

Validation and Functional Analysis of Reference and Tissue-Specific Genes in Adipose Tissue of Freshwater Drum, Aplodinotus grunniens, under Starvation and Hypothermia Stress

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Animals and Experimental Design
2.3. Sample Collection
2.4. Extraction of Total RNA and Synthesis of cDNA
2.5. Screening of Reference and Tissue-Specific Genes
2.6. RT-PCR Analysis
2.7. Statistical Analysis
3. Results
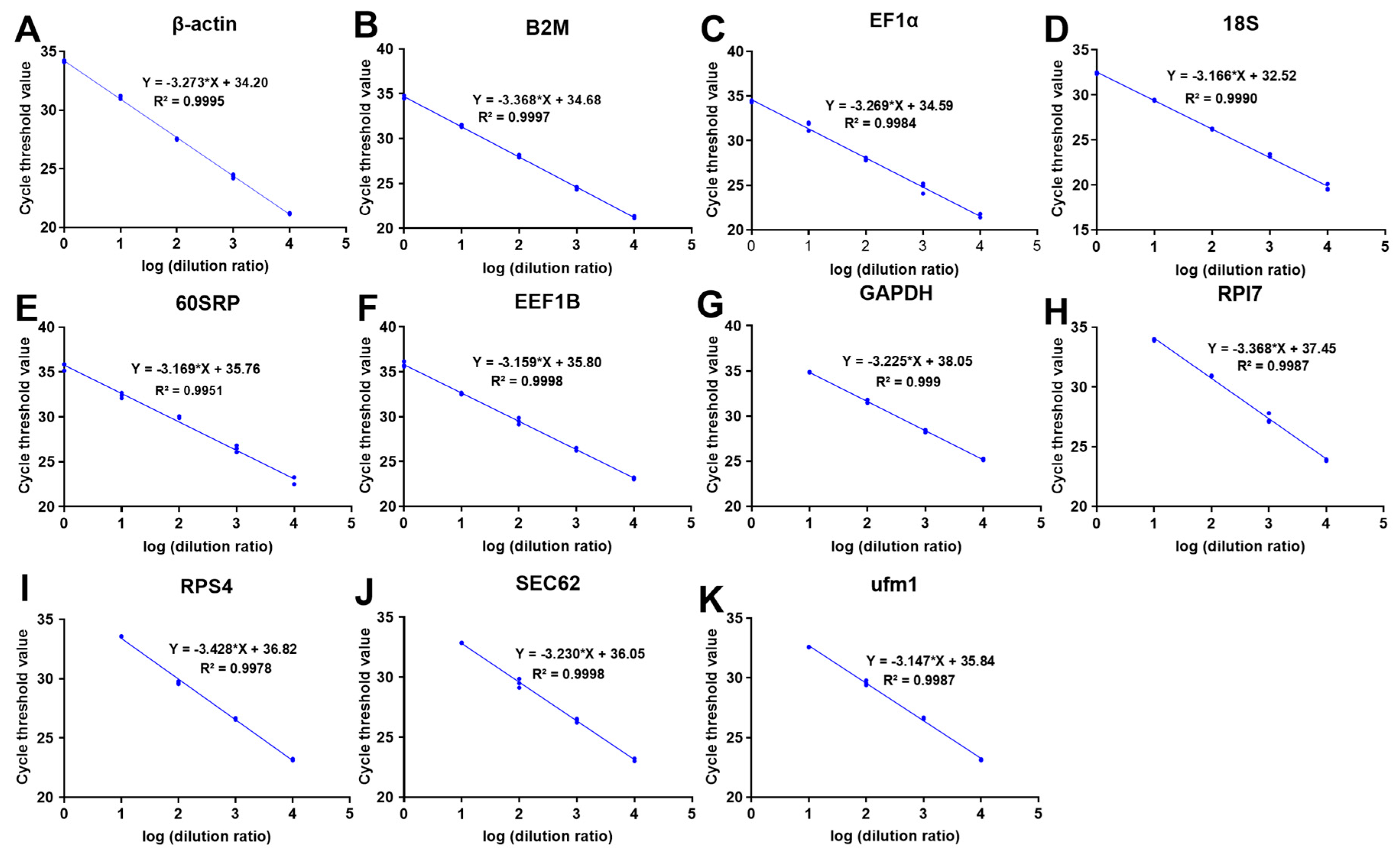
3.1. RNA Quality and Primer Amplification Assessment
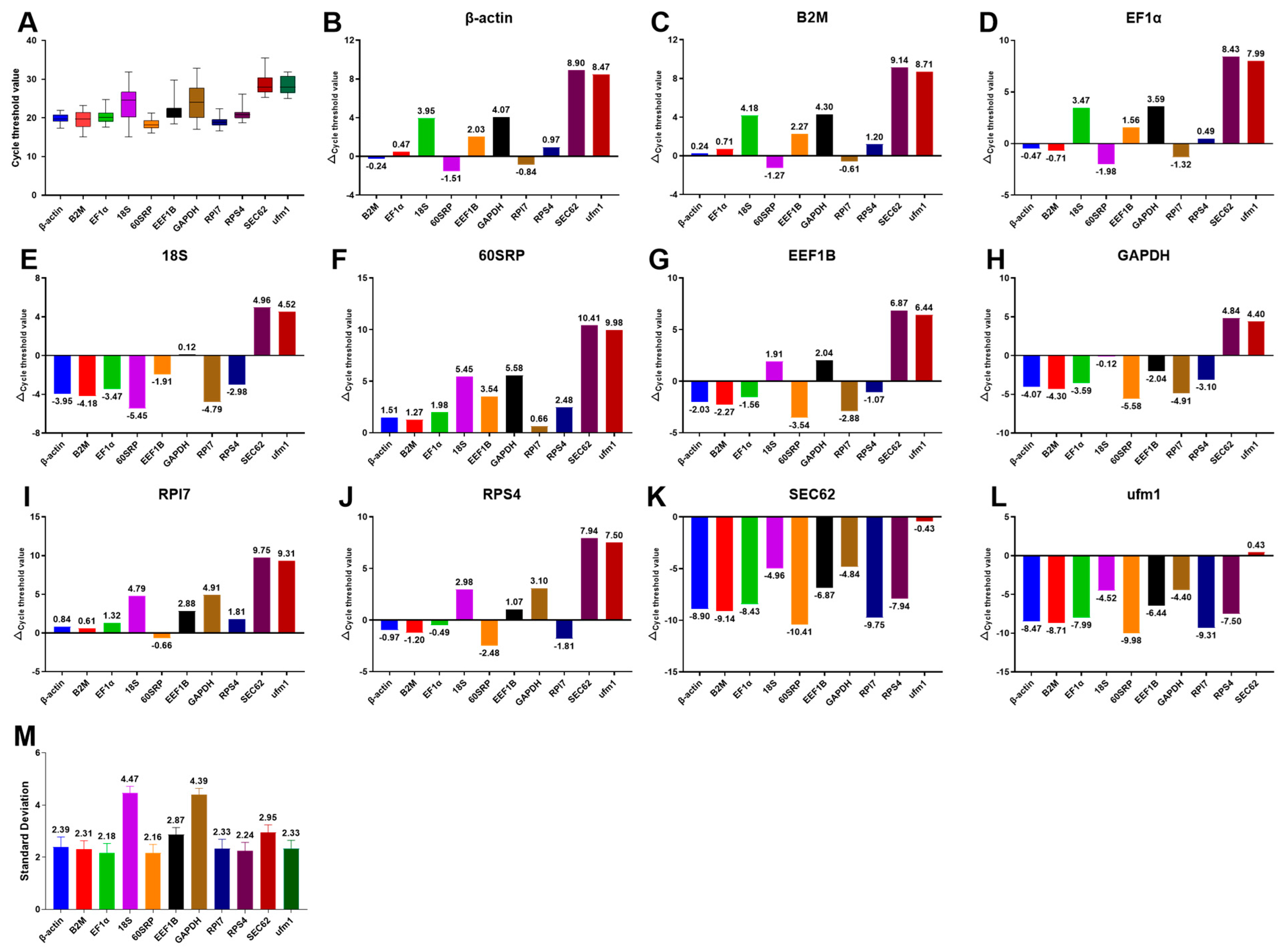
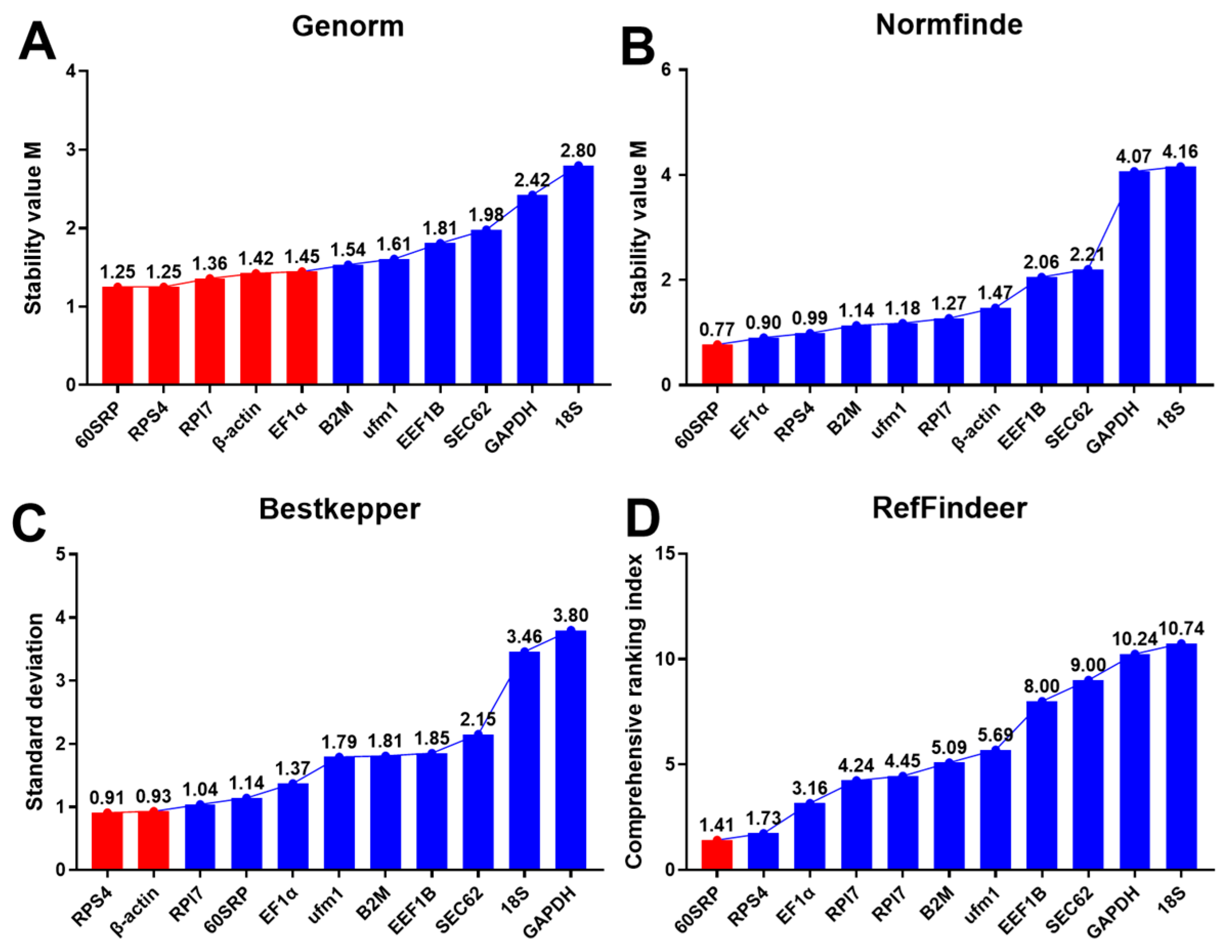
3.2. Expression Ranges and Stability of Candidate Reference Genes
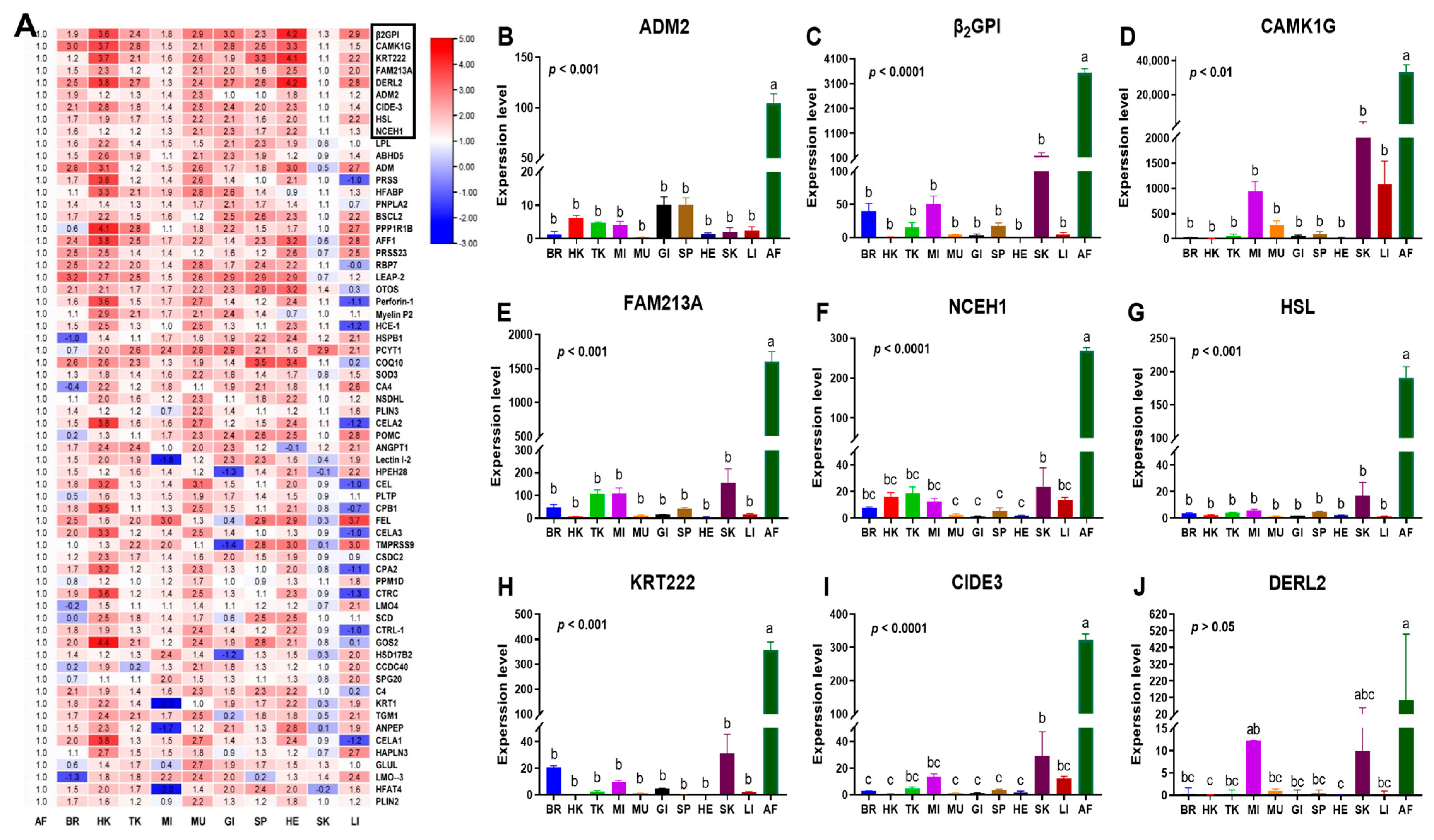
3.3. Identification and Expression of Adipose Tissue-Specific Genes
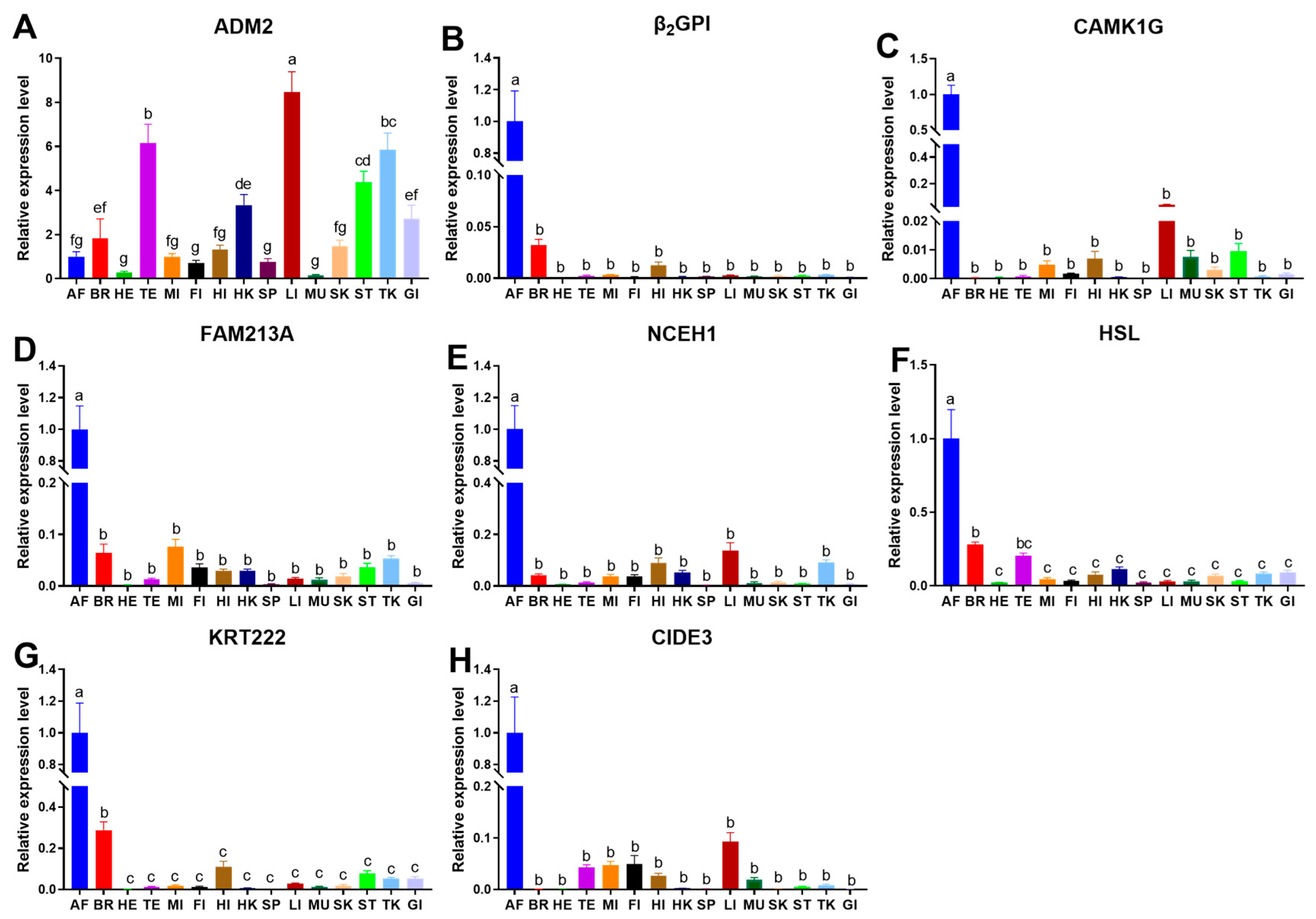
3.4. Expression of Adipose Tissue-Specific Genes in Different Tissues
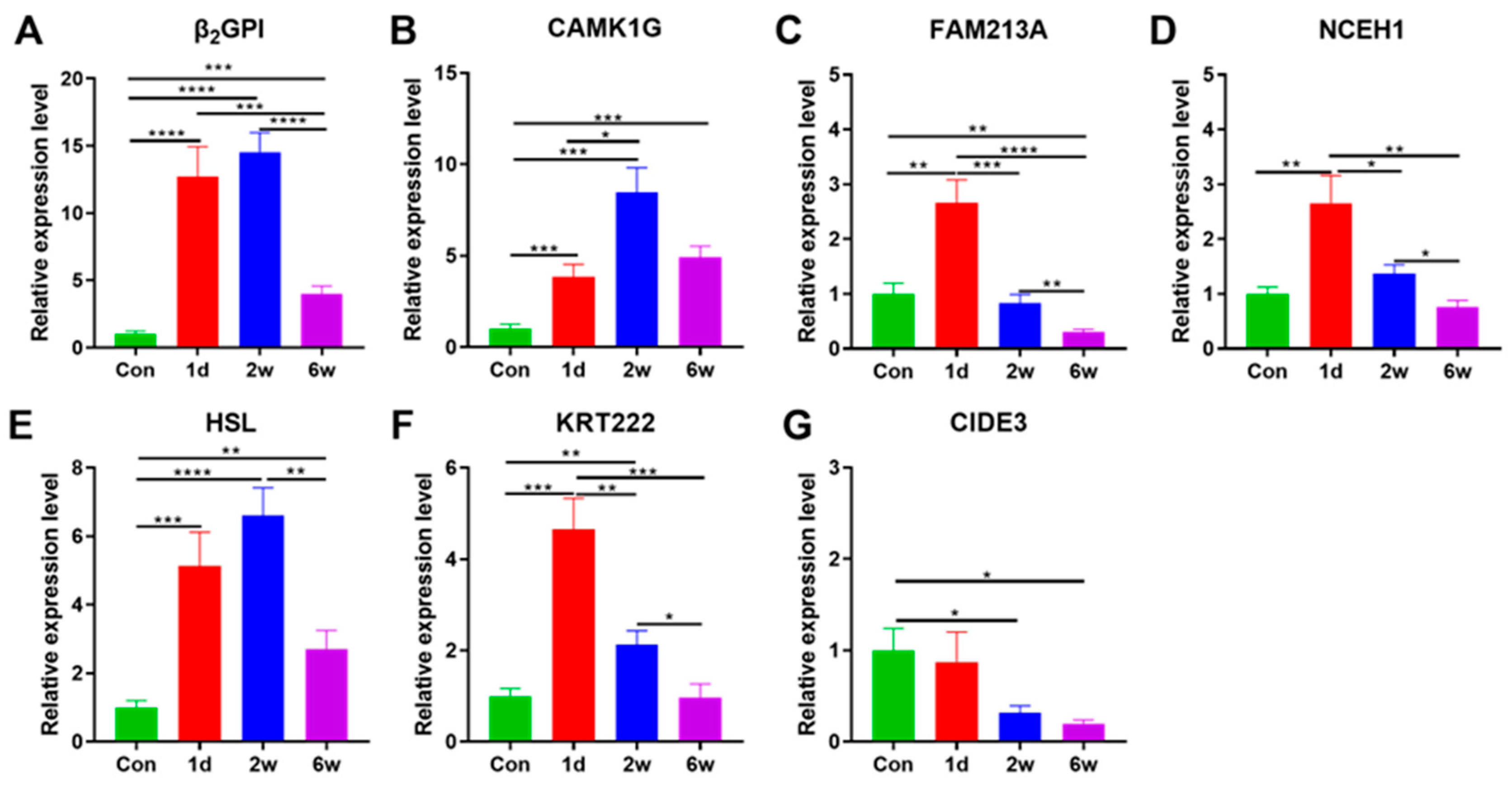
3.5. Expression Characteristics of Adipose Tissue-Specific Genes under Starvation
3.6. Expression Characteristics of Adipose Tissue-Specific Genes under Hypothermia
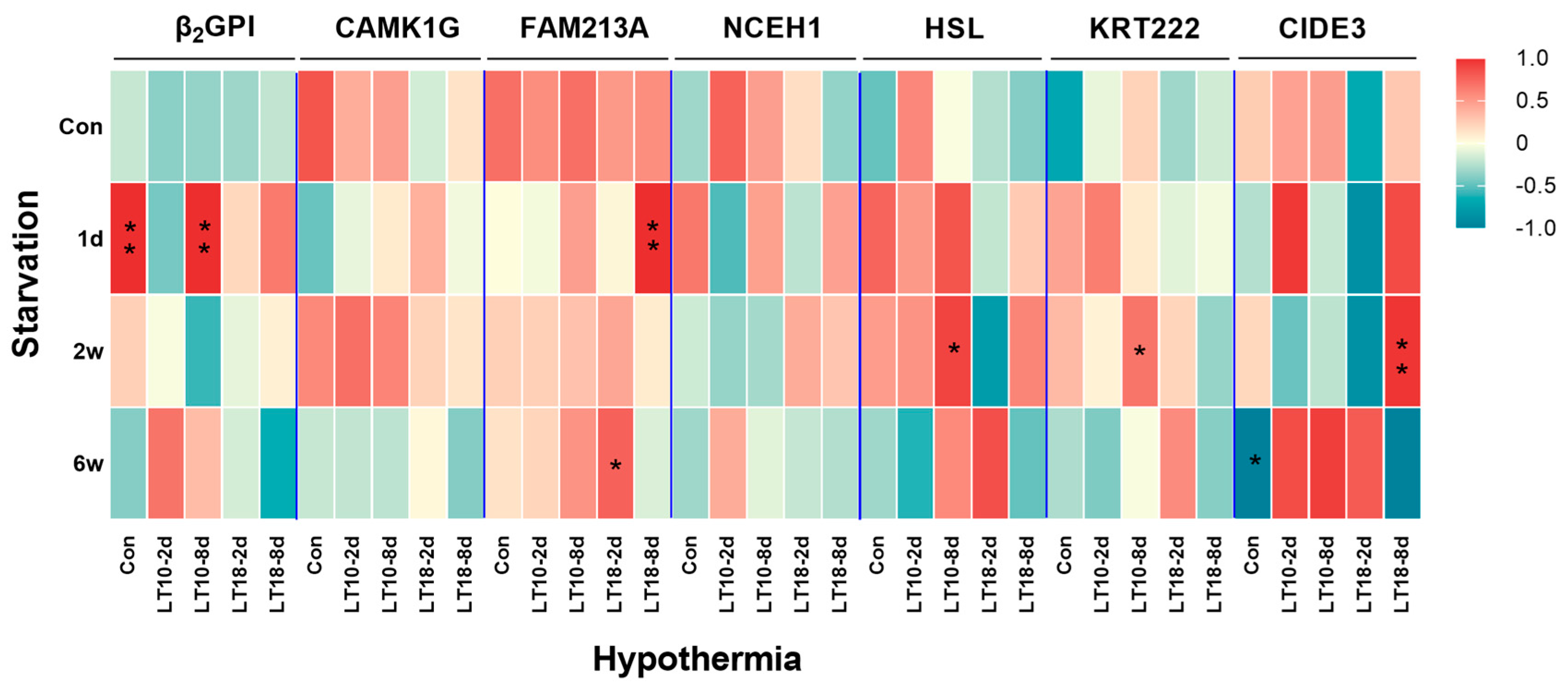
3.7. Comprehensive Analysis of Tissue-Specific Genes under Hypothermia and Starvation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Weil, C.; Lefèvre, F.; Bugeon, J. Characteristics and Metabolism of Different Adipose Tissues in Fish. Rev. Fish Biol. Fish. 2012, 23, 157–173. [Google Scholar] [CrossRef]
- Liu, D.; Mai, K.; Ai, Q. Tumor Necrosis Factor Alpha Is a Potent Regulator in Fish Adipose Tissue. Aquaculture 2015, 436, 65–71. [Google Scholar] [CrossRef]
- Fenzl, A.; Kiefer, F.W. Brown adipose tissue and thermogenesis. Horm. Mol. Biol. Clin. Investig. 2014, 19, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Chatzifotis, S.; Panagiotidou, M.; Papaioannou, N.; Pavlidis, M.; Nengas, I.; Mylonas, C.C. Effect of Dietary Lipid Levels on Growth, Feed Utilization, Body Composition and Serum Metabolites of Meagre (Argyrosomus regius) Juveniles. Aquaculture 2010, 307, 65–70. [Google Scholar] [CrossRef]
- Du, Z.Y.; Clouet, P.; Zheng, W.H.; Degrace, P.; Tian, L.X.; Liu, Y.J. Biochemical Hepatic Alterations and Body Lipid Composition in the Herbivorous Grass Carp (Ctenopharyngodon idella) Fed High-Fat Diets. Br. J. Nutr. 2006, 95, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.Y. Effects of HSL gene on fat deposition in common carp and its regulatory mechanism. Shanghai Ocean Univ. 2020, 2, 70. (In Chinese) [Google Scholar] [CrossRef]
- Zeng, B.H.; Jin, L.J.; Wang, Q.; Su, H.D. Pathology and prevention of liver injury in aquatic animals. Chongqing Aquac. 2014, 3, 38–41. (In Chinese) [Google Scholar]
- Walks, D.; Lavau, M.; Presta, E.; Yang, M.U.; Björntorp, P. Refeeding after Fasting in the Rat: Effects of Dietary-Induced Obesity on Energy Balance Regulation. Am. J. Clin. Nutr. 1983, 37, 387–395. [Google Scholar] [CrossRef]
- Wilson, R.; Deasy, W.; Stathis, C.; Hayes, A.; Cooke, M. Intermittent Fasting with or without Exercise Prevents Weight Gain and Improves Lipids in Diet-Induced Obese Mice. Nutrients 2018, 10, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar, N. Physiological and Hormonal Changes during Prolonged Starvation in Fish. Can. J. Fish. Aquat. Sci. 2014, 71, 1447–1458. [Google Scholar] [CrossRef]
- van Dijk, P.L.M.; Hardewig, I.; HOlker, F. Energy Reserves during Food Deprivation and Compensatory Growth in Juvenile Roach: The Importance of Season and Temperature. J. Fish Biol. 2005, 66, 167–181. [Google Scholar] [CrossRef]
- Kooka, K.; Yamamura, O. Winter Energy Allocation and Deficit of Juvenile Walleye Pollock Theragra Chalcogramma in the Doto Area, Northern Japan. Environ. Biol. Fishes 2011, 94, 389–402. [Google Scholar] [CrossRef]
- Lu, D.L. Mechanisms of Fasting Enhance Cold Tolerance in Zebrafish (Danio rerio). Ph.D. Thesis, East China Normal University, Shanghai, China, 2019; p. 165. (In Chinese). [Google Scholar]
- El-Sayed, A.-F.M.; Kawanna, M. Optimum Water Temperature Boosts the Growth Performance of Nile Tilapia (Oreochromis niloticus) Fry Reared in a Recycling System. Aquac. Res. 2008, 39, 670–672. [Google Scholar] [CrossRef]
- Snyder, R.J.; Hennessey, T.M. Cold Tolerance and Homeoviscous Adaptation in Freshwater Alewives (Alosa pseudoharengus). Fish Physiol. Biochem. 2003, 29, 117–126. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.; Jiang, S.; Xiong, Y.; Sun, C.; Zhou, Q.; Jiang, Z.; Liu, B.; Zhang, H. Anthraquinone Extract from Rheum Officinale Bail Improves Growth Performance and Toll–Relish Signaling-Regulated Immunity and Hyperthermia Tolerance in Freshwater Prawn Macrobrachium nipponense. 3 Biotech 2020, 10, 526. [Google Scholar] [CrossRef]
- Hsieh, S.L.; Kuo, C.-M. Stearoyl–CoA Desaturase Expression and Fatty Acid Composition in Milkfish (Chanos chanos) and Grass Carp (Ctenopharyngodon idella) during Cold Acclimation. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 141, 95–101. [Google Scholar] [CrossRef]
- Hazel, J.R. Influence of Thermal Acclimation on Membrane Lipid Composition of Rainbow Trout Liver. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1979, 236, R91–R101. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Mechanisms of Action of (n-3) Fatty Acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [Green Version]
- Spector, A.A.; Yorek, M.A. Membrane Lipid Composition and Cellular Function. J. Lipid Res. 1985, 26, 1015–1035. [Google Scholar] [CrossRef]
- Cao, Y.C.; Wang, Z.X. Study on the mechanism of low temperature adaption in fish. Ⅱ.Effects of acclimation temperature on fitty acid composition and cholesterl content in mitochondrial membranes of muscle tissue of the grass carp and mud carp. J. South China Agric. Univ. 1991, S1, 45–49. (In Chinese) [Google Scholar]
- Zhang, X.D.; Ye, C.X.; Xian, J.A.; Wang, A.L. Research progress of fish cold stress. Feed. Ind. 2013, 34, 17–22. (In Chinese) [Google Scholar]
- Liu, W.; Sun, Z.Q.; Xie, H.W. The Progress of Gene Tissue Specificity Researches. Prog. Biochem. Biophys. 2016, 43, 5–13. (In Chinese) [Google Scholar]
- Zhang, Y.N. Screening on Pathways and Genes Related to Ovary Development of Oriental River Prawn (Macrobrachium nipponense) Based on Trancriptme and Metabolome. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2020. (In Chinese). [Google Scholar] [CrossRef]
- Gao, Y.F. Screening of Growth and Feeding Genes and Association Analysis between of wnt5b with Growth-Related Traits in Bighead carp. Master’s Thesis, Dalian Ocean University, Dalian, China, 2022. (In Chinese). [Google Scholar] [CrossRef]
- Bo, Y.Y. Exploited of Immune-Related Gene and Constructed of Microarray in the Common Carp. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2013. (In Chinese). [Google Scholar]
- Hernández-Gómez, R.E.; Contreras-Sánchez, W.M.; Hernández-Franyutti, A.; Perera-García, M.A.; Torres-Martínez, A. Testicular Structure and Development of the Male Germinal Epithelium in the Freshwater Drum Aplodinotus grunniens (Perciformes: Sciaenidae) from the Usumacinta River, Southern Mexico. Acta Zool. 2021, 103, 414–432. [Google Scholar] [CrossRef]
- Song, C.; Wen, H.; Liu, G.; Ma, X.; Lv, G.; Wu, N.; Chen, J.; Xue, M.; Li, H.; Xu, P. Gut Microbes Reveal Pseudomonas Medicates Ingestion Preference via Protein Utilization and Cellular Homeostasis Under Feed Domestication in Freshwater Drum, Aplodinotus grunniens. Front. Microbiol. 2022, 13, 861705. [Google Scholar] [CrossRef]
- Chen, J.; Li, H.; Xu, P.; Tang, Y.; Su, S.; Liu, G.; Wu, N.; Xue, M.; Yu, F.; Feng, W.; et al. Hypothermia-Mediated Apoptosis and Inflammation Contribute to Antioxidant and Immune Adaption in Freshwater Drum, Aplodinotus Grunniens. Antioxidants 2022, 11, 1657. [Google Scholar] [CrossRef]
- Chen, J.; Song, C.; Wen, H.; Liu, G.; Wu, N.; Li, H.; Xue, M.; Xu, P. miR-1/AMPK-Mediated Glucose and Lipid Metabolism under Chronic Hypothermia in the Liver of Freshwater Drum, Aplodinotus Grunniens. Metabolites 2022, 12, 697. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Wen, H.; Xu, P.; Chen, J.; Xue, M.; Li, J.; Wang, M.; Song, C.; Li, H. PPAR Signaling Maintains Metabolic Homeostasis under Hypothermia in Freshwater Drum (Aplodinotus grunniens). Metabolites 2023, 13, 102. [Google Scholar] [CrossRef] [PubMed]
- Peters, I.R.; Peeters, D.; Helps, C.R.; Day, M.J. Development and Application of Multiple Internal Reference (Housekeeper) Gene Assays for Accurate Normalisation of Canine Gene Expression Studies. Vet. Immunol. Immunopathol. 2007, 117, 55–66. [Google Scholar] [CrossRef]
- Mitter, K.; Kotoulas, G.; Magoulas, A.; Mulero, V.; Sepulcre, P.; Figueras, A.; Novoa, B.; Sarropoulou, E. Corrigendum to “Evaluation of Candidate Reference Genes for QPCR during Ontogenesis and of Immune-Relevant Tissues of European Seabass (Dicentrarchus labrax)” [Comp. Biochem. Physiol. 153B (2009) 340–347]. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 154, 372. [Google Scholar] [CrossRef]
- Bustin, S. Absolute Quantification of mRNA Using Real-Time Reverse Transcription Polymerase Chain Reaction Assays. J. Mol. Endocrinol. 2000, 25, 169–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazari, F.; Parham, A.; Maleki, A.F. GAPDH, β-Actin and Β2-Microglobulin, as Three Common Reference Genes, Are Not Reliable for Gene Expression Studies in Equine Adipose- and Marrow-Derived Mesenchymal Stem Cells. J. Anim. Sci. Technol. 2015, 57, 18. [Google Scholar] [CrossRef] [Green Version]
- Glare, E.M. Beta-Actin and GAPDH Housekeeping Gene Expression in Asthmatic Airways Is Variable and Not Suitable for Normalising mRNA Levels. Thorax 2002, 57, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Li, M.; Wang, K.; Qin, Q.; Chen, X. Characterization of Large Yellow Croaker (Pseudosciaena crocea) β-Actin Promoter Supports β-Actin Gene as an Internal Control for Gene Expression Modulation and Its Potential Application in Transgenic Studies in Fish. Fish Shellfish. Immunol. 2011, 30, 1072–1079. [Google Scholar] [CrossRef]
- Ruan, W.; Lai, M. Actin, a Reliable Marker of Internal Control? Clin. Chim. Acta 2007, 385, 1–5. [Google Scholar] [CrossRef]
- Tossounian, M.A.; Zhang, B.; Gout, I. The writers, readers, and erasers in redox regulation of GAPDH. Antioxidants 2020, 9, 1288. [Google Scholar] [CrossRef]
- Zhang, W.-X.; Fan, J.; Ma, J.; Rao, Y.-S.; Zhang, L.; Yan, Y.-E. Selection of Suitable Reference Genes for Quantitative Real-Time PCR Normalization in Three Types of Rat Adipose Tissue. Int. J. Mol. Sci. 2016, 17, 968. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Hu, J. Development and Validation of Endogenous Reference Genes for Expression Profiling of Medaka (Oryzias latipes) Exposed to Endocrine Disrupting Chemicals by Quantitative Real-Time RT-PCR. Toxicol. Sci. 2006, 95, 356–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Cheng, J.; Fan, K.; Xia, Y.; Zhang, Z.; Liu, Y.; Liu, P. Evaluation of Potential Reference Genes by Quantitative RT-qPCR Analysis of Takifugu rubripes under Normal Conditions and after Cryptocaryon Irritans Infection. Aquac. Res. 2021, 52, 5814–5828. [Google Scholar] [CrossRef]
- Rassier, G.T.; Silveira, T.L.R.; Remião, M.H.; Daneluz, L.O.; Martins, A.W.S.; Dellagostin, E.N.; Ortiz, H.G.; Domingues, W.B.; Komninou, E.R.; Kütter, M.T.; et al. Evaluation of qPCR Reference Genes in GH-Overexpressing Transgenic Zebrafish (Danio rerio). Sci. Rep. 2020, 10, 12692. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, C.; Zeng, Q.; Agrawal, I.; Zhu, X.; Gong, Z. Genome-Wide Identification of Suitable Zebrafish Danio Rerio Reference Genes for Normalization of Gene Expression Data by RT-qPCR. J. Fish Biol. 2016, 88, 2095–2110. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Hernandez, N.; Véliz, D.; Vega-Retter, C. Selection of Suitable Reference Genes for Gene Expression Analysis in Gills and Liver of Fish under Field Pollution Conditions. Sci. Rep. 2019, 9, 3459. [Google Scholar] [CrossRef] [Green Version]
- Bettacchioli, E.; Nafai, S.; Renaudineau, Y. News and Meta-Analysis Regarding Anti-Beta 2 Glycoprotein I Antibodies and Their Determination. Clin. Immunol. 2019, 205, 106–115. [Google Scholar] [CrossRef]
- Puri, V.; Konda, S.; Ranjit, S.; Aouadi, M.; Chawla, A.; Chouinard, M.; Chakladar, A.; Czech, M.P. Fat-Specific Protein 27, a Novel Lipid Droplet Protein That Enhances Triglyceride Storage. J. Biol. Chem. 2007, 282, 34213–34218. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.-L.; Lou, J.-C.; Lyu, W.; Zhang, B. Update on the Synergistic Effect of HSL and Insulin in the Treatment of Metabolic Disorders. Ther. Adv. Endocrinol. Metab. 2019, 10, 204201881987730. [Google Scholar] [CrossRef]
- Lampidonis, A.D.; Rogdakis, E.; Voutsinas, G.E.; Stravopodis, D.J. The Resurgence of Hormone-Sensitive Lipase (HSL) in Mammalian Lipolysis. Gene 2011, 477, 1–11. [Google Scholar] [CrossRef]
- Sekiya, M.; Yamamuro, D.; Ohshiro, T.; Honda, A.; Takahashi, M.; Kumagai, M.; Sakai, K.; Nagashima, S.; Tomoda, H.; Igarashi, M.; et al. Absence of Nceh1 Augments 25-Hydroxycholesterol-Induced ER Stress and Apoptosis in Macrophages. J. Lipid Res. 2014, 55, 2082–2092. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, M.; Osuga, J.; Isshiki, M.; Sekiya, M.; Okazaki, H.; Takase, S.; Takanashi, M.; Ohta, K.; Kumagai, M.; Nishi, M.; et al. Targeting of Neutral Cholesterol Ester Hydrolase to the Endoplasmic Reticulum via Its N-Terminal Sequence. J. Lipid Res. 2010, 51, 274–285. [Google Scholar] [CrossRef] [Green Version]
- Takemoto-Kimura, S.; Ishihara-Ageta, N.; Nonaka, M.; Okuno, H.; Bito, H. Regulation of Dendritogenesis via a Lipid Raft-Associated Ca2+/Calmodulin-Dependent Protein Kinase CLICK-III/CaMKIγ. Neurosci. Res. 2007, 58, S20. [Google Scholar] [CrossRef]
- Li, Z.; Tian, Y.; Wang, L.; Li, Z.; Chen, S.; Li, L.; Liu, Y.; Li, W.; Pang, Z.; Ma, W.; et al. Comparative Transcriptomics Analyses and Revealing Candidate Networks and Genes Involved in Lordosis of the Yunlong Grouper (Epinephelus Moara ♀ × Epinephelus Lanceolatus ♂). Aquaculture 2022, 550, 737866. [Google Scholar] [CrossRef]
- Herrmann, H.; Hesse, M.; Reichenzeller, M.; Aebi, U.; Magin, T.M. Functional complexity of intermediate filament cytoskeletons: From structure to assembly to gene ablation. Int. Rev. Cytol. 2003, 223, 83–175. [Google Scholar] [CrossRef] [PubMed]
- Kirfel, J.; Magin, T.M.; Reichelt, J. Keratins: A Structural Scaffold with Emerging Functions. Cell. Mol. Life Sci. 2003, 60, 56–71. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.K.; Ha, M.; Han, M.E.; Heo, H.J.; Myung, K.; Lee, Y.; Oh, S.O.; Kim, Y.H. FAM213A is linked to prognostic significance in acute myeloid leukemia through regulation of oxidative stress and myelopoiesis. Hematol Oncol. 2020, 38, 381–389. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Wei, Y.-Y.; Yang, C.-C.; Liu, C.-J.; Yeh, L.-Y.; Chou, C.-H.; Chang, K.-W.; Lin, S.-C. miR-125b Suppresses Oral Oncogenicity by Targeting the Anti-Oxidative Gene PRXL2A. Redox Biol. 2019, 22, 101140. [Google Scholar] [CrossRef]
- Matafome, P.; Seiça, R. Function and Dysfunction of Adipose Tissue. Adv. Neurobiol. 2017, 19, 3–31. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.J.; Zhang, J.; Huang, J.S.; Shi, G.; Pan, C.H.; Xie, R.T.; Chen, G.; Zhang, J.D.; Wang, Z.L.; Tang, B.G. Effects of low temperature stress on the expression of genes related to lipid metabolism of juvenile cobia, Rachycentron canadum. Haiyang Xuebao 2021, 43, 116–122. (In Chinese) [Google Scholar]
- Xu, C. Study on the Triglycerol Catabolism, Dietary Lipid and α-Lipoic acid on Chinese Mitten Crab Eriocheir Sinensis. Ph.D. Thesis, East China Normal University, Shanghai, China, 2018. (In Chinese). [Google Scholar]
- Dietrich, M.A.; Hliwa, P.; Adamek, M.; Steinhagen, D.; Karol, H.; Ciereszko, A. Acclimation to Cold and Warm Temperatures Is Associated with Differential Expression of Male Carp Blood Proteins Involved in Acute Phase and Stress Responses, and Lipid Metabolism. Fish Shellfish. Immunol. 2018, 76, 305–315. [Google Scholar] [CrossRef]
- Mateus, A.P.; Costa, R.; Gisbert, E.; Pinto, P.I.S.; Andree, K.B.; Estévez, A.; Power, D.M. Thermal Imprinting Modifies Bone Homeostasis in Cold Challenged Sea Bream (Sparus aurata L.). J. Exp. Biol. 2017, 220, 3442–3454. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.J.; Shao, T.; Yang, J.P.; Gong, Q.; Li, L.J. The effect of starvation on lipid metabolism of darkbarbel catfish, Pelteobagrus vachelli. Acta Hydrobiol. Sin. 2015, 39, 58–65. (In Chinese) [Google Scholar]
- Larsson, A.; Lewander, K. Metabolic Effects of Starvation in the Eel, Anguilla anguilla L. Comp. Biochem. Physiol. Part A Physiol. 1973, 44, 367–374. [Google Scholar] [CrossRef]
- Ince, B.W.; Thorpe, A. The Effects of Starvation and Force-Feeding on the Metabolism of the Northern Pike, Esox lucius L. J. Fish Biol. 1976, 8, 79–88. [Google Scholar] [CrossRef]
- Long, Y.; Ge, G.D.; Li, X.X.; Cui, Z.B. Regulatory mechanisms of low-temperature stress response in fish. Acta Hydrobiol. Sin. 2021, 45, 1405–1414. (In Chinese) [Google Scholar]
- Zhao, C.Y.; Zhou, X.; Bing, X.W.; Wang, G.Q. Effects of starvation on digestive enzyme activities and some immune indexes in broodstock red swamp crawfish Procambarus clarkii. J. Dalian Fish Univ. 2010, 25, 85–87. (In Chinese) [Google Scholar] [CrossRef]
- Li, D.P.; Liu, S.Y.; Xie, C.X.; Zhang, X.Z. Effects of water temperature on serum content of reactive oxygen species and antiox idant defense system in Chinese sturgeon, Acipenser sinensis. Acta Hydrobiol. Sin. 2008, 32, 327–332. (In Chinese) [Google Scholar] [CrossRef]
- Chen, H.G.; Ma, S.W.; Lin, Q.; Gan, J.L.; Cai, W.G.; Jia, X.P. Effects of tribu tyltin chloride (TBTCl) on SOD activities, MDA contents and GPx activities in gill and liver of the black porgy (Sparus macro cephalus). South China Fish. Sci. 2009, 5, 23–27. (In Chinese) [Google Scholar]
- Du, Z.Y.; Liu, Y.J.; Tian, L.X.; Cao, J.M.; Liang, G.Y.; He, J.G. Effects of starvation on visceral weight and main biochemical compsition of the muscle, liver and serum in the Janpanese sea bass (Lateolabrax japonicus). Acta Zool. Sin. 2003, 49, 458–465. (In Chinese) [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank | Gene | Primer Sequence (5′—3′) | Amplification Efficiency | R2 |
|---|---|---|---|---|
| XP_008328442.1 | β-actin | F: AGGCTGTGCTGTCCCTGTAT R: GCTGTGGTGGTGAAGGAGTAG | 102.08 | 0.9995 |
| AEI83278.1 | B2M | F: CCTGGAAAGTTCGGCAGTAG R: TCCACGTTCTTGGTCAGATG | 98.11 | 0.9997 |
| KKF31758.1 | EF1α | F: TGACAACTTCAACGCTCAGG R: ATGGGCTTCTGTGGAATGAG | 102.26 | 0.9984 |
| AAL31950.1 | 18S | F: TCAGCGTGTGTCTACCCTTC R: CCTCACTAAACCATCCAATCG | 106.95 | 0.9990 |
| XP_018517365.1 | 60SRP | F: CAAAGGACATCAAGGCCATC R: GAGCCACTACAGCACCACCT | 106.80 | 0.9951 |
| CAG01324.1 | EEF1B | F: GATGAGGGTGGGCTTCTTG R: ATGTTGACCTGTTCGGCTCT | 107.28 | 0.9998 |
| XP_010741722.1 | GAPDH | F: ATGACCCTTTCATCGACCTG R: GCTTCACCCCATTTGATGATT | 104.21 | 0.9990 |
| XP_010732148.1 | RPl7 | F: GATGCTGGCTGAGAAGAAGG R: GCCGTTGATACCTCTGATCC | 98.11 | 0.9987 |
| XP_010747631.1 | RPS4 | F: GACAAGCTGACCGGAGTGTT R: CCAGCAGGGTAGGTGATGTC | 95.76 | 0.978 |
| XP_010738284.2 | SEC62 | F: GCCATCACTTCTGGTTCCTC R: CCATCCTTCTTTTCGCTGTC | 103.98 | 0.9998 |
| XP_005808493.1 | ufm1 | F: GCCGTTCACAGCAGTTTG R: GTCTCCTTGTCCTCCCACTCT | 107.86 | 0.9987 |
| XP_010742647.2 | β2GP1 | F: GGCAGTATCCTCACCCCATC R: CCTTCTGAGGTCCATCCAGC | 99.17 | 0.9988 |
| KKF21127.1 | CAMK1G | F: TACATGCTCGGCTCCACTCT R: TCTCCTTCACGCTCAACTCG | 108.54 | 0.9986 |
| KKF23363.1 | KRT222 | F: GAGAGTGCAGAAGGTCACGG R: GGGGAGGCTGTCCTGTTTAG | 94.77 | 0.9774 |
| XP_010735223.1 | ADM2 | F: GCATGAAAGCAGCCTTGTCG R: CATGTTCCCAAGACGCAACC | 107.28 | 0.9541 |
| XP_018535573.1 | CIDE3 | F: ACCCCACATCCAAACAGCAT R: TTTTTGGCAGCGTAACAGCG | 92.21 | 0.9628 |
| XP_019122735.1 | HSL | F: TTGCTGAGATGAGGGTGGA R: ACAGGCTGGTCTATGTTCC | 107.33 | 0.9657 |
| XP_010730495.2 | NCEH1 | F: TATTAACGGTGGCGTTCGCT R: AAAGAAGCCAGGTGCATCGT | 110.28 | 0.9964 |
| XP_010741055.2 | FAM213A | F: CCCGTGAAAGAAAGATGG R: GTCCAATGACGAACACCC | 107.57 | 0.9736 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, M.; Wen, H.; Xu, P.; Chen, J.; Wang, Q.; Tang, Y.; Ma, X.; Lv, G.; Li, H.; Song, C. Validation and Functional Analysis of Reference and Tissue-Specific Genes in Adipose Tissue of Freshwater Drum, Aplodinotus grunniens, under Starvation and Hypothermia Stress. Cells 2023, 12, 1328. https://doi.org/10.3390/cells12091328
Xue M, Wen H, Xu P, Chen J, Wang Q, Tang Y, Ma X, Lv G, Li H, Song C. Validation and Functional Analysis of Reference and Tissue-Specific Genes in Adipose Tissue of Freshwater Drum, Aplodinotus grunniens, under Starvation and Hypothermia Stress. Cells. 2023; 12(9):1328. https://doi.org/10.3390/cells12091328
Chicago/Turabian StyleXue, Miaomiao, Haibo Wen, Pao Xu, Jianxiang Chen, Qingyong Wang, Yongkai Tang, Xueyan Ma, Guohua Lv, Hongxia Li, and Changyou Song. 2023. "Validation and Functional Analysis of Reference and Tissue-Specific Genes in Adipose Tissue of Freshwater Drum, Aplodinotus grunniens, under Starvation and Hypothermia Stress" Cells 12, no. 9: 1328. https://doi.org/10.3390/cells12091328
APA StyleXue, M., Wen, H., Xu, P., Chen, J., Wang, Q., Tang, Y., Ma, X., Lv, G., Li, H., & Song, C. (2023). Validation and Functional Analysis of Reference and Tissue-Specific Genes in Adipose Tissue of Freshwater Drum, Aplodinotus grunniens, under Starvation and Hypothermia Stress. Cells, 12(9), 1328. https://doi.org/10.3390/cells12091328