
Ectopic Rod Photoreceptor Development in Mice with Genetic Deficiency of WNT2B
 , , , and
, , , and 
Abstract
:1. Introduction
2. Methods
2.1. Generation of Wnt2b Knockout (KO) Mice
2.2. Eye Dissection and Examination of Anterior Segments and Lens
2.3. Analysis of Retinal Vasculature in Retinal Flat Mounts
2.4. Retinal Histology Characterization in Eye Cross Sections with H&E Staining
2.5. Immunohistochemistry
2.6. Fundus and OCT Imaging
2.7. Polymerase Chain Reaction (PCR) and Genotyping
- Genotyping for Wnt2b:
- Genotyping for CMV-Cre:
2.8. RNA Isolation and Real-Time Quantitative PCR (RT-qPCR)
2.9. Statistical Analysis
3. Results
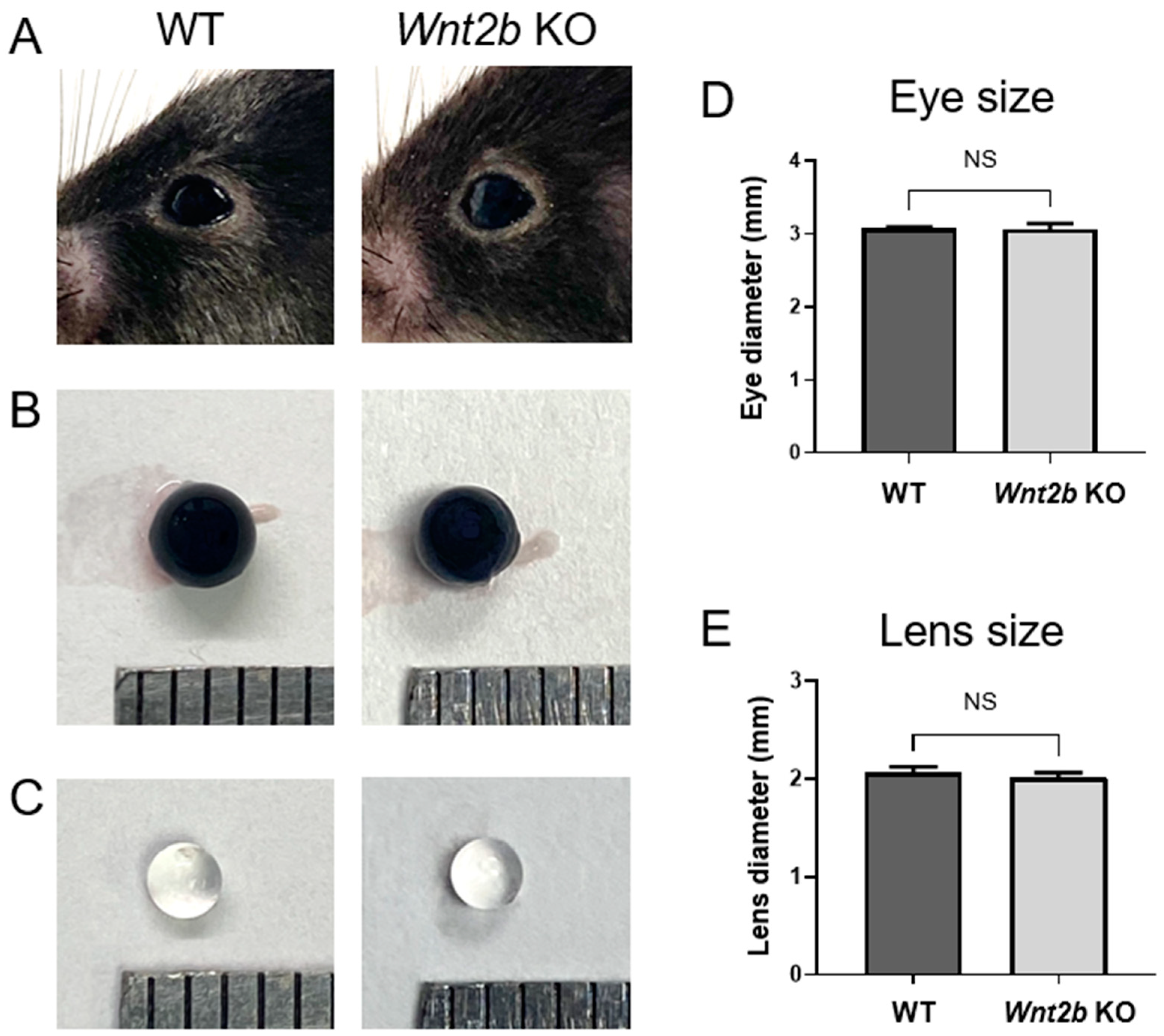
3.1. Normal Appearance of Anterior Segments of Wnt2b KO Mice
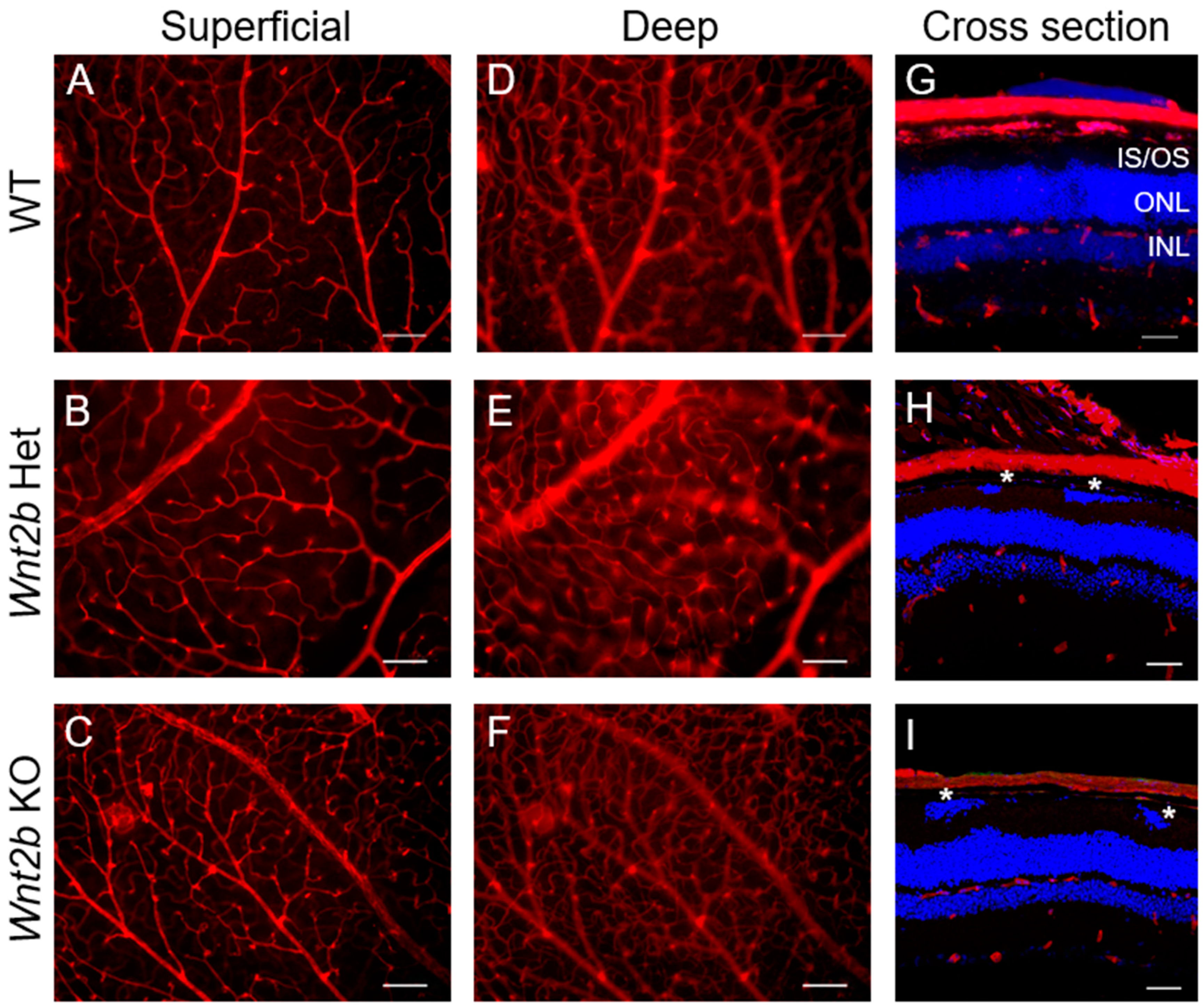
3.2. The Retinal Vasculature of Wnt2b KO Mice Appears Normal
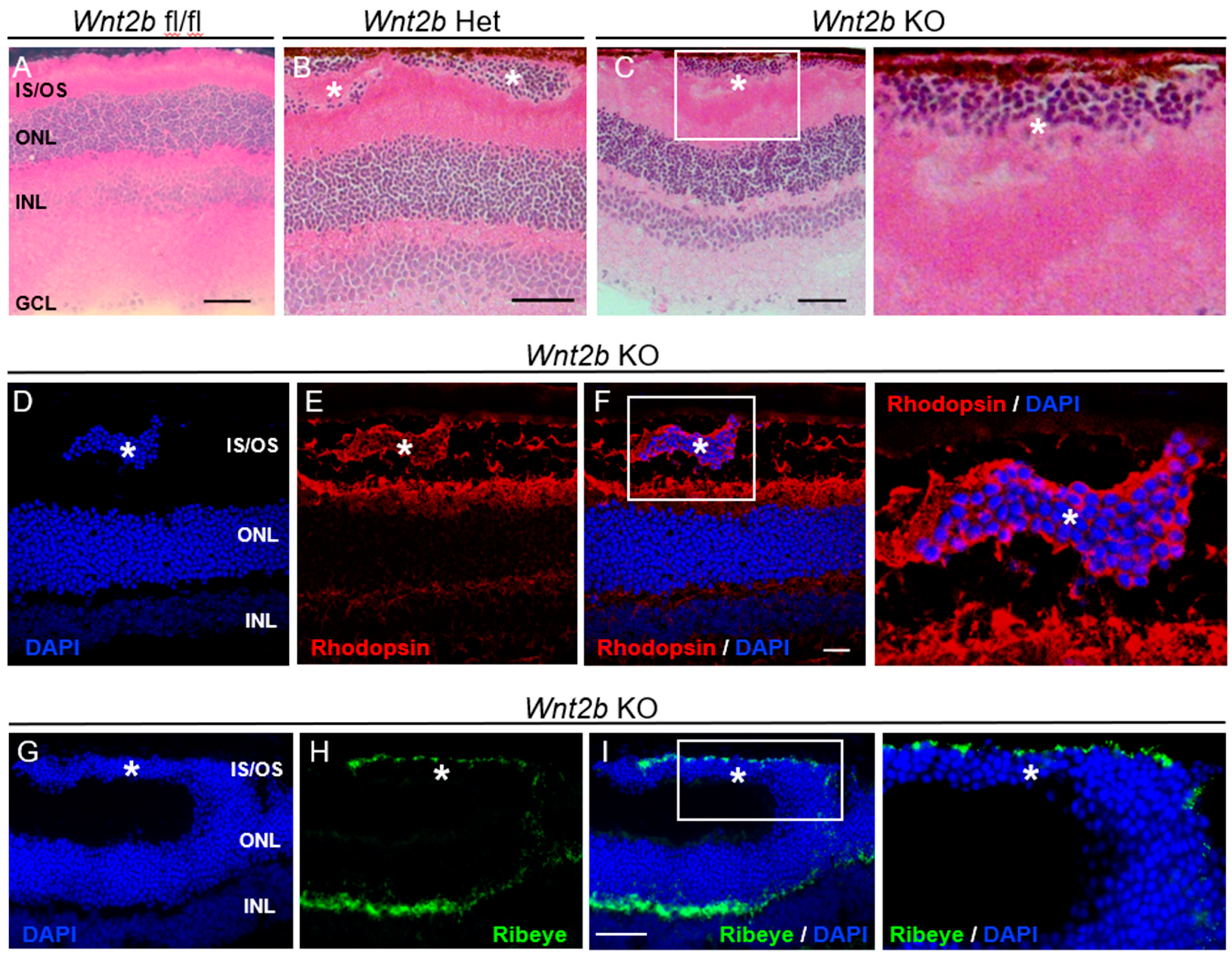
3.3. Ectopic Formation of Rod Photoreceptor Clusters in the Subretinal Space in Adult Wnt2b Mutant Mice
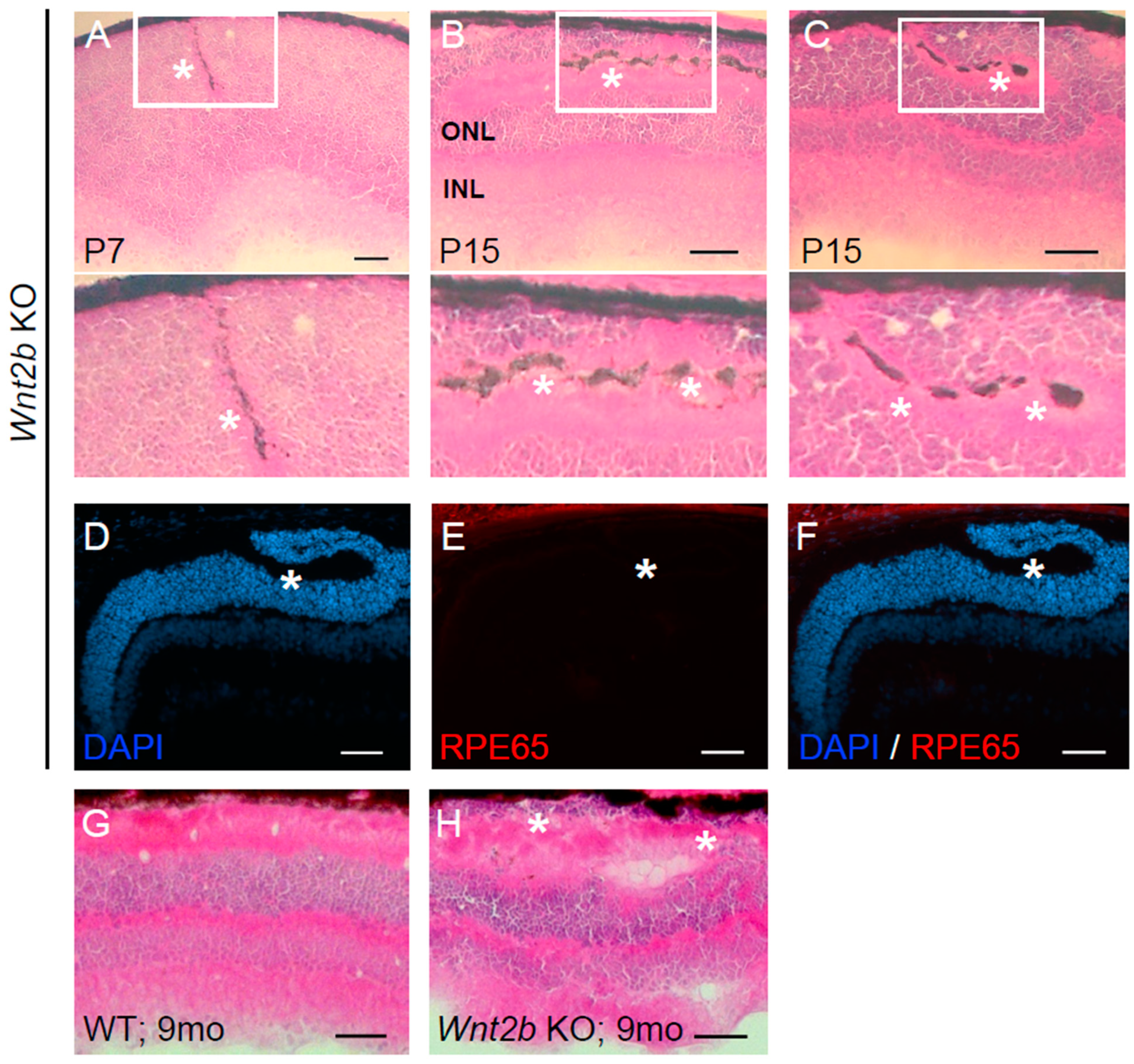
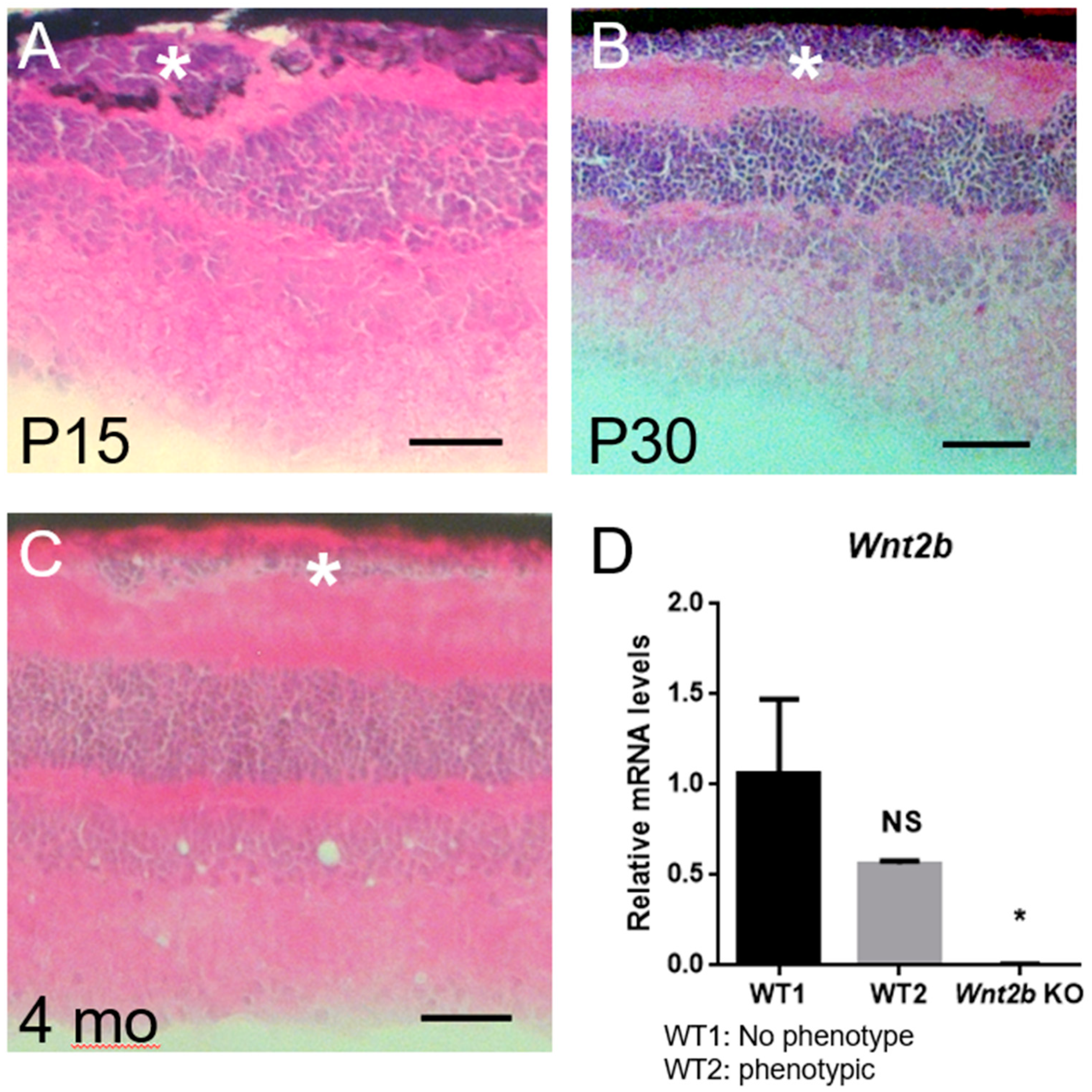
3.4. Ectopic Subretinal Cell Clusters Form during Wnt2b Mutant Mice Development
3.5. Absence of Retinal Thinning and Degeneration in Aged Wnt2b KO Mice
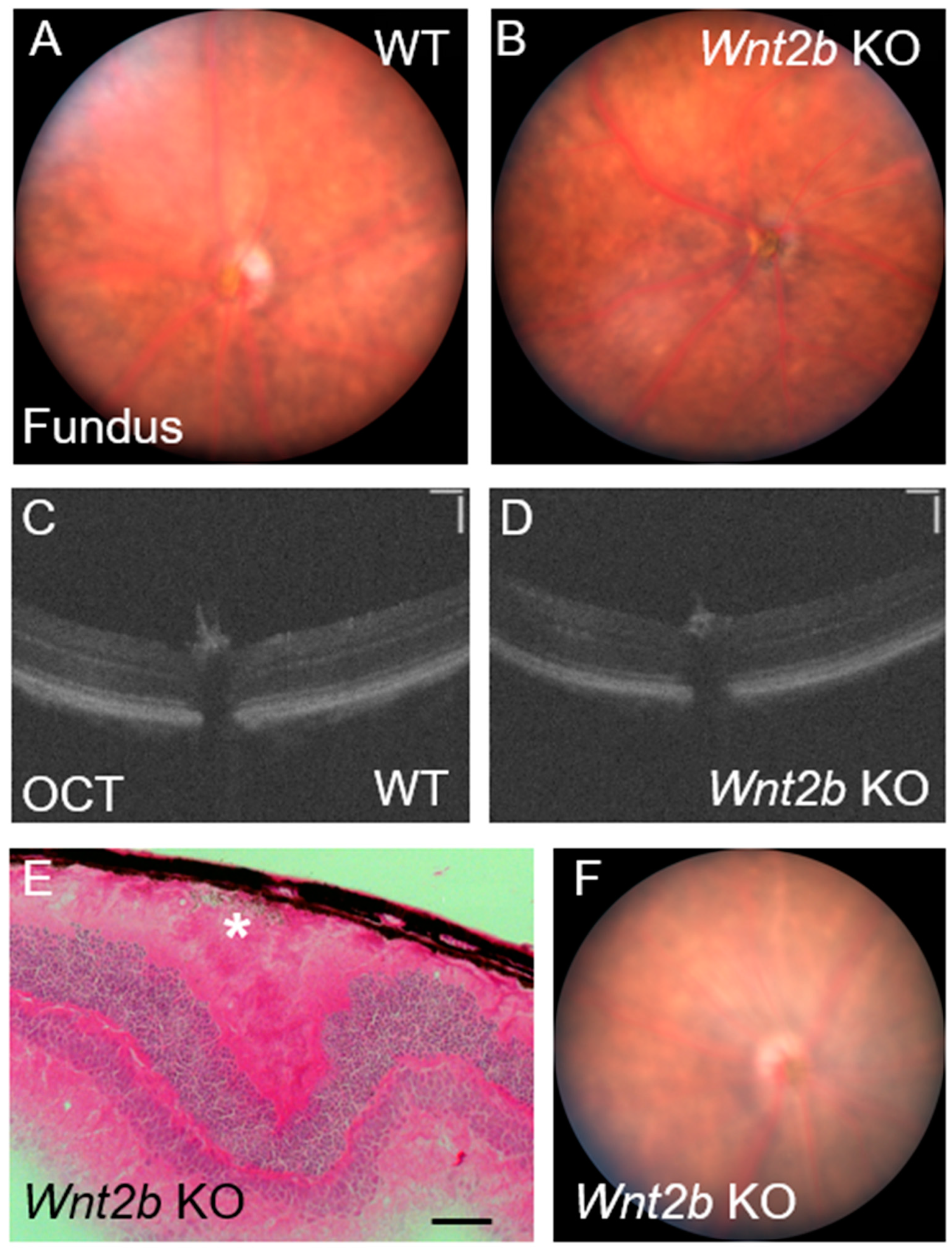
3.6. Lack of Phenotype Detection in Fundus Imaging and OCT in Live Mice
3.7. Occasional Presence of Phenotype in WT Mice
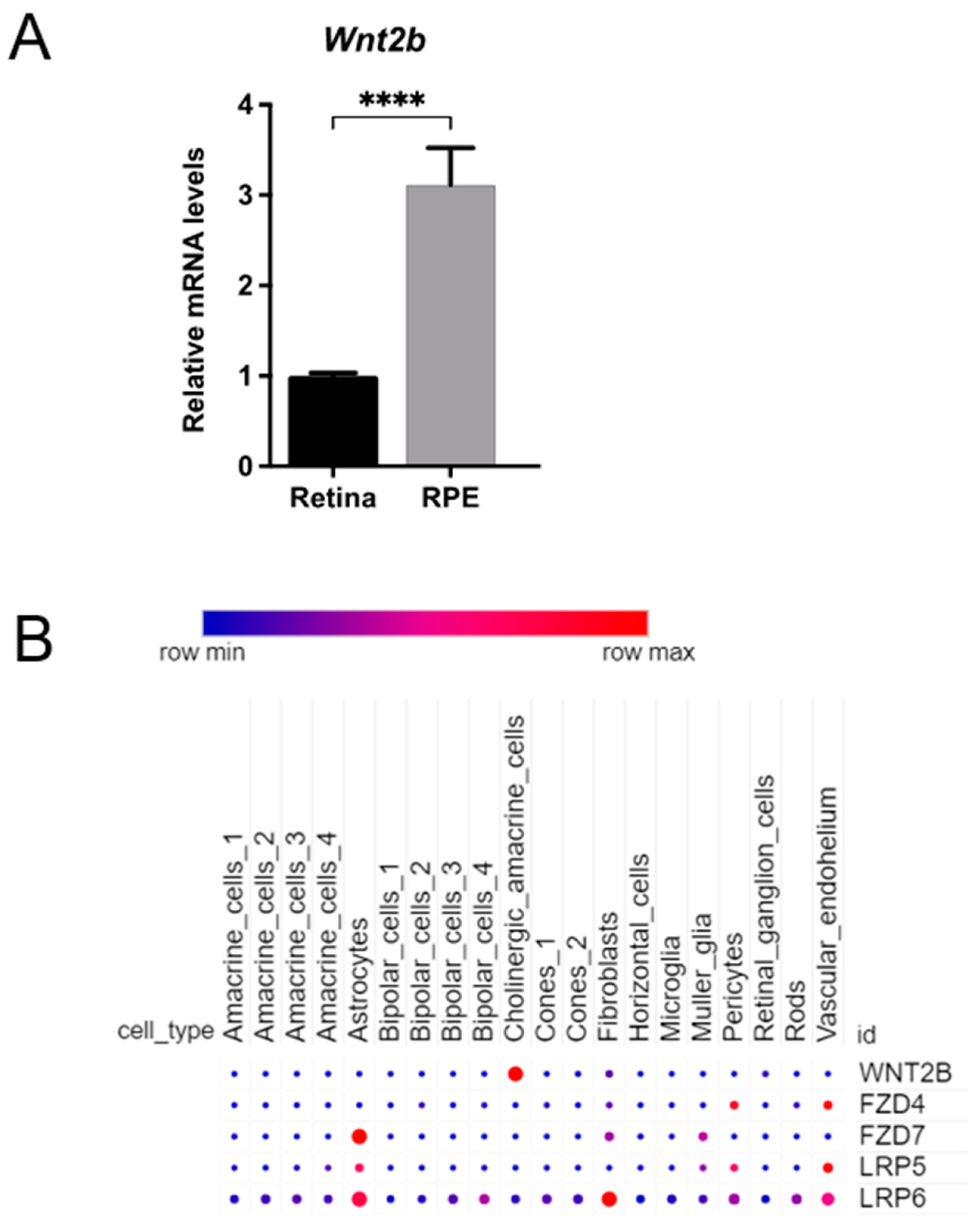
3.8. Expression of Wnt2b mRNA in the RPE and Retina
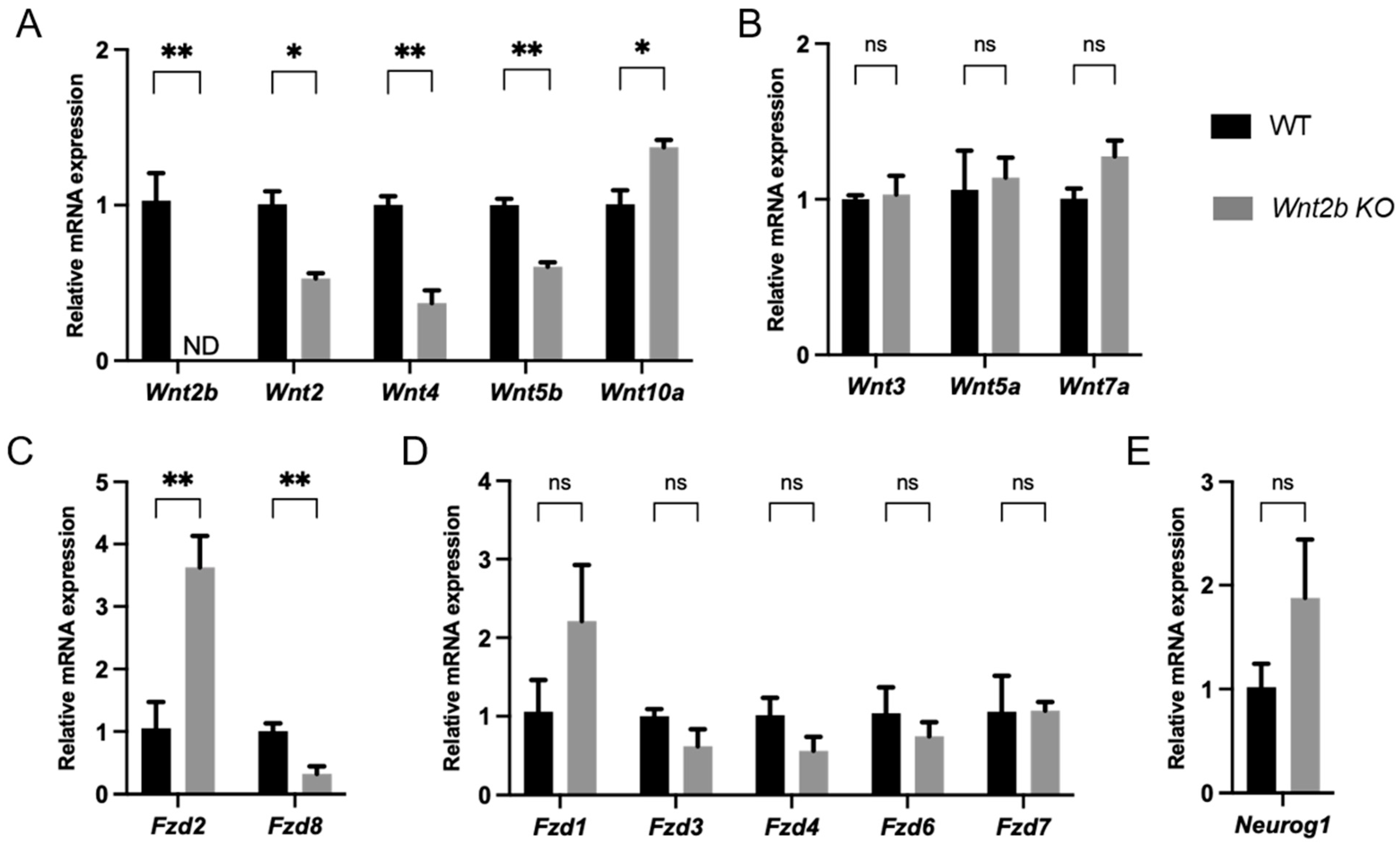
3.9. Expression of Wnt Ligands and Frizzled (Fzd) Receptors Was Altered in Wnt2b KO Retinas
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fujimura, N. WNT/beta-Catenin Signaling in Vertebrate Eye Development. Front. Cell Dev. Biol. 2016, 4, 138. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Xu, S.; Wang, Y.; Mazerolle, C.; Thurig, S.; Coles, B.L.; Ren, J.-C.; Taketo, M.M.; van der Kooy, D.; Wallace, V.A. Ciliary margin transdifferentiation from neural retina is controlled by canonical Wnt signaling. Dev. Biol. 2007, 308, 54–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cvekl, A.; Zhang, X. Signaling and Gene Regulatory Networks in Mammalian Lens Development. Trends Genet. 2017, 33, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Steinfeld, J.; Steinfeld, I.; Coronato, N.; Hampel, M.-L.; Layer, P.G.; Araki, M.; Vogel-Höpker, A. RPE specification in the chick is mediated by surface ectoderm-derived BMP and Wnt signalling. Development 2013, 140, 4959–4969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, N.; Taketo, M.M.; Mori, M.; Korinek, V.; Kozmik, Z. Spatial and temporal regulation of Wnt/beta-catenin signaling is essential for development of the retinal pigment epithelium. Dev. Biol. 2009, 334, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, C.-H.; Huang, S.; Chen, J. Wnt Signaling in vascular eye diseases. Prog. Retin. Eye Res. 2018, 70, 110–133. [Google Scholar] [CrossRef]
- Steinfeld, J.; Steinfeld, I.; Bausch, A.; Coronato, N.; Hampel, M.L.; Depner, H.; Layer, P.G.; Vogel-Höpker, A. BMP-induced reprogramming of the neural retina into retinal pigment epithelium requires Wnt signalling. Biol. Open 2017, 6, 979–992. [Google Scholar]
- Grocott, T.; Johnson, S.; Bailey, A.P.; Streit, A. Neural crest cells organize the eye via TGF-beta and canonical Wnt signalling. Nat. Commun. 2011, 2, 265. [Google Scholar] [CrossRef] [Green Version]
- Seitz, R.; Weber, G.; Albrecht, S.; Fuchshofer, R.; Tamm, E.R.; Ohlmann, A. Cross-Inhibition of Norrin and TGF-beta Signaling Modulates Development of Retinal and Choroidal Vasculature. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2240–2251. [Google Scholar] [CrossRef] [Green Version]
- Katoh, M. WNT2B: Comparative integromics and clinical applications (Review). Int. J. Mol. Med. 2005, 16, 1103–1108. [Google Scholar] [CrossRef]
- Ortiz-Masià, D.; Salvador, P.; Macias-Ceja, D.C.; Gisbert-Ferrándiz, L.; Esplugues, J.V.; Manyé, J.; Alós, R.; Navarro-Vicente, F.; Mamie, C.; Scharl, M.; et al. WNT2b Activates Epithelial-mesenchymal Transition Through FZD4: Relevance in Penetrating Crohn s Disease. J. Crohn’s Colitis 2020, 14, 230–239. [Google Scholar] [CrossRef]
- Cho, S.H.; Cepko, C.L. Wnt2b/beta-catenin-mediated canonical Wnt signaling determines the peripheral fates of the chick eye. Development 2006, 133, 3167–3177. [Google Scholar] [CrossRef] [Green Version]
- Ohta, K.; Ito, A.; Kuriyama, S.; Lupo, G.; Kosaka, M.; Ohnuma, S.-I.; Nakagawa, S.; Tanaka, H. Tsukushi functions as a Wnt signaling inhibitor by competing with Wnt2b for binding to transmembrane protein Frizzled4. Proc. Natl. Acad. Sci. USA 2011, 108, 14962–14967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, F.; Takeichi, M.; Nakagawa, S. Wnt2b inhibits differentiation of retinal progenitor cells in the absence of Notch activity by downregulating the expression of proneural genes. Development 2005, 132, 2759–2770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, A.E.; Zhou, F.; Shah, M.S.; Murphy, Q.; Rickner, H.; Kelsen, J.; Boyle, J.; Doyle, J.J.; Gangwani, B.; Thiagarajah, J.R.; et al. Neonatal-Onset Chronic Diarrhea Caused by Homozygous Nonsense WNT2B Mutations. Am. J. Hum. Genet. 2018, 103, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.J.; Jimenez, L.; Azova, S.; Kremen, J.; Chan, Y.-M.; Elhusseiny, A.M.; Saeed, H.; Goldsmith, J.; Al-Ibraheemi, A.; O’Connell, A.E.; et al. Novel variants in the stem cell niche factor WNT2B define the disease phenotype as a congenital enteropathy with ocular dysgenesis. Eur. J. Hum. Genet. 2021, 29, 998–1007. [Google Scholar] [CrossRef]
- Islam, F.; Htun, S.; Lai, L.W.; Krall, M.; Poranki, M.; Martin, P.M.; Sobreira, N.; Wohler, E.S.; Yu, J.; Moore, A.T.; et al. Exome sequencing in patients with microphthalmia, anophthalmia, and coloboma (MAC) from a consanguineous population. Clin. Genet. 2020, 98, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Tsukiyama, T.; Yamaguchi, T.P. Mice lacking Wnt2b are viable and display a postnatal olfactory bulb phenotype. Neurosci. Lett. 2012, 512, 48–52. [Google Scholar] [CrossRef] [Green Version]
- Mattapallil, M.J.; Wawrousek, E.F.; Chan, C.C.; Zhao, H.; Roychoudhury, J.; Ferguson, T.A.; Caspi, R.R. The Rd8 mutation of the Crb1 gene is present in vendor lines of C57BL/6N mice and embryonic stem cells, and confounds ocular induced mutant phenotypes. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2921–2927. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.H.; Wang, Z.; Sun, Y.; Chen, J. Animal models of ocular angiogenesis: From development to pathologies. FASEB J. 2017, 31, 4665–4681. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Yemanyi, F.; Blomfield, A.K.; Bora, K.; Huang, S.; Liu, C.-H.; Britton, W.R.; Cho, S.S.; Tomita, Y.; Fu, Z.; et al. Amino acid transporter SLC38A5 regulates developmental and pathological retinal angiogenesis. eLife 2022, 11, e73105. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Liu, C.H.; Wang, Z.; Fu, Z.; Britton, W.R.; Blomfield, A.K.; Kamenecka, T.M.; Kamenecka, J.L.; Solt, L.A.; Chen, J. REV-ERBalpha regulates age-related and oxidative stress-induced degeneration in retinal pigment epithelium via NRF2. Redox Biol. 2022, 51, 102261. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Li, J.; Sun, Y.; Fu, Z.; Liu, C.-H.; Evans, L.; Tian, K.; Saba, N.; Fredrick, T.; Morss-Walton, P.; et al. Optimization of an Image-Guided Laser-Induced Choroidal Neovascularization Model in Mice. PLoS ONE 2015, 10, e0132643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehalow, A.K. CRB1 is essential for external limiting membrane integrity and photoreceptor morphogenesis in the mammalian retina. PLoS ONE 2015, 10, e0132643. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.H.; Yemanyi, F.; Bora, K.; Kushwah, N.; Blomfield, A.K.; Kamenecka, T.M.; SanGiovanni, J.P.; Sun, Y.; Solt, L.A.; Chen, J. Genetic deficiency and pharmacological modulation of RORalpha regulate laser-induced choroidal neovascularization. Aging 2023, 15, 37–52. [Google Scholar] [CrossRef]
- Ye, X.; Wang, Y.; Nathans, J. The Norrin/Frizzled4 signaling pathway in retinal vascular development and disease. Trends Mol. Med. 2010, 16, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Macosko, E.Z.; Basu, A.; Satija, R.; Nemesh, J.; Shekhar, K.; Goldman, M.; Tirosh, I.; Bialas, A.R.; Kamitaki, N.; Martersteck, E.M.; et al. Highly Parallel Genome-wide Expression Profiling of Individual Cells Using Nanoliter Droplets. Cell 2015, 161, 1202–1214. [Google Scholar] [CrossRef] [Green Version]
- Yan, R.-T.; Li, X.; Wang, S.-Z. Photoreceptor-like cells in transgenic mouse eye. Investig. Ophthalmol. Vis. Sci. 2013, 54, 4766–4775. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Ma, W.; Zhuo, Y.; Yan, R.T.; Wang, S.Z. Using neurogenin to reprogram chick RPE to produce photoreceptor-like neurons. Investig. Ophthalmol. Vis. Sci. 2010, 51, 516–525. [Google Scholar] [CrossRef] [Green Version]
- Yan, R.-T.; He, L.; Zhan, W.; Wang, S.-Z. Induction of ectopic retina-like tissue by transgenic expression of neurogenin. PLoS ONE 2015, 10, e0116171. [Google Scholar] [CrossRef] [Green Version]
- Amram, B.; Cohen-Tayar, Y.; David, A.; Ashery-Padan, R. The retinal pigmented epithelium—From basic developmental biology research to translational approaches. Int. J. Dev. Biol. 2017, 61, 225–234. [Google Scholar] [CrossRef]
- Flanagan, D.J.; Phesse, T.J.; Barker, N.; Schwab, R.H.M.; Amin, N.; Malaterre, J.; Stange, D.E.; Nowell, C.J.; Currie, S.A.; Saw, J.T.S.; et al. Frizzled7 functions as a Wnt receptor in intestinal epithelial Lgr5+ stem cells. Stem Cell Rep. 2015, 4, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, S.; Zou, C.; Levine, E.M. Retinal pigment epithelium development, plasticity, and tissue homeostasis. Exp. Eye Res. 2014, 123, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Bharti, K.; Gasper, M.; Ou, J.; Brucato, M.; Clore-Gronenborn, K.; Pickel, J.; Arnheiter, H. A regulatory loop involving PAX6, MITF, and WNT signaling controls retinal pigment epithelium development. PLoS Genet. 2012, 8, e1002757. [Google Scholar] [CrossRef] [Green Version]
- Hawes, N.L.; Chang, B.; Hageman, G.S.; Nusinowitz, S.; Nishina, P.M.; Schneider, B.S.; Smith, R.S.; Roderick, T.H.; Davisson, M.T.; Heckenlively, J.R. Retinal degeneration 6 (rd6): A new mouse model for human retinitis punctata albescens. Investig. Ophthalmol. Vis. Sci. 2000, 41, 3149–3157. [Google Scholar]
- Bumsted, K.M.; Barnstable, C.J. Dorsal retinal pigment epithelium differentiates as neural retina in the microphthalmia (mi/mi) mouse. Investig. Ophthalmol. Vis. Sci. 2000, 41, 903–908. [Google Scholar]
- Song, H.; Bush, R.A.; Vijayasarathy, C.; Fariss, R.N.; Kjellstrom, S.; Sieving, P.A. Transgenic expression of constitutively active RAC1 disrupts mouse rod morphogenesis. Investig. Ophthalmol. Vis. Sci. 2014, 55, 2659–2668. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Hu, S.; Okray, Z.; Ren, X.; De Geest, N.; Claeys, A.; Yan, J.; Bellefroid, E.; Hassan, B.A.; Quan, X.-J. The Drosophila neurogenin Tap functionally interacts with the Wnt-PCP pathway to regulate neuronal extension and guidance. Development 2016, 143, 2760–2766. [Google Scholar] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward Sequence 5′-3′ | Reverse Sequence 5′-3′ |
|---|---|---|
| Wnt2b | GCCCGAGTGATCTGTGACAA | CACTCTCGGATCCATTCCCG |
| Wnt2 | CTCGGTGGAATCTGGCTCTG | CACATTGTCACACATCACCCT |
| Wnt3 | CCGCTTCTGTCTAGGGTCTG | GTAGAGAGTGCAGGCAAGGG |
| Wnt4 | AGACGTGCGAGAAACTCAAAG | GGAACTGGTATTGGCACTCCT |
| Wnt5a | CAACTGGCAGGACTTTCTCAA | CATCTCCGATGCCGGAACT |
| Wnt5b | GGTTCCACTGGTGTTGCTTT | AGACTTTTGTGAGGCGGAGA |
| Wnt7a | CACTTGTGGTCTCAGGGGTT | GCATCTGAGTTTCACCAGCA |
| Wnt10a | GCTCAACGCCAACACAGTG | CGAAAACCTCGGCTGAAGATG |
| Fzd1 | CAGCAGTACAACGGCGAAC | GTCCTCCTGATTCGTGTGGC |
| Fzd2 | CATGCCCAACCTTCTTGGC | CAGCGGGTAGAACTGATGCAC |
| Fzd3 | ATGGCTGTGAGCTGGATTGTC | GGCACATCCTCAAGGTTATAGGT |
| Fzd4 | AGACGTGCGAACTCAAAG | GGAACTGGTATTGGCACTCCT |
| Fzd6 | ATGGAAAGGTCCCCGTTTCTG | GGGAAGAACGTCATGTTGTAAGT |
| Fzd7 | GCCACACGAACCAAGAGGAC | CGGGTGCGTACATAGAGCATAA |
| Fzd8 | ATGGAGTGGGGTTACCTGTTG | CACCGTGATCTCTTGGCAC |
| Neurog1 | AGTAGTCCCTCGGCTTCAGA | TATGGGATGAAACAGGGCGT |
| Gapdh | AACAGCAACTCCCACTCTTC | CCTGTTGCTGTAGCCGTATT |
| Mouse Age | Genotype | ||
|---|---|---|---|
| WT or Wnt2b fl/fl * | Wnt2b Het | Wnt2b KO | |
| P7 | 0/4 | 1/4 | 1/3 |
| P15 | 2/6 | 2/3 | 1/4 |
| P30 | 1/4 | 1/2 | 1/4 |
| 2–5 mo | 1/6 | 4/5 | 3/6 |
| 9 mo | 0/3 | 0/1 | 1/3 |
| Total | 4/23 (17%) | 8/15 (53%) | 7/20 (35%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blomfield, A.K.; Maurya, M.; Bora, K.; Pavlovich, M.C.; Yemanyi, F.; Huang, S.; Fu, Z.; O’Connell, A.E.; Chen, J. Ectopic Rod Photoreceptor Development in Mice with Genetic Deficiency of WNT2B. Cells 2023, 12, 1033. https://doi.org/10.3390/cells12071033
Blomfield AK, Maurya M, Bora K, Pavlovich MC, Yemanyi F, Huang S, Fu Z, O’Connell AE, Chen J. Ectopic Rod Photoreceptor Development in Mice with Genetic Deficiency of WNT2B. Cells. 2023; 12(7):1033. https://doi.org/10.3390/cells12071033
Chicago/Turabian StyleBlomfield, Alexandra K., Meenakshi Maurya, Kiran Bora, Madeline C. Pavlovich, Felix Yemanyi, Shuo Huang, Zhongjie Fu, Amy E. O’Connell, and Jing Chen. 2023. "Ectopic Rod Photoreceptor Development in Mice with Genetic Deficiency of WNT2B" Cells 12, no. 7: 1033. https://doi.org/10.3390/cells12071033
APA StyleBlomfield, A. K., Maurya, M., Bora, K., Pavlovich, M. C., Yemanyi, F., Huang, S., Fu, Z., O’Connell, A. E., & Chen, J. (2023). Ectopic Rod Photoreceptor Development in Mice with Genetic Deficiency of WNT2B. Cells, 12(7), 1033. https://doi.org/10.3390/cells12071033