphetamine Injection
2.3. Locomotor Activity Rhythm
2.4. Measurement of Abdominal Temperature
2.5. Detection of Per1 Expression Rhythms in the Skin of Freely Moving Mice
2.6. Detection of MAP-Induced Per1 Expression in the Scalp Hairs Using Micro-PMT
2.7. Detection of MAP-Induced Per1 Expression in a Whisker Hair Using Micro-PMT
2.7.1. Solution Method
2.7.2. Direct Methods
2.8. LUC Expression in Scalp and Whisker Hairs by Immunohistological Analysis
2.9. Statistics
3. Results
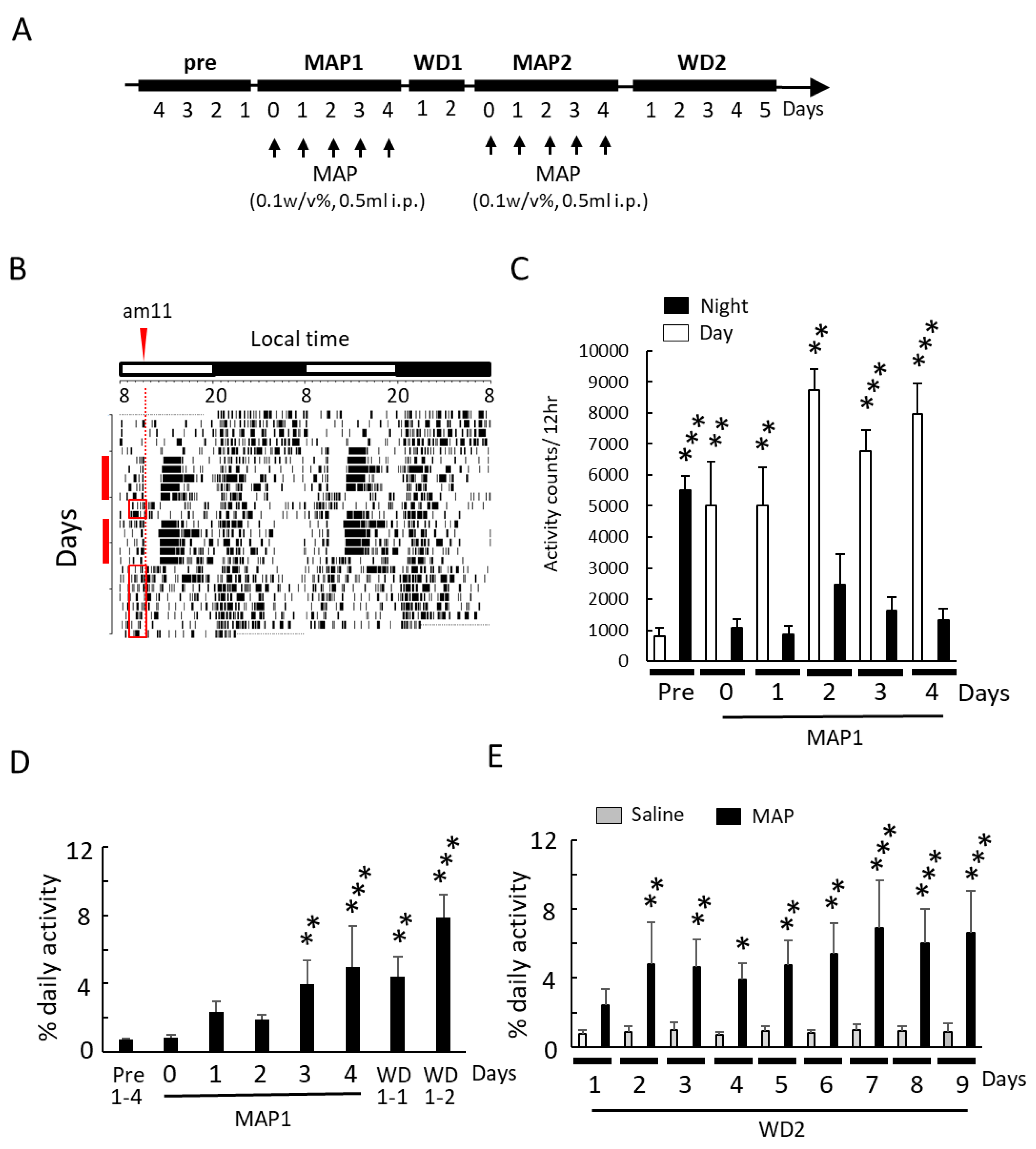
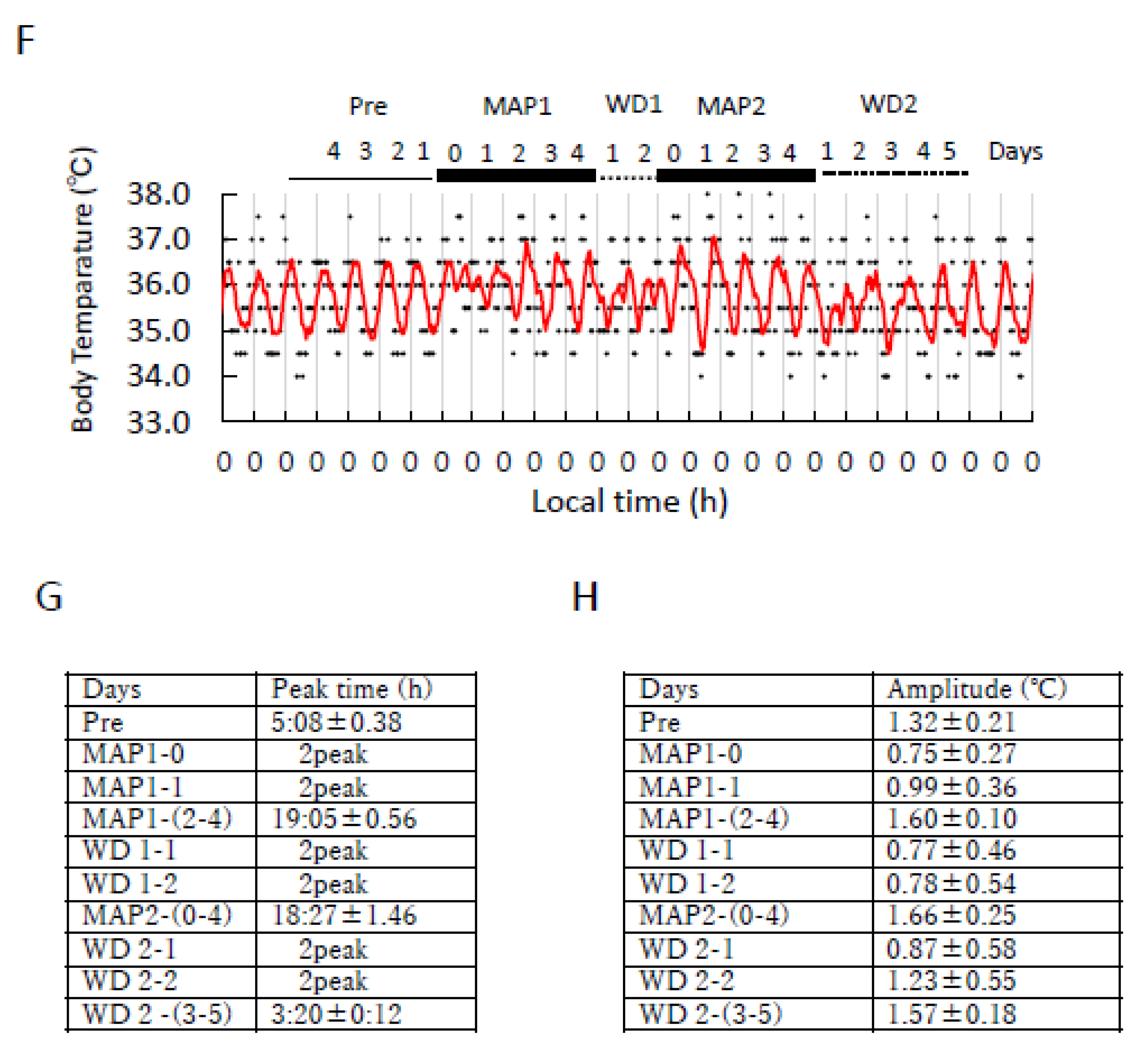
3.1. Effect of Daily MAP Injection on Locomotor Activity and Body Temperature Rhythm
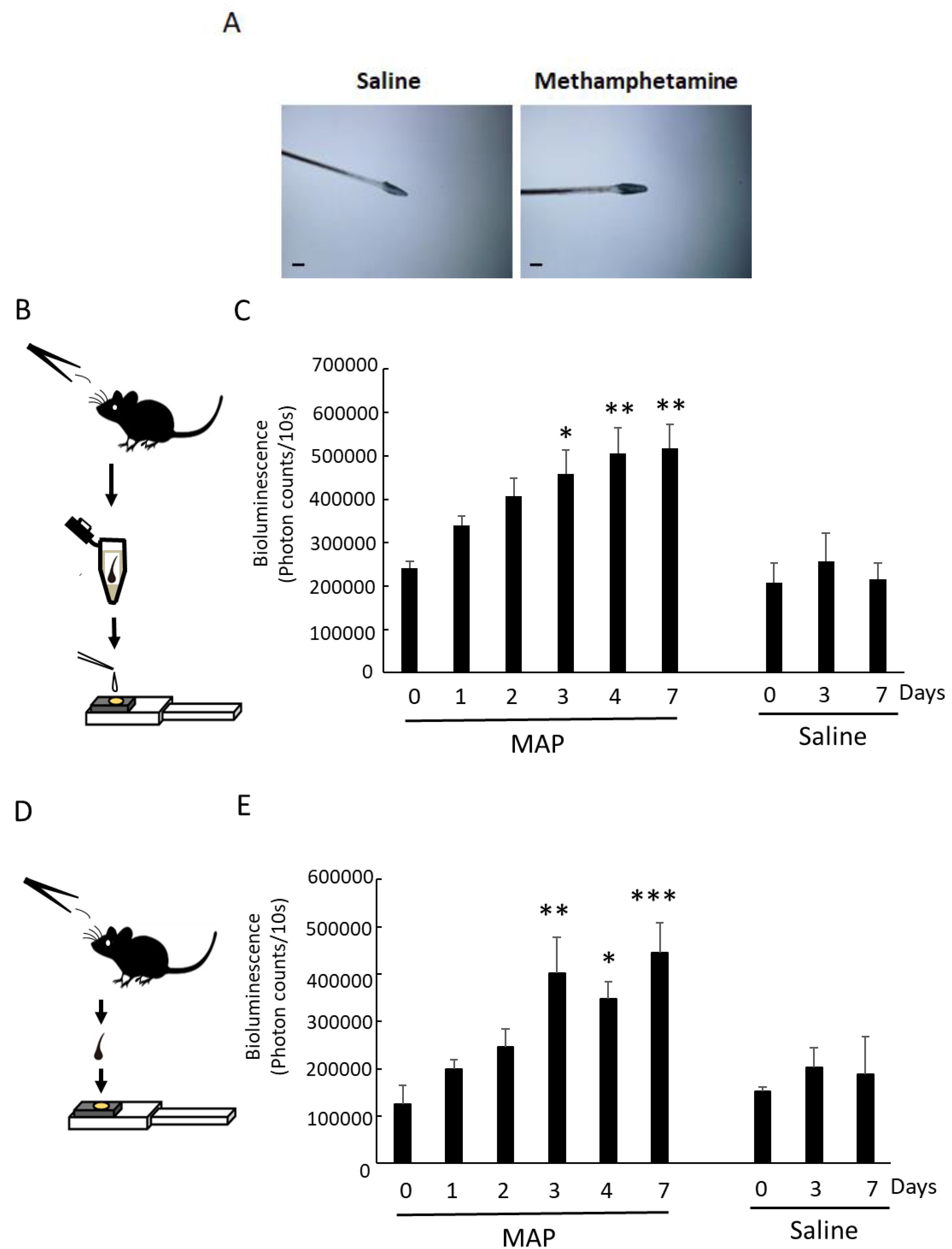
3.2. MAP-Induced Per1 Gene Expression in the Scalp Hairs
3.3. MAP-Induced Per1 Gene Expression in a Whisker Hair
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, R.Y.; Eichler, V.B. Loss of a circadian adrenal corticosterone rhythm following suprachiasmatic lesions in the rat. Brain Res. 1972, 42, 201–206. [Google Scholar] [CrossRef]
- Stephan, F.K.; Zucker, I. Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proc. Natl. Acad. Sci. USA 1972, 69, 1583–1586. [Google Scholar] [CrossRef] [Green Version]
- Honma, K.; Honma, S.; Hiroshige, T. Disorganization of the rat activity rhythm by chronic treatment with methamphetamine. Physiol. Behav. 1986, 38, 687–695. [Google Scholar] [CrossRef]
- Tataroglu, O.; Davidson, A.J.; Benvenuto, L.J.; Menaker, M. The methamphetamine-sensitive circadian oscillator (MASCO) in mice. J. Biol. Rhythms 2006, 21, 185–194. [Google Scholar] [CrossRef]
- Honma, K.; Honma, S. The SCN-independent clocks, methamphetamine and food restriction. Eur. J. Neurosci. 2009, 30, 1707–1717. [Google Scholar] [CrossRef]
- Honma, S.; Honma, K.; Hiroshige, T. Methamphetamine effects on rat circadian clock depend on actograph. Physiol. Behav. 1991, 49, 787–795. [Google Scholar] [CrossRef]
- Shibata, S.; Minamoto, Y.; Ono, M.; Watanabe, S. Aging impairs methamphetamine-induced free-running and anticipatory locomotor activity rhythms in rats. Neurosci. Lett. 1994, 172, 107–110. [Google Scholar] [CrossRef]
- Iijima, M.; Nikaido, T.; Akiyama, M.; Moriya, T.; Shibata, S. Methamphetamine-induced, suprachiasmatic nucleus-independent circadian rhythms of activity and mPer gene expression in the striatum of the mouse. Eur. J. Neurosci. 2002, 6, 921–929. [Google Scholar] [CrossRef]
- Mohawk, J.A.; Pezuk, P.; Menaker, M. Methamphetamine and dopamine receptor D1 regulate entrainment of murine circadian oscillators. PLoS ONE 2013, 8, e62463. [Google Scholar] [CrossRef] [Green Version]
- Honma, S.; Honma, K. Phase-dependent phase shift of methamphetamine-induced circadian rhythm by haloperidol in SCN-lesioned rats. Brain Res. 1995, 674, 283–290. [Google Scholar] [CrossRef]
- Shibata, S.; Ono, M.; Fukuhara, N.; Watanabe, S. Involvement of dopamine, N-methyl-D-aspartate and sigma receptor mechanisms in methamphetamine-induced anticipatory activity rhythm in rats. J. Pharmacol. Exp. Ther. 1995, 274, 688–694. [Google Scholar]
- Kanou, H.; Nagasawa, K.; Ishii, Y.; Chishima, A.; Hayashi, J.; Haga, S.; Sutherland, K.; Ishikawa, M.; Ozaki, M.; Shirato, H.; et al. Period1 gene expression in the olfactory bulb and liver of freely moving streptozotocin-treated diabetic mouse. Biochem. Biophys. Res. Commun. 2021, 560, 14–20. [Google Scholar] [CrossRef]
- Hamada, K.; Ishii, Y.; Yoshida, Y.; Nakaya, M.; Sato, Y.; Kanai, M.; Kikuchi, Y.; Yamaguchi, T.; Iijima, N.; Sutherland, K.; et al. The analysis of Period1 gene expression in vivo and in vitro using a micro PMT system. Biophys. Res. Commun. 2021, 577, 64–70. [Google Scholar] [CrossRef]
- Hida, A.; Koike, N.; Hirose, M.; Hattori, M.; Sakaki, Y.; Tei, H. The human and mouse Period1 genes: Five well-conserved E-boxes additively contribute to the enhancement of mPer1 transcription. Genomics 2000, 65, 224–233. [Google Scholar] [CrossRef]
- Hamada, K.; Oota, A.; Ito, R.; Kasahara, S.; Nakajima, K.; Kikuchi, Y.; Sutherland, K.; Ishikawa, M.; Shirato, H.; Ozaki, M.; et al. Double recording system of Period1 gene expression rhythm in the olfactory bulb and liver in freely moving mouse. Biophys. Res. Commun. 2020, 529, 898–903. [Google Scholar] [CrossRef]
- Ito, R.; Hamada, K.; Kasahara, S.; Kikuchi, Y.; Nakajima, K.; Sutherland, K.; Shirato, H.; Ozaki, M.; Ishikawa, M.; Hamada, T. Mouse period1 gene expression recording from olfactory bulb under free moving conditions with a portable optic fiber device. Luminescence 2020, 35, 1248–1253. [Google Scholar] [CrossRef]
- Hamada, T.; Sutherland, K.; Ishikawa, M.; Miyamoto, N.; Honma, S.; Shirato, H.; Honma, K.-I. In vivo imaging of clock gene expression in multiple tissues of freely moving mice. Nat. Commun. 2016, 7, 11705. [Google Scholar] [CrossRef] [Green Version]
- Hamada, T.; Honma, S.; Honma, K. Light responsiveness of clock genes, Per1 and Per2, in the olfactory bulb of mice. Biochem. Biophys. Res. Commun. 2011, 409, 727–731. [Google Scholar] [CrossRef]
- Mistlberger, R.E. Circadian food-anticipatory activity: Formal models and physiological mechanisms. Neurosci. Biobehav. Rev. 1994, 18, 171–195. [Google Scholar] [CrossRef]
- Kuribara, H.; Tadokoro, S. Reverse tolerance to ambulation-increasing effects of methamphetamine and morphine in 6 mouse strains. Jpn. J. Pharmacol. 1989, 49, 197–203. [Google Scholar] [CrossRef]
- Tanioka, M.; Yamada, H.; Do, M.; Bando, H.; Yamaguchi, Y.; Nishigori, C.; Okamur, H. Molecular clocks in mouse skin. J. Investig. Dermatol. 2009, 129, 1225–1231. [Google Scholar] [CrossRef]
- Cook, C.E.; Jeffcoat, A.R.; Sadler, B.M.; Hill, J.M.; Voyksner, R.D.; Pugh, D.E.; White, W.R.; Perez-Reyes, M. Pharmacokinetics of oral methamphetamine and effects of repeated daily dosing in humans. Drug Metab. Dispos. 1992, 20, 856–862. [Google Scholar]
- Thompson, J.F.; Hayes, L.S.; Lloyd, D.B. Modulation of firefly luciferase stability and impact on studies of gene regulation. Gene 1991, 103, 171–177. [Google Scholar] [CrossRef]
- Ignowski, J.M.; Schaffer, D.V. Kinetic Analysis and Modeling of Firefly Luciferase as a Quantitative reporter gene in live mammalian cells. Biotechnol. Bioeng. 2004, 86, 827–834. [Google Scholar] [CrossRef]
- Buhr, E.D.; Yoo, S.H.; Takahashi, J.S. Temperature as a universal resetting cue for mammalian circadian oscillators. Science 2010, 330, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Zanellom, S.B.; Jackson, D.M.; Holic, M.F. Expression of the circadian clock genes clock and period1 in human skin. J. Vest. Dermatol. 2000, 115, 757–760. [Google Scholar]
- Bjarnason, G.A.; Jordan, R.C.; Wood, P.A.; Li, Q.; Lincoln, D.W.; Sothern, R.B.; Hrushesky, W.J.; Ben-David, Y. Circadian expression of clock genes in human oral mucosa and skin: Association with specific cell-cycle phases. Am. J. Pathol. 2001, 158, 1793–1801. [Google Scholar] [CrossRef]
- Kawara, S.; Mydlarski, R.; Mamelak, A.J.; Freed, I.; Wang, B.; Watanabe, H.; Shivji, G.; Tavadia, S.K.; Suzuki, H.; Bjarnason, G.A.; et al. Low-dose ultraviolet B rays alter the mRNA expression of the circadian clock genes in cultured human keratinocytes. J. Investig. Dermatol. 2002, 19, 1220–1223. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.A.; Fleury-Olela, F.; Nagoshi, E.; Hauser, C.; Juge, C.; Meier, C.A.; Chicheportiche, R.; Dayer, J.-M.; Albrecht, U.; Schibler, U. The period length of fibroblast circadian gene expression varies widely among human individuals. PLoS Biol. 2005, 3, e338. [Google Scholar] [CrossRef]
- Pikus, M.V.; Van Spyk, E.N.; Pham, K.; Geyfman, M.; Kumar, V.; Takahashi, J.S.; Andersen, B. The circadian clock in skin: Implications for adult stem cells, tissue regeneration, cancer, aging, and immunity. J. Biol. Rhythms 2015, 30, 163–182. [Google Scholar] [CrossRef] [Green Version]
- Geyfman, M.; Kumar, V.; Liu, Q.; Ruiz, R.; Gordon, W.; Espitia, F.; Cam, E.; Millar, S.E.; Smyth, P.; Ihler, A.; et al. Brain and nuscle Arnt-like protein-1 (BMAL1) controls circadian cell proliferation and susceptibility to UVB-induced DNA damage in the epidermis. Proc. Natl. Acad. Sci. USA 2012, 109, 11758–11763. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Crair, M.C. How do barrels form in somatosensory cortex? Ann. N. Y. Acad. Sci. 2011, 1225, 119–129. [Google Scholar] [CrossRef]
- Natsubori, A.; Honma, K.; Honma, S. Dual regulation of clock gene Per2 expression in discrete brain areas by the circadian pacemaker and methamphetamine-induced oscillator in rats. Eur. J. Neurosci. 2014, 39, 229–240. [Google Scholar] [CrossRef]
- Akashi, M.; Soma, H.; Yamamoto, T.; Tsugitomi, A.; Yamashita, S.; Yamamoto, T.; Nishida, E.; Yasuda, A.; Liao, J.K.; Node, K. Noninvasive method for assessing the human circadian clock using hair follicle cells. Proc. Natl. Acad. Sci. USA 2010, 107, 15643–15648. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, A.; Matsumura, R.; Matsuzaki, T.; Nakamura, W.; Node, K.; Akashi, M. A simple method using ex vivo culture of hair follicle tissue to investigate intrinsic circadian characteristics in humans. Sci. Rep. 2017, 7, 6824. [Google Scholar] [CrossRef] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, R.; Kanai, M.; Yoshida, Y.; Fukushima, S.; Nogami, M.; Yamaguchi, T.; Iijima, N.; Sutherland, K.; Haga, S.; Ozaki, M.; et al. Analysis of the Anticipatory Behavior Formation Mechanism Induced by Methamphetamine Using a Single Hair. Cells 2023, 12, 654. https://doi.org/10.3390/cells12040654
Sato R, Kanai M, Yoshida Y, Fukushima S, Nogami M, Yamaguchi T, Iijima N, Sutherland K, Haga S, Ozaki M, et al. Analysis of the Anticipatory Behavior Formation Mechanism Induced by Methamphetamine Using a Single Hair. Cells. 2023; 12(4):654. https://doi.org/10.3390/cells12040654
Chicago/Turabian StyleSato, Riku, Megumi Kanai, Yukina Yoshida, Shiori Fukushima, Masahiro Nogami, Takeshi Yamaguchi, Norio Iijima, Kenneth Sutherland, Sanae Haga, Michitaka Ozaki, and et al. 2023. "Analysis of the Anticipatory Behavior Formation Mechanism Induced by Methamphetamine Using a Single Hair" Cells 12, no. 4: 654. https://doi.org/10.3390/cells12040654
APA StyleSato, R., Kanai, M., Yoshida, Y., Fukushima, S., Nogami, M., Yamaguchi, T., Iijima, N., Sutherland, K., Haga, S., Ozaki, M., Hamada, K., & Hamada, T. (2023). Analysis of the Anticipatory Behavior Formation Mechanism Induced by Methamphetamine Using a Single Hair. Cells, 12(4), 654. https://doi.org/10.3390/cells12040654