
Prolonged Subculturing of Aspergillus fumigatus on Galleria Extract Agar Results in Altered Virulence and Sensitivity to Antifungal Agents


Abstract
:1. Introduction
2. Materials and Methods
2.1. Aspergillus fumigatus Culture Conditions
2.2. Generation of Passaged Strains of Aspergillus fumigatus
2.3. Virulence Assessment of Passaged Strains In Vivo
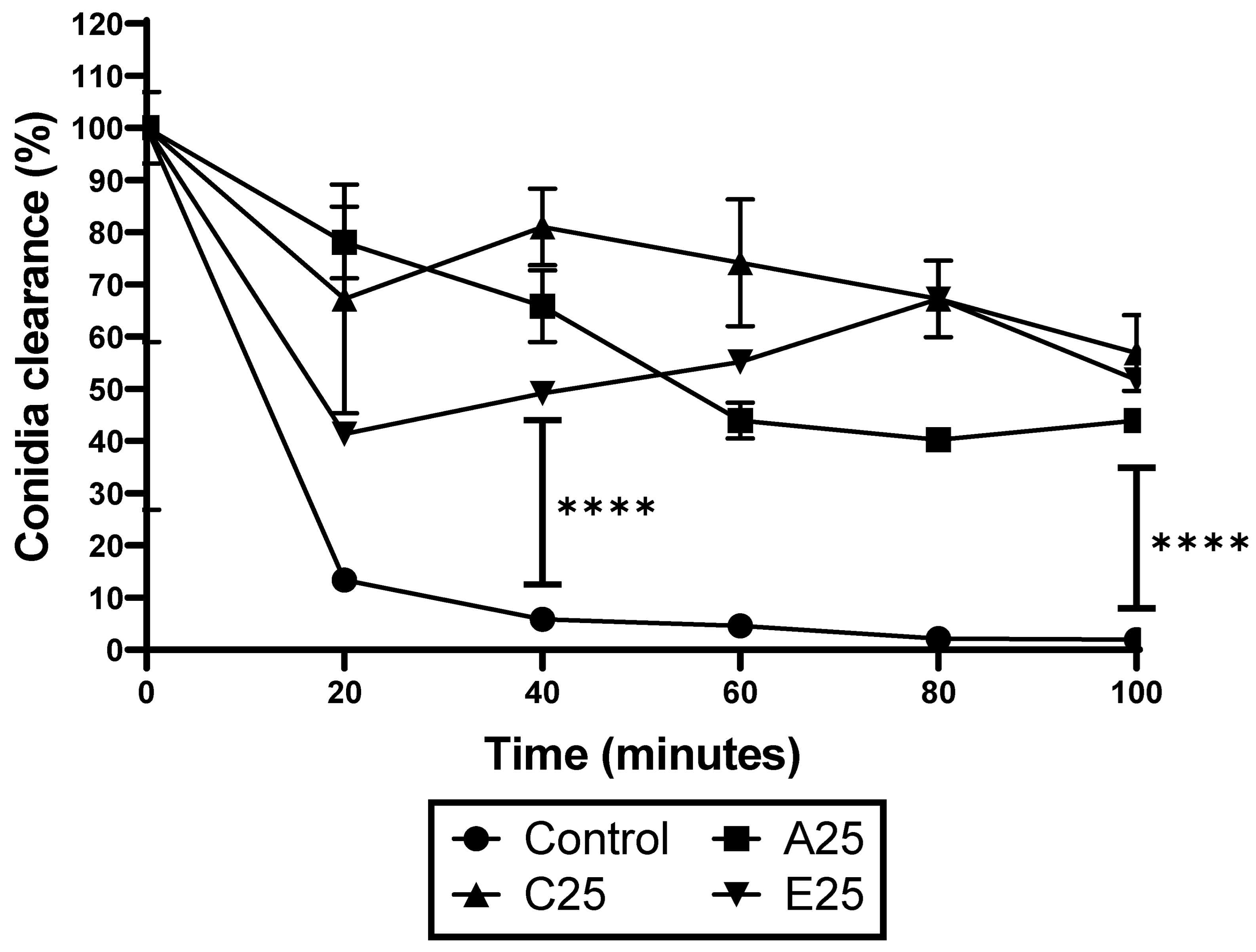
2.4. Hemocyte Kill Assay
2.5. Susceptibility Testing
2.6. Gliotoxin Extraction and Quantification
2.7. Total Secreted Siderophore Quantification
2.8. Protein Extraction and Purification from A. fumigatus Hyphae
2.9. Mass Spectrometry
2.10. Data Analysis
2.11. Statistical Analysis
3. Results
3.1. Characterization of Growth Characteristics of Serially Subcultured A. fumigatus Strains
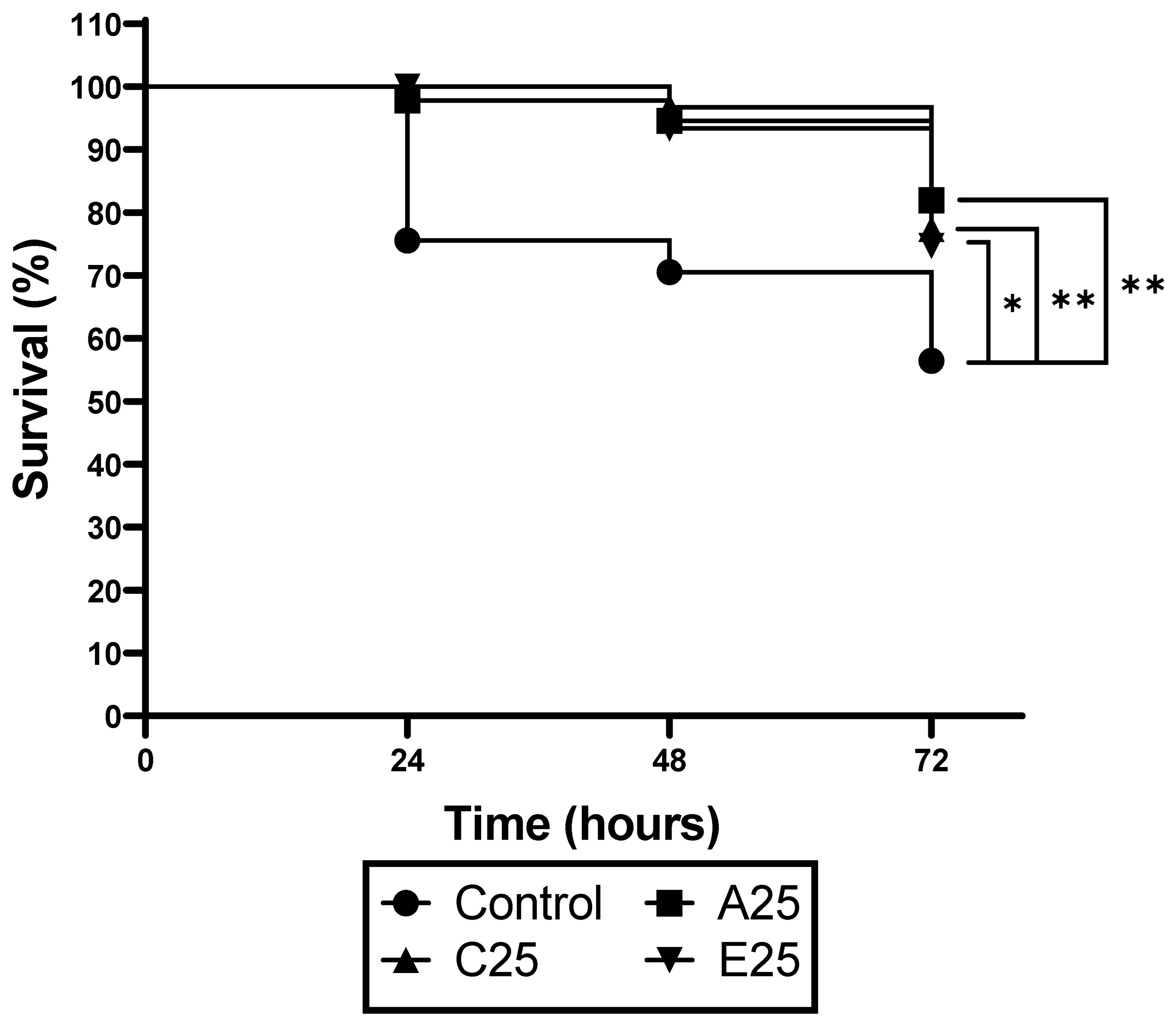
3.2. Passaged A. fumigatus Strains Show Reduced Virulence in G. mellonella Larvae
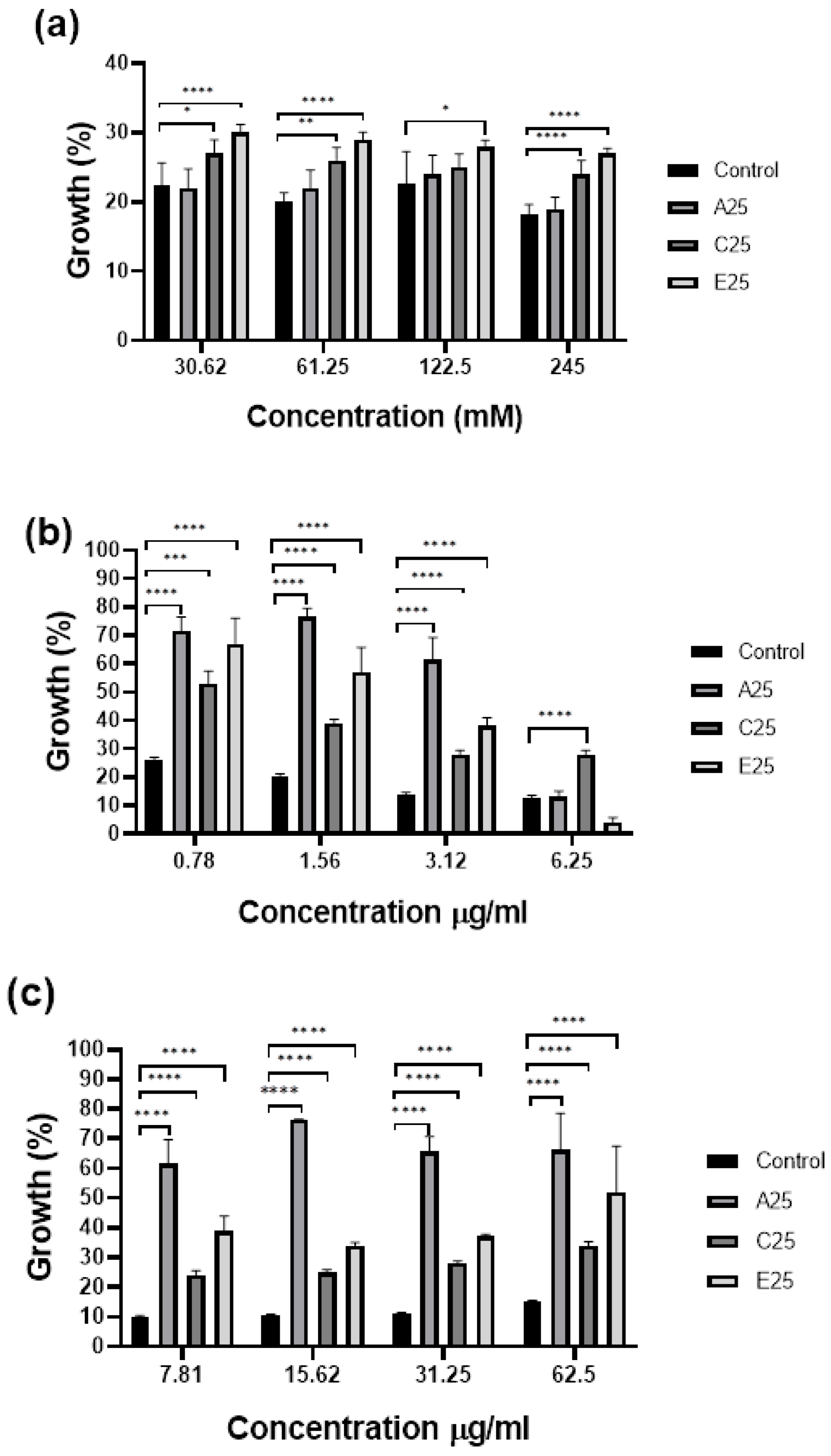
3.3. Passaged A. fumigatus Strains Show Altered Susceptibility to Antifungal Agents
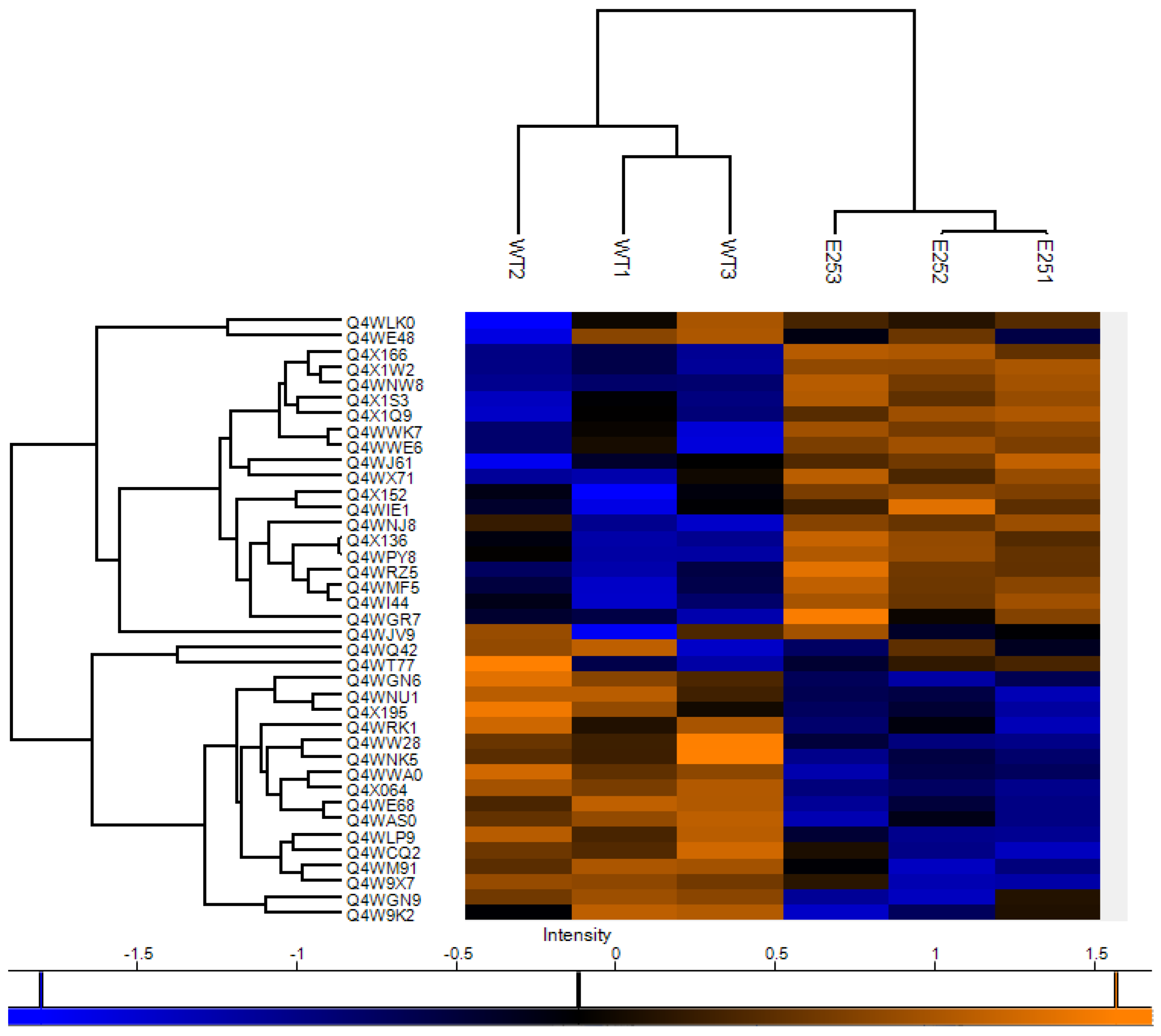
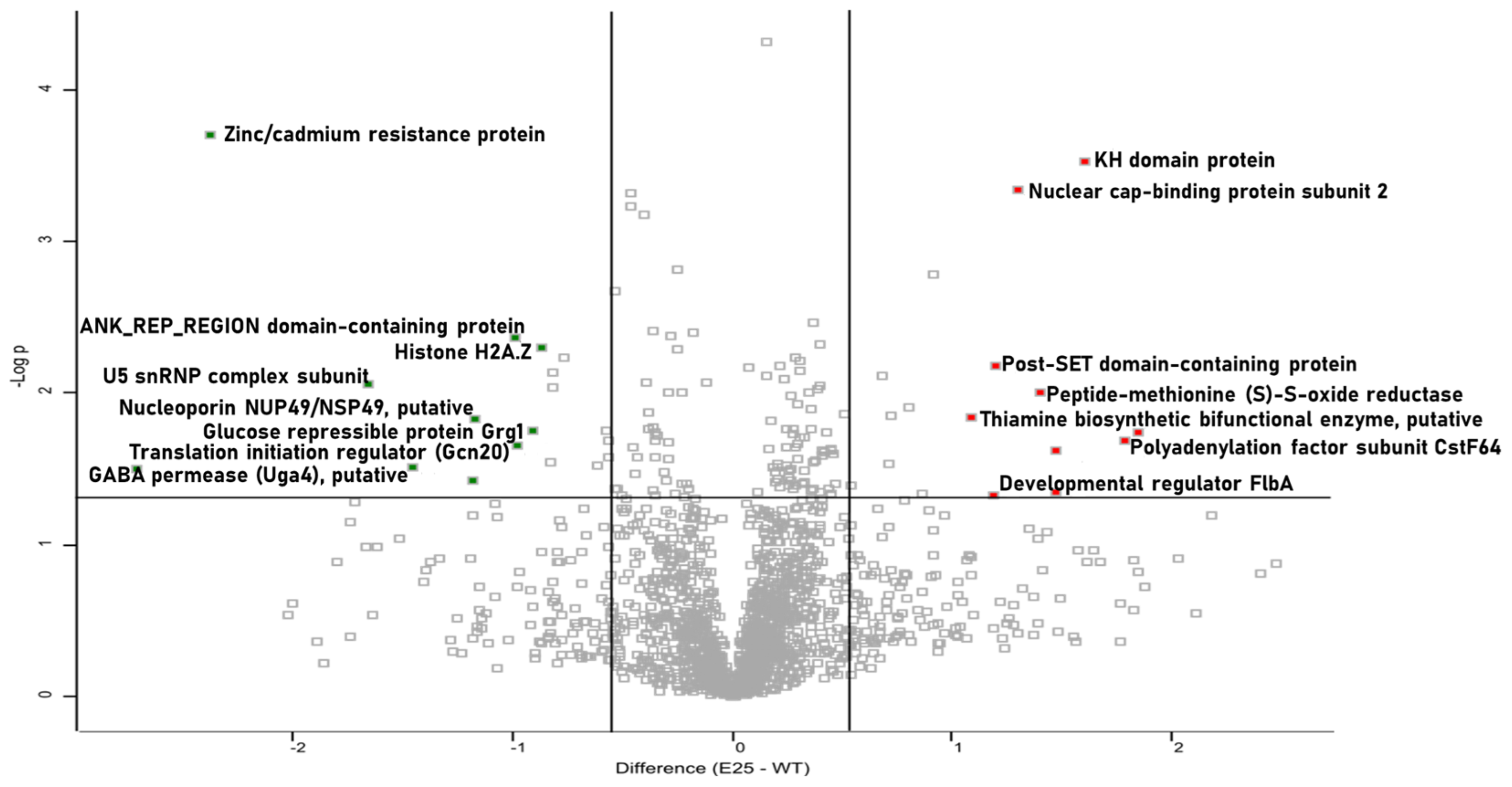
3.4. Proteomic Characterization of Passaged A. fumigatus Strain E25
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mousavi, B.; Hedayati, M.; Hedayati, N.; Ilkit, M.; Syedmousavi, S. Aspergillus Species in Indoor Environments and Their Possible Occupational and Public Health Hazards. Curr. Med. Mycol. 2016, 2, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokas, A.; Mead, M.E.; Steenwyk, J.L.; Oberlies, N.H.; Goldman, G.H. Evolving Moldy Murderers: Aspergillus Section Fumigati as a Model for Studying the Repeated Evolution of Fungal Pathogenicity. PLoS Pathog. 2020, 16, e1008315. [Google Scholar] [CrossRef] [Green Version]
- Raksha; Singh, G.; Urhekar, A.D. Virulence Factors Detection in Aspergillus Isolates from Clinical and Environmental Samples. J. Clin. Diagn. Res. 2017, 11, DC13–DC18. [Google Scholar] [CrossRef]
- Van Waeyenberghe, L.; Baré, J.; Pasmans, F.; Claeys, M.; Bert, W.; Haesebrouck, F.; Houf, K.; Martel, A. Interaction of Aspergillus fumigatus Conidia with Acanthamoeba Castellanii Parallels Macrophage-Fungus Interactions. Environ. Microbiol. Rep. 2013, 5, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Hillmann, F.; Novohradská, S.; Mattern, D.J.; Forberger, T.; Heinekamp, T.; Westermann, M.; Winckler, T.; Brakhage, A.A. Virulence Determinants of the Human Pathogenic Fungus Aspergillus fumigatus Protect against Soil Amoeba Predation. Environ. Microbiol. 2015, 17, 2858–2869. [Google Scholar] [CrossRef]
- Ben-Ami, R.; Lewis, R.E.; Kontoyiannis, D.P. Enemy of the (Immunosuppressed) State: An Update on the Pathogenesis of Aspergillus fumigatus Infection. Br. J. Haematol. 2010, 150, 406–417. [Google Scholar] [CrossRef]
- El-Baba, F.; Gao, Y.; Soubani, A.O. Pulmonary Aspergillosis: What the Generalist Needs to Know. Am. J. Med. 2020, 133, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W.; Pleuvry, A.; Cole, D.C. Global Burden of Allergic Bronchopulmonary Aspergillosis with Asthma and Its Complication Chronic Pulmonary Aspergillosis in Adults. Med. Mycol. 2013, 51, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Maleki, M.; Mortezaee, V.; Hassanzad, M.; Mahdaviani, S.A.; Poorabdollah, M.; Mehrian, P.; Behnampour, N.; Mirenayat, M.S.; Abastabar, M.; Tavakoli, M.; et al. Prevalence of Allergic Bronchopulmonary Aspergillosis in Cystic Fibrosis Patients Using Two Different Diagnostic Criteria. Eur. Ann. Allergy Clin. Immunol. 2020, 52, 104–111. [Google Scholar] [CrossRef] [PubMed]
- LiPuma, J.J. The Changing Microbial Epidemiology in Cystic Fibrosis. Clin. Microbiol. Rev. 2010, 23, 299–323. [Google Scholar] [CrossRef] [Green Version]
- Ross, B.S. Aspergillus fumigatus in Cystic Fibrosis: Persistence and Adaptation to CF Conditions Through Unique Stress Response Mutation and Convergent Hypoxia Fitness. Ph.D. Thesis, Dartmouth College, Hanover, NH, USA, 2022. [Google Scholar]
- Ballard, E.; Melchers, W.J.G.; Zoll, J.; Brown, A.J.P.; Verweij, P.E.; Warris, A. In-Host Microevolution of Aspergillus fumigatus: A Phenotypic and Genotypic Analysis. Fungal Genet. Biol. 2018, 113, 1–13. [Google Scholar] [CrossRef]
- Denning, D.W.; Cadranel, J.; Beigelman-Aubry, C.; Ader, F.; Chakrabarti, A.; Blot, S.; Ullmann, A.J.; Dimopoulos, G.; Lange, C. Chronic Pulmonary Aspergillosis: Rationale and Clinical Guidelines for Diagnosis and Management. Eur. Respir. J. 2016, 47, 45–68. [Google Scholar] [CrossRef] [Green Version]
- Im, Y.; Jhun, B.W.; Kang, E.-S.; Koh, W.-J.; Jeon, K. Impact of Treatment Duration on Recurrence of Chronic Pulmonary Aspergillosis. J. Infect. 2021, 83, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Howard, S.J.; Pasqualotto, A.C.; Anderson, M.J.; Leatherbarrow, H.; Albarrag, A.M.; Harrison, E.; Gregson, L.; Bowyer, P.; Denning, D.W. Major Variations in Aspergillus fumigatus Arising within Aspergillomas in Chronic Pulmonary Aspergillosis. Mycoses 2013, 56, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, H.; Li, R.; Bu, D.; Wan, Z. Mutations in the Cyp51A Gene and Susceptibility to Itraconazole in Aspergillus fumigatus Serially Isolated from a Patient with Lung Aspergilloma. J. Antimicrob. Chemother. 2005, 55, 31–37. [Google Scholar] [CrossRef]
- Colabardini, A.C.; Wang, F.; Miao, Z.; Pardeshi, L.; Valero, C.; de Castro, P.A.; Akiyama, D.Y.; Tan, K.; Nora, L.C.; Silva-Rocha, R.; et al. Chromatin Profiling Reveals Heterogeneity in Clinical Isolates of the Human Pathogen Aspergillus fumigatus. PLoS Genet. 2022, 18, e1010001. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Heterogeneity Confounds Establishment of “a” Model Microbial Strain. mBio 2017, 8, e00135-17. [Google Scholar] [CrossRef] [Green Version]
- Kirkland, M.E.; Stannard, M.; Kowalski, C.H.; Mould, D.; Caffrey-Carr, A.; Temple, R.M.; Ross, B.S.; Lofgren, L.A.; Stajich, J.E.; Cramer, R.A.; et al. Host Lung Environment Limits Aspergillus fumigatus Germination through an SskA-Dependent Signaling Response. mSphere 2021, 6, e0092221. [Google Scholar] [CrossRef]
- Scully, L.R.; Bidochka, M.J. The Host Acts as a Genetic Bottleneck during Serial Infections: An Insect-Fungal Model System. Curr. Genet. 2006, 50, 335–345. [Google Scholar] [CrossRef]
- Curtis, A.; Binder, U.; Kavanagh, K. Galleria mellonella Larvae as a Model for Investigating Fungal—Host Interactions. Front. Fungal Biol. 2022, 3, 893494. [Google Scholar] [CrossRef]
- Trevijano-Contador, N.; Zaragoza, O. Immune Response of Galleria mellonella against Human Fungal Pathogens. J. Fungi 2018, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.F.; Tansie, S.M.; Shahan, J.R.; Seipelt-Thiemann, R.L.; McClelland, E.E. Serial Passage of Cryptococcus neoformans in Galleria mellonella Results in Increased Capsule and Intracellular Replication in Hemocytes, but Not Increased Resistance to Hydrogen Peroxide. Pathogens 2020, 9, 732. [Google Scholar] [CrossRef]
- Slater, J.L.; Gregson, L.; Denning, D.W.; Warn, P.A. Pathogenicity of Aspergillus fumigatus Mutants Assessed in Galleria mellonella Matches That in Mice. Med. Mycol. 2011, 49 (Suppl. 1), S107–S113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheehan, G.; Clarke, G.; Kavanagh, K. Characterisation of the Cellular and Proteomic Response of Galleria mellonella Larvae to the Development of Invasive Aspergillosis. BMC Microbiol. 2018, 18, 63. [Google Scholar] [CrossRef] [PubMed]
- Margalit, A.; Sheehan, D.; Carolan, J.C.; Kavanagh, K. Exposure to the Pseudomonas Aeruginosa Secretome Alters the Proteome and Secondary Metabolite Production of Aspergillus fumigatus. Microbiology 2022, 168, 001164. [Google Scholar] [CrossRef]
- Margalit, A.; Carolan, J.C.; Sheehan, D.; Kavanagh, K. The Aspergillus fumigatus Secretome Alters the Proteome of Pseudomonas Aeruginosa to Stimulate Bacterial Growth: Implications for Co-Infection. Mol. Cell. Proteom. 2020, 19, 1346–1359. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2021, 50, D543–D552. [Google Scholar] [CrossRef]
- Shin, K.-S.; Park, H.-S.; Kim, Y.-H.; Yu, J.-H. Comparative Proteomic Analyses Reveal That FlbA Down-Regulates GliT Expression and SOD Activity in Aspergillus fumigatus. J. Proteom. 2013, 87, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ponts, N. H2A.Z and Chromatin Remodelling Complexes: A Focus on Fungi. Crit. Rev. Microbiol. 2020, 46, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Wang, Y.; Qi, S.; Gai, K.; He, Q.; Wang, Y. Histone Variant H2A.Z Antagonizes the Positive Effect of the Transcriptional Activator CPC1 to Regulate Catalase-3 Expression under Normal and Oxidative Stress Conditions. Free Radic. Biol. Med. 2018, 121, 136–148. [Google Scholar] [CrossRef]
- Vicentefranqueira, R.; Amich, J.; Marín, L.; Sánchez, C.I.; Leal, F.; Calera, J.A. The Transcription Factor ZafA Regulates the Homeostatic and Adaptive Response to Zinc Starvation in Aspergillus fumigatus. Genes 2018, 9, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Wilkinson, H.H.; Correa, A.; Lewis, Z.A.; Bell-Pedersen, D.; Ebbole, D.J. Transcriptional Response to Glucose Starvation and Functional Analysis of a Glucose Transporter of Neurospora Crassa. Fungal Genet. Biol. 2004, 41, 1104. [Google Scholar] [CrossRef] [PubMed]
- Shekhova, E.; Kniemeyer, O.; Brakhage, A.A. Induction of Mitochondrial Reactive Oxygen Species Production by Itraconazole, Terbinafine, and Amphotericin B as a Mode of Action against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2017, 61, e00978-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, J.; Rowan, R.; McCann, M.; Kavanagh, K. Exposure to Caspofungin Activates Cap and Hog Pathways in Candida Albicans. Med. Mycol. 2009, 47, 697–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Gene Name | Peptides | Sequence Coverage (%) | Score | Fold Change |
|---|---|---|---|---|---|
| Polyadenylation factor subunit CstF64, putative | AFUA_2G09100 | 3 | 18.9 | 17.95 | 3.58 |
| Complex I intermediate associated protein (Cia30), putative | AFUA_3G06220 | 4 | 15.2 | 14.82 | 3.44 |
| KH domain protein | AFUA_4G07220 | 6 | 19.1 | 22.51 | 3.04 |
| Uncharacterized protein | AFUA_3G08440 | 3 | 22.6 | 9.074 | 2.77 |
| 50S ribosomal protein L3 | AFUA_4G06000 | 4 | 20.5 | 19.30 | 2.77 |
| Peptide-methionine (S)-S-oxide reductase | AFUA_2G03140 | 3 | 49.1 | 26.35 | 2.63 |
| Nuclear cap-binding protein subunit 2 | AFUA_2G08570 | 5 | 25 | 7.55 | 2.46 |
| Post-SET domain-containing protein | AFUA_6G10080 | 2 | 23.3 | 8.33 | 2.28 |
| Developmental regulator FlbA | AFUA_2G11180 | 4 | 7.4 | 10.65 | 2.27 |
| Thiamine biosynthetic bifunctional enz, putative | AFUA_2G08970 | 8 | 27.2 | 19.26 | 2.11 |
| RING-type E3 ubiquitin transferase | AFUA_2G11040 | 4 | 29.2 | 6.07 | 1.88 |
| Small nuclear ribonucleoprotein E | AFUA_7G05980 | 4 | 32.6 | 9.08 | 1.81 |
| Phosphatidylglycerol/phosphatidylinositol transfer protein | npc2 | 10 | 35.4 | 29.44 | 1.74 |
| DlpA domain protein | AFUA_4G10940 | 5 | 23.4 | 15.65 | 1.64 |
| RSC complex subunit (RSC1), putative | AFUA_3G05560 | 4 | 7.8 | 9.43 | 1.63 |
| Probable mannosyl-oligosaccharide alpha-1,2-mannosidase 1B | mns1B | 14 | 47.5 | 64.36 | 1.60 |
| ATP-dependent RNA helicase dbp2 | dbp2 | 17 | 43 | 109.23 | −1.53 |
| Rhomboid protein 2 | rbd2 | 2 | 15.4 | 19.36 | −1.69 |
| Integral ER membrane protein Scs2, putative | AFUA_4G06950 | 9 | 40.2 | 53.75 | −1.75 |
| Translation elongation factor eEF-3, putative | AFUA_7G03660 | 68 | 77.1 | 323.31 | −1.76 |
| Guanyl-nucleotide exchange factor (Sec7), putative | AFUA_7G05700 | 8 | 5.8 | 27.80 | −1.76 |
| Histone H2A.Z | htz1 | 6 | 51.4 | 65.54 | −1.82 |
| Glucose repressible protein Grg1, putative | AFUA_5G14210 | 2 | 34.8 | 68.31 | −1.87 |
| Translation initiation regulator (Gcn20), putative | AFUA_4G06070 | 9 | 20.2 | 37.21 | −1.96 |
| ANK_REP_REGION domain-containing protein | AFUA_5G14930 | 20 | 48 | 190.54 | −1.98 |
| Nucleoporin NUP49/NSP49, putative | AFUA_6G10730 | 4 | 10.8 | 10.05 | −2.24 |
| Aminotransferase, putative | AFUA_6G02030 | 6 | 23 | 16.58 | −2.26 |
| GABA permease (Uga4), putative | AFUA_4G03370 | 4 | 8.9 | 12.47 | −2.72 |
| U5 snRNP complex subunit, putative | AFUA_7G02280 | 7 | 32.6 | 19.75 | −3.14 |
| Zinc/cadmium resistance protein | AFUA_2G14570 | 1 | 3 | 9.01 | −5.17 |
| Uncharacterized protein | AFUA_1G16030 | 16 | 38.2 | 62.94 | −6.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curtis, A.; Walshe, K.; Kavanagh, K. Prolonged Subculturing of Aspergillus fumigatus on Galleria Extract Agar Results in Altered Virulence and Sensitivity to Antifungal Agents. Cells 2023, 12, 1065. https://doi.org/10.3390/cells12071065
Curtis A, Walshe K, Kavanagh K. Prolonged Subculturing of Aspergillus fumigatus on Galleria Extract Agar Results in Altered Virulence and Sensitivity to Antifungal Agents. Cells. 2023; 12(7):1065. https://doi.org/10.3390/cells12071065
Chicago/Turabian StyleCurtis, Aaron, Kieran Walshe, and Kevin Kavanagh. 2023. "Prolonged Subculturing of Aspergillus fumigatus on Galleria Extract Agar Results in Altered Virulence and Sensitivity to Antifungal Agents" Cells 12, no. 7: 1065. https://doi.org/10.3390/cells12071065
APA StyleCurtis, A., Walshe, K., & Kavanagh, K. (2023). Prolonged Subculturing of Aspergillus fumigatus on Galleria Extract Agar Results in Altered Virulence and Sensitivity to Antifungal Agents. Cells, 12(7), 1065. https://doi.org/10.3390/cells12071065