
Bioinformatic miRNA-mRNAs Analysis Revels to miR-934 as a Potential Regulator of the Epithelial–Mesenchymal Transition in Triple-Negative Breast Cancer


 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression Profiles of miRNAs and mRNAs
2.2. Enrichment Analysis of Pathways and Biological Processes
2.3. Maintenance of the Breast Cancer Cell Line and Transfection of the miR-934 Inhibitor
2.4. RNA Isolation and qRT-PCR Analysis of miR-934 Expression
2.5. Cell Migration Assays
2.6. Statistical Analysis
3. Results
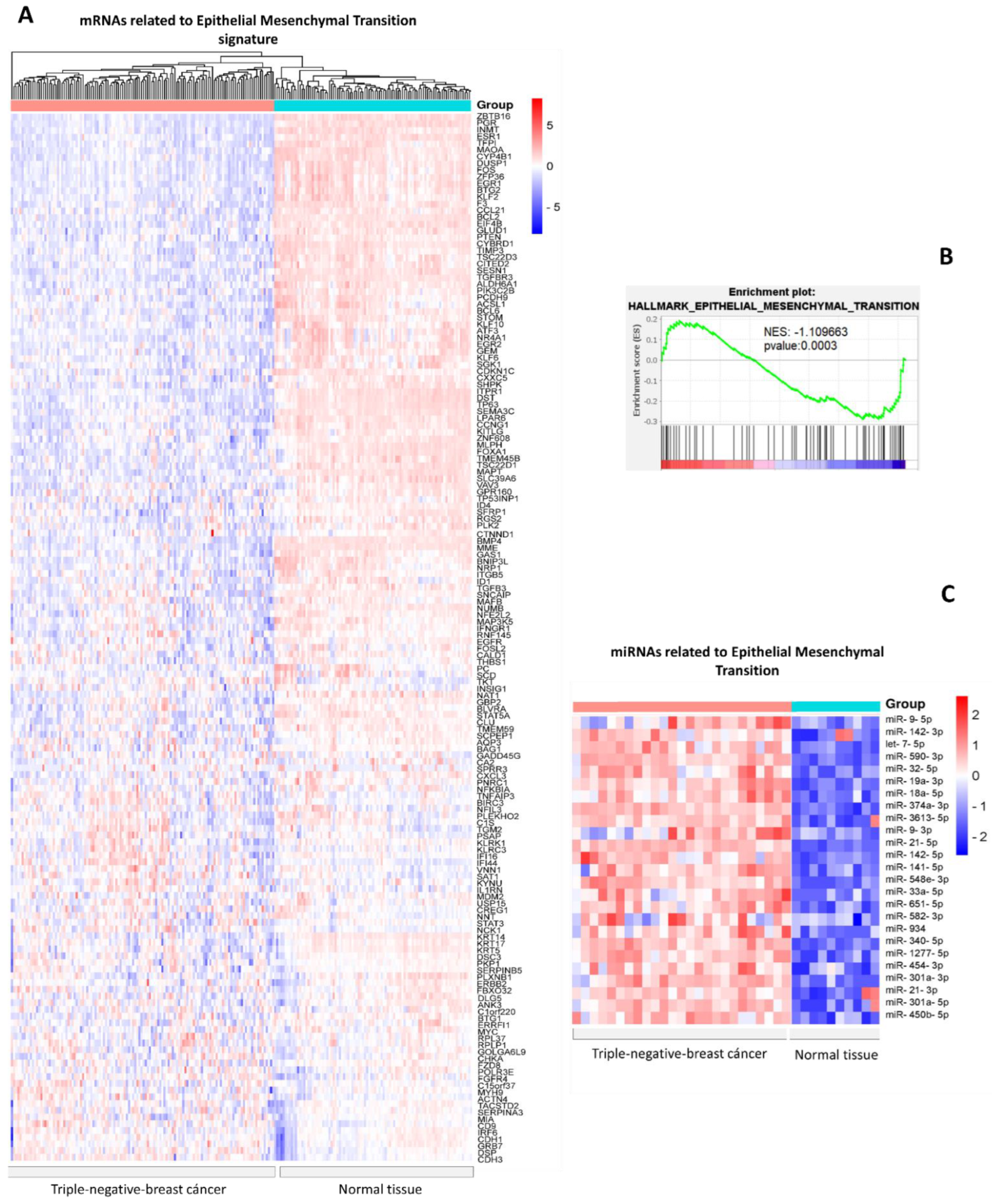
3.1. Identification of Differentially Expressed miRNAs in Triple-Negative Breast Cancer versus Normal Tissues Related to Epithelial–Mesenchymal Transition mRNAs
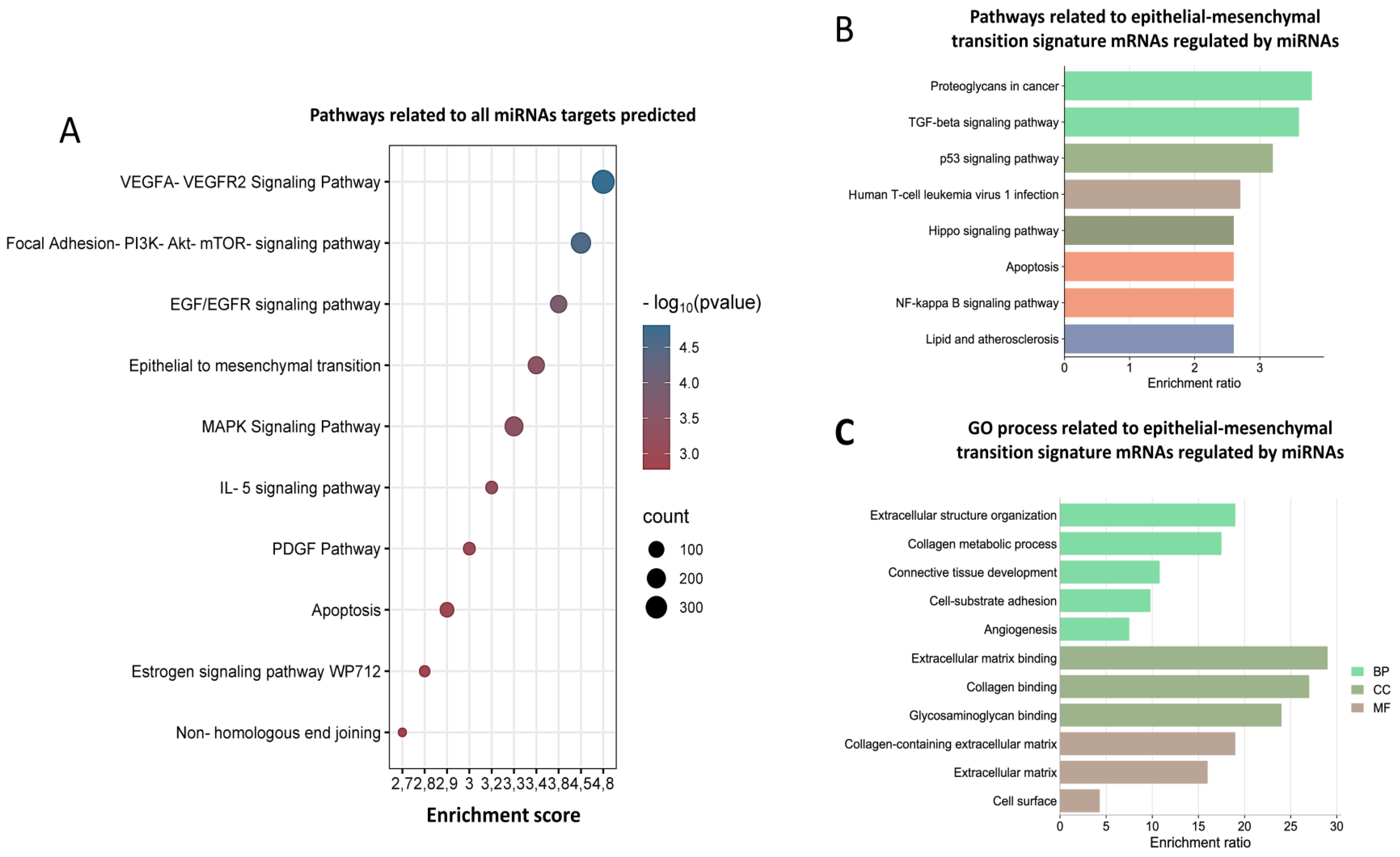
3.2. Biological Processes Regulating miRNAs Related to Epithelial–Mesenchymal Transition
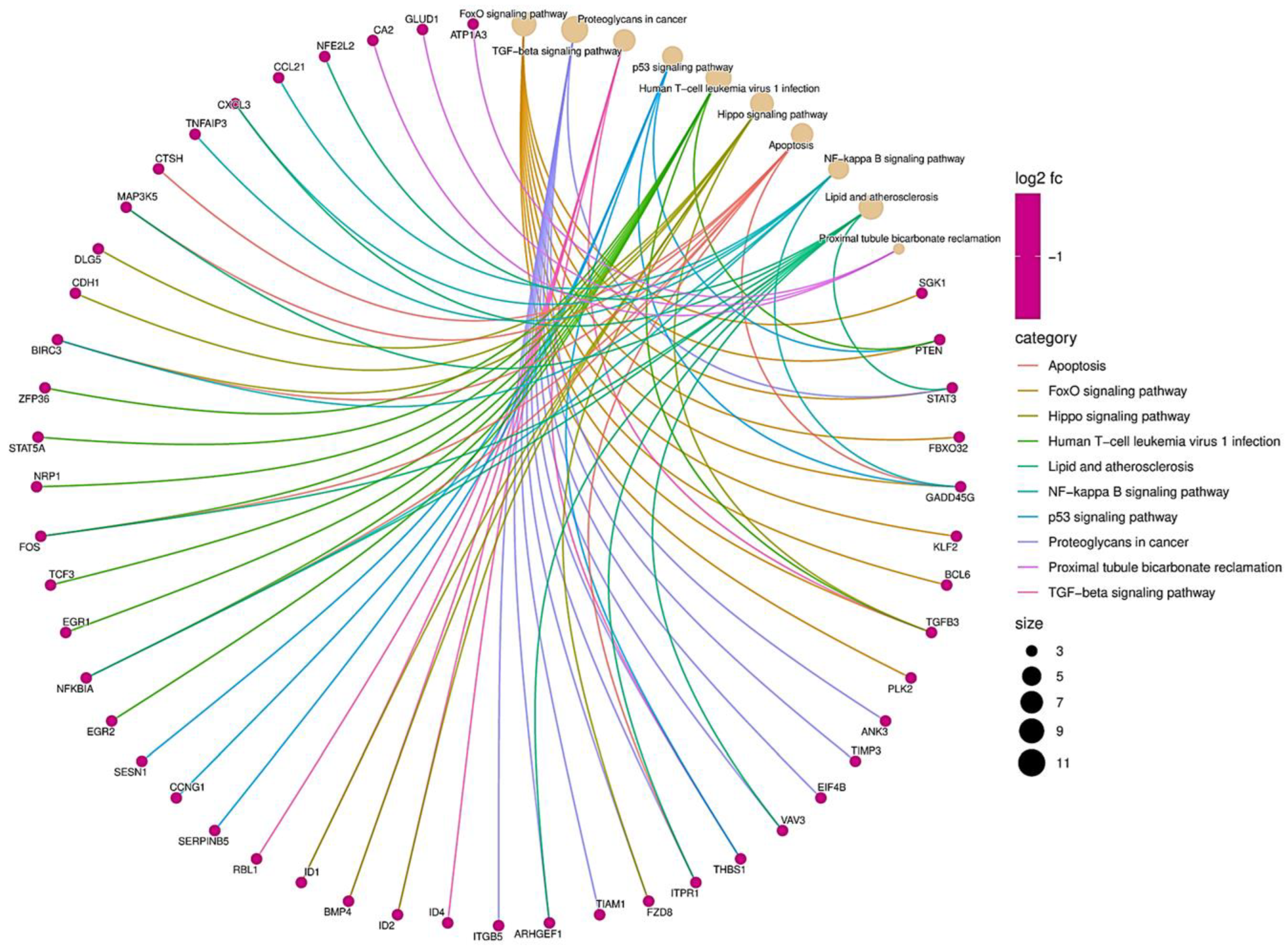
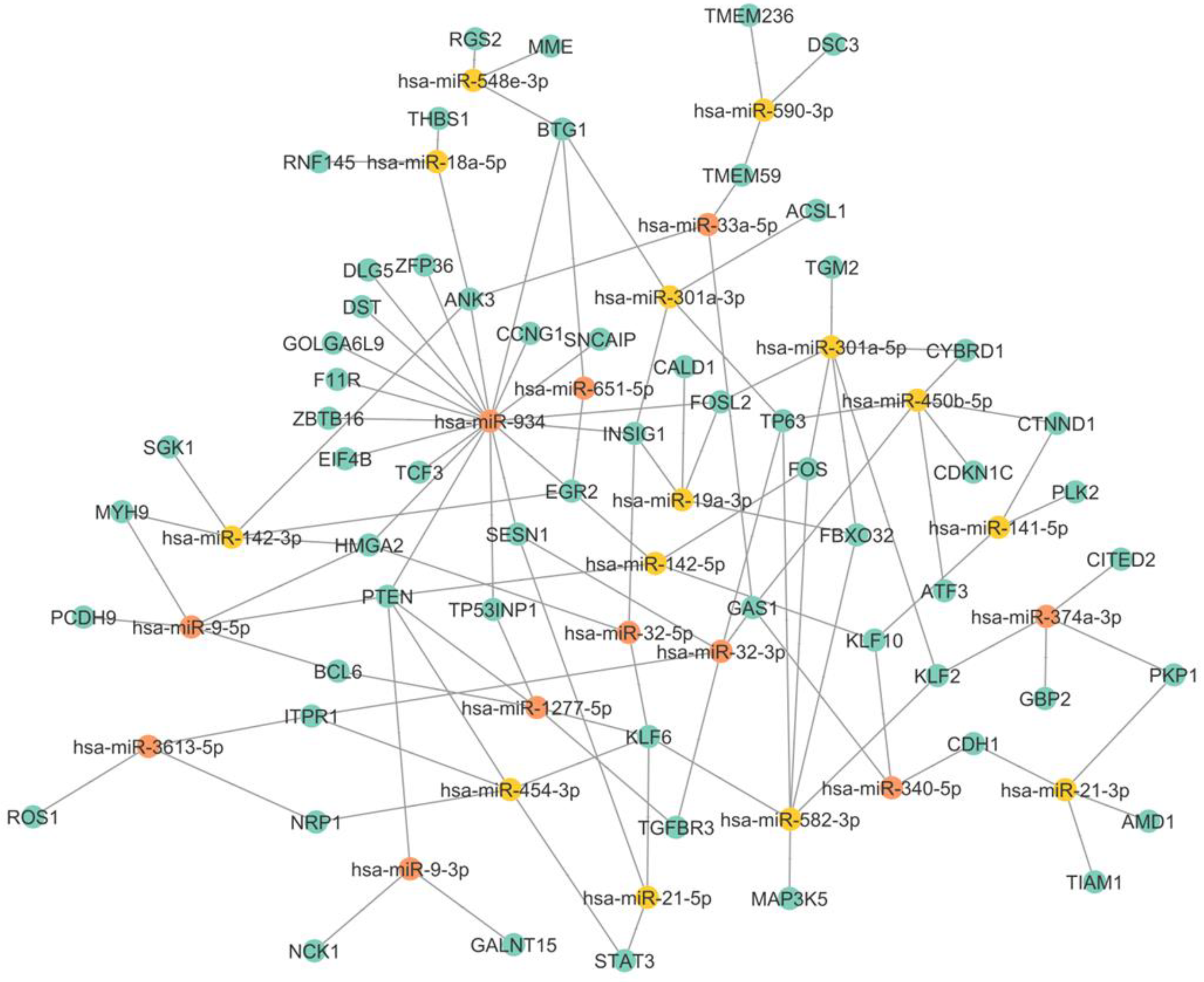
3.3. miRNA–mRNA Interaction Networks Show miR-934 as a Possible Key Regulator of Epithelial–Mesenchymal Transition
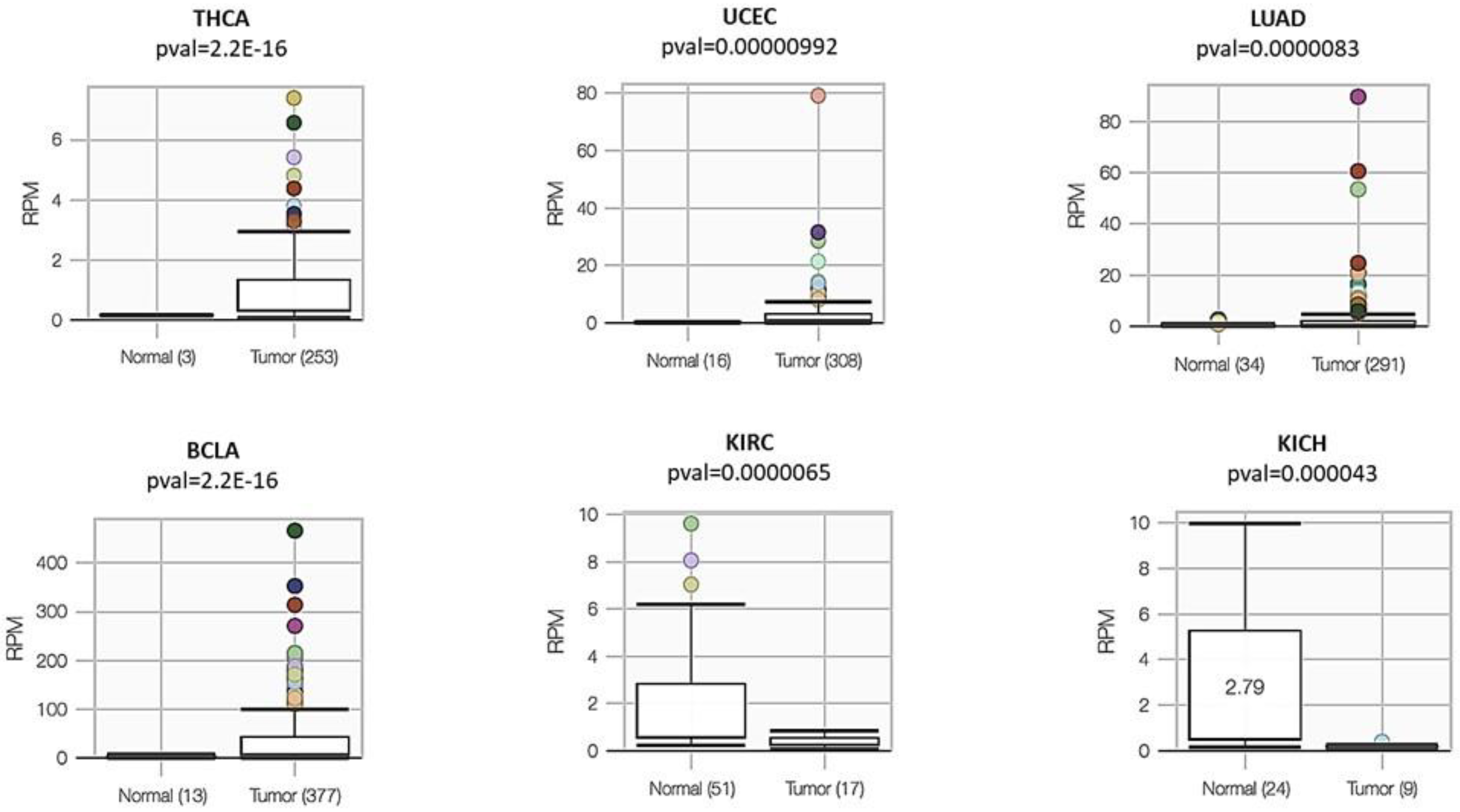
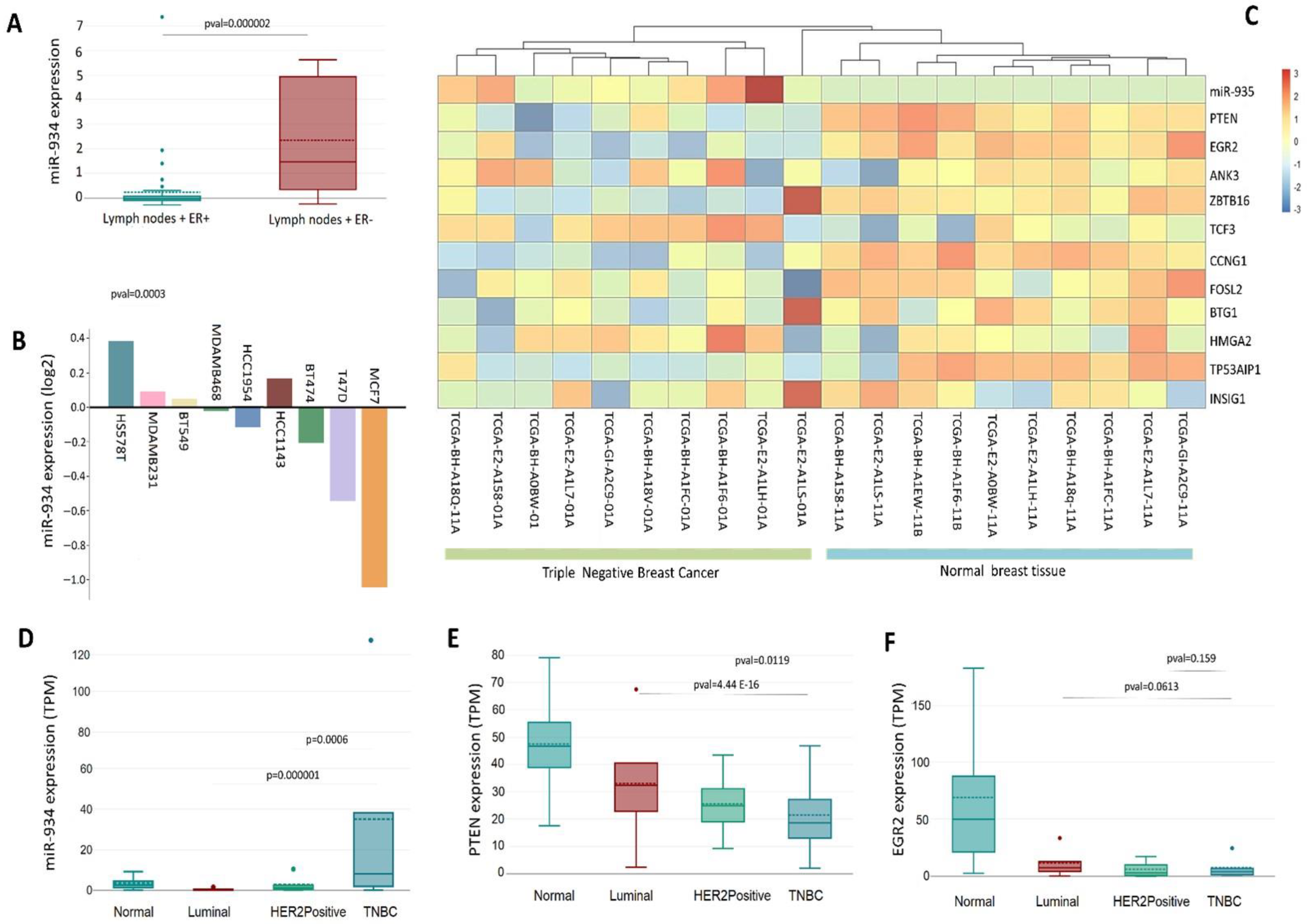
3.4. A Landscape of miR-934 Expression in Different Types of Cancers
3.5. The Upregulation of miR-934 Is Specific to the Molecular Subtype of Triple-Negative Breast Cancer
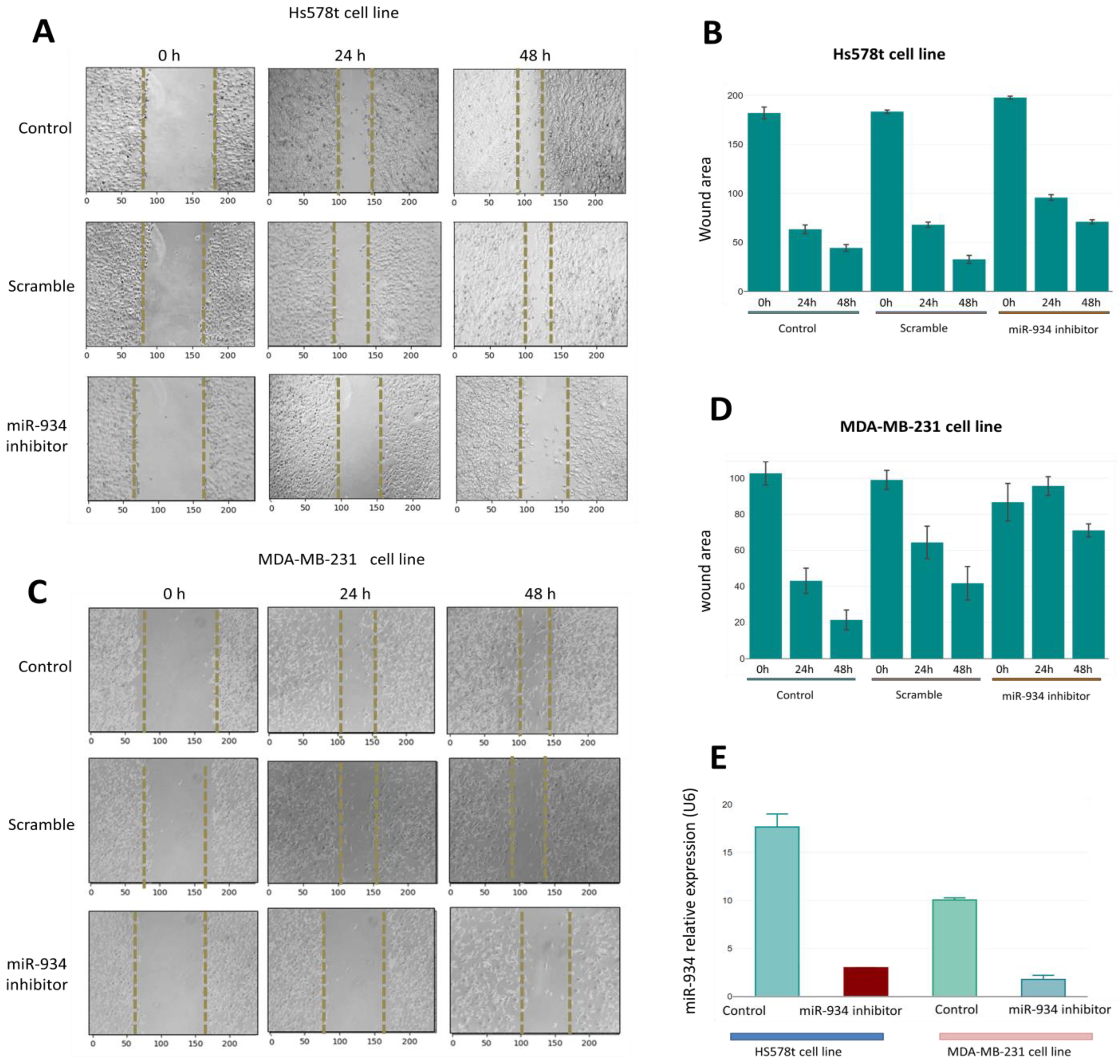
3.6. miR-934 Inhibition Partly Suppresses the Migration Capacity in Triple-Negative Breast Cancer Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J.; Downs, B.M.; Cope, L.M.; Fackler, M.J.; Zhang, X.; Song, C.; VandenBussche, C.; Zhang, K.; Han, Y.; Liu, Y.; et al. Automated and Rapid Detection of Cancer in Suspicious Axillary Lymph Nodes in Patients with Breast Cancer. NPJ Breast Cancer 2021, 7, 89. [Google Scholar] [CrossRef]
- Derakhshan, F.; Reis-Filho, J.S. Pathogenesis of Triple-Negative Breast Cancer. Annu. Rev. Pathol. Mech. Dis. 2022, 17, 181–204. [Google Scholar] [CrossRef]
- Wein, L.; Loi, S. Mechanisms of Resistance of Chemotherapy in Early-Stage Triple Negative Breast Cancer (TNBC). Breast 2017, 34, S27–S30. [Google Scholar] [CrossRef] [PubMed]
- Vijay, G.V.; Zhao, N.; Den Hollander, P.; Toneff, M.J.; Joseph, R.; Pietila, M.; Taube, J.H.; Sarkar, T.R.; Ramirez-Pena, E.; Werden, S.J.; et al. GSK3β Regulates Epithelial-Mesenchymal Transition and Cancer Stem Cell Properties in Triple-Negative Breast Cancer. Breast Cancer Res. 2019, 21, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loibl, S.; Marmé, F.; Martin, M.; Untch, M.; Bonnefoi, H.; Kim, S.-B.; Bear, H.; McCarthy, N.; Melé Olivé, M.; Gelmon, K.; et al. Palbociclib for Residual High-Risk Invasive HR-Positive and HER2-Negative Early Breast Cancer—The Penelope-B Trial. J. Clin. Oncol. 2021, 39, 1518–1530. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New Insights into the Mechanisms of Epithelial–Mesenchymal Transition and Implications for Cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Pastushenko, I.; Blanpain, C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welch, D.R.; Hurst, D.R. Defining the Hallmarks of Metastasis. Cancer Res. 2019, 79, 3011–3027. [Google Scholar] [CrossRef]
- Seo, H.; González-Avalos, E.; Zhang, W.; Ramchandani, P.; Yang, C.; Lio, C.-W.J.; Rao, A.; Hogan, P.G. BATF and IRF4 Cooperate to Counter Exhaustion in Tumor-Infiltrating CAR T Cells. Nat. Immunol. 2021, 22, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Pisarska, M.D.; Bresee, C.; Ida Chen, Y.-D.; Lester, J.; Afshar, Y.; Alexander, C.; Karlan, B.Y. BRCA1 Germline Mutations May Be Associated with Reduced Ovarian Reserve. Fertil. Steril. 2014, 102, 1723–1728. [Google Scholar] [CrossRef] [Green Version]
- Martinez, O.; Sosabowski, J.; Maher, J.; Papa, S. New Developments in Imaging Cell-Based Therapy. J. Nucl. Med. 2019, 60, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Prvanović, M.; Nedeljković, M.; Tanić, N.; Tomić, T.; Terzić, T.; Milovanović, Z.; Maksimović, Z.; Tanić, N. Role of PTEN, PI3K, and MTOR in Triple-Negative Breast Cancer. Life 2021, 11, 1247. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, O.; Parfenyev, S.; Daks, A.; Shuvalov, O.; Barlev, N.A. The Role of PTEN in Epithelial–Mesenchymal Transition. Cancers 2022, 14, 3786. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.-S.; Huang, H.-T.; Qiu, J.; Sun, J.; Ge, R.-F.; Jiang, L.-W. MiR-224-5p Targets EGR2 to Promote the Development of Papillary Thyroid Carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4890–4900. [Google Scholar] [CrossRef]
- Lu, Y.; Hu, X.; Yang, X. MiR-934 Promotes Breast Cancer Metastasis by Regulation of PTEN and Epithelial–Mesenchymal Transition. Tissue Cell 2021, 71, 101581. [Google Scholar] [CrossRef]
- Muñoz-Rodríguez, J.L.; Vrba, L.; Futscher, B.W.; Hu, C.; Komenaka, I.K.; Meza-Montenegro, M.M.; Gutierrez-Millan, L.E.; Daneri-Navarro, A.; Thompson, P.A.; Martinez, M.E. Differentially Expressed MicroRNAs in Postpartum Breast Cancer in Hispanic Women. PLoS ONE 2015, 10, e0124340. [Google Scholar] [CrossRef] [Green Version]
- Castilla, M.Á.; López-García, M.Á.; Atienza, M.R.; Rosa-Rosa, J.M.; Díaz-Martín, J.; Pecero, M.L.; Vieites, B.; Romero-Pérez, L.; Benítez, J.; Calcabrini, A.; et al. VGLL1 Expression Is Associated with a Triple-Negative Basal-like Phenotype in Breast Cancer. Endocr. Relat. Cancer 2014, 21, 587–599. [Google Scholar] [CrossRef] [Green Version]
- Sarrió, D.; Rodriguez-Pinilla, S.M.; Hardisson, D.; Cano, A.; Moreno-Bueno, G.; Palacios, J. Epithelial-Mesenchymal Transition in Breast Cancer Relates to the Basal-like Phenotype. Cancer Res. 2008, 68, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Jechlinger, M.; Grunert, S.; Tamir, I.H.; Janda, E.; Lüdemann, S.; Waerner, T.; Seither, P.; Weith, A.; Beug, H.; Kraut, N. Expression Profiling of Epithelial Plasticity in Tumor Progression. Oncogene 2003, 22, 7155–7169. [Google Scholar] [CrossRef] [Green Version]
- Chin, K.; DeVries, S.; Fridlyand, J.; Spellman, P.T.; Roydasgupta, R.; Kuo, W.-L.; Lapuk, A.; Neve, R.M.; Qian, Z.; Ryder, T.; et al. Genomic and Transcriptional Aberrations Linked to Breast Cancer Pathophysiologies. Cancer Cell 2006, 10, 529–541. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Washington, M.K.; Crawford, H.C. Loss of FOXA1/2 Is Essential for the Epithelial-to-Mesenchymal Transition in Pancreatic Cancer. Cancer Res. 2010, 70, 2115–2125. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Chen, Y.; Mo, C.; Wang, D.; Chen, J.; Mao, X.; Guo, S.; Zhuang, J.; Qiu, S. Association of DSC3 MRNA Down-Regulation in Prostate Cancer with Promoter Hypermethylation and Poor Prognosis. PLoS ONE 2014, 9, e92815. [Google Scholar] [CrossRef]
- Sahab, Z.J.; Hall, M.D.; Me Sung, Y.; Dakshanamurthy, S.; Ji, Y.; Kumar, D.; Byers, S.W. Tumor Suppressor RARRES1 Interacts with Cytoplasmic Carboxypeptidase AGBL2 to Regulate the α-Tubulin Tyrosination Cycle. Cancer Res. 2011, 71, 1219–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Liu, J.; Chao, J.; Scheuerman, M.P.; Rahimi, S.A.; Lee, L.Y.; Li, S. PTEN Suppresses Epithelial–Mesenchymal Transition and Cancer Stem Cell Activity by Downregulating Abi1. Sci. Rep. 2020, 10, 12685. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.R.; Oyan, A.M.; Rostad, K.; Hellem, M.R.; Liu, J.; Li, L.; Micklem, D.R.; Haugen, H.; Lorens, J.B.; Rotter, V.; et al. P63 Attenuates Epithelial to Mesenchymal Potential in an Experimental Prostate Cell Model. PLoS ONE 2013, 8, e62547. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, H.; Zhang, D.; Fan, D.; Zhang, Y.; Dong, X.; Liu, S.; Yang, Z.; Ni, C.; Li, Y.; et al. TP 53 INP 1 Inhibits Hypoxia-induced Vasculogenic Mimicry Formation via the ROS /Snail Signalling Axis in Breast Cancer. J. Cell. Mol. Med. 2018, 22, 3475–3488. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Zhang, Y.; Chen, Y.; Liu, P.; An, T.; Zhang, J.; Yang, H.; Zhu, W.; Yang, X. FOXO1 Inhibits the Invasion and Metastasis of Hepatocellular Carcinoma by Reversing ZEB2-Induced Epithelial-Mesenchymal Transition. Oncotarget 2017, 8, 1703–1713. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.; Kremerskothen, J.; Lin, X.; Zhang, X.; Rong, X.; Zhang, D.; Wang, E. WWC3 Inhibits Epithelial–Mesenchymal Transition of Lung Cancer by Activating Hippo-YAP Signaling. Onco. Targets. Ther. 2018, 11, 2581–2591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.; Veronese, A.; Pichiorri, F.; Lee, T.J.; Jeon, Y.-J.; Volinia, S.; Pineau, P.; Marchio, A.; Palatini, J.; Suh, S.-S.; et al. P53 Regulates Epithelial–Mesenchymal Transition through MicroRNAs Targeting ZEB1 and ZEB2. J. Exp. Med. 2011, 208, 875–883. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.; Hu, B.; Ma, X.; Yang, Z.; Yu, M.; Sun, H.; Huang, A.; Zhang, X.; Wang, J.; Hu, Z.; et al. New Insight into BIRC3: A Novel Prognostic Indicator and a Potential Therapeutic Target for Liver Cancer. J. Cell. Biochem. 2019, 120, 6035–6045. [Google Scholar] [CrossRef]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.-P.; Tong, F.; et al. A Collection of Breast Cancer Cell Lines for the Study of Functionally Distinct Cancer Subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorling, L.; Carvalho, S.; Allen, J.; González-Neira, A.; Luccarini, C.; Wahlström, C.; Pooley, K.A.; Parsons, M.T.; Fortuno, C.; Wang, Q.; et al. Breast Cancer Risk Genes—Association Analysis in More than 113,000 Women. N. Engl. J. Med. 2021, 384, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Huber, S.; Schramm, C.; Lehr, H.A.; Mann, A.; Schmitt, S.; Becker, C.; Protschka, M.; Galle, P.R.; Neurath, M.F.; Blessing, M. Cutting Edge: TGF-β Signaling Is Required for the In Vivo Expansion and Immunosuppressive Capacity of Regulatory CD4 + CD25 + T Cells. J. Immunol. 2004, 173, 6526–6531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akrida, I.; Bravou, V.; Papadaki, H. The Deadly Cross-Talk between Hippo Pathway and Epithelial–Mesenchymal Transition (EMT) in Cancer. Mol. Biol. Rep. 2022, 49, 10065–10076. [Google Scholar] [CrossRef]
- Huo, L.; Wang, Y.; Gong, Y.; Krishnamurthy, S.; Wang, J.; Diao, L.; Liu, C.-G.; Liu, X.; Lin, F.; Symmans, W.F.; et al. MicroRNA Expression Profiling Identifies Decreased Expression of MiR-205 in Inflammatory Breast Cancer. Mod. Pathol. 2016, 29, 330–346. [Google Scholar] [CrossRef] [Green Version]
- Søkilde, R.; Persson, H.; Ehinger, A.; Pirona, A.C.; Fernö, M.; Hegardt, C.; Larsson, C.; Loman, N.; Malmberg, M.; Rydén, L.; et al. Refinement of Breast Cancer Molecular Classification by MiRNA Expression Profiles. BMC Genom. 2019, 20, 503. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras-Rodríguez, J.A.; Puente-Rivera, J.; Córdova-Esparza, D.M.; Nuñez-Olvera, S.I.; Silva-Cázares, M.B. Bioinformatic miRNA-mRNAs Analysis Revels to miR-934 as a Potential Regulator of the Epithelial–Mesenchymal Transition in Triple-Negative Breast Cancer. Cells 2023, 12, 834. https://doi.org/10.3390/cells12060834
Contreras-Rodríguez JA, Puente-Rivera J, Córdova-Esparza DM, Nuñez-Olvera SI, Silva-Cázares MB. Bioinformatic miRNA-mRNAs Analysis Revels to miR-934 as a Potential Regulator of the Epithelial–Mesenchymal Transition in Triple-Negative Breast Cancer. Cells. 2023; 12(6):834. https://doi.org/10.3390/cells12060834
Chicago/Turabian StyleContreras-Rodríguez, Jorge Alberto, Jonathan Puente-Rivera, Diana Margarita Córdova-Esparza, Stephanie I. Nuñez-Olvera, and Macrina Beatriz Silva-Cázares. 2023. "Bioinformatic miRNA-mRNAs Analysis Revels to miR-934 as a Potential Regulator of the Epithelial–Mesenchymal Transition in Triple-Negative Breast Cancer" Cells 12, no. 6: 834. https://doi.org/10.3390/cells12060834
APA StyleContreras-Rodríguez, J. A., Puente-Rivera, J., Córdova-Esparza, D. M., Nuñez-Olvera, S. I., & Silva-Cázares, M. B. (2023). Bioinformatic miRNA-mRNAs Analysis Revels to miR-934 as a Potential Regulator of the Epithelial–Mesenchymal Transition in Triple-Negative Breast Cancer. Cells, 12(6), 834. https://doi.org/10.3390/cells12060834