
Atherogenesis in Apoe−/− and Ldlr−/− Mice with a Genetically Resistant Background



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Ligation and Morphometric Analysis of Common Carotid Arteries
2.3. Measurement of Primary Atherosclerosis
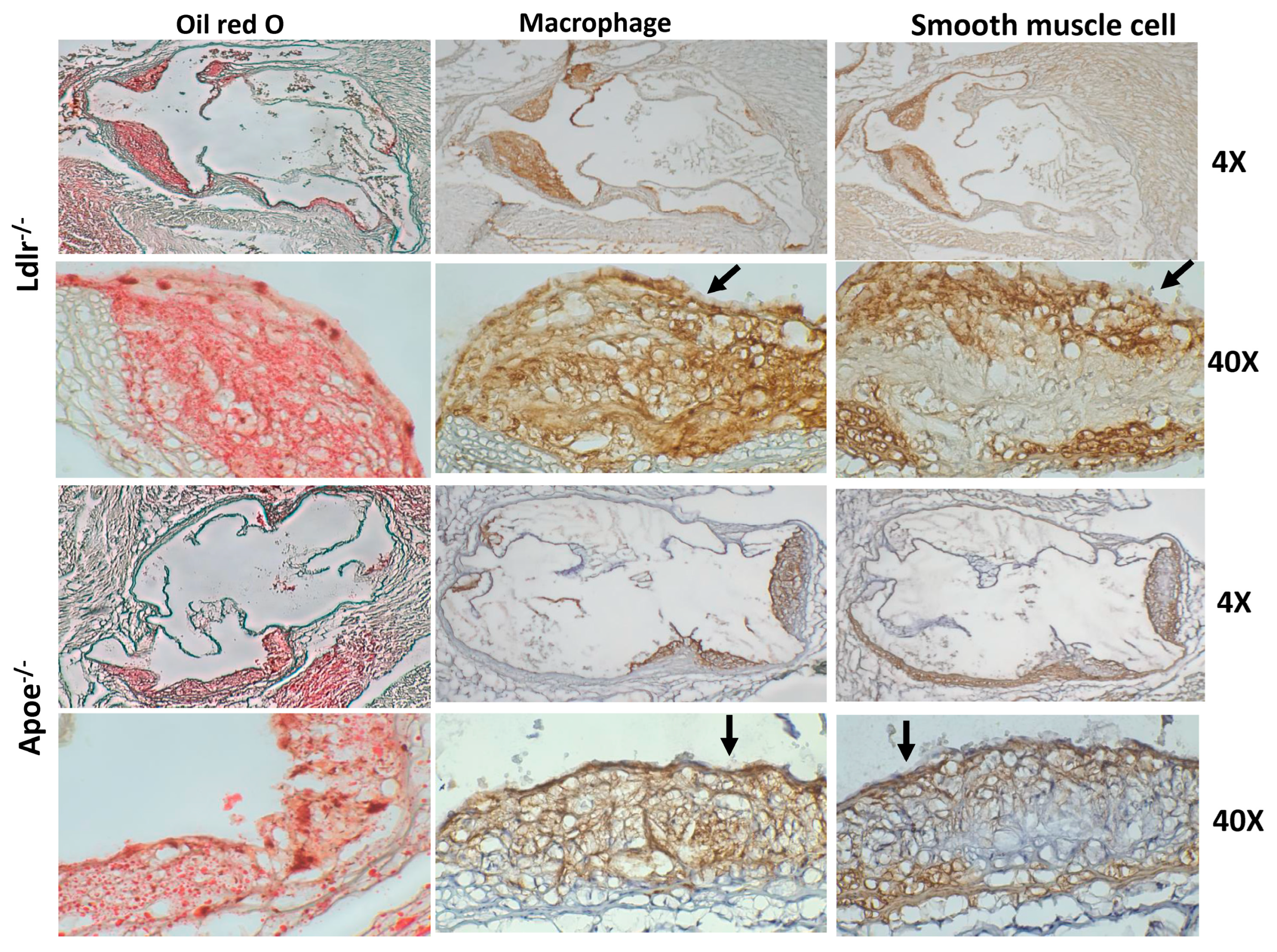
2.4. Immunohistochemical Analysis
2.5. Measurement of Malondialdehyde and MCP-1
2.6. Small Dense LDL Assay
2.7. Statistical Analysis
3. Results
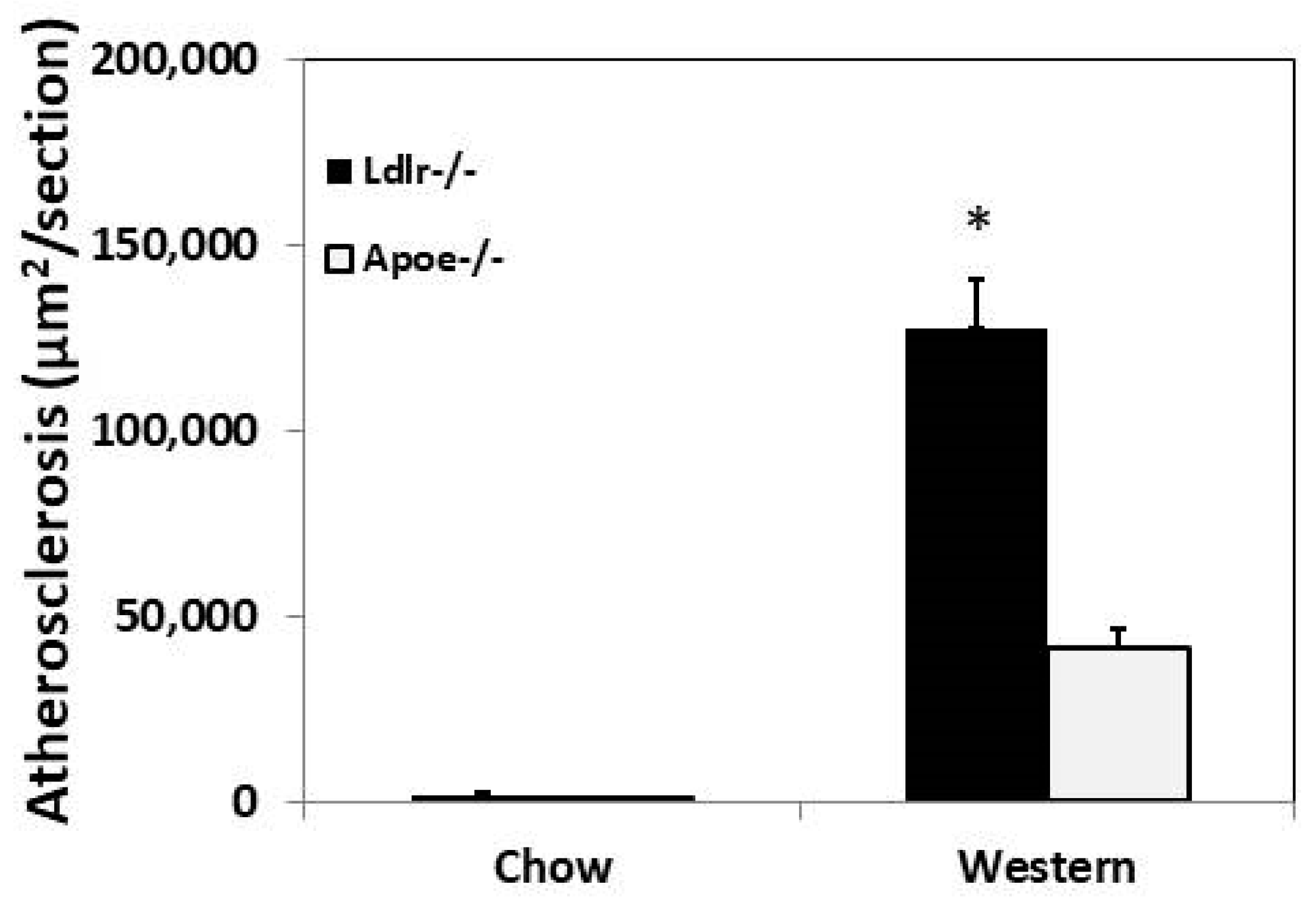
3.1. Primary Atherosclerosis
3.2. Lesion Formation in Ligated Carotid Artery
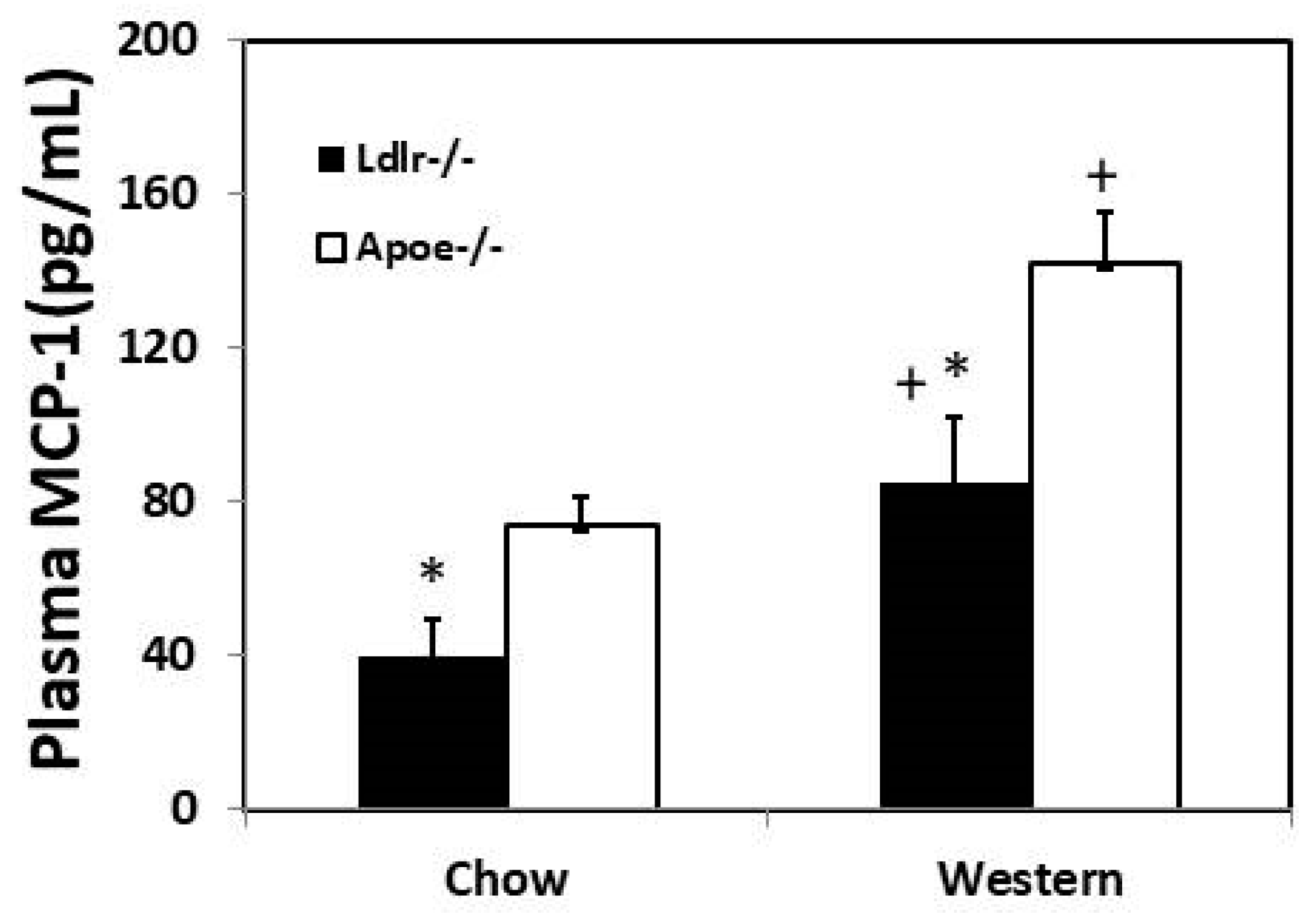
3.3. Plasma MCP-1 Concentration
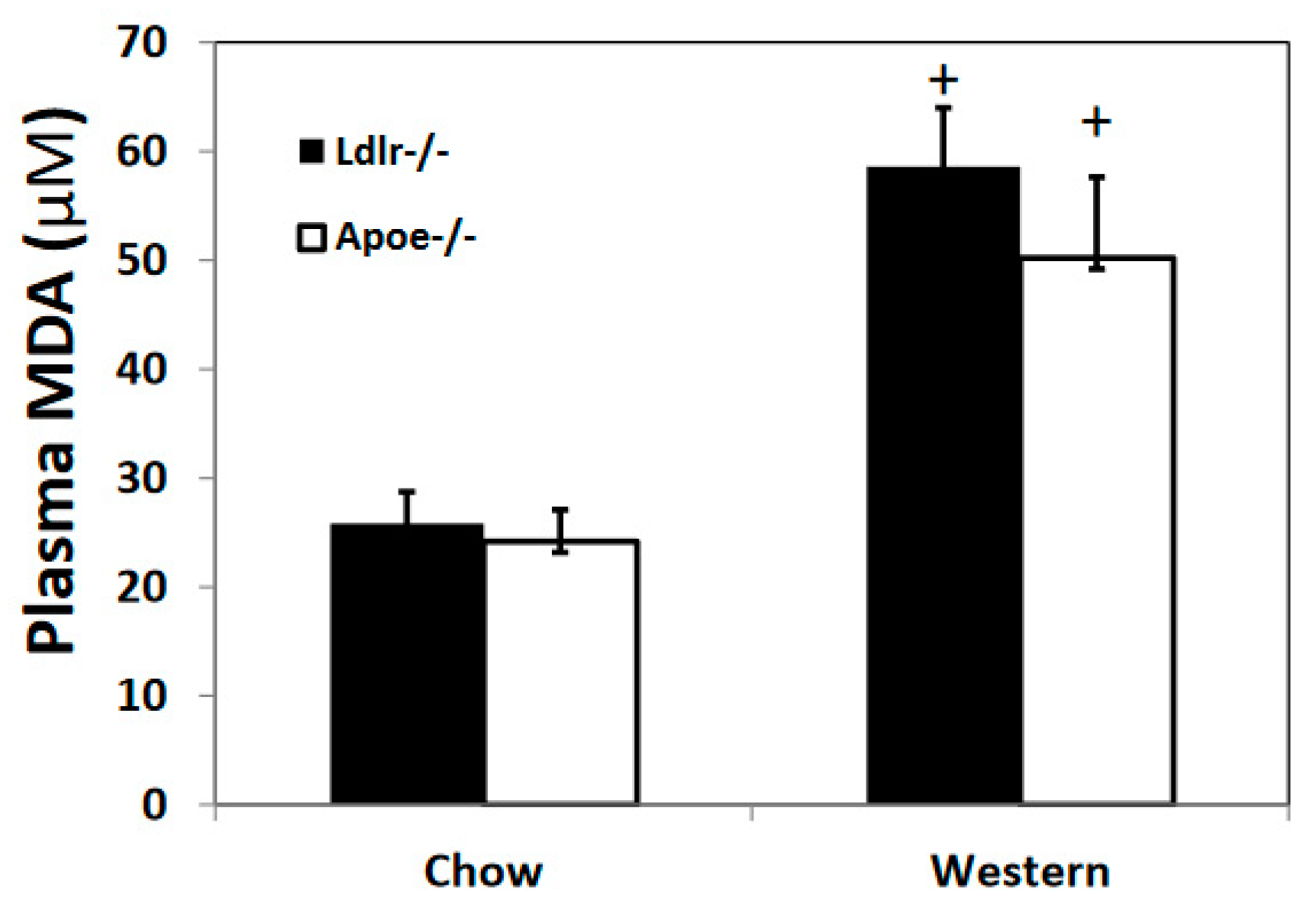
3.4. Oxidative Stress Biomarker
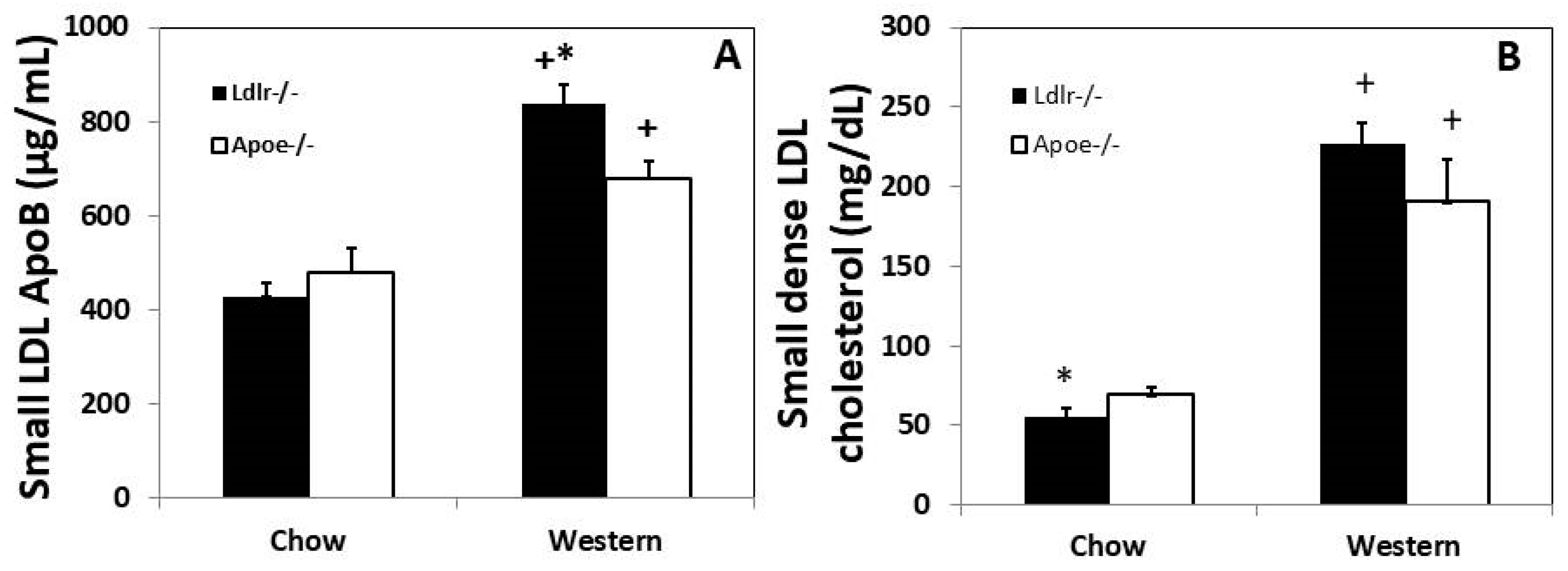
3.5. Plasma Small Dense LDL
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 31 December 2022).
- Ridker, P.M.; Rifai, N.; Cook, N.R.; Bradwin, G.; Buring, J.E. Non-HDL Cholesterol, Apolipoproteins A-I and B100, Standard Lipid Measures, Lipid Ratios, and CRP as Risk Factors for Cardiovascular Disease in Women. JAMA 2005, 294, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Sharrett, A.R.; Ballantyne, C.M.; Coady, S.A.; Heiss, G.; Sorlie, P.D.; Catellier, D.; Patsch, W.; Atherosclerosis Risk in Communities Study Group. Coronary Heart Disease Prediction from Lipoprotein Cholesterol Levels, Triglycerides, Lipoprotein(a), Apolipoproteins A-I and B, and HDL Density Subfractions: The Atherosclerosis Risk in Communities (ARIC) Study. Circulation 2001, 104, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Tcheandjieu, C.; Zhu, X.; Hilliard, A.T.; Clarke, S.L.; Napolioni, V.; Ma, S.; Lee, K.M.; Fang, H.; Chen, F.; Lu, Y.; et al. Large-Scale Genome-Wide Association Study of Coronary Artery Disease in Genetically Diverse Populations. Nat. Med. 2022, 28, 1679–1692. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, T.R.; Kjekshus, J.; Berg, K.; Haghfelt, T.; Faergeman, O.; Faergeman, G.; Pyorala, K.; Miettinen, T.; Wilhelmsen, L.; Olsson, A.G.; et al. Randomised Trial of Cholesterol Lowering in 4444 Patients with Coronary Heart Disease: The Scandinavian Simvastatin Survival Study (4S). Atheroscler. Suppl. 2004, 5, 81–87. [Google Scholar] [CrossRef]
- Chaudhary, R.; Garg, J.; Shah, N.; Sumner, A. PCSK9 Inhibitors: A New Era of Lipid Lowering Therapy. World J. Cardiol. 2017, 9, 76–91. [Google Scholar] [CrossRef]
- Fernández-Friera, L.; Fuster, V.; López-Melgar, B.; Oliva, B.; García-Ruiz, J.M.; Mendiguren, J.; Bueno, H.; Pocock, S.; Ibáñez, B.; Fernández-Ortiz, A.; et al. Normal LDL-Cholesterol Levels Are Associated With Subclinical Atherosclerosis in the Absence of Risk Factors. J. Am. Coll. Cardiol. 2017, 70, 2979–2991. [Google Scholar] [CrossRef]
- Shi, W.; Li, J.; Bao, K.; Chen, M.-H.; Liu, Z. Ldlr-Deficient Mice with an Atherosclerosis-Resistant Background Develop Severe Hyperglycemia and Type 2 Diabetes on a Western-Type Diet. Biomedicines 2022, 10, 1429. [Google Scholar] [CrossRef]
- Véniant, M.M.; Sullivan, M.A.; Kim, S.K.; Ambroziak, P.; Chu, A.; Wilson, M.D.; Hellerstein, M.K.; Rudel, L.L.; Walzem, R.L.; Young, S.G. Defining the Atherogenicity of Large and Small Lipoproteins Containing Apolipoprotein B100. J. Clin. Investig. 2000, 106, 1501–1510. [Google Scholar] [CrossRef]
- Liu, S.; Li, J.; Chen, M.H.; Liu, Z.; Shi, W. Variation in Type 2 Diabetes-Related Phenotypes among Apolipoprotein E-Deficient Mouse Strains. PLoS ONE 2015, 10, e0120935. [Google Scholar] [CrossRef]
- Tian, J.; Pei, H.; Sanders, J.M.; Angle, J.F.; Sarembock, I.J.; Matsumoto, A.H.; Helm, G.A.; Shi, W. Hyperlipidemia Is a Major Determinant of Neointimal Formation in LDL Receptor-Deficient Mice. Biochem. Biophys. Res. Commun. 2006, 345, 1004–1009. [Google Scholar] [CrossRef]
- Getz, G.S.; Reardon, C.A. Do the Apoe−/− and Ldlr−/− Mice Yield the Same Insight on Atherogenesis? Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1734–1741. [Google Scholar] [CrossRef] [PubMed]
- Roselaar, S.E.; Kakkanathu, P.X.; Daugherty, A. Lymphocyte Populations in Atherosclerotic Lesions of ApoE −/− and LDL Receptor −/− Mice. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Sjöland, H.; Eitzman, D.T.; Gordon, D.; Westrick, R.; Nabel, E.G.; Ginsburg, D. Atherosclerosis Progression in LDL Receptor–Deficient and Apolipoprotein E–Deficient Mice Is Independent of Genetic Alterations in Plasminogen Activator Inhibitor-1. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 846–852. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Huangfu, C.; Chang, Z.; Zhou, W.; Grainger, A.T.; Liu, Z.; Shi, W. Inflammation and Enhanced Atherogenesis in the Carotid Artery with Altered Blood Flow in an Atherosclerosis-Resistant Mouse Strain. Physiol. Rep. 2021, 9, e14829. [Google Scholar] [CrossRef]
- Zhao, J.; Huangfu, C.; Chang, Z.; Grainger, A.T.; Liu, Z.; Shi, W. Atherogenesis in the Carotid Artery with and without Interrupted Blood Flow of Two Hyperlipidemic Mouse Strains. J. Vasc. Res. 2019, 56, 241–254. [Google Scholar] [CrossRef]
- Shi, W.; Haberland, M.E.; Jien, M.L.; Shih, D.M.; Lusis, A.J. Endothelial Responses to Oxidized Lipoproteins Determine Genetic Susceptibility to Atherosclerosis in Mice. Circulation 2000, 102, 75–81. [Google Scholar] [CrossRef]
- Grainger, A.T.; Pilar, N.; Li, J.; Chen, M.-H.; Abramson, A.M.; Becker-Pauly, C.; Shi, W. Identification of Mep1a as a Susceptibility Gene for Atherosclerosis in Mice. Genetics 2021, 219, iyab160. [Google Scholar] [CrossRef]
- Chang, Z.; Huangfu, C.; Grainger, A.T.; Zhang, J.; Guo, Q.; Shi, W. Accelerated Atherogenesis in Completely Ligated Common Carotid Artery of Apolipoprotein E-Deficient Mice. Oncotarget 2017, 8, 110289–110299. [Google Scholar] [CrossRef]
- Hirano, T.; Ito, Y.; Saegusa, H.; Yoshino, G. A Novel and Simple Method for Quantification of Small, Dense LDL. J. Lipid. Res. 2003, 44, 2193–2201. [Google Scholar] [CrossRef]
- Sampson, M.; Wolska, A.; Warnick, R.; Lucero, D.; Remaley, A.T. A New Equation Based on the Standard Lipid Panel for Calculating Small Dense Low-Density Lipoprotein-Cholesterol and Its Use as a Risk-Enhancer Test. Clin. Chem. 2021, 67, 987–997. [Google Scholar] [CrossRef]
- Teupser, D.; Persky, A.D.; Breslow, J.L. Induction of Atherosclerosis by Low-Fat, Semisynthetic Diets in LDL Receptor-Deficient C57BL/6J and FVB/NJ Mice: Comparison of Lesions of the Aortic Root, Brachiocephalic Artery, and Whole Aorta (En Face Measurement). Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, W.; Zhang, J.; Lu, Y.; Wu, W.; Yan, H.; Wang, Y. Hyperlipidemia and Atherosclerotic Lesion Development in Ldlr-Deficient Mice on a Long-Term High-Fat Diet. PLoS ONE 2012, 7, e35835. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.; Ni, C.W.; Rezvan, A.; Suo, J.; Budzyn, K.; Llanos, A.; Harrison, D.; Giddens, D.; Jo, H. Partial Carotid Ligation Is a Model of Acutely Induced Disturbed Flow, Leading to Rapid Endothelial Dysfunction and Atherosclerosis. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1535–H1543. [Google Scholar] [CrossRef] [PubMed]
- Liao, F.; Andalibi, A.; deBeer, F.C.; Fogelman, A.M.; Lusis, A.J. Genetic Control of Inflammatory Gene Induction and NF-Kappa B-like Transcription Factor Activation in Response to an Atherogenic Diet in Mice. J. Clin. Investig. 1993, 91, 2572–2579. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Oyama, T.; Sugie, S.; Shimizu, M. Different Susceptibilities between Apoe- and Ldlr-Deficient Mice to Inflammation-Associated Colorectal Carcinogenesis. Int. J. Mol. Sci. 2016, 17, 1806. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Ackermann, S.; Ma, Z.; Mohanta, S.K.; Zhang, C.; Li, Y.; Nietzsche, S.; Westermann, M.; Peng, L.; Hu, D.; et al. ApoE Attenuates Unresolvable Inflammation by Complex Formation with Activated C1q. Nat. Med. 2019, 25, 496–506. [Google Scholar] [CrossRef]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Ishigami, M.; Swertfeger, D.K.; Hui, M.S.; Granholm, N.A.; Hui, D.Y. Apolipoprotein E Inhibition of Vascular Smooth Muscle Cell Proliferation but Not the Inhibition of Migration Is Mediated Through Activation of Inducible Nitric Oxide Synthase. Arter. Thromb. Vasc. Biol. 2000, 20, 1020–1026. [Google Scholar] [CrossRef]
- Fuller, D.T.; Grainger, A.T.; Manichaikul, A.; Shi, W. Genetic linkage of oxidative stress with cardiometabolic traits in an intercross derived from hyperlipidemic mouse strains. Atherosclerosis 2019, 293, 1–10. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Tribble, D.L.; Rizzo, M.; Chait, A.; Lewis, D.M.; Blanche, P.J.; Krauss, R.M. Enhanced oxidative susceptibility and reduced antioxidant content of metabolic precursors of small, dense low-density lipoproteins. Am. J. Med. 2001, 110, 103–110. [Google Scholar] [CrossRef]
- Yan, X.; Kuipers, F.; Havekes, L.M.; Havinga, R.; Dontje, B.; Poelstra, K.; Scherphof, G.L.; Kamps, J.A. The role of apolipoprotein E in the elimination of liposomes from blood by hepatocytes in the mouse. Biochem. Biophys. Res. Commun. 2005, 328, 57–62. [Google Scholar] [CrossRef]
- Kowal, R.C.; Herz, J.; Goldstein, J.L.; Esser, V.; Brown, M.S. Low density lipoprotein receptor-related protein mediates uptake of cholesteryl esters derived from apoprotein E-enriched lipoproteins. Proc. Natl. Acad. Sci. USA 1989, 86, 5810–5814. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, S.; Brown, M.S.; Goldstein, J.L.; Gerard, R.D.; E Hammer, R.; Herz, J. Hypercholesterolemia in low density lipoprotein receptor knockout mice and its reversal by adenovirus-mediated gene delivery. J. Clin. Investig. 1993, 92, 883–893. [Google Scholar] [CrossRef]
- Zsigmond, E.; Fuke, Y.; Li, L.; Kobayashi, K.; Chan, L. Resistance of chylomicron and VLDL remnants to post-heparin lipolysis in ApoE-deficient mice: The role of apoE in lipoprotein lipase-mediated lipolysis in vivo and in vitro. J. Lipid Res. 1998, 39, 1852–1861. [Google Scholar] [CrossRef] [PubMed]
- Harmon, K.J.; Couper, L.L.; Lindner, V. Strain-Dependent Vascular Remodeling Phenotypes in Inbred Mice. Am. J. Pathol. 2000, 156, 1741–1748. [Google Scholar] [CrossRef]
- Bonthu, S.; Heistad, D.D.; Chappell, D.A.; Lamping, K.G.; Faraci, F. Atherosclerosis, Vascular Remodeling, and Impairment of Endothelium-Dependent Relaxation in Genetically Altered Hyperlipidemic Mice. Arter. Thromb. Vasc. Biol. 1997, 17, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Lindner, V. Remodeling with Neointima Formation in the Mouse Carotid Artery After Cessation of Blood Flow. Arter. Thromb. Vasc. Biol. 1997, 17, 2238–2244. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Brown, M.D.; Wang, X.; Wong, J.; Kallmes, D.F.; Matsumoto, A.H.; Helm, G.A.; Drake, T.A.; Lusis, A.J. Genetic Backgrounds but Not Sizes of Atherosclerotic Lesions Determine Medial Destruction in the Aortic Root of Apolipoprotein E–Deficient Mice. Arter. Thromb. Vasc. Biol. 2003, 23, 1901–1906. [Google Scholar] [CrossRef]
- Varnava, A.M.; Mills, P.G.; Davies, M.J. Relationship Between Coronary Artery Remodeling and Plaque Vulnerability. Circulation 2002, 105, 939–943. [Google Scholar] [CrossRef]
- Ota, H.; Magalhaes, M.A.; Torguson, R.; Negi, S.; Kollmer, M.R.; Spad, M.-A.; Gai, J.; Satler, L.F.; Suddath, W.O.; Pichard, A.D.; et al. The influence of lipid-containing plaque composition assessed by near-infrared spectroscopy on coronary lesion remodelling. Eur. Hear. J. Cardiovasc. Imaging 2015, 17, 821–831. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torikai, H.; Chen, M.-H.; Jin, L.; He, J.; Angle, J.F.; Shi, W. Atherogenesis in Apoe−/− and Ldlr−/− Mice with a Genetically Resistant Background. Cells 2023, 12, 1255. https://doi.org/10.3390/cells12091255
Torikai H, Chen M-H, Jin L, He J, Angle JF, Shi W. Atherogenesis in Apoe−/− and Ldlr−/− Mice with a Genetically Resistant Background. Cells. 2023; 12(9):1255. https://doi.org/10.3390/cells12091255
Chicago/Turabian StyleTorikai, Hideyuki, Mei-Hua Chen, Li Jin, Jiang He, John F. Angle, and Weibin Shi. 2023. "Atherogenesis in Apoe−/− and Ldlr−/− Mice with a Genetically Resistant Background" Cells 12, no. 9: 1255. https://doi.org/10.3390/cells12091255
APA StyleTorikai, H., Chen, M. -H., Jin, L., He, J., Angle, J. F., & Shi, W. (2023). Atherogenesis in Apoe−/− and Ldlr−/− Mice with a Genetically Resistant Background. Cells, 12(9), 1255. https://doi.org/10.3390/cells12091255