
N-Glycosylation of LRP6 by B3GnT2 Promotes Wnt/β-Catenin Signalling
 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Plasmids
2.2. Medaka (Oryzias latipes) cDNA Library Screening for LRP6 Modifiers
2.3. Cell Culture, Cell Transfection
2.4. TOPFLASH Reporter Assay
2.5. Western Blot, Immunoprecipitation (IP), Lectin Blot
2.6. RNA Extraction, RT-PCR, Semi-qPCR
2.7. Immunofluorescence Analysis
2.8. Mass Spectrometry Analysis
2.9. Cell Surface LRP6 Nano-BiT Assay
2.10. Lentiviral-Mediated B3GnT2 Gene Knockout
2.10.1. sgRNA Design for Inactivation of B3GnT2 in HEK293T Cells
2.10.2. Cloning Lenti-CRISPRV2-B3GnT2-sgRNA Plasmid
2.10.3. Infecting HEK 293T Target Cells with LentiCRISPR-B3GnT2-sgRNA Lentivirus
2.10.4. Generation of Single-Cell Clones and Genotyping
2.10.5. Sanger Sequencing of B3GnT2 Gene KO Cell Clones
3. Results
3.1. Identification of Medaka B3GnT2l as A Novel LRP6 Modifier
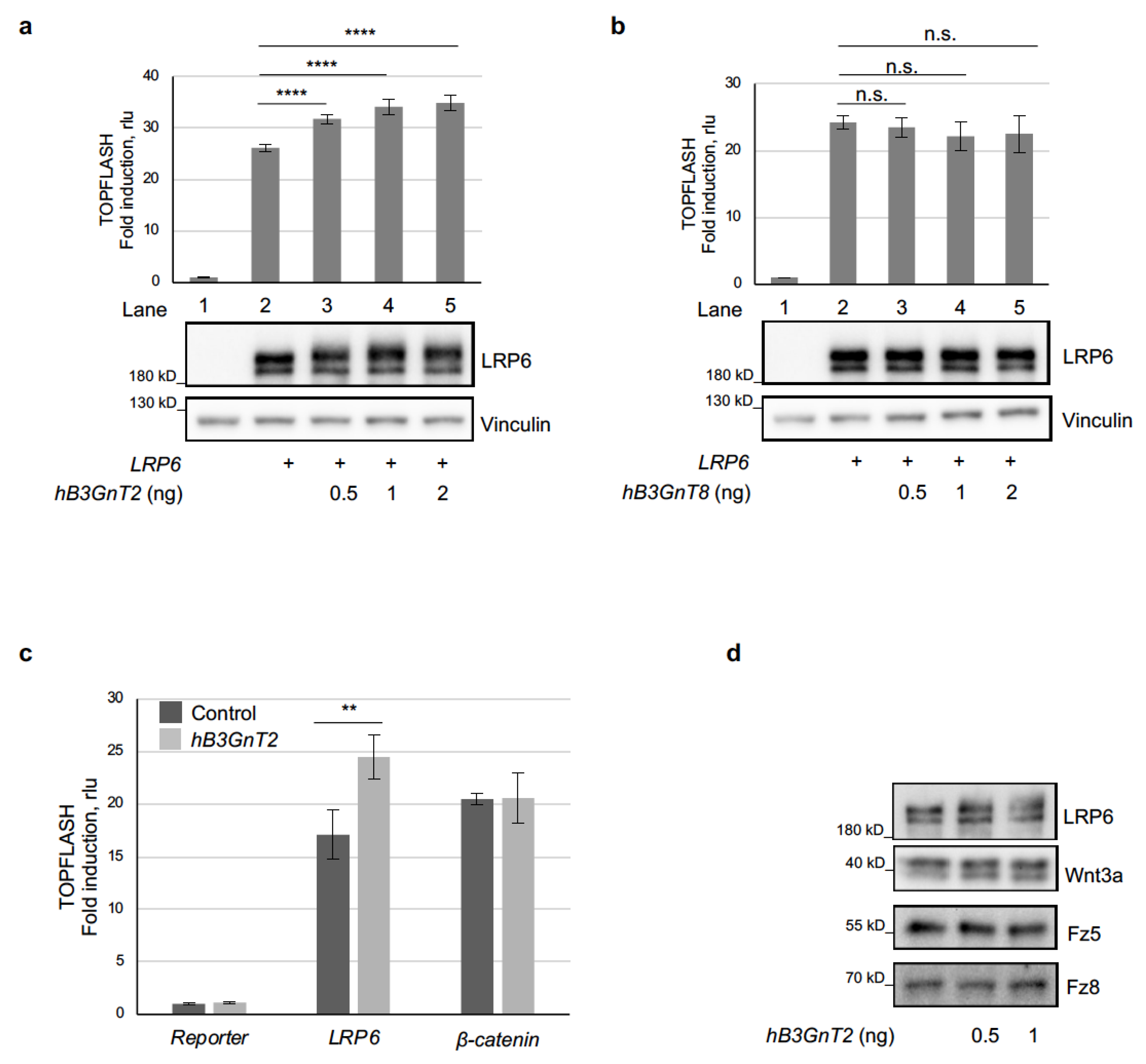
3.2. Human B3GnT2 Promotes Wnt/β-Catenin Signalling and Acts at the Level of LRP6
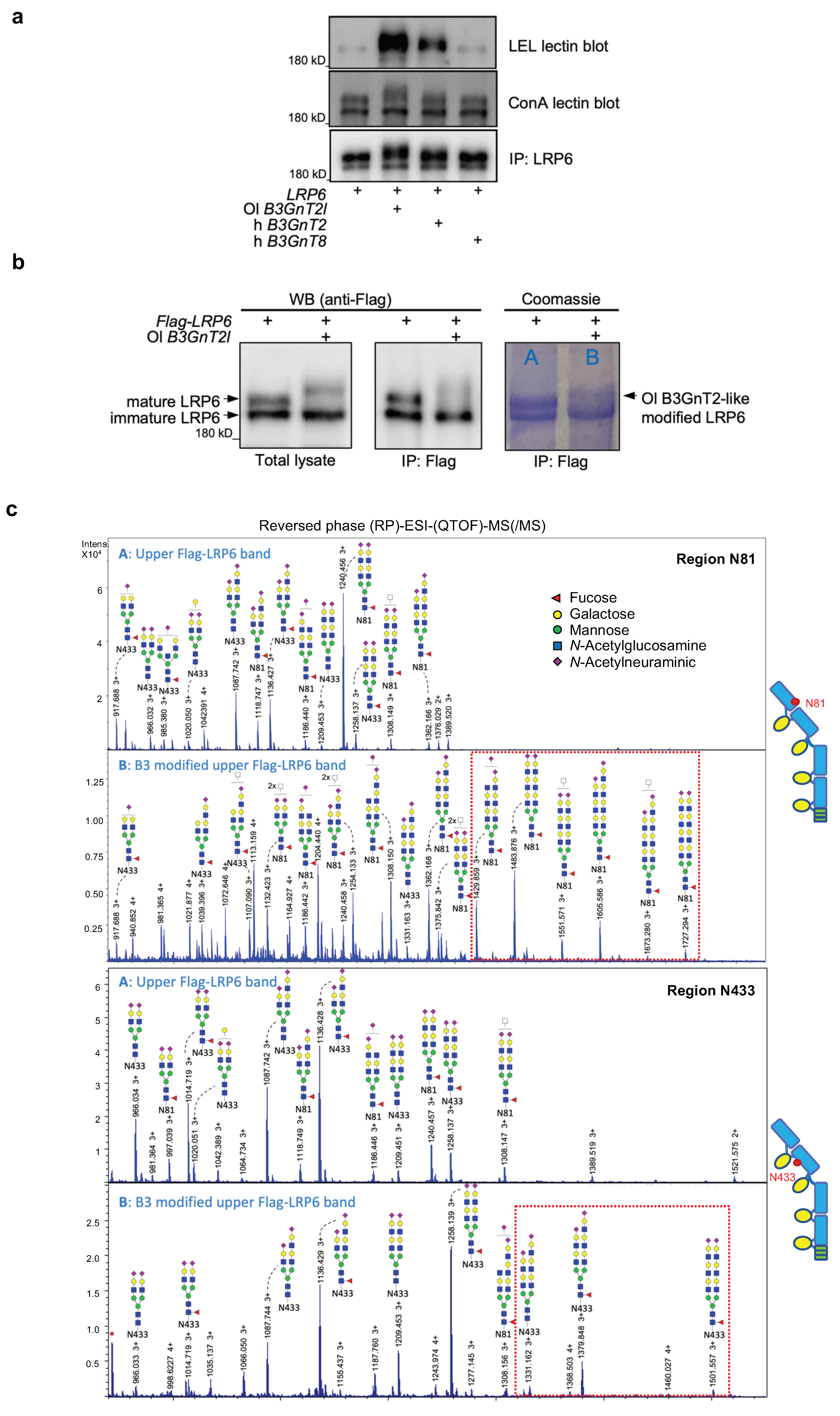
3.3. B3GnT2 Extends Polylactosamine Chains of LRP6 N-glycans
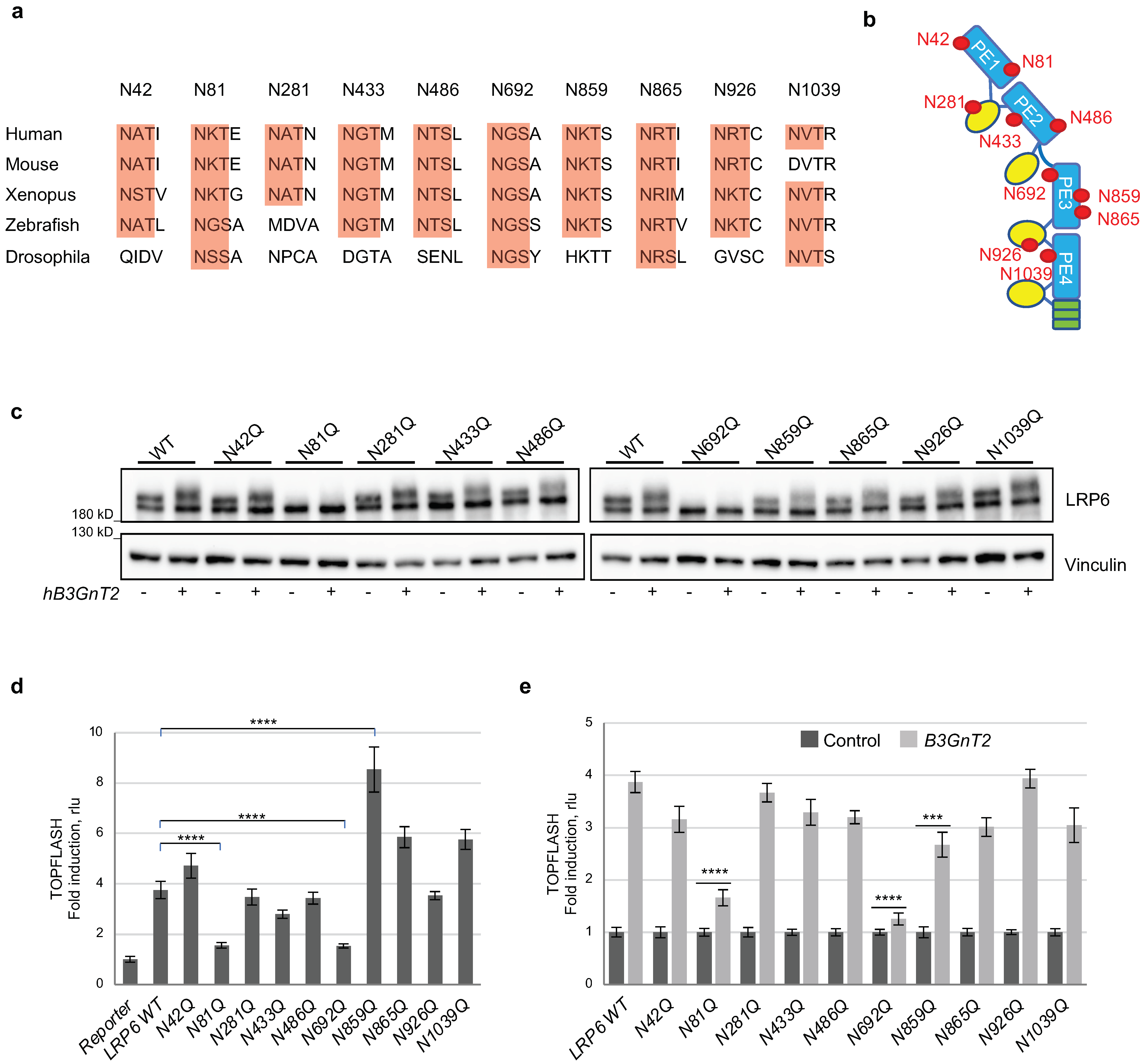
3.4. Multiple N-glycan Sites of LRP6 Are Relevant for B3GnT2-Mediated LRP6 Modification
3.5. B3GnT2 Promotes Cell Surface Levels of LRP6
3.6. Human B3GnT2 Is Required for Wnt/β-Catenin Signalling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niehrs, C. The complex world of WNT receptor signalling. Nat. Rev. Mol. Cell Biol. 2012, 13, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Wiese, K.E.; Nusse, R.; van Amerongen, R. Wnt signalling: Conquering complexity. Development 2018, 145, 12. [Google Scholar] [CrossRef] [Green Version]
- Holstein, T.W. The evolution of the Wnt pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a007922. [Google Scholar] [CrossRef]
- Acebron, S.P.; Niehrs, C. Beta-Catenin-Independent Roles of Wnt/LRP6 Signaling. Trends Cell Biol. 2016, 26, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Koca, Y.; Collu, G.M.; Mlodzik, M. Wnt-frizzled planar cell polarity signaling in the regulation of cell motility. Curr. Top. Dev. Biol. 2022, 150, 255–297. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.J.; Acebron, S.P. Wnt signalling in cell division: From mechanisms to tissue engineering. Trends Cell Biol. 2022, 32, 1035–1048. [Google Scholar] [CrossRef]
- Tejeda-Munoz, N.; De Robertis, E.M. Wnt, GSK3, and Macropinocytosis. Subcell Biochem. 2022, 98, 169–187. [Google Scholar] [CrossRef]
- Davidson, G. LRPs in WNT Signalling. Handb. Exp. Pharmacol. 2021, 269, 45–73. [Google Scholar] [CrossRef]
- Niehrs, C. Function and biological roles of the Dickkopf family of Wnt modulators. Oncogene 2006, 25, 7469–7481. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Semenov, M.; Tamai, K.; Zeng, X. LDL receptor-related proteins 5 and 6 in Wnt/beta-catenin signaling: Arrows point the way. Development 2004, 131, 1663–1677. [Google Scholar] [CrossRef] [Green Version]
- Matoba, K.; Mihara, E.; Tamura-Kawakami, K.; Miyazaki, N.; Maeda, S.; Hirai, H.; Thompson, S.; Iwasaki, K.; Takagi, J. Conformational Freedom of the LRP6 Ectodomain Is Regulated by N-glycosylation and the Binding of the Wnt Antagonist Dkk1. Cell Rep. 2017, 18, 32–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Biechele, T.; Wei, Z.; Morrone, S.; Moon, R.T.; Wang, L.; Xu, W. Crystal structures of the extracellular domain of LRP6 and its complex with DKK1. Nat. Struct. Mol. Biol. 2011, 18, 1204–1210. [Google Scholar] [CrossRef] [PubMed]
- Joiner, D.M.; Ke, J.; Zhong, Z.; Xu, H.E.; Williams, B.O. LRP5 and LRP6 in development and disease. Trends Endocrinol. Metab. 2013, 24, 31–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, V.E.; Chu, M.L.; Choi, H.J.; Tran, D.; Abo, A.; Weis, W.I. Structural basis of Wnt signaling inhibition by Dickkopf binding to LRP5/6. Dev. Cell 2011, 21, 862–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ettenberg, S.A.; Charlat, O.; Daley, M.P.; Liu, S.; Vincent, K.J.; Stuart, D.D.; Schuller, A.G.; Yuan, J.; Ospina, B.; Green, J.; et al. Inhibition of tumorigenesis driven by different Wnt proteins requires blockade of distinct ligand-binding regions by LRP6 antibodies. Proc. Natl. Acad. Sci. USA 2010, 107, 15473–15478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.; Tamai, K.; Doble, B.; Li, S.; Huang, H.; Habas, R.; Okamura, H.; Woodgett, J.; He, X. A dual-kinase mechanism for Wnt co-receptor phosphorylation and activation. Nature 2005, 438, 873–877. [Google Scholar] [CrossRef] [Green Version]
- Davidson, G.; Wu, W.; Shen, J.; Bilic, J.; Fenger, U.; Stannek, P.; Glinka, A.; Niehrs, C. Casein kinase 1 gamma couples Wnt receptor activation to cytoplasmic signal transduction. Nature 2005, 438, 867–872. [Google Scholar] [CrossRef]
- Piao, S.; Lee, S.H.; Kim, H.; Yum, S.; Stamos, J.L.; Xu, Y.; Lee, S.J.; Lee, J.; Oh, S.; Han, J.K.; et al. Direct inhibition of GSK3beta by the phosphorylated cytoplasmic domain of LRP6 in Wnt/beta-catenin signaling. PLoS ONE 2008, 3, e4046. [Google Scholar] [CrossRef] [Green Version]
- Tamai, K.; Zeng, X.; Liu, C.; Zhang, X.; Harada, Y.; Chang, Z.; He, X. A mechanism for Wnt coreceptor activation. Mol. Cell 2004, 13, 149–156. [Google Scholar] [CrossRef]
- Hsieh, J.C.; Lee, L.; Zhang, L.; Wefer, S.; Brown, K.; DeRossi, C.; Wines, M.E.; Rosenquist, T.; Holdener, B.C. Mesd encodes an LRP5/6 chaperone essential for specification of mouse embryonic polarity. Cell 2003, 112, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Culi, J.; Mann, R.S. Boca, an endoplasmic reticulum protein required for wingless signaling and trafficking of LDL receptor family members in Drosophila. Cell 2003, 112, 343–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culi, J.; Springer, T.A.; Mann, R.S. Boca-dependent maturation of beta-propeller/EGF modules in low-density lipoprotein receptor proteins. EMBO J. 2004, 23, 1372–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Bubeck, D.; MacDonald, B.T.; Liang, W.X.; Mao, J.H.; Malinauskas, T.; Llorca, O.; Aricescu, A.R.; Siebold, C.; He, X.; et al. Structural and functional studies of LRP6 ectodomain reveal a platform for Wnt signaling. Dev. Cell 2011, 21, 848–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrami, L.; Kunz, B.; Iacovache, I.; van der Goot, F.G. Palmitoylation and ubiquitination regulate exit of the Wnt signaling protein LRP6 from the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2008, 105, 5384–5389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, H.; Lee, S.K.; Jho, E.H. Mest/Peg1 inhibits Wnt signalling through regulation of LRP6 glycosylation. Biochem. J. 2011, 436, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Vijayakumar, S.; de la Torre, T.V.; Rotolo, S.; Bafico, A. Analysis of endogenous LRP6 function reveals a novel feedback mechanism by which Wnt negatively regulates its receptor. Mol. Cell Biol. 2007, 27, 7291–7301. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wang, B.; Su, Y.; He, D.; Mo, H.; Zheng, M.; Meng, Z.; Ren, L.; Zhang, X.; Ren, D.; et al. ALG8 Fuels Stemness Through Glycosylation of the WNT/Beta-Catenin Signaling Pathway in Colon Cancer. DNA Cell Biol. 2022, 41, 1075–1083. [Google Scholar] [CrossRef]
- Hong, S.; Feng, L.; Yang, Y.; Jiang, H.; Hou, X.; Guo, P.; Marlow, F.L.; Stanley, P.; Wu, P. In Situ Fucosylation of the Wnt Co-receptor LRP6 Increases Its Endocytosis and Reduces Wnt/beta-Catenin Signaling. Cell Chem. Biol. 2020, 27, 1140–1150.e1144. [Google Scholar] [CrossRef]
- Jeong, W.; Kim, S.; Lee, U.; Zhong, Z.A.; Savitsky, M.; Kwon, H.; Kim, J.; Lee, T.; Cho, J.W.; Williams, B.O.; et al. LDL receptor-related protein LRP6 senses nutrient levels and regulates Hippo signaling. EMBO Rep. 2020, 21, e50103. [Google Scholar] [CrossRef]
- Hirata, T.; Kizuka, Y. N-Glycosylation. Adv. Exp. Med. Biol. 2021, 1325, 3–24. [Google Scholar] [CrossRef]
- Narimatsu, H. Human glycogene cloning: Focus on beta 3-glycosyltransferase and beta 4-glycosyltransferase families. Curr. Opin. Struct. Biol. 2006, 16, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.L.; Kohler, J.J.; Pfeffer, S.R. Association of beta-1,3-N-acetylglucosaminyltransferase 1 and beta-1,4-galactosyltransferase 1, trans-Golgi enzymes involved in coupled poly-N-acetyllactosamine synthesis. Glycobiology 2009, 19, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Kadirvelraj, R.; Yang, J.Y.; Kim, H.W.; Sanders, J.H.; Moremen, K.W.; Wood, Z.A. Comparison of human poly-N-acetyl-lactosamine synthase structure with GT-A fold glycosyltransferases supports a modular assembly of catalytic subsites. J. Biol. Chem. 2021, 296, 100110. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Crequer-Grandhomme, A.; Javier, N.; Singh, A.; Chen, H.; Manzanillo, P.; Lo, M.C.; Huang, X. Structures and mechanism of human glycosyltransferase beta1,3-N-acetylglucosaminyltransferase 2 (B3GNT2), an important player in immune homeostasis. J. Biol. Chem. 2021, 296, 100042. [Google Scholar] [CrossRef] [PubMed]
- Venkitachalam, S.; Revoredo, L.; Varadan, V.; Fecteau, R.E.; Ravi, L.; Lutterbaugh, J.; Markowitz, S.D.; Willis, J.E.; Gerken, T.A.; Guda, K. Biochemical and functional characterization of glycosylation-associated mutational landscapes in colon cancer. Sci. Rep. 2016, 6, 23642. [Google Scholar] [CrossRef] [Green Version]
- Gromova, I.; Gromov, P.; Celis, J.E. A novel member of the glycosyltransferase family, beta 3 Gn-T2, highly downregulated in invasive human bladder transitional cell carcinomas. Mol. Carcinog. 2001, 32, 61–72. [Google Scholar] [CrossRef]
- Togayachi, A.; Kozono, Y.; Ishida, H.; Abe, S.; Suzuki, N.; Tsunoda, Y.; Hagiwara, K.; Kuno, A.; Ohkura, T.; Sato, N.; et al. Polylactosamine on glycoproteins influences basal levels of lymphocyte and macrophage activation. Proc. Natl. Acad. Sci. USA 2007, 104, 15829–15834. [Google Scholar] [CrossRef] [Green Version]
- Tsoi, L.C.; Spain, S.L.; Knight, J.; Ellinghaus, E.; Stuart, P.E.; Capon, F.; Ding, J.; Li, Y.; Tejasvi, T.; Gudjonsson, J.E.; et al. Identification of 15 new psoriasis susceptibility loci highlights the role of innate immunity. Nat. Genet. 2012, 44, 1341–1348. [Google Scholar] [CrossRef] [Green Version]
- Okada, Y.; Terao, C.; Ikari, K.; Kochi, Y.; Ohmura, K.; Suzuki, A.; Kawaguchi, T.; Stahl, E.A.; Kurreeman, F.A.; Nishida, N.; et al. Meta-analysis identifies nine new loci associated with rheumatoid arthritis in the Japanese population. Nat. Genet. 2012, 44, 511–516. [Google Scholar] [CrossRef]
- Australo-Anglo-American Spondyloarthritis, C.; Reveille, J.D.; Sims, A.M.; Danoy, P.; Evans, D.M.; Leo, P.; Pointon, J.J.; Jin, R.; Zhou, X.; Bradbury, L.A.; et al. Genome-wide association study of ankylosing spondylitis identifies non-MHC susceptibility loci. Nat. Genet. 2010, 42, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Joung, J.; Kirchgatterer, P.C.; Singh, A.; Cho, J.H.; Nety, S.P.; Larson, R.C.; Macrae, R.K.; Deasy, R.; Tseng, Y.Y.; Maus, M.V.; et al. CRISPR activation screen identifies BCL-2 proteins and B3GNT2 as drivers of cancer resistance to T cell-mediated cytotoxicity. Nat. Commun. 2022, 13, 1606. [Google Scholar] [CrossRef] [PubMed]
- Eckert, A.F.; Gao, P.; Wesslowski, J.; Wang, X.; Rath, J.; Nienhaus, K.; Davidson, G.; Nienhaus, G.U. Measuring ligand-cell surface receptor affinities with axial line-scanning fluorescence correlation spectroscopy. Elife 2020, 9, e55286. [Google Scholar] [CrossRef] [PubMed]
- Souren, M.; Martinez-Morales, J.R.; Makri, P.; Wittbrodt, B.; Wittbrodt, J. A global survey identifies novel upstream components of the Ath5 neurogenic network. Genome Biol. 2009, 10, R92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Su, Y.; Wesslowski, J.; Hagemann, A.I.; Ramialison, M.; Wittbrodt, J.; Scholpp, S.; Davidson, G. Tyrosine phosphorylation of LRP6 by Src and Fer inhibits Wnt/beta-catenin signalling. EMBO Rep. 2014, 15, 1254–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Kratz, A.S.; Salama, M.; Elabd, S.; Heinrich, T.; Wittbrodt, J.; Blattner, C.; Davidson, G. Expression screening using a Medaka cDNA library identifies evolutionarily conserved regulators of the p53/Mdm2 pathway. BMC Biotechnol. 2015, 15, 92. [Google Scholar] [CrossRef] [Green Version]
- Davidson, G.; Shen, J.; Huang, Y.L.; Su, Y.; Karaulanov, E.; Bartscherer, K.; Hassler, C.; Stannek, P.; Boutros, M.; Niehrs, C. Cell cycle control of wnt receptor activation. Dev. Cell 2009, 17, 788–799. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, K.; Kurata-Miura, K.; Ujita, M.; Angata, K.; Nakagawa, S.; Sekine, S.; Nishi, T.; Fukuda, M. Expression cloning of cDNA encoding a human beta-1,3-N-acetylglucosaminyltransferase that is essential for poly-N-acetyllactosamine synthesis. Proc. Natl. Acad. Sci. USA 1997, 94, 14294–14299. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, N.; Natsume, A.; Togayachi, A.; Endo, T.; Akashima, T.; Yamada, Y.; Imai, N.; Nakagawa, S.; Koizumi, S.; Sekine, S.; et al. Identification and characterization of three novel beta 1,3-N-acetylglucosaminyltransferases structurally related to the beta 1,3-galactosyltransferase family. J. Biol. Chem. 2001, 276, 3498–3507. [Google Scholar] [CrossRef] [Green Version]
- Ishida, H.; Togayachi, A.; Sakai, T.; Iwai, T.; Hiruma, T.; Sato, T.; Okubo, R.; Inaba, N.; Kudo, T.; Gotoh, M.; et al. A novel beta1,3-N-acetylglucosaminyltransferase (beta3Gn-T8), which synthesizes poly-N-acetyllactosamine, is dramatically upregulated in colon cancer. FEBS Lett. 2005, 579, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Togayachi, A.; Kozono, Y.; Kuno, A.; Ohkura, T.; Sato, T.; Hirabayashi, J.; Ikehara, Y.; Narimatsu, H. β3GnT2 (B3GNT2), a Major Polylactosamine Synthase: Analysis of B3gnt2-Deficient Mice. In Functional Glycomics; Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2010; pp. 185–204. [Google Scholar]
- Liu, G.; Bafico, A.; Harris, V.K.; Aaronson, S.A. A novel mechanism for Wnt activation of canonical signaling through the LRP6 receptor. Mol. Cell Biol. 2003, 23, 5825–5835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarentino, A.L.; Gomez, C.M.; Plummer, T.H., Jr. Deglycosylation of asparagine-linked glycans by peptide:N-glycosidase F. Biochemistry 1985, 24, 4665–4671. [Google Scholar] [CrossRef] [PubMed]
- Freeze, H.H. Endoglycosidase and glycoamidase release of N-linked oligosaccharides. Curr. Protoc. Mol. Biol. 2001, Chapter 17, Unit17.13A. [Google Scholar] [CrossRef]
- Seko, A.; Yamashita, K. Activation of beta1,3-N-acetylglucosaminyltransferase-2 (beta3Gn-T2) by beta3Gn-T8. Possible involvement of beta3Gn-T8 in increasing poly-N-acetyllactosamine chains in differentiated HL-60 cells. J. Biol. Chem. 2008, 283, 33094–33100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkle, R.K.; Cummings, R.D. Lectin affinity chromatography of glycopeptides. Methods Enzymol. 1987, 138, 232–259. [Google Scholar] [CrossRef]
- Takeda-Uchimura, Y.; Nishitsuji, K.; Ikezaki, M.; Akama, T.O.; Ihara, Y.; Allain, F.; Uchimura, K. Beta3Gn-T7 Is a Keratan Sulfate beta1,3 N-Acetylglucosaminyltransferase in the Adult Brain. Front. Neuroanat. 2022, 16, 813841. [Google Scholar] [CrossRef] [PubMed]
- Littlechild, S.L.; Young, R.D.; Caterson, B.; Yoshida, H.; Yamazaki, M.; Sakimura, K.; Quantock, A.J.; Akama, T.O. Keratan Sulfate Phenotype in the beta-1,3-N-Acetylglucosaminyltransferase-7-Null Mouse Cornea. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1641–1651. [Google Scholar] [CrossRef] [Green Version]
- Seko, A.; Yamashita, K. beta1,3-N-Acetylglucosaminyltransferase-7 (beta3Gn-T7) acts efficiently on keratan sulfate-related glycans. FEBS Lett. 2004, 556, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Oh-Hashi, K.; Furuta, E.; Fujimura, K.; Hirata, Y. Application of a novel HiBiT peptide tag for monitoring ATF4 protein expression in Neuro2a cells. Biochem. Biophys. Rep. 2017, 12, 40–45. [Google Scholar] [CrossRef]
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 729–749. [Google Scholar] [CrossRef]
- Meyer, A.; Van de Peer, Y. From 2R to 3R: Evidence for a fish-specific genome duplication (FSGD). Bioessays 2005, 27, 937–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petit, D.; Teppa, R.E.; Harduin-Lepers, A. A phylogenetic view and functional annotation of the animal beta1,3-glycosyltransferases of the GT31 CAZy family. Glycobiology 2021, 31, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Kitayama, K.; Hayashida, Y.; Nishida, K.; Akama, T.O. Enzymes responsible for synthesis of corneal keratan sulfate glycosaminoglycans. J. Biol. Chem. 2007, 282, 30085–30096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongsma, M.L.M.; de Waard, A.A.; Raaben, M.; Zhang, T.; Cabukusta, B.; Platzer, R.; Blomen, V.A.; Xagara, A.; Verkerk, T.; Bliss, S.; et al. The SPPL3-defined glycosphingolipid repertoire orchestrates HLA class I-mediated immune responses. Immunity 2021, 54, 387. [Google Scholar] [CrossRef]
- Yamamoto, H.; Sakane, H.; Yamamoto, H.; Michiue, T.; Kikuchi, A. Wnt3a and Dkk1 regulate distinct internalization pathways of LRP6 to tune the activation of beta-catenin signaling. Dev. Cell 2008, 15, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, A.; Yamamoto, H.; Sato, A. Selective activation mechanisms of Wnt signaling pathways. Trends Cell Biol. 2009, 19, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; van der Zon, G.; Ma, J.; Mei, H.; Cabukusta, B.; Agaser, C.C.; Madunic, K.; Wuhrer, M.; Zhang, T.; Ten Dijke, P. ST3GAL5-catalyzed gangliosides inhibit TGF-beta-induced epithelial-mesenchymal transition via TbetaRI degradation. EMBO J. 2023, 42, e110553. [Google Scholar] [CrossRef]
- Li, Y.; Lu, W.; He, X.; Bu, G. Modulation of LRP6-mediated Wnt signaling by molecular chaperone Mesd. FEBS Lett. 2006, 580, 5423–5428. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, R.; Wang, X.; Safi, S.; Braunegger, N.; Hipgrave Ederveen, A.; Rottmann, M.; Wittbrodt, J.; Wuhrer, M.; Wesslowski, J.; Davidson, G. N-Glycosylation of LRP6 by B3GnT2 Promotes Wnt/β-Catenin Signalling. Cells 2023, 12, 863. https://doi.org/10.3390/cells12060863
Xu R, Wang X, Safi S, Braunegger N, Hipgrave Ederveen A, Rottmann M, Wittbrodt J, Wuhrer M, Wesslowski J, Davidson G. N-Glycosylation of LRP6 by B3GnT2 Promotes Wnt/β-Catenin Signalling. Cells. 2023; 12(6):863. https://doi.org/10.3390/cells12060863
Chicago/Turabian StyleXu, Ruiyao, Xianxian Wang, Sadia Safi, Nico Braunegger, Agnes Hipgrave Ederveen, Michelle Rottmann, Joachim Wittbrodt, Manfred Wuhrer, Janine Wesslowski, and Gary Davidson. 2023. "N-Glycosylation of LRP6 by B3GnT2 Promotes Wnt/β-Catenin Signalling" Cells 12, no. 6: 863. https://doi.org/10.3390/cells12060863
APA StyleXu, R., Wang, X., Safi, S., Braunegger, N., Hipgrave Ederveen, A., Rottmann, M., Wittbrodt, J., Wuhrer, M., Wesslowski, J., & Davidson, G. (2023). N-Glycosylation of LRP6 by B3GnT2 Promotes Wnt/β-Catenin Signalling. Cells, 12(6), 863. https://doi.org/10.3390/cells12060863