


NET Proteome in Established Type 1 Diabetes Is Enriched in Metabolic Proteins

 , and
, and
Abstract
:1. Introduction
2. Research Design and Methods
2.1. Human Subjects and Ethics Statement
2.2. Isolation of Primary Human Neutrophils
2.3. Quantification and Visualization of NETs by Immunofluorescence
2.4. Plasma NET Markers Measurements
2.5. Multiplex Cytokine Assays
2.6. NET Protein Harvesting
2.7. LC-MS/MS Analysis
2.8. Bioenergetic Profile of Neutrophils
2.9. Lactate Measurements
2.10. Bioinformatic and Statistical Analyses
3. Results
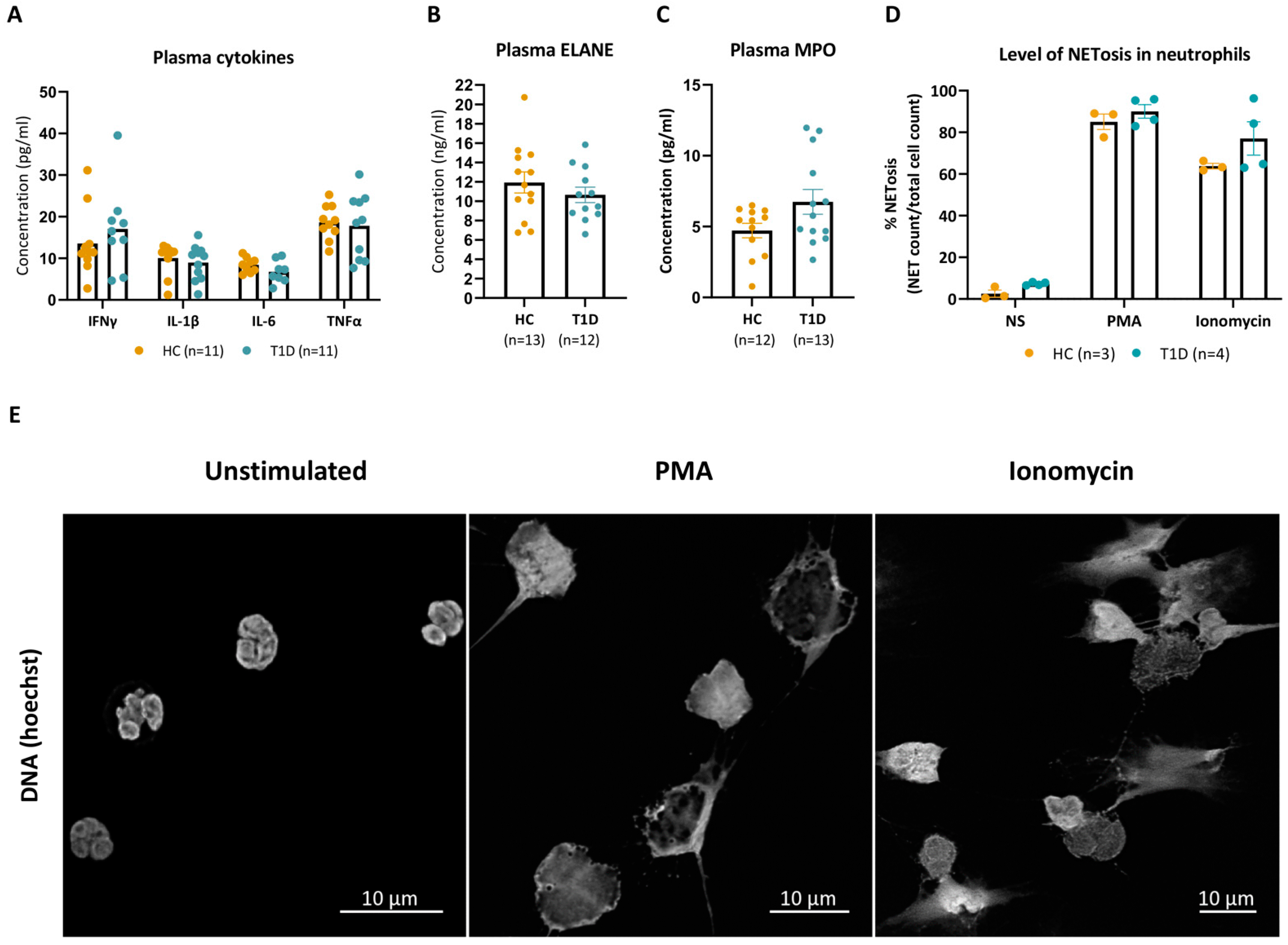
3.1. PMA and Ionomycin Induce Comparable Levels of NETosis in Neutrophils Isolated from T1D and HC Donors
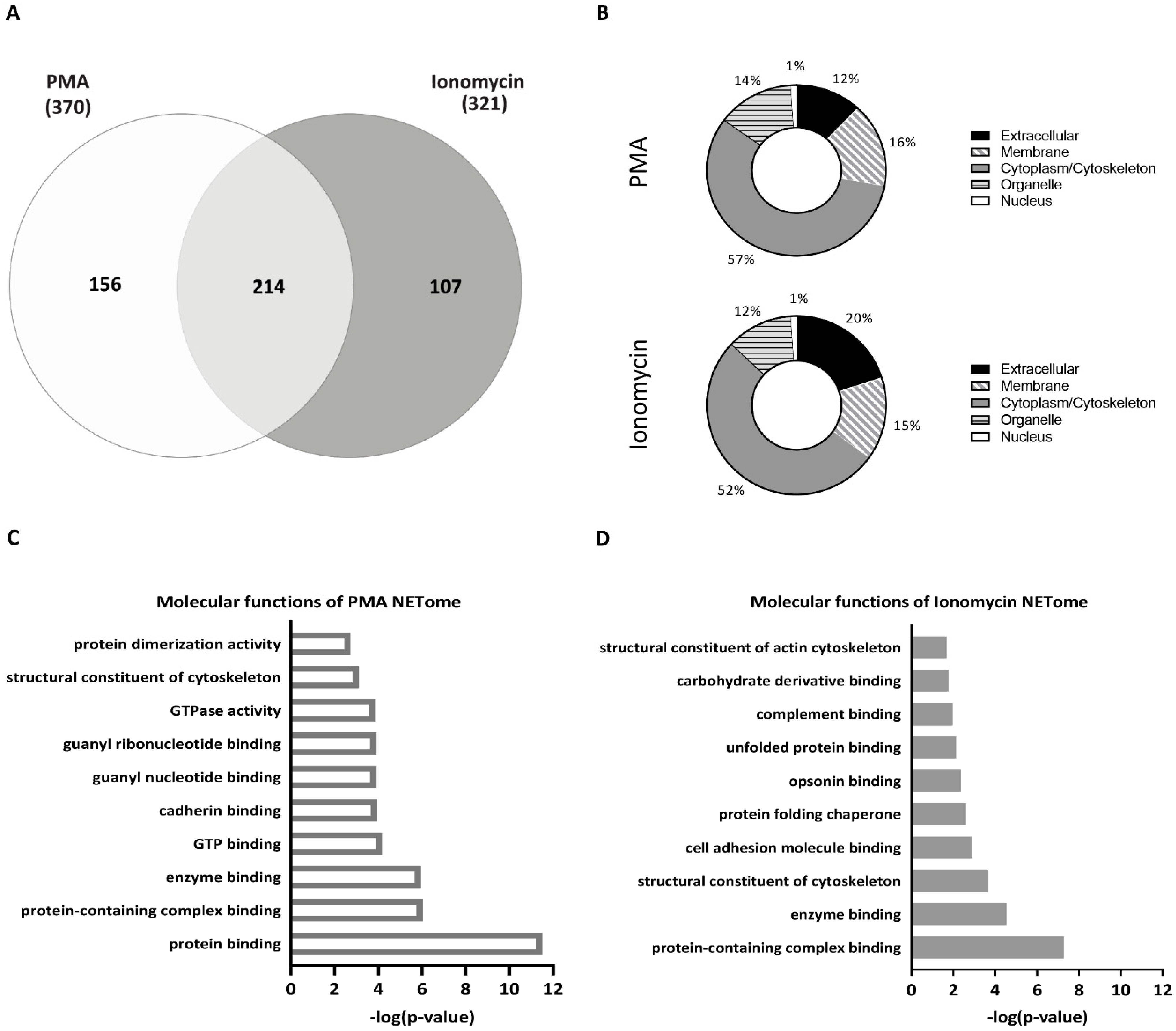
3.2. NET Proteomes of Peripheral Neutrophils Are Different upon PMA and Ionomycin Stimulation
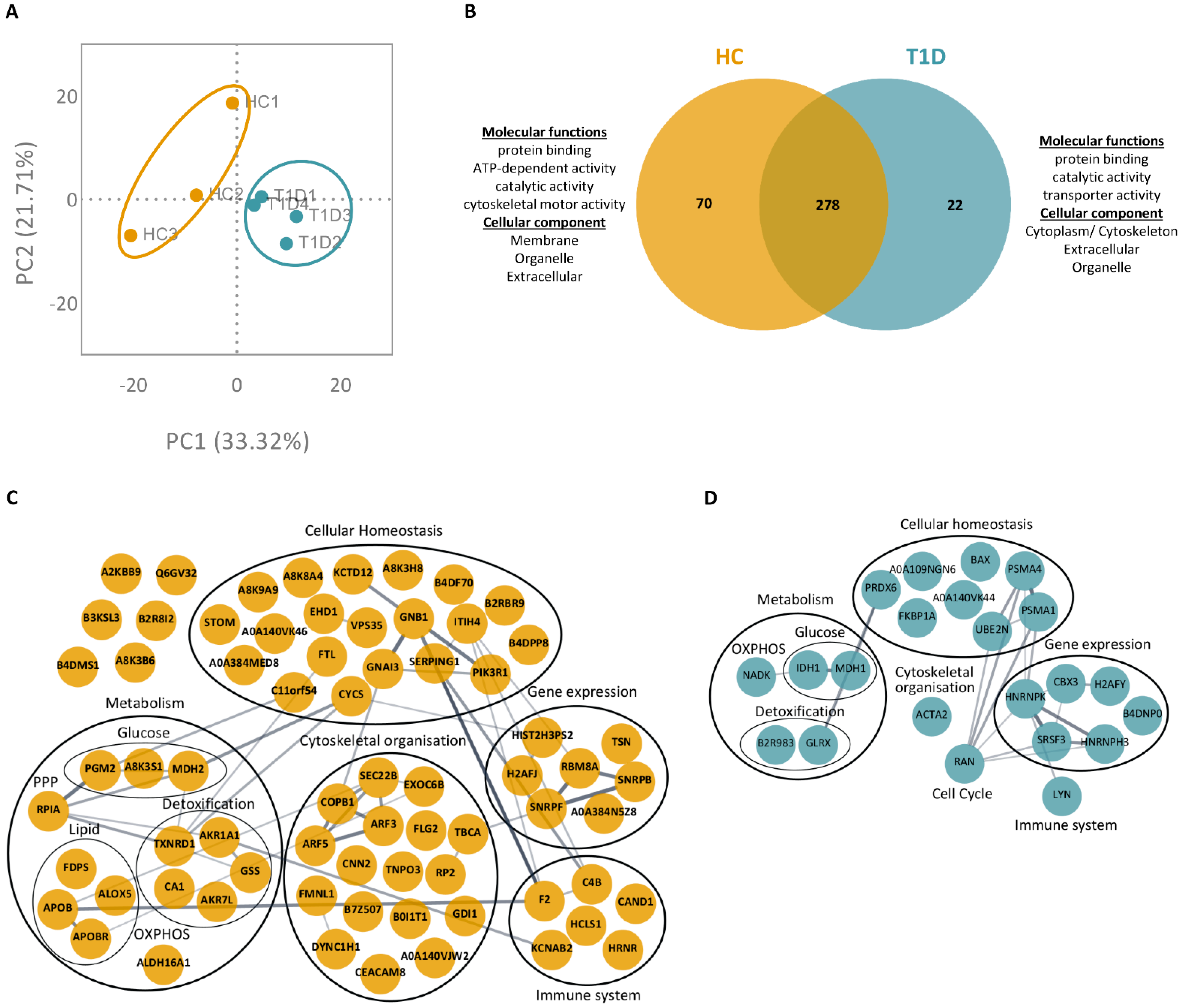
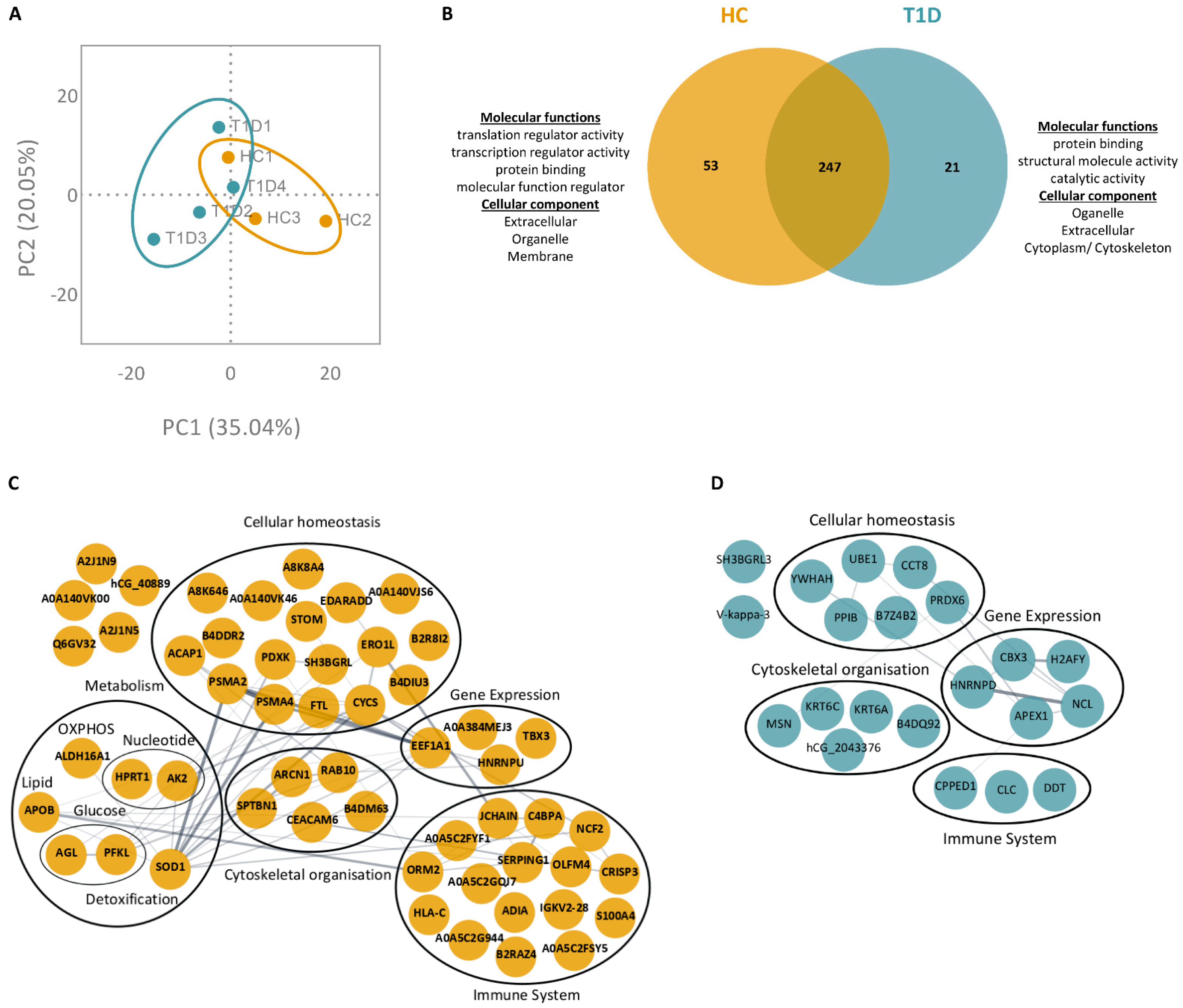
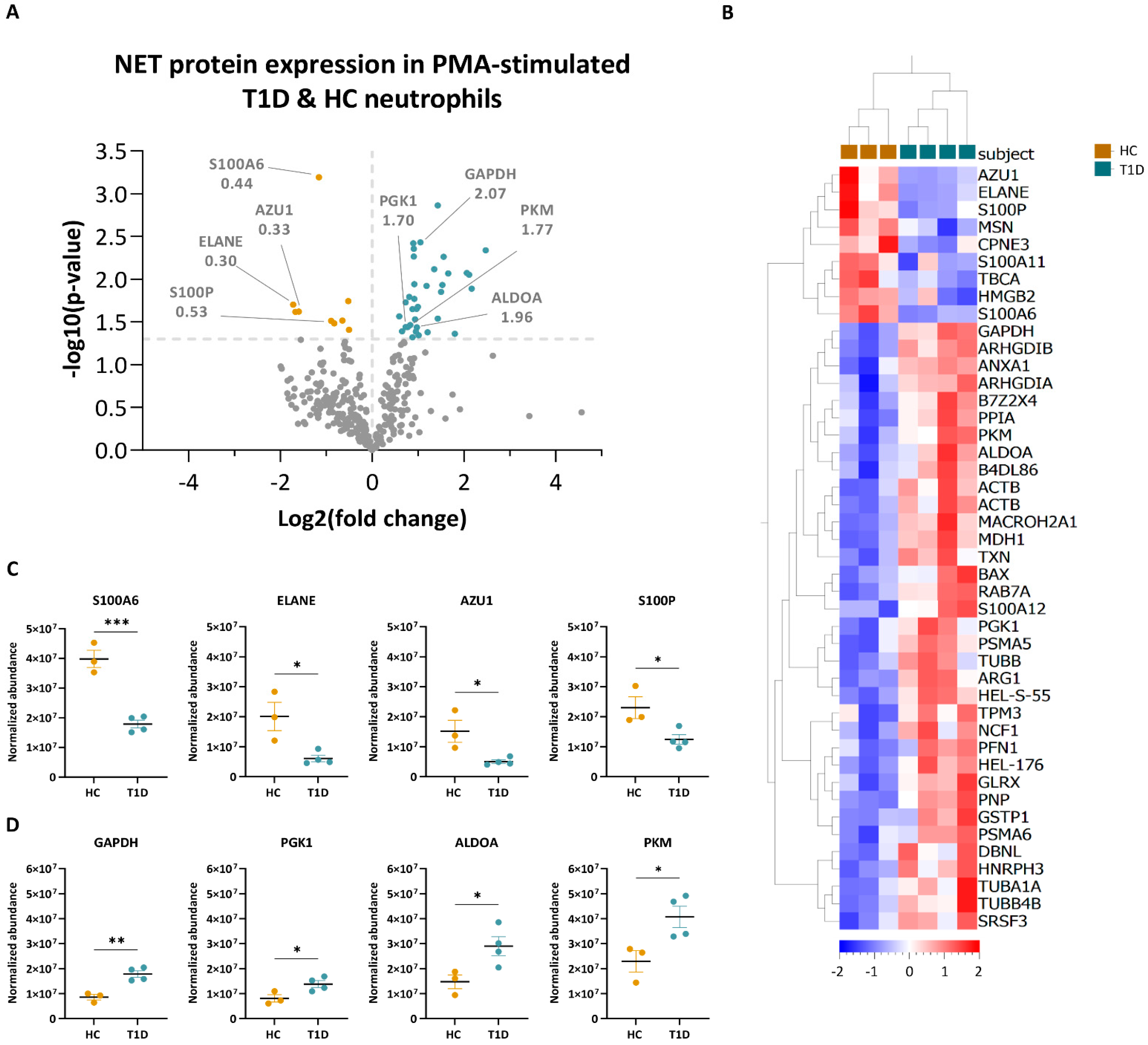
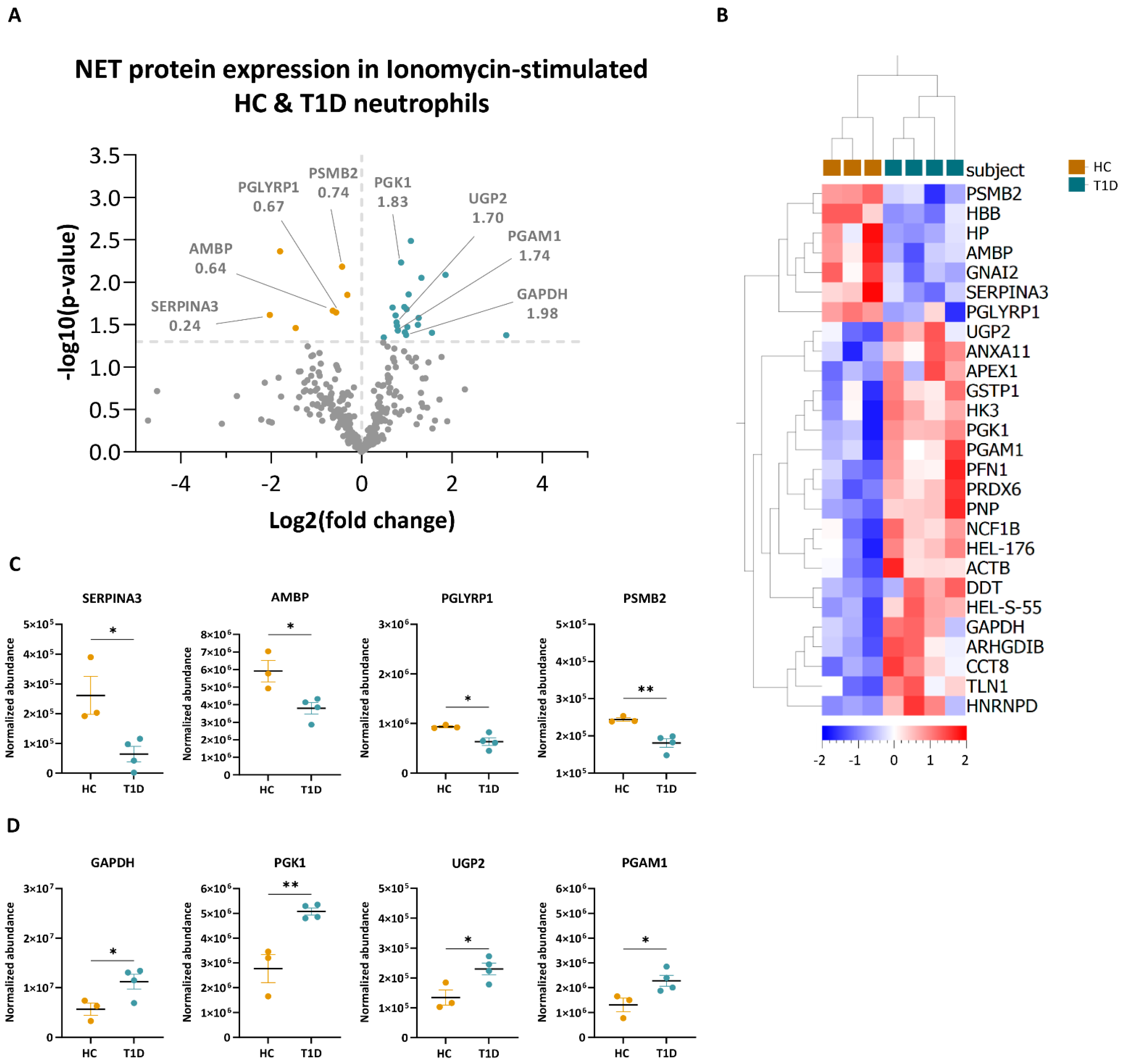
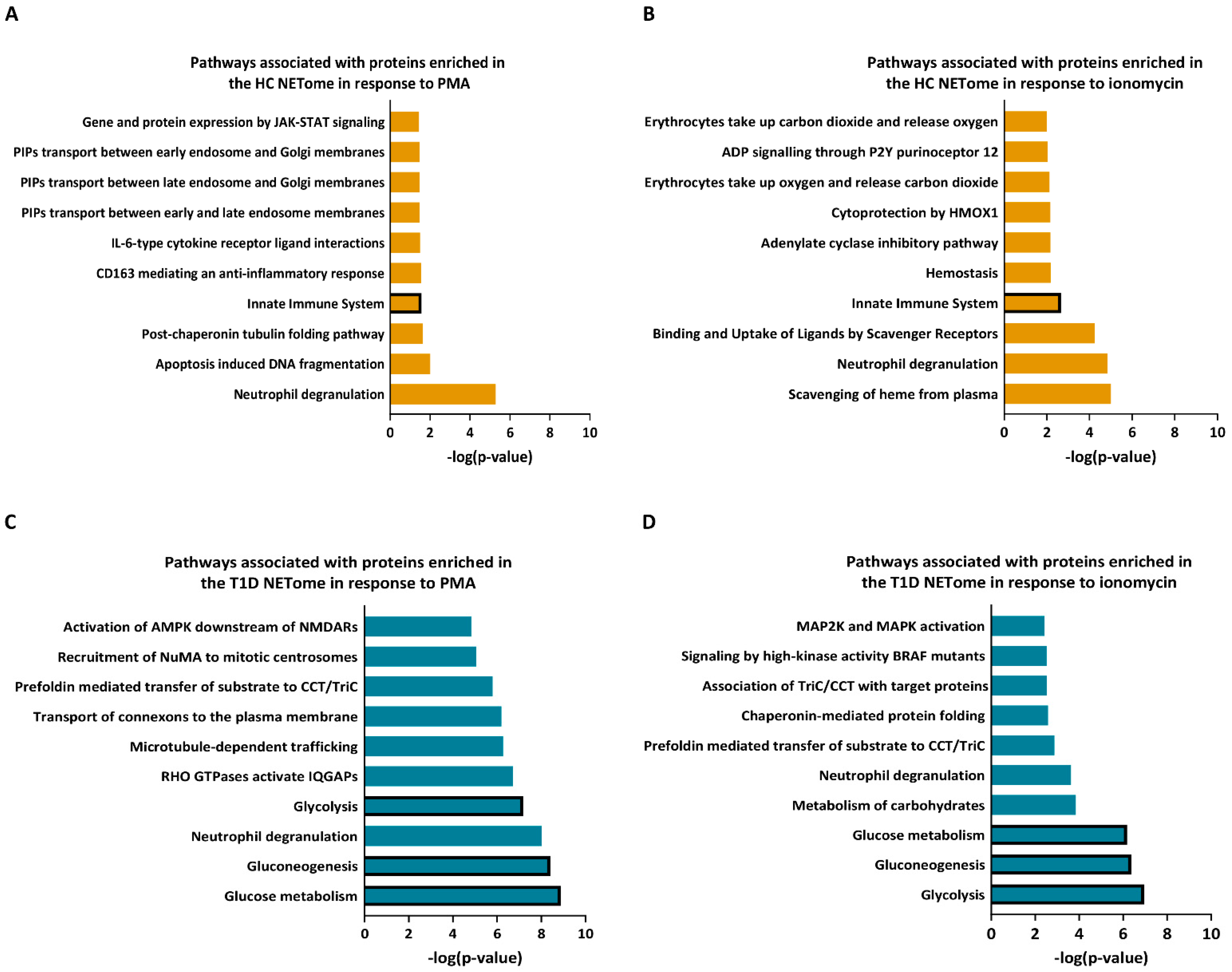
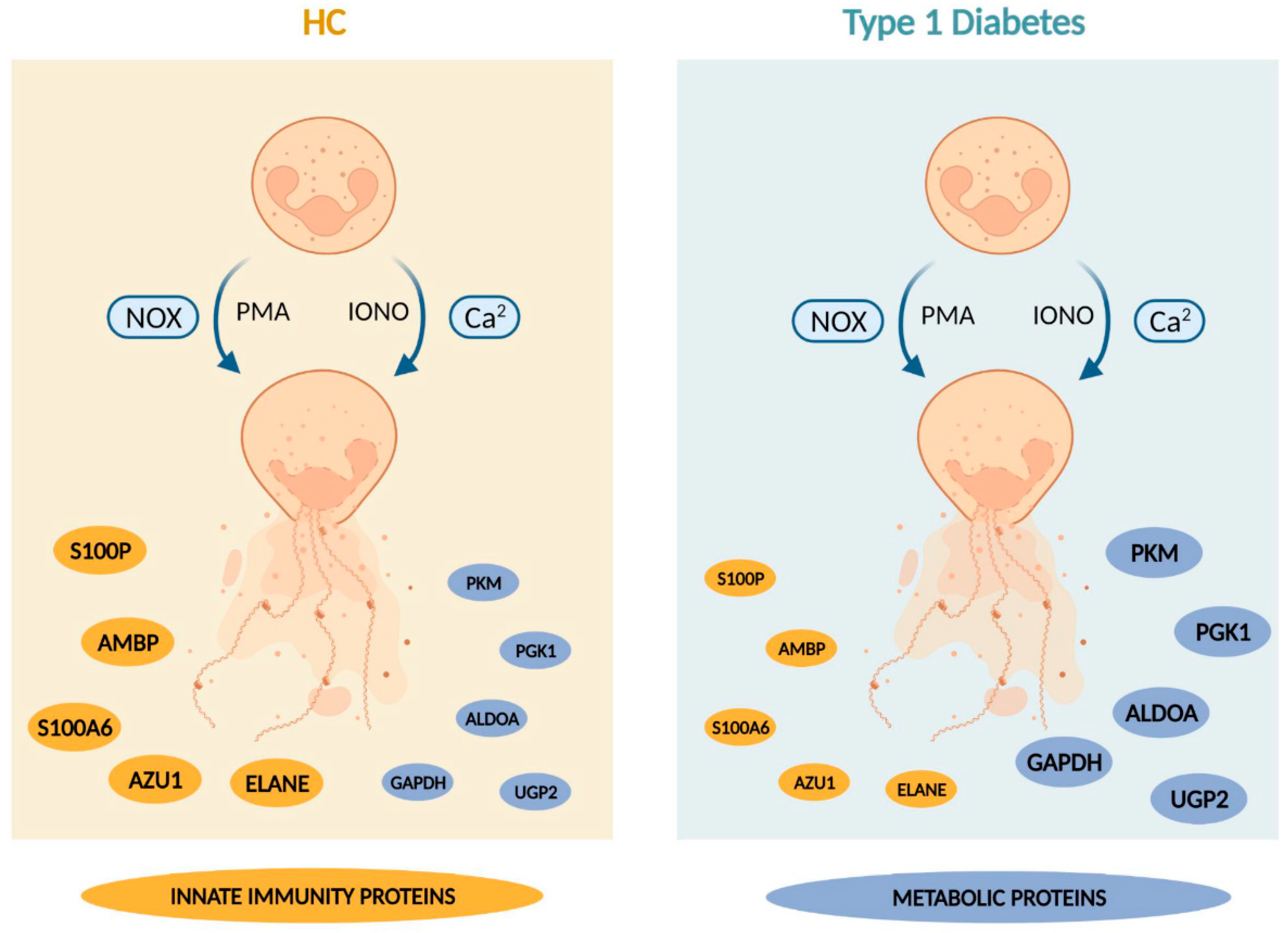
3.3. PMA- and Ionomycin-Induced NET Proteomes of T1D Neutrophils Are Distinct from Those of HC
3.4. Enzymes Involved in the Glucose Metabolism Pathway Are More Abundant in the NET Proteomes of T1D Subjects Compared to Those of HC Subjects
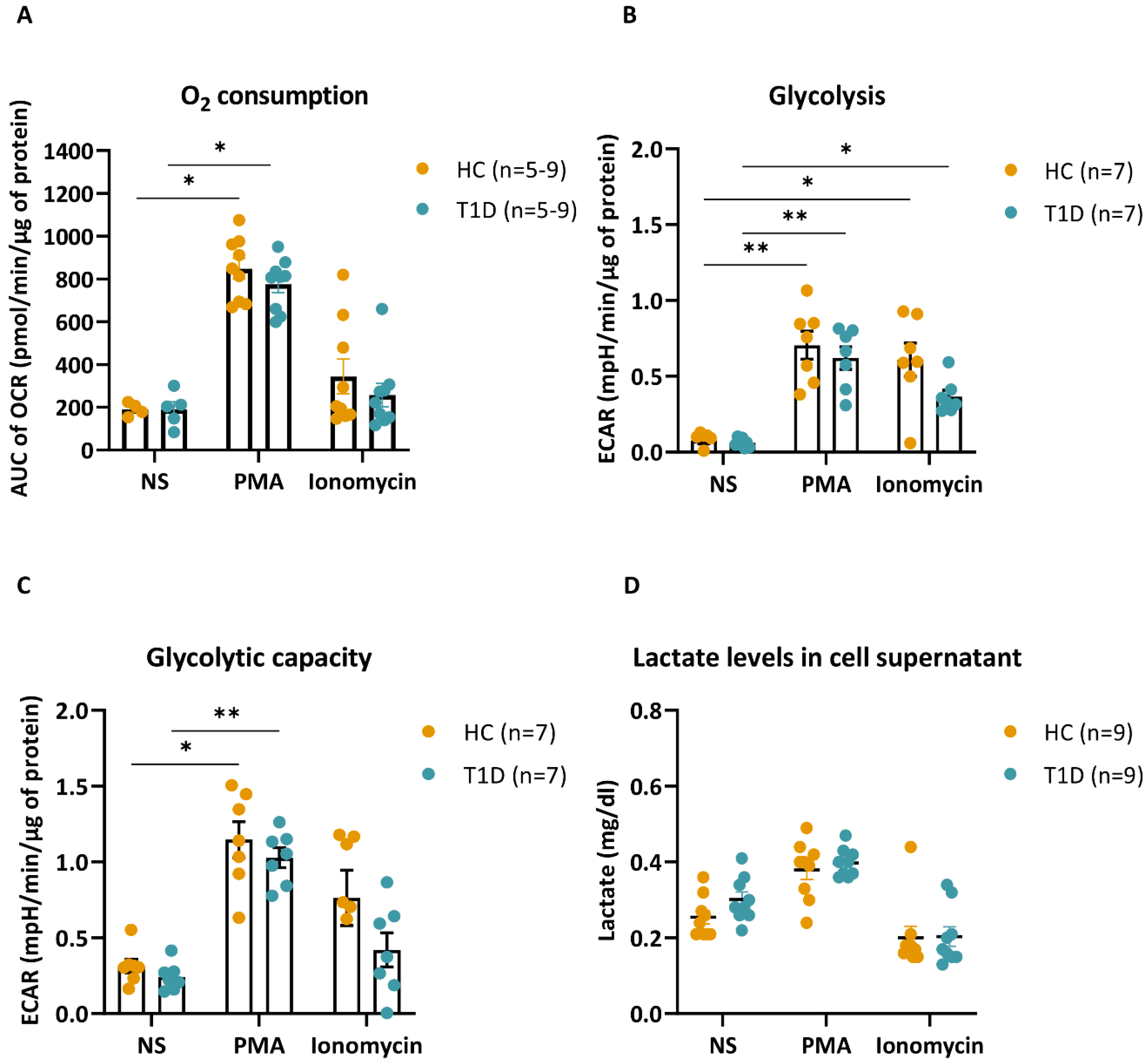
3.5. PMA- and Ionomycin-Stimulated Neutrophils of T1D Subjects Have Similar Metabolic Profiles as HC
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AAV | ANCA-associated vasculitis |
| AGL | 4-alpha-glucanotransferase |
| ALDH16A1 | Aldehyde dehydrogenase family 16 member A1 |
| ALDOA | Fructose-bisphosphate aldolase A |
| ALOX5 | Polyunsaturated fatty acid 5-lipoxygenase |
| AMBP | Protein AMBP |
| ANCA | Anti-cytoplasmic antibody |
| APOB | Apolipoprotein B |
| APOBR | Apolipoprotein B receptor |
| AZU1 | Azurocidin |
| ECAR | Extracellular acidification rate |
| FAO | Fatty acid oxidation |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GO | Gene ontology |
| HC | Healthy control |
| IDH1 | Isocitrate dehydrogenase |
| IL-1β | Interleukin 1 beta |
| IL-6 | Interleukin 6 |
| LC-MS/MS | Liquid chromatography tandem mass spectrometry |
| LPS | Lipopolysaccharide |
| MPO | Myeloperoxidase |
| NADK | Nicotinamide adenine dinucleotide kinase |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| ELANE | Neutrophil elastase |
| NETome | NET proteome |
| NOD | Non-obese diabetic |
| OCR | Oxygen consumption rate |
| OXPHOS | Oxidative phosphorylation |
| PADI | Peptidyl arginine deiminase |
| PFKL | ATP-dependent 6-phosphofructokinase |
| PGAM1 | Phosphoglycerate mutase 1 |
| PGK1 | Phosphoglycerate kinase |
| PGLYRP1 | Peptidoglycan recognition protein 1 |
| PGM2 | Phosphoglucomutase-2 |
| PKM | Pyruvate kinase PKM |
| PMA | Phorbol 12-myristate 13-acetate |
| PR3 | Proteinase 3 |
| PSMB2 | Proteasome subunit beta |
| RA | Rheumatoid arthritis |
| S100A6 | Protein S100-A6 |
| S100P | Protein S100-P |
| SERPINA3 | Alpha-1-antichymotrypsin |
| SLE | Systemic lupus erythematosus |
| T1D | Type 1 diabetes |
| TCA | Tricarboxylic acid |
| TNF-α | Tumor necrosis factor alpha |
| UGP2 | UTP-glucose-1-phosphate uridylyltransferase |
References
- Zirpel, H.; Roep, B.O. Islet-Resident Dendritic Cells and Macrophages in Type 1 Diabetes: In Search of Bigfoot’s Print. Front. Endocrinol. 2021, 12, 666795. [Google Scholar] [CrossRef]
- Vecchio, F.; Buono, N.L.; Stabilini, A.; Nigi, L.; Dufort, M.J.; Geyer, S.; Rancoita, P.M.; Cugnata, F.; Mandelli, A.; Valle, A.; et al. Abnormal neutrophil signature in the blood and pancreas of presymptomatic and symptomatic type 1 diabetes. JCI Insight 2018, 3, 122146. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, Y.; Zhong, L.; Ye, D.; Zhang, J.; Tu, Y.; Bornstein, S.R.; Zhou, Z.; Lam, K.S.L.; Xu, A. Increased Neutrophil Elastase and Proteinase 3 and Augmented NETosis Are Closely Associated With β-Cell Autoimmunity in Patients with Type 1 Diabetes. Diabetes 2014, 63, 4239–4248. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.L.; Demers, M.; Martinod, K.; Gallant, M.; Wang, Y.; Goldfine, A.B.; Kahn, C.R.; Wagner, D.D. Diabetes primes neutrophils to undergo NETosis which severely impairs wound healing. Nat. Med. 2015, 21, 815–819. [Google Scholar] [CrossRef]
- Aukrust, S.G.; Holte, K.B.; Opstad, T.B.; Seljeflot, I.; Berg, T.J.; Helseth, R. NETosis in Long-Term Type 1 Diabetes Mellitus and Its Link to Coronary Artery Disease. Front. Immunol. 2021, 12, 799539. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.G.; Ostuni, R.; Hidalgo, A. Heterogeneity of neutrophils. Nat. Rev. Immunol. 2019, 19, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef]
- He, Y.; Yang, F.-Y.; Sun, E.-W. Neutrophil Extracellular Traps in Autoimmune Diseases. Chin. Med. J. 2018, 131, 1513–1519. [Google Scholar] [CrossRef]
- Jorch, S.K.; Kubes, P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nat. Med. 2017, 23, 279–287. [Google Scholar] [CrossRef]
- Skoglund, C.; Appelgren, D.; Johansson, I.; Casas, R.; Ludvigsson, J. Increase of Neutrophil Extracellular Traps, Mitochondrial DNA and Nuclear DNA in Newly Diagnosed Type 1 Diabetes Children but Not in High-Risk Children. Front. Immunol. 2021, 12, 628564. [Google Scholar] [CrossRef]
- Parackova, Z.; Zentsova, I.; Vrabcova, P.; Klocperk, A.; Sumnik, Z.; Pruhova, S.; Petruzelkova, L.; Hasler, R.; Sediva, A. Neutrophil Extracellular Trap Induced Dendritic Cell Activation Leads to Th1 Polarization in Type 1 Diabetes. Front. Immunol. 2020, 11, 661. [Google Scholar] [CrossRef]
- Kenny, E.F.; Herzig, A.; Krüger, R.; Muth, A.; Mondal, S.; Thompson, P.R.; Brinkmann, V.; von Bernuth, H.; Zychlinsky, A. Diverse stimuli engage different neutrophil extracellular trap pathways. eLife 2017, 6, e24437. [Google Scholar] [CrossRef] [PubMed]
- Parker, H.; Dragunow, M.; Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Requirements for NADPH oxidase and myeloperoxidase in neutrophil extracellular trap formation differ depending on the stimulus. J. Leukoc. Biol. 2012, 92, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Douda, D.N.; Khan, M.A.; Grasemann, H.; Palaniyar, N. SK3 channel and mitochondrial ROS mediate NADPH oxidase-independent NETosis induced by calcium influx. Proc. Natl. Acad. Sci. USA 2015, 112, 2817–2822. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.A.; Lyon, M.; Simpson, D.; Mason, D.; Beynon, R.J.; Moots, R.J.; Wright, H.L. Caught in a Trap? Proteomic Analysis of Neutrophil Extracellular Traps in Rheumatoid Arthritis and Systemic Lupus Erythematosus. Front. Immunol. 2019, 10, 423. [Google Scholar] [CrossRef] [PubMed]
- Bruschi, M.; Petretto, A.; Santucci, L.; Vaglio, A.; Pratesi, F.; Migliorini, P.; Bertelli, R.; Lavarello, C.; Bartolucci, M.; Candiano, G.; et al. Neutrophil Extracellular Traps protein composition is specific for patients with Lupus nephritis and includes methyl-oxidized αenolase (methionine sulfoxide 93). Sci. Rep. 2019, 9, 7934. [Google Scholar] [CrossRef]
- Buitinga, M.; Callebaut, A.; Marques Câmara Sodré, F.; Crèvecoeur, I.; Blahnik-Fagan, G.; Yang, M.-L.; Bugliani, M.; Arribas-Layton, D.; Marré, M.; Cook, D.P.; et al. Inflammation-Induced Citrullinated Glucose-Regulated Protein 78 Elicits Immune Responses in Human Type 1 Diabetes. Diabetes 2018, 67, 2337–2348. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.-P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Njeim, R.; Azar, W.S.; Fares, A.H.; Azar, S.T.; Kfoury Kassouf, H.; Eid, A.A. NETosis contributes to the pathogenesis of diabetes and its complications. J. Mol. Endocrinol. 2020, 65, R65–R76. [Google Scholar] [CrossRef]
- Berezin, A. Neutrophil extracellular traps: The core player in vascular complications of diabetes mellitus. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 3017–3023. [Google Scholar] [CrossRef] [PubMed]
- Garciafigueroa, Y.; Phillips, B.E.; Engman, C.; Trucco, M.; Giannoukakis, N. Neutrophil-Associated Inflammatory Changes in the Pre-Diabetic Pancreas of Early-Age NOD Mice. Front. Endocrinol. 2021, 12, 565981. [Google Scholar] [CrossRef]
- Stojkov, D.; Gigon, L.; Peng, S.; Lukowski, R.; Ruth, P.; Karaulov, A.; Rizvanov, A.; Barlev, N.A.; Yousefi, S.; Simon, H.-U. Physiological and Pathophysiological Roles of Metabolic Pathways for NET Formation and Other Neutrophil Functions. Front. Immunol. 2022, 13, 826515. [Google Scholar] [CrossRef] [PubMed]
- Klocperk, A.; Vcelakova, J.; Vrabcova, P.; Zentsova, I.; Petruzelkova, L.; Sumnik, Z.; Pruhova, S.; Sediva, A.; Parackova, Z. Elevated Biomarkers of NETosis in the Serum of Pediatric Patients with Type 1 Diabetes and Their First-Degree Relatives. Front. Immunol. 2021, 12, 699386. [Google Scholar] [CrossRef]
- Valle, A.; Giamporcaro, G.M.; Scavini, M.; Stabilini, A.; Grogan, P.; Bianconi, E.; Sebastiani, G.; Masini, M.; Maugeri, N.; Porretti, L.; et al. Reduction of circulating neutrophils precedes and accompanies type 1 diabetes. Diabetes 2013, 62, 2072–2077. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yao, Y.; Sumi, Y.; Li, A.; To, U.K.; Elkhal, A.; Inoue, Y.; Woehrle, T.; Zhang, Q.; Hauser, C.; et al. Purinergic Signaling: A Fundamental Mechanism in Neutrophil Activation. Sci. Signal. 2010, 3, ra45. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, D. Purinergic Regulation of Neutrophil Function. Front. Immunol. 2018, 9, 399. Available online: https://www.frontiersin.org/article/10.3389/fimmu.2018.00399 (accessed on 16 March 2022). [CrossRef] [PubMed]
- Seifert, R.; Rosenthal, W.; Schultz, G. Guanine nucleotides stimulate NADPH oxidase in membranes of human neutrophils. FEBS Lett. 1986, 205, 161–165. [Google Scholar] [CrossRef]
- Tackenberg, H.; Möller, S.; Filippi, M.-D.; Laskay, T. The Small GTPase Cdc42 Is a Major Regulator of Neutrophil Effector Functions. Front. Immunol. 2020, 11, 1197. Available online: https://www.frontiersin.org/article/10.3389/fimmu.2020.01197 (accessed on 7 April 2022). [CrossRef]
- Werno, C.; Zhou, J.; Brüne, B. A23187, ionomycin and thapsigargin upregulate mRNA of HIF-1α via endoplasmic reticulum stress rather than a rise in intracellular calcium. J. Cell. Physiol. 2008, 215, 708–714. [Google Scholar] [CrossRef]
- Vorobjeva, N.V.; Chernyak, B.V. NETosis: Molecular Mechanisms, Role in Physiology and Pathology. Biochem. Mosc. 2020, 85, 1178–1190. [Google Scholar] [CrossRef]
- Kramer, P.A.; Ravi, S.; Chacko, B.; Johnson, M.S.; Darley-Usmar, V.M. A review of the mitochondrial and glycolytic metabolism in human platelets and leukocytes: Implications for their use as bioenergetic biomarkers. Redox Biol. 2014, 2, 206–210. [Google Scholar] [CrossRef]
- Amini, P.; Stojkov, D.; Felser, A.; Jackson, C.B.; Courage, C.; Schaller, A.; Gelman, L.; Soriano, M.E.; Nuoffer, J.-M.; Scorrano, L.; et al. Neutrophil extracellular trap formation requires OPA1-dependent glycolytic ATP production. Nat. Commun. 2018, 9, 2958. [Google Scholar] [CrossRef]
- Riffelmacher, T.; Clarke, A.; Richter, F.C.; Stranks, A.; Pandey, S.; Danielli, S.; Hublitz, P.; Yu, Z.; Johnson, E.; Schwerd, T.; et al. Autophagy-Dependent Generation of Free Fatty Acids Is Critical for Normal Neutrophil Differentiation. Immunity 2017, 47, 466–480.e5. [Google Scholar] [CrossRef]
- Kumar, S.; Dikshit, M. Metabolic Insight of Neutrophils in Health and Disease. Front. Immunol. 2019, 10, 2099. [Google Scholar] [CrossRef] [PubMed]
- Fossati, G.; Moulding, D.A.; Spiller, D.G.; Moots, R.J.; White, M.R.H.; Edwards, S.W. The Mitochondrial Network of Human Neutrophils: Role in Chemotaxis, Phagocytosis, Respiratory Burst Activation, and Commitment to Apoptosis. J. Immunol. 2003, 170, 1964–1972. [Google Scholar] [CrossRef]
- Koppaka, V.; Thompson, D.C.; Chen, Y.; Ellermann, M.; Nicolaou, K.C.; Juvonen, R.O.; Petersen, D.; Deitrich, R.A.; Hurley, T.D.; Vasiliou, V. Aldehyde Dehydrogenase Inhibitors: A Comprehensive Review of the Pharmacology, Mechanism of Action, Substrate Specificity, and Clinical Application. Pharmacol. Rev. 2012, 64, 520–539. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-S.; Kim, S.H.; Lee, S.; Kang, J.H.; Lee, S.-H.; Cheong, J.-H.; Kim, S.-Y. Gastric cancer depends on aldehyde dehydrogenase 3A1 for fatty acid oxidation. Sci. Rep. 2019, 9, 16313. [Google Scholar] [CrossRef] [PubMed]
- Endo, J.; Sano, M.; Katayama, T.; Hishiki, T.; Shinmura, K.; Morizane, S.; Matsuhashi, T.; Katsumata, Y.; Zhang, Y.; Ito, H.; et al. Metabolic Remodeling Induced by Mitochondrial Aldehyde Stress Stimulates Tolerance to Oxidative Stress in the Heart. Circ. Res. 2009, 105, 1118–1127. [Google Scholar] [CrossRef]
- Schmidt, C.A.; Fisher-Wellman, K.H.; Neufer, P.D. From OCR and ECAR to energy: Perspectives on the design and interpretation of bioenergetics studies. J. Biol. Chem. 2021, 297, 101140. [Google Scholar] [CrossRef]
- Fresneda Alarcon, M.; McLaren, Z.; Wright, H.L. Neutrophils in the Pathogenesis of Rheumatoid Arthritis and Systemic Lupus Erythematosus: Same Foe Different M.O. Front. Immunol. 2021, 12, 649693. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Forbes, R.A.; Verma, A. Hypoxia-inducible Factor 1 Activation by Aerobic Glycolysis Implicates the Warburg Effect in Carcinogenesis*. J. Biol. Chem. 2002, 277, 23111–23115. [Google Scholar] [CrossRef] [PubMed]
- Lum, J.J.; Bui, T.; Gruber, M.; Gordan, J.D.; DeBerardinis, R.J.; Covello, K.L.; Simon, M.C.; Thompson, C.B. The transcription factor HIF-1α plays a critical role in the growth factor-dependent regulation of both aerobic and anaerobic glycolysis. Genes Dev. 2007, 21, 1037–1049. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type 1 Diabetes | Healthy Control | |
|---|---|---|
| Number of donors | 14 | 12 |
| Age (years) * | 36.3 (19–61) | 34.5 (24–65) |
| Disease duration (years) * | 17.8 (4–38) | na |
| Gender F/M | 8/6 | 5/7 |
| Glycemia (mg/dL) * | 132.7 (76–175) | na |
| HbA1c (%) * | 7.8 (5.9–13.7) | na |
| Time In Range (TIR, %) * | 53.5 (19–88) | na |
| Insulin total daily dose (U/day) * | 33.8 (24.9–84) | na |
| GADA (U/mL) pos/neg ** | 5/9 | 0/12 |
| IA2-A (U/mL) pos/neg *** | 5/9 | 0/12 |
| IAA (% binding) pos/neg **** | 11/3 | 0/12 |
| ZnT8A (% binding) pos/neg ***** | 1/13 | 0/12 |
| Cellular Localization | Protein Name | Gene Name | Accession Number |
|---|---|---|---|
| Granules | Azurocidin | AZU1 | P20160 |
| Cathepsin G | CTSG | P08311 | |
| Lactotransferrin | LTF | P02788 | |
| Myeloperoxidase | MPO | P05164 | |
| Neutrophil defensin 1 | DEFA1 | P59665 | |
| Neutrophil Elastase | ELANE | P08246 | |
| Nucleus | Neutrophil gelatinase-associated lipocalin | NGAL | B2ZDQ1 |
| Histone H2B | HIST1H2BK | O60814 | |
| Myeloid cell nuclear differentiation antigen | MNDA | P41218 | |
| Protein-arginine deiminase type-4 | PADI4 | Q9UM07 | |
| Cytoplasm | Protein S100-A6 | S100A6 | P06703 |
| Protein S100-A8 | S100A8 | P05109 | |
| Protein S100-A9 | S100A9 | P06702 | |
| Protein S100-A11 | S100A11 | P31949 | |
| Protein S100-A12 | S100A12 | P80511 | |
| Protein S100-P | S100P | P25815 | |
| Cytoskeletal | Actin (cytoplasmic) | ACTB | P60709 |
| Myosin-9 | MYH9 | P35579 | |
| Peroxisomal | Catalase | CAT | P04040 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bissenova, S.; Ellis, D.; Callebaut, A.; Eelen, G.; Derua, R.; Buitinga, M.; Mathieu, C.; Gysemans, C.; Overbergh, L. NET Proteome in Established Type 1 Diabetes Is Enriched in Metabolic Proteins. Cells 2023, 12, 1319. https://doi.org/10.3390/cells12091319
Bissenova S, Ellis D, Callebaut A, Eelen G, Derua R, Buitinga M, Mathieu C, Gysemans C, Overbergh L. NET Proteome in Established Type 1 Diabetes Is Enriched in Metabolic Proteins. Cells. 2023; 12(9):1319. https://doi.org/10.3390/cells12091319
Chicago/Turabian StyleBissenova, Samal, Darcy Ellis, Aïsha Callebaut, Guy Eelen, Rita Derua, Mijke Buitinga, Chantal Mathieu, Conny Gysemans, and Lut Overbergh. 2023. "NET Proteome in Established Type 1 Diabetes Is Enriched in Metabolic Proteins" Cells 12, no. 9: 1319. https://doi.org/10.3390/cells12091319
APA StyleBissenova, S., Ellis, D., Callebaut, A., Eelen, G., Derua, R., Buitinga, M., Mathieu, C., Gysemans, C., & Overbergh, L. (2023). NET Proteome in Established Type 1 Diabetes Is Enriched in Metabolic Proteins. Cells, 12(9), 1319. https://doi.org/10.3390/cells12091319