

Activated Human Adipose Tissue Transplantation Promotes Sensorimotor Recovery after Acute Spinal Cord Contusion in Rats
 , ,
, ,  ,
,  ,
,  and
and 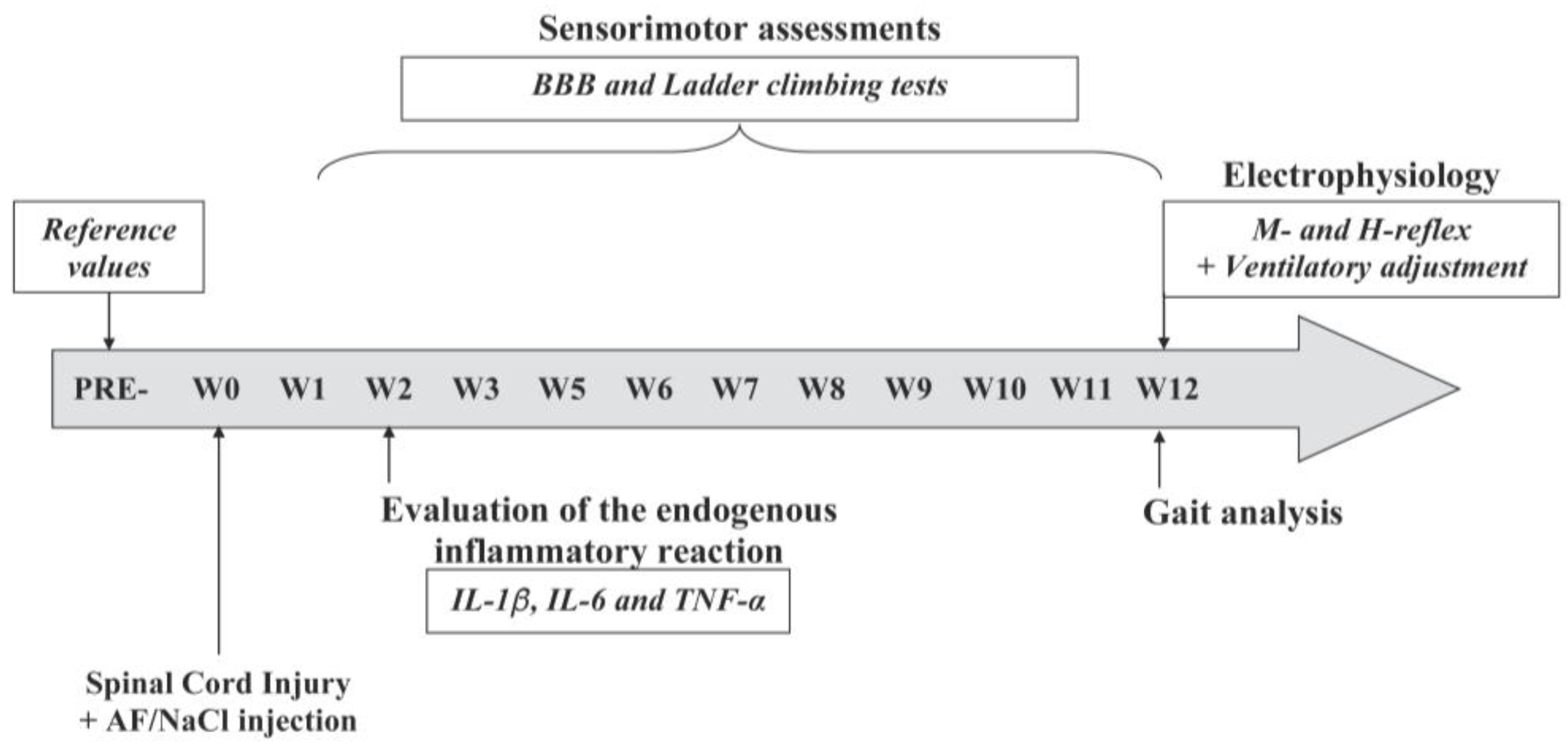{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ethical Considerations
2.3. Fat Collection, Purification, and Mechanical Activation
2.4. Experimental Groups and Design of the Study
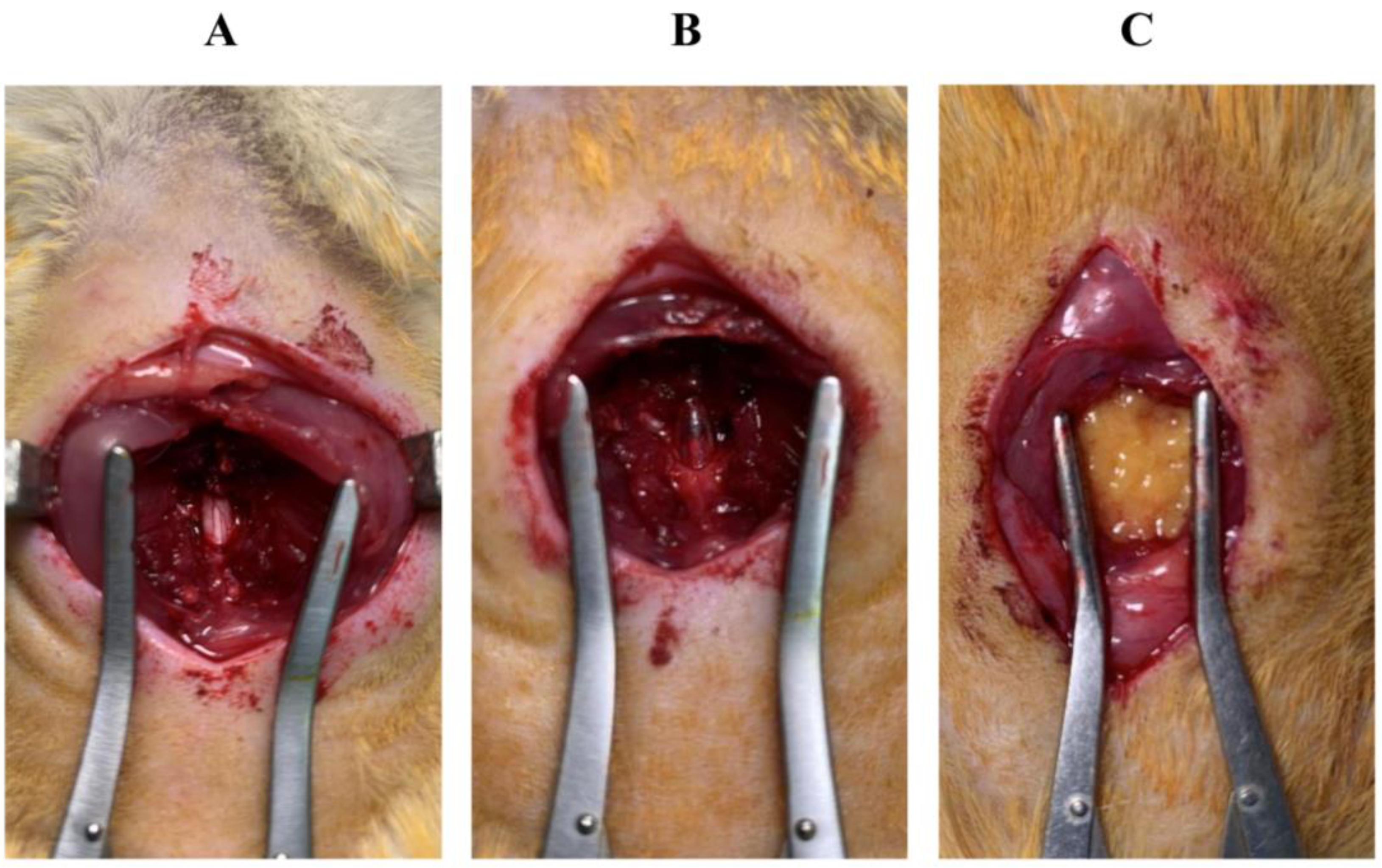
2.5. Surgical Procedure for the SCI
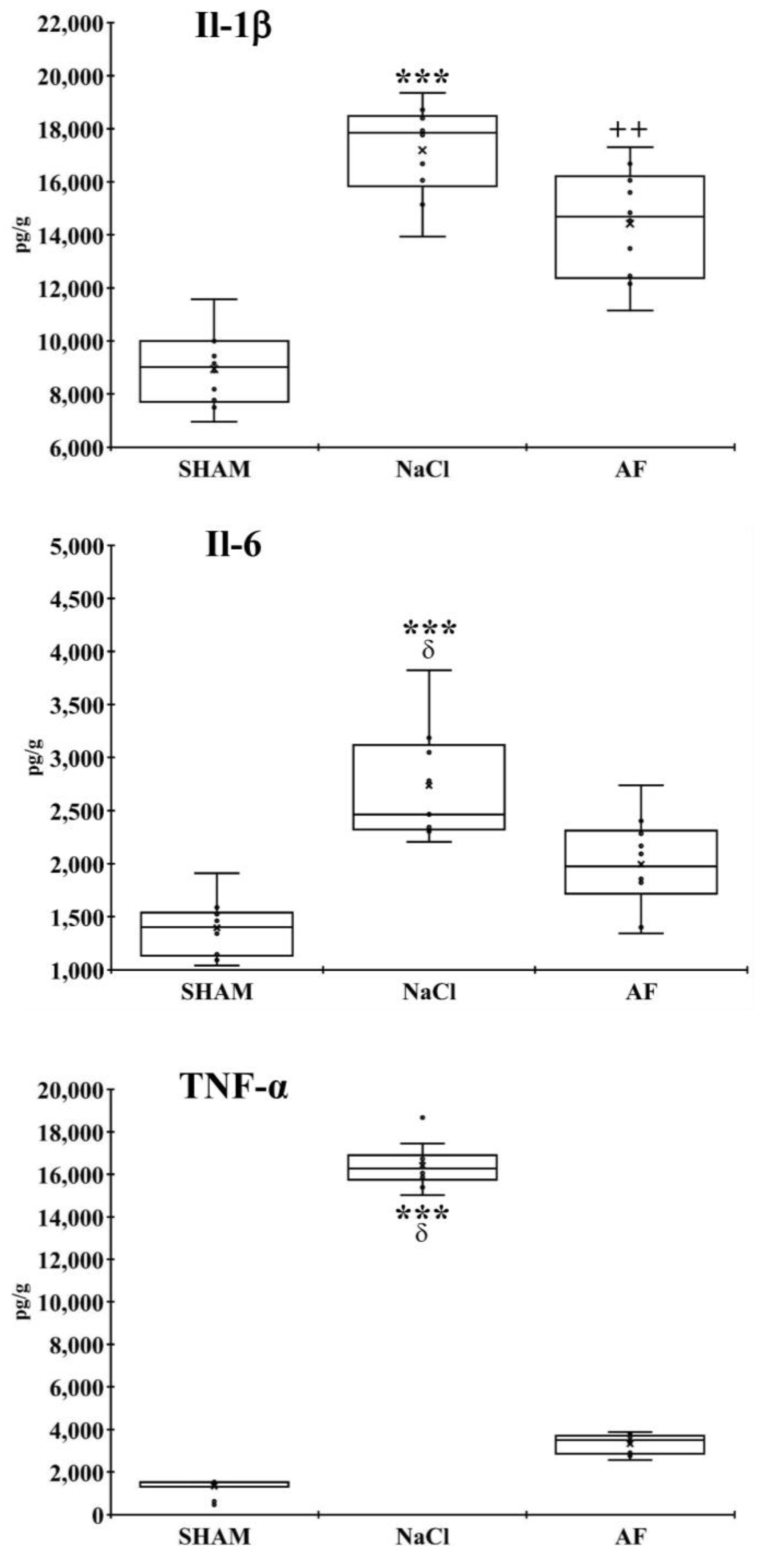
2.6. Endogenous Inflammation
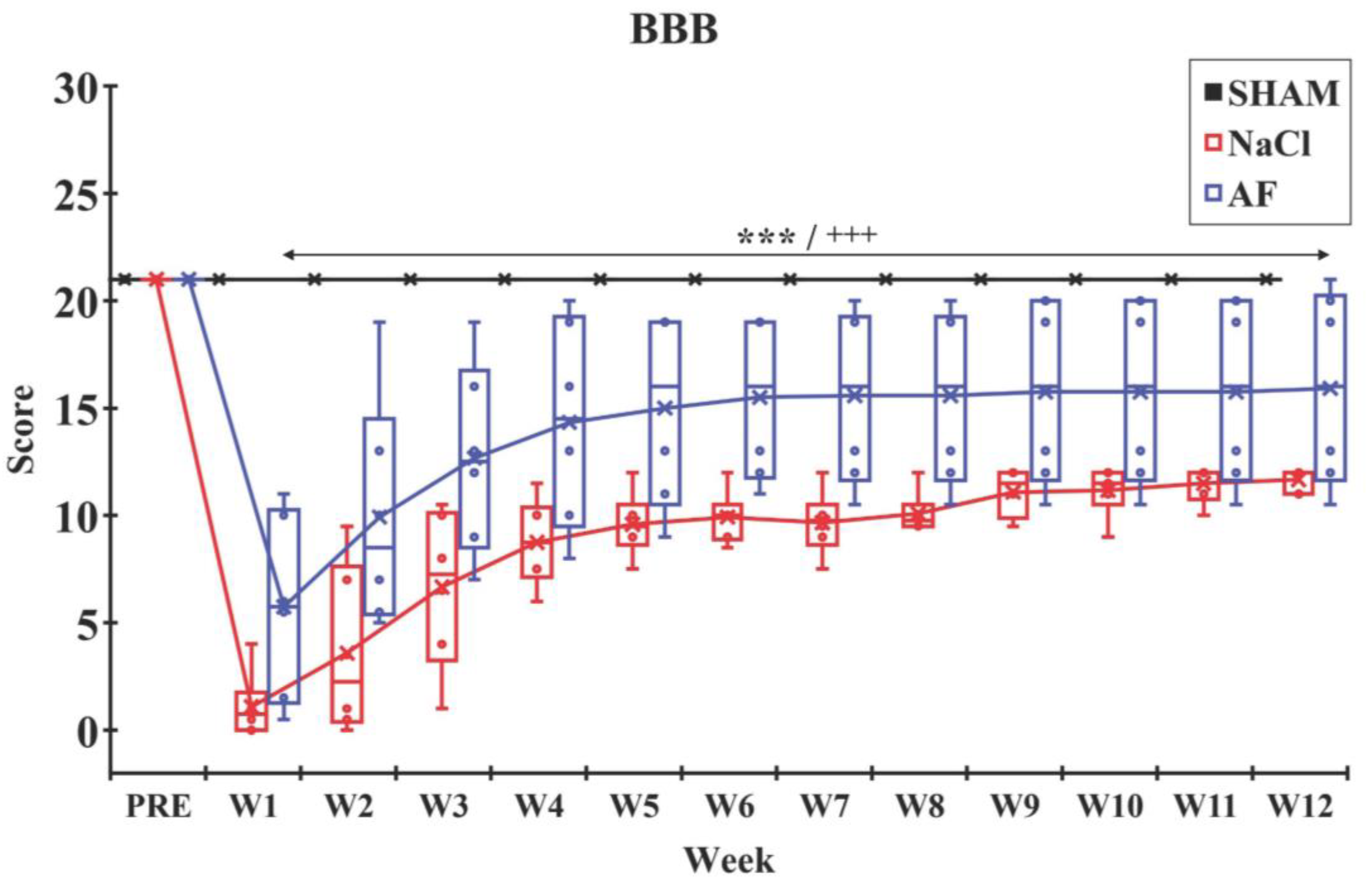
2.7. Evaluation of Hindlimb Recovery
- − Average speed: The speed of forward locomotion across the runway, measured in centimeters per second (cm/s).
- − Stride length: The distance between successive placements of the same paw, measured in centimeters (cm).
- − Stance phase: The duration of paw contact with the glass plate, measured in seconds (s).
- − Swing phase: The duration during which the paw was not in contact with the glass plate, measured in seconds (s).
- − Step cycle: The time between two successive placements of a single paw, calculated as the sum of the stance and swing phases, measured in seconds (s).
- − Phase dispersion: This parameter evaluated the synchrony of the initial contact between pairs of limbs. It indicates the timing between the first contacts of paw pairs (e.g., RF-LH) relative to the step cycle of the anchor paw. Phase dispersion is expressed as a percentage (%).
- − Step sequence patterns: The stepping patterns were categorized as normal when the animal sequentially placed its four paws in an alternating, cruciate, or rotating pattern.
- − Regulatory index (RI): This index measures the degree of coordination among the limbs and is expressed as a percentage (%).
2.8. Electrophysiological Recordings
2.9. Euthanasia
2.10. Statistical Analysis
3. Results
3.1. Expression of Mesenchymal Markers
3.2. Animals
3.3. Endogenous Inflammation
3.4. Behavioral Tests
3.5. Electrophysiological Recordings
4. Discussion
4.1. Activated Fat Reduced Neuroinflammation after SCI
4.2. Activated Fat Improves Sensorimotor Recovery
4.3. Activated Fat Restores Sensorimotor Loops and Their Supraspinal Command
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Golestani, A.; Shobeiri, P.; Sadeghi-Naini, M.; Jazayeri, S.B.; Maroufi, S.F.; Ghodsi, Z.; Dabbagh Ohadi, M.A.; Mohammadi, E.; Rahimi-Movaghar, V.; Ghodsi, S.M. Epidemiology of Traumatic Spinal Cord Injury in Developing Countries from 2009 to 2020: A Systematic Review and Meta-Analysis. Neuroepidemiology 2022, 56, 219–239. [Google Scholar] [CrossRef] [PubMed]
- Shavelle, R.M.; DeVivo, M.J.; Brooks, J.C.; Strauss, D.J.; Paculdo, D.R. Improvements in Long-Term Survival after Spinal Cord Injury? Arch. Phys. Med. Rehabil. 2015, 96, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, J.D.; Buzzell, A.; Gmünder, H.P.; Hug, K.; Jordan, X.; Moser, A.; Schubert, M.; Zwahlen, M.; Brinkhof, M.W.G.; the SwiSCI Study Group and the Swiss National Cohort. Comparison of All-Cause and Cause-Specific Mortality of Persons with Traumatic Spinal Cord Injuries to the General Swiss Population: Results from a National Cohort Study. Neuroepidemiology 2019, 52, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Dodd, W.; Motwani, K.; Small, C.; Pierre, K.; Patel, D.; Malnik, S.; Lucke-Wold, B.; Porche, K. Spinal Cord Injury and Neurogenic Lower Urinary Tract Dysfunction: Whatdo We Know and Where Are We Going? J. Mens. Health 2022, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- DeVivo, M.J.; Chen, Y.; Wen, H. Cause of Death Trends among Persons with Spinal Cord Injury in the United States: 1960–2017. Arch. Phys. Med. Rehabil. 2022, 103, 634–641. [Google Scholar] [CrossRef]
- Sekhon, L.H.S.; Fehlings, M.G. Epidemiology, Demographics, and Pathophysiology of Acute Spinal Cord Injury. Spine 2001, 26, S2–S12. [Google Scholar] [CrossRef]
- Krueger, H.; Noonan, V.K.; Trenaman, L.M.; Joshi, P.; Rivers, C.S. The Economic Burden of Traumatic Spinal Cord Injury in Canada. Chronic Dis. Inj. Can. 2013, 33, 113–122. [Google Scholar] [CrossRef]
- Malekzadeh, H.; Golpayegani, M.; Ghodsi, Z.; Sadeghi-Naini, M.; Asgardoon, M.; Baigi, V.; Vaccaro, A.R.; Rahimi-Movaghar, V. Direct Cost of Illness for Spinal Cord Injury: A Systematic Review. Glob. Spine J. 2022, 12, 1267–1281. [Google Scholar] [CrossRef]
- Cao, Y.; Krause, J. Economic Costs of Traumatic Spinal Cord Injury—A Population Based Study. Arch. Phys. Med. Rehabil. 2021, 102, e58–e59. [Google Scholar] [CrossRef]
- Cao, Y.; Krause, J.S. The Association between Secondary Health Conditions and Indirect Costs after Spinal Cord Injury. Spinal Cord 2021, 59, 306–310. [Google Scholar] [CrossRef]
- Hartkopp, A.; Brønnum-Hansen, H.; Seidenschnur, A.-M.; Biering-Sorensen, F. Suicide in a Spinal Cord Injured Population: Its Relation to Functional Status. Arch. Phys. Med. Rehabil. 1998, 79, 1356–1361. [Google Scholar] [CrossRef] [PubMed]
- Hawasli, A.H.; Rutlin, J.; Roland, J.L.; Murphy, R.K.J.; Song, S.-K.; Leuthardt, E.C.; Shimony, J.S.; Ray, W.Z. Spinal Cord Injury Disrupts Resting-State Networks in the Human Brain. J. Neurotrauma 2018, 35, 864–873. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ge, Y.; Li, J.; Dou, W.; Pan, Y. Impact of Injury Duration on a Sensorimotor Functional Network in Complete Spinal Cord Injury. J. Neurosci. Res. 2022, 100, 1765–1774. [Google Scholar] [CrossRef]
- Cao, Q.; Zhang, Y.P.; Iannotti, C.; DeVries, W.H.; Xu, X.-M.; Shields, C.B.; Whittemore, S.R. Functional and Electrophysiological Changes after Graded Traumatic Spinal Cord Injury in Adult Rat. Exp. Neurol. 2005, 191, S3–S16. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kim, J.H.; Oh, S.-K.; Baek, S.R.; Min, J.; Kim, Y.W.; Kim, S.T.; Woo, C.-W.; Jeon, S.R. Analysis of Equivalent Parameters of Two Spinal Cord Injury Devices: The New York University Impactor versus the Infinite Horizon Impactor. Spine J. 2016, 16, 1392–1403. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic Spinal Cord Injury: An Overview of Pathophysiology, Models and Acute Injury Mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Hachem, L.D.; Fehlings, M.G. Pathophysiology of Spinal Cord Injury. Neurosurg. Clin. N. Am. 2021, 32, 305–313. [Google Scholar] [CrossRef]
- Darrow, D.; Balser, D.; Netoff, T.I.; Krassioukov, A.; Phillips, A.; Parr, A.; Samadani, U. Epidural Spinal Cord Stimulation Facilitates Immediate Restoration of Dormant Motor and Autonomic Supraspinal Pathways after Chronic Neurologically Complete Spinal Cord Injury. J. Neurotrauma 2019, 36, 2325–2336. [Google Scholar] [CrossRef]
- Hofer, A.-S.; Schwab, M.E. Enhancing Rehabilitation and Functional Recovery after Brain and Spinal Cord Trauma with Electrical Neuromodulation. Curr. Opin. Neurol. 2019, 32, 828–835. [Google Scholar] [CrossRef]
- Rao, Y.; Zhu, W.; Liu, H.; Jia, C.; Zhao, Q.; Wang, Y. Clinical Application of Olfactory Ensheathing Cells in the Treatment of Spinal Cord Injury. J. Int. Med. Res. 2013, 41, 473–481. [Google Scholar] [CrossRef]
- Varma, A.K.; Das, A.; Wallace, G.; Barry, J.; Vertegel, A.A.; Ray, S.K.; Banik, N.L. Spinal Cord Injury: A Review of Current Therapy, Future Treatments, and Basic Science Frontiers. Neurochem. Res. 2013, 38, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Silva, N.A.; Sousa, N.; Reis, R.L.; Salgado, A.J. From Basics to Clinical: A Comprehensive Review on Spinal Cord Injury. Prog. Neurobiol. 2014, 114, 25–57. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic Spinal Cord Injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.P.; Warren, P.M.; Silver, J. The Biology of Regeneration Failure and Success after Spinal Cord Injury. Physiol. Rev. 2018, 98, 881–917. [Google Scholar] [CrossRef]
- Rabinstein, A.A. Traumatic Spinal Cord Injury. Contin. Lifelong Learn. Neurol. 2018, 24, 551–566. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yang, Y.; He, L.; Pang, M.; Luo, C.; Liu, B.; Rong, L. High-Dose Methylprednisolone for Acute Traumatic Spinal Cord Injury: A Meta-Analysis. Neurology 2019, 93, e841–e850. [Google Scholar] [CrossRef] [PubMed]
- Kwon, B. Pathophysiology and Pharmacologic Treatment of Acute Spinal Cord Injury. Spine J. 2004, 4, 451–464. [Google Scholar] [CrossRef]
- Pêgo, A.P.; Kubinova, S.; Cizkova, D.; Vanicky, I.; Mar, F.M.; Sousa, M.M.; Sykova, E. Regenerative Medicine for the Treatment of Spinal Cord Injury: More than Just Promises? J. Cell. Mol. Med. 2012, 16, 2564–2582. [Google Scholar] [CrossRef]
- Assinck, P.; Duncan, G.J.; Hilton, B.J.; Plemel, J.R.; Tetzlaff, W. Cell Transplantation Therapy for Spinal Cord Injury. Nat. Neurosci. 2017, 20, 637–647. [Google Scholar] [CrossRef]
- Flack, J.; Sharma, K.; Xie, J. Delving into the Recent Advancements of Spinal Cord Injury Treatment: A Review of Recent Progress. Neural Regen. Res. 2022, 17, 283. [Google Scholar] [CrossRef]
- Zipser, C.M.; Cragg, J.J.; Guest, J.D.; Fehlings, M.G.; Jutzeler, C.R.; Anderson, A.J.; Curt, A. Cell-Based and Stem-Cell-Based Treatments for Spinal Cord Injury: Evidence from Clinical Trials. Lancet Neurol. 2022, 21, 659–670. [Google Scholar] [CrossRef]
- Carelli, S.; Messaggio, F.; Canazza, A.; Hebda, D.M.; Caremoli, F.; Latorre, E.; Grimoldi, M.G.; Colli, M.; Bulfamante, G.; Tremolada, C.; et al. Characteristics and Properties of Mesenchymal Stem Cells Derived from Microfragmented Adipose Tissue. Cell Transpl. 2015, 24, 1233–1252. [Google Scholar] [CrossRef]
- Carelli, S.; Colli, M.; Vinci, V.; Caviggioli, F.; Klinger, M.; Gorio, A. Mechanical Activation of Adipose Tissue and Derived Mesenchymal Stem Cells: Novel Anti-Inflammatory Properties. Int. J. Mol. Sci. 2018, 19, 267. [Google Scholar] [CrossRef]
- Hey, G.; Willman, M.; Patel, A.; Goutnik, M.; Willman, J.; Lucke-Wold, B. Stem Cell Scaffolds for the Treatment of Spinal Cord Injury—A Review. Biomechanics 2023, 3, 322–342. [Google Scholar] [CrossRef]
- Siddiqui, A.M.; Khazaei, M.; Fehlings, M.G. Translating Mechanisms of Neuroprotection, Regeneration, and Repair to Treatment of Spinal Cord Injury. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2015; Volume 218, pp. 15–54. ISBN 978-0-444-63565-5. [Google Scholar]
- Tabakow, P.; Jarmundowicz, W.; Czapiga, B.; Fortuna, W.; Miedzybrodzki, R.; Czyz, M.; Huber, J.; Szarek, D.; Okurowski, S.; Szewczyk, P.; et al. Transplantation of Autologous Olfactory Ensheathing Cells in Complete Human Spinal Cord Injury. Cell Transpl. 2013, 22, 1591–1612. [Google Scholar] [CrossRef] [PubMed]
- Coelho, M.; Oliveira, T.; Fernandes, R. State of the Art Paper Biochemistry of Adipose Tissue: An Endocrine Organ. Arch. Med. Sci. 2013, 2, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, A.; Oh, K.-J.; Lee, S.C.; Kim, W.K.; Bae, K.-H. The Role of Adipose Tissue Mitochondria: Regulation of Mitochondrial Function for the Treatment of Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 4924. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Funcke, J.-B.; Zi, Z.; Zhao, S.; Straub, L.G.; Zhu, Y.; Zhu, Q.; Crewe, C.; An, Y.A.; Chen, S.; et al. Adipocyte Iron Levels Impinge on a Fat-Gut Crosstalk to Regulate Intestinal Lipid Absorption and Mediate Protection from Obesity. Cell Metab. 2021, 33, 1624–1639.e9. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, H.; Li, X.; Zhao, Y. Current Understanding of the Role of microRNAs from Adipose-Derived Extracellular Vesicles in Obesity. Biochem. Soc. Trans. 2022, 50, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, G.; Giuffrida, R.; Lo Furno, D.; Parrinello, N.; Forte, S.; Gulino, R.; Colarossi, C.; Schinocca, L.; Giuffrida, R.; Cardile, V.; et al. Potential Effect of CD271 on Human Mesenchymal Stromal Cell Proliferation and Differentiation. Int. J. Mol. Sci. 2015, 16, 15609–15624. [Google Scholar] [CrossRef]
- Palumbo, P.; Miconi, G.; Cinque, B.; La Torre, C.; Lombardi, F.; Zoccali, G.; Orsini, G.; Leocata, P.; Giuliani, M.; Cifone, M.G. In Vitro Evaluation of Different Methods of Handling Human Liposuction Aspirate and Their Effect on Adipocytes and Adipose Derived Stem Cells. J. Cell. Physiol. 2015, 230, 1974–1981. [Google Scholar] [CrossRef] [PubMed]
- Carelli, S.; Giallongo, T.; Rey, F.; Colli, M.; Tosi, D.; Bulfamante, G.; Di Giulio, A.M.; Gorio, A. Neuroprotection, Recovery of Function and Endogenous Neurogenesis in Traumatic Spinal Cord Injury Following Transplantation of Activated Adipose Tissue. Cells 2019, 8, 329. [Google Scholar] [CrossRef] [PubMed]
- Gimble, J.M.; Guilak, F. Adipose-Derived Adult Stem Cells: Isolation, Characterization, and Differentiation Potential. Cytotherapy 2003, 5, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Guo, X.; Zhao, S.; Jin, Y.; Zhou, F.; Yuan, P.; Cao, L.; Wang, J.; Qiu, Y.; Sun, C.; et al. Differentiation of Human Adipose-Derived Stem Cells into Neuron/Motoneuron-like Cells for Cell Replacement Therapy of Spinal Cord Injury. Cell Death Dis. 2019, 10, 597. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage Cells from Human Adipose Tissue: Implications for Cell-Based Therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef]
- Tsuji, W.; Rubin, J.P.; Marra, K.G. Adipose-Derived Stem Cells: Implications in Tissue Regeneration. World J. Stem Cells 2014, 6, 312. [Google Scholar] [CrossRef] [PubMed]
- Betz, V.M.; Sitoci-Ficici, K.H.; Uckermann, O.; Leipnitz, E.; Iltzsche, A.; Thirion, C.; Salomon, M.; Zwipp, H.; Schackert, G.; Betz, O.B.; et al. Gene-Activated Fat Grafts for the Repair of Spinal Cord Injury: A Pilot Study. Acta Neurochir. 2016, 158, 367–378. [Google Scholar] [CrossRef]
- Zhang, H.-T.; Luo, J.; Sui, L.-S.; Ma, X.; Yan, Z.-J.; Lin, J.-H.; Wang, Y.-S.; Chen, Y.-Z.; Jiang, X.-D.; Xu, R.-X. Effects of Differentiated Versus Undifferentiated Adipose Tissue-Derived Stromal Cell Grafts on Functional Recovery After Spinal Cord Contusion. Cell. Mol. Neurobiol. 2009, 29, 1283–1292. [Google Scholar] [CrossRef]
- Tien, N.L.B.; Hoa, N.D.; Thanh, V.V.; Van Thach, N.; Ngoc, V.T.N.; Dinh, T.C.; Phuong, T.N.T.; Toi, P.L.; Chu, D.T. Autologous Transplantation of Adipose-Derived Stem Cells to Treat Acute Spinal Cord Injury: Evaluation of Clinical Signs, Mental Signs, and Quality of Life. Open Access Maced. J. Med. Sci. 2019, 7, 4399–4405. [Google Scholar] [CrossRef]
- Walker, L.M.; Publicover, S.J.; Preston, M.R.; Said Ahmed, M.A.A.; El Haj, A.J. Calcium-Channel Activation and Matrix Protein Upregulation in Bone Cells in Response to Mechanical Strain. J. Cell. Biochem. 2000, 79, 648–661. [Google Scholar] [CrossRef]
- Vogel, V.; Sheetz, M. Local Force and Geometry Sensing Regulate Cell Functions. Nat. Rev. Mol. Cell Biol. 2006, 7, 265–275. [Google Scholar] [CrossRef]
- Pruitt, B.L.; Dunn, A.R.; Weis, W.I.; Nelson, W.J. Mechano-Transduction: From Molecules to Tissues. PLoS Biol. 2014, 12, e1001996. [Google Scholar] [CrossRef] [PubMed]
- El Haj, A.J.; Walker, L.M.; Preston, M.R.; Publicover, S.J. Mechanotransduction Pathways in Bone: Calcium Fluxes and the Role of Voltage-Operated Calcium Channels. Med. Biol. Eng. Comput. 1999, 37, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wei, S.; Chiu, J. Mechanical Regulation of Epigenetics in Vascular Biology and Pathobiology. J. Cell. Mol. Med. 2013, 17, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, M.; Alluin, O.; Trimaille, T.; Gigmes, D.; Marqueste, T.; Decherchi, P. Delayed Injection of a Physically Cross-Linked PNIPAAm-g-PEG Hydrogel in Rat Contused Spinal Cord Improves Functional Recovery. ACS Omega 2020, 5, 10247–10259. [Google Scholar] [CrossRef]
- Basso, D.M.; Beattie, M.S.; Bresnahan, J.C. A Sensitive and Reliable Locomotor Rating Scale for Open Field Testing in Rats. J. Neurotrauma 1995, 12, 1–21. [Google Scholar] [CrossRef]
- Metz, G.A.; Whishaw, I.Q. Cortical and Subcortical Lesions Impair Skilled Walking in the Ladder Rung Walking Test: A New Task to Evaluate Fore- and Hindlimb Stepping, Placing, and Co-Ordination. J. Neurosci. Methods 2002, 115, 169–179. [Google Scholar] [CrossRef]
- Semler, J.; Wellmann, K.; Wirth, F.; Stein, G.; Angelova, S.; Ashrafi, M.; Schempf, G.; Ankerne, J.; Ozsoy, O.; Ozsoy, U.; et al. Objective Measures of Motor Dysfunction after Compression Spinal Cord Injury in Adult Rats: Correlations with Locomotor Rating Scores. J. Neurotrauma 2011, 28, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, D.J.; Popovich, P.G. Inflammation and Its Role in Neuroprotection, Axonal Regeneration and Functional Recovery after Spinal Cord Injury. Exp. Neurol. 2008, 209, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Bonnet, M.; Trimaille, T.; Brezun, J.-M.; Feron, F.; Gigmes, D.; Marqueste, T.; Decherchi, P. Motor and Sensitive Recovery after Injection of a Physically Cross-Linked PNIPAAm-g-PEG Hydrogel in Rat Hemisectioned Spinal Cord. Mater. Sci. Eng. C 2020, 107, 110354. [Google Scholar] [CrossRef] [PubMed]
- Scheff, S.W.; Roberts, K.N. Infinite Horizon Spinal Cord Contusion Model. In Animal Models of Acute Neurological Injuries; Springer Protocols Handbooks Book Series; Humana Press: Totowa, NJ, USA, 2009; pp. 423–432. [Google Scholar]
- Li, Y.; Decherchi, P.; Raisman, G. Transplantation of Olfactory Ensheathing Cells into Spinal Cord Lesions Restores Breathing and Climbing. J. Neurosci. 2003, 23, 727–731. [Google Scholar] [CrossRef]
- Hamers, F.P.T.; Lankhorst, A.J.; Van Laar, T.J.; Veldhuis, W.B.; Gispen, W.H. Automated Quantitative Gait Analysis during Overground Locomotion in the Rat: Its Application to Spinal Cord Contusion and Transection Injuries. J. Neurotrauma 2001, 18, 187–201. [Google Scholar] [CrossRef]
- Cheng, H.; Almström, S.; Giménez-Llort, L.; Chang, R.; Ove Ögren, S.; Hoffer, B.; Olson, L. Gait Analysis of Adult Paraplegic Rats after Spinal Cord Repair. Exp. Neurol. 1997, 148, 544–557. [Google Scholar] [CrossRef] [PubMed]
- Seblani, M.; Ertlen, C.; Coyle, T.; Decherchi, P.; Brezun, J.-M. Combined Effect of Trifluoperazine and Sodium Cromoglycate on Reducing Acute Edema and Limiting Lasting Functional Impairments after Spinal Cord Injury in Rats. Exp. Neurol. 2024, 372, 114612. [Google Scholar] [CrossRef] [PubMed]
- Mendes, C.S.; Bartos, I.; Márka, Z.; Akay, T.; Márka, S.; Mann, R.S. Quantification of Gait Parameters in Freely Walking Rodents. BMC Biol. 2015, 13, 50. [Google Scholar] [CrossRef] [PubMed]
- Nawrotek, K.; Marqueste, T.; Modrzejewska, Z.; Zarzycki, R.; Rusak, A.; Decherchi, P. Thermogelling Chitosan Lactate Hydrogel Improves Functional Recovery after a C2 Spinal Cord Hemisection in Rat. J. Biomed. Mater. Res. A 2017, 105, 2004–2019. [Google Scholar] [CrossRef]
- Gueye, Y.; Marqueste, T.; Maurel, F.; Khrestchatisky, M.; Decherchi, P.; Feron, F. Cholecalciferol (Vitamin D₃) Improves Functional Recovery When Delivered during the Acute Phase after a Spinal Cord Trauma. J. Steroid Biochem. Mol. Biol. 2015, 154, 23–31. [Google Scholar] [CrossRef]
- Skinner, R.D.; Houle, J.D.; Reese, N.B.; Berry, C.L.; Garcia-Rill, E. Effects of Exercise and Fetal Spinal Cord Implants on the H-Reflex in Chronically Spinalized Adult Rats. Brain Res. 1996, 729, 127–131. [Google Scholar] [CrossRef]
- Bianco, J.; Gueye, Y.; Marqueste, T.; Alluin, O.; Risso, J.-J.; Garcia, S.; Lavault, M.-N.; Khrestchatisky, M.; Feron, F.; Decherchi, P. Vitamin D₃ Improves Respiratory Adjustment to Fatigue and H-Reflex Responses in Paraplegic Adult Rats. Neuroscience 2011, 188, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Pertici, V.; Trimaille, T.; Laurin, J.; Felix, M.-S.; Marqueste, T.; Pettmann, B.; Chauvin, J.-P.; Gigmes, D.; Decherchi, P. Repair of the Injured Spinal Cord by Implantation of a Synthetic Degradable Block Copolymer in Rat. Biomaterials 2014, 35, 6248–6258. [Google Scholar] [CrossRef]
- Caron, G.; Decherchi, P.; Marqueste, T. Does Metabosensitive Afferent Fibers Activity Differ from Slow- and Fast-Twitch Muscles? Exp. Brain Res. 2015, 233, 2549–2554. [Google Scholar] [CrossRef]
- Hellenbrand, D.J.; Quinn, C.M.; Piper, Z.J.; Morehouse, C.N.; Fixel, J.A.; Hanna, A.S. Inflammation after Spinal Cord Injury: A Review of the Critical Timeline of Signaling Cues and Cellular Infiltration. J. Neuroinflamm. 2021, 18, 284. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.-Y.; Huang, F.-F.; Xie, Y.-Y.; Wang, W.-Q.; Wang, L.-D.; Mu, D.; Cui, Y.; Wang, B. Dynamic Changes in the Systemic Immune Responses of Spinal Cord Injury Model Mice. Neural Regen. Res. 2021, 16, 382. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D.; Giusti, P.; Facci, L. Microglia and Mast Cells: Two Tracks on the Road to Neuroinflammation. FASEB J. 2012, 26, 3103–3117. [Google Scholar] [CrossRef] [PubMed]
- Dumont, C.M.; Margul, D.J.; Shea, L.D. Tissue Engineering Approaches to Modulate the Inflammatory Milieu Following Spinal Cord Injury. Cells Tissues Organs 2016, 202, 52–66. [Google Scholar] [CrossRef]
- Menezes, K.; Rosa, B.G.; Freitas, C.; Da Cruz, A.S.; De Siqueira Santos, R.; Nascimento, M.A.; Alves, D.V.L.; Bonamino, M.; Rossi, M.I.; Borojevic, R.; et al. Human Mesenchymal Stromal/Stem Cells Recruit Resident Pericytes and Induce Blood Vessels Maturation to Repair Experimental Spinal Cord Injury in Rats. Sci. Rep. 2020, 10, 19604. [Google Scholar] [CrossRef]
- Villanova Junior, J.A.; Fracaro, L.; Rebelatto, C.L.K.; Da Silva, A.J.; Barchiki, F.; Senegaglia, A.C.; Dominguez, A.C.; De Moura, S.A.B.; Pimpão, C.T.; Brofman, P.R.S.; et al. Recovery of Motricity and Micturition after Transplantation of Mesenchymal Stem Cells in Rats Subjected to Spinal Cord Injury. Neurosci. Lett. 2020, 734, 135134. [Google Scholar] [CrossRef] [PubMed]
- Vialle, E.N.; Fracaro, L.; Barchiki, F.; Dominguez, A.C.; Arruda, A.D.O.; Olandoski, M.; Brofman, P.R.S.; Kuniyoshi Rebelatto, C.L. Human Adipose-Derived Stem Cells Reduce Cellular Damage after Experimental Spinal Cord Injury in Rats. Biomedicines 2023, 11, 1394. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Jones, E.Y.; Fugger, L.; Strominger, J.L.; Siebold, C. MHC Class II Proteins and Disease: A Structural Perspective. Nat. Rev. Immunol. 2006, 6, 271–282. [Google Scholar] [CrossRef]
- Tu, Z.; Li, Q.; Bu, H.; Lin, F. Mesenchymal Stem Cells Inhibit Complement Activation by Secreting Factor H. Stem Cells Dev. 2010, 19, 1803–1809. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lam, K.S.L.; Yau, M.; Xu, A. Post-Translational Modifications of Adiponectin: Mechanisms and Functional Implications. Biochem. J. 2008, 409, 623–633. [Google Scholar] [CrossRef]
- Kusminski, C.M.; McTernan, P.G.; Schraw, T.; Kos, K.; O’Hare, J.P.; Ahima, R.; Kumar, S.; Scherer, P.E. Adiponectin Complexes in Human Cerebrospinal Fluid: Distinct Complex Distribution from Serum. Diabetologia 2007, 50, 634–642. [Google Scholar] [CrossRef]
- Fernández-Martos, C.M.; González, P.; Rodriguez, F.J. Acute Leptin Treatment Enhances Functional Recovery after Spinal Cord Injury. PLoS ONE 2012, 7, e35594. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Tian, X.; Mo, B.; Xu, H.; Zhang, L.; Huang, L.; Yao, S.; Huang, Z.; Wang, Y.; Xie, H.; et al. Adipose Mesenchymal Stem Cell Transplantation Alleviates Spinal Cord Injury-Induced Neuroinflammation Partly by Suppressing the Jagged1/Notch Pathway. Stem Cell Res. Ther. 2020, 11, 212. [Google Scholar] [CrossRef]
- Scheff, S.W.; Rabchevsky, A.G.; Fugaccia, I.; Main, J.A.; Lumpp, J.E. Experimental Modeling of Spinal Cord Injury: Characterization of a Force-Defined Injury Device. J. Neurotrauma 2003, 20, 179–193. [Google Scholar] [CrossRef]
- Bovonsunthonchai, S.; Hiengkaew, V.; Vachalathiti, R.; Vongsirinavarat, M.; Tretriluxana, J. Effect of Speed on the Upper and Contralateral Lower Limb Coordination during Gait in Individuals with Stroke. Kaohsiung J. Med. Sci. 2012, 28, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-C.; Lim, J.-H. The Effects of Coordinative Locomotor Training on Coordination and Gait in Chronic Stroke Patients: A Randomized Controlled Pilot Trial. J. Exerc. Rehabil. 2018, 14, 1010–1016. [Google Scholar] [CrossRef]
- Naeimi, A.; Zaminy, A.; Amini, N.; Balabandi, R.; Golipoor, Z. Effects of Melatonin-Pretreated Adipose-Derived Mesenchymal Stem Cells (MSC) in an Animal Model of Spinal Cord Injury. BMC Neurosci. 2022, 23, 65. [Google Scholar] [CrossRef]
- Li, F.; Liu, H.; Zhang, K.; Xiao, D.-J.; Wang, C.; Wang, Y.-S. Adipose-Derived Stromal Cells Improve Functional Recovery after Spinal Cord Injury through TGF-Β1/Smad3/PLOD2 Pathway Activation. Aging 2021, 13, 4370–4387. [Google Scholar] [CrossRef]
- Li, J.; Ji, Z.; Wang, Y.; Li, T.; Luo, J.; Li, J.; Shi, X.; Li, L.; He, L.; Wu, W. Human Adipose-Derived Stem Cells Combined with Nano-Hydrogel Promote Functional Recovery after Spinal Cord Injury in Rats. Biology 2022, 11, 781. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, L.; Liu, M.; Gao, G.; Zhao, W.; Fu, Q.; Wang, Y. Implantation of Adipose-Derived Mesenchymal Stem Cell Sheets Promotes Axonal Regeneration and Restores Bladder Function after Spinal Cord Injury. Stem Cell Res. Ther. 2022, 13, 503. [Google Scholar] [CrossRef]
- Keikhaei, R.; Abdi, E.; Darvishi, M.; Ghotbeddin, Z.; Hamidabadi, H.G. Combined Treatment of High-intensity Interval Training with Neural Stem Cell Generation on Contusive Model of Spinal Cord Injury in Rats. Brain Behav. 2023, 13, e3043. [Google Scholar] [CrossRef]
- Hur, J.W.; Cho, T.-H.; Park, D.-H.; Lee, J.-B.; Park, J.-Y.; Chung, Y.-G. Intrathecal Transplantation of Autologous Adipose-Derived Mesenchymal Stem Cells for Treating Spinal Cord Injury: A Human Trial. J. Spinal Cord Med. 2016, 39, 655–664. [Google Scholar] [CrossRef]
- Tomita, K.; Madura, T.; Sakai, Y.; Yano, K.; Terenghi, G.; Hosokawa, K. Glial Differentiation of Human Adipose-Derived Stem Cells: Implications for Cell-Based Transplantation Therapy. Neuroscience 2013, 236, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, E.; Dickman, R. Growth Factors and Their Peptide Mimetics for Treatment of Traumatic Brain Injury. Bioorg. Med. Chem. 2023, 90, 117368. [Google Scholar] [CrossRef] [PubMed]
- Andia, I.; Maffulli, N.; Burgos-Alonso, N. Stromal Vascular Fraction Technologies and Clinical Applications. Expert Opin. Biol. Ther. 2019, 19, 1289–1305. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Chen, S.; Zhao, Z.; Luo, Y.; Hou, Y.; Li, H.; He, L.; Zhou, L.; Wu, W. Effect of VEGF on Inflammatory Regulation, Neural Survival, and Functional Improvement in Rats Following a Complete Spinal Cord Transection. Front. Cell. Neurosci. 2017, 11, 381. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Kang, S.M.; Kim, E.; Kim, C.-H.; Song, H.-T.; Lee, J.E. Adiponectin Receptor-Mediated Signaling Ameliorates Cerebral Cell Damage and Regulates the Neurogenesis of Neural Stem Cells at High Glucose Concentrations: An In Vivo and In Vitro Study. Cell Death Dis. 2015, 6, e1844. [Google Scholar] [CrossRef]
- Bloemer, J.; Pinky, P.D.; Govindarajulu, M.; Hong, H.; Judd, R.; Amin, R.H.; Moore, T.; Dhanasekaran, M.; Reed, M.N.; Suppiramaniam, V. Role of Adiponectin in Central Nervous System Disorders. Neural Plast. 2018, 2018, 4593530. [Google Scholar] [CrossRef]
- Formolo, D.A.; Cheng, T.; Yu, J.; Kranz, G.S.; Yau, S.-Y. Central Adiponectin Signaling—A Metabolic Regulator in Support of Brain Plasticity. Brain Plast. 2022, 8, 79–96. [Google Scholar] [CrossRef]
- Ouchi, N.; Walsh, K. Adiponectin as an Anti-Inflammatory Factor. Clin. Chim. Acta 2007, 380, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Shibata, R.; Ouchi, N.; Kihara, S.; Sato, K.; Funahashi, T.; Walsh, K. Adiponectin Stimulates Angiogenesis in Response to Tissue Ischemia through Stimulation of AMP-Activated Protein Kinase Signaling. J. Biol. Chem. 2004, 279, 28670–28674. [Google Scholar] [CrossRef]
- Miao, J.; Shen, L.; Tang, Y.; Wang, Y.; Tao, M.; Jin, K.; Zhao, Y.; Yang, G. Overexpression of Adiponectin Improves Neurobehavioral Outcomes after Focal Cerebral Ischemia in Aged Mice. CNS Neurosci. Ther. 2013, 19, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Wang, S.; Ma, Y.; Lim, G.; Kim, H.; Mao, J. Leptin Enhances NMDA-Induced Spinal Excitation in Rats: A Functional Link between Adipocytokine and Neuropathic Pain. Pain 2011, 152, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Li, X.; Sun, G.; Li, S.; Liang, S.; Li, Z.; Li, B.; Xia, M. Protective Effect of Leptin-Mediated Caveolin-1 Expression on Neurons after Spinal Cord Injury. Cell Calcium 2018, 76, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Decker, J.T.; Margul, D.J.; Smith, D.R.; Cummings, B.J.; Anderson, A.J.; Shea, L.D. Local Immunomodulation with Anti-Inflammatory Cytokine-Encoding Lentivirus Enhances Functional Recovery after Spinal Cord Injury. Mol. Ther. 2018, 26, 1756–1770. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Glaser, J.; Liu, M.T.; Lane, T.E.; Keirstead, H.S. Reducing Inflammation Decreases Secondary Degeneration and Functional Deficit after Spinal Cord Injury. Exp. Neurol. 2003, 184, 456–463. [Google Scholar] [CrossRef]
- Alexander, J.K.; Popovich, P.G. Neuroinflammation in Spinal Cord Injury: Therapeutic Targets for Neuroprotection and Regeneration. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2009; Volume 175, pp. 125–137. ISBN 978-0-12-374511-8. [Google Scholar]
- Kotaka, K.; Nagai, J.; Hensley, K.; Ohshima, T. Lanthionine Ketimine Ester Promotes Locomotor Recovery after Spinal Cord Injury by Reducing Neuroinflammation and Promoting Axon Growth. Biochem. Biophys. Res. Commun. 2017, 483, 759–764. [Google Scholar] [CrossRef]
- Brambilla, R.; Bracchi-Ricard, V.; Hu, W.-H.; Frydel, B.; Bramwell, A.; Karmally, S.; Green, E.J.; Bethea, J.R. Inhibition of Astroglial Nuclear Factor κB Reduces Inflammation and Improves Functional Recovery after Spinal Cord Injury. J. Exp. Med. 2005, 202, 145–156. [Google Scholar] [CrossRef]
- Hiersemenzel, L.-P.; Curt, A.; Dietz, V. From Spinal Shock to Spasticity: Neuronal Adaptations to a Spinal Cord Injury. Neurology 2000, 54, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Decherchi, P.; Dousset, E.; Jammes, Y. Respiratory and Cardiovascular Responses Evoked by Tibialis Anterior Muscle Afferent Fibers in Rats. Exp. Brain Res. 2007, 183, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Pertici, V.; Amendola, J.; Laurin, J.; Gigmes, D.; Madaschi, L.; Carelli, S.; Marqueste, T.; Gorio, A.; Decherchi, P. The Use of Poly(N-[2-Hydroxypropyl]-Methacrylamide) Hydrogel to Repair a T10 Spinal Cord Hemisection in Rat: A Behavioural, Electrophysiological and Anatomical Examination. ASN Neuro 2013, 5, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Thompson, F.J.; Reier, P.J.; Lucas, C.C.; Parmer, R. Altered Patterns of Reflex Excitability Subsequent to Contusion Injury of the Rat Spinal Cord. J. Neurophysiol. 1992, 68, 1473–1486. [Google Scholar] [CrossRef]
- Lee, J.K.; Emch, G.S.; Johnson, C.S.; Wrathall, J.R. Effect of Spinal Cord Injury Severity on Alterations of the H-Reflex. Exp. Neurol. 2005, 196, 430–440. [Google Scholar] [CrossRef]
- Reese, N.B.; Skinner, R.D.; Mitchell, D.; Yates, C.; Barnes, C.N.; Kiser, T.S.; Garcia-Rill, E. Restoration of Frequency-Dependent Depression of the H-Reflex by Passive Exercise in Spinal Rats. Spinal Cord 2006, 44, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Yates, C.C.; Charlesworth, A.; Reese, N.B.; Skinner, R.D.; Garcia-Rill, E. The Effects of Passive Exercise Therapy Initiated Prior to or after the Development of Hyperreflexia Following Spinal Transection. Exp. Neurol. 2008, 213, 405–409. [Google Scholar] [CrossRef]
- Bennett, D.J.; Li, Y.; Harvey, P.J.; Gorassini, M. Evidence for Plateau Potentials in Tail Motoneurons of Awake Chronic Spinal Rats with Spasticity. J. Neurophysiol. 2001, 86, 1972–1982. [Google Scholar] [CrossRef]
- Schindler-Ivens, S.; Shields, R.K. Low Frequency Depression of H-Reflexes in Humans with Acute and Chronic Spinal-Cord Injury. Exp. Brain Res. 2000, 133, 233–241. [Google Scholar] [CrossRef]
- Yuan, X.; Yuan, W.; Ding, L.; Shi, M.; Luo, L.; Wan, Y.; Oh, J.; Zhou, Y.; Bian, L.; Deng, D.Y.B. Cell-Adaptable Dynamic Hydrogel Reinforced with Stem Cells Improves the Functional Repair of Spinal Cord Injury by Alleviating Neuroinflammation. Biomaterials 2021, 279, 121190. [Google Scholar] [CrossRef]
- Mukhamedshina, Y.O.; Akhmetzyanova, E.R.; Kostennikov, A.A.; Zakirova, E.Y.; Galieva, L.R.; Garanina, E.E.; Rogozin, A.A.; Kiassov, A.P.; Rizvanov, A.A. Adipose-Derived Mesenchymal Stem Cell Application Combined with Fibrin Matrix Promotes Structural and Functional Recovery Following Spinal Cord Injury in Rats. Front. Pharmacol. 2018, 9, 343. [Google Scholar] [CrossRef] [PubMed]
- Pousti, F.; Ahmadi, R.; Mirahmadi, F.; Hosseinmardi, N.; Rohampour, K. Adiponectin Modulates Synaptic Plasticity in Hippocampal Dentate Gyrus. Neurosci. Lett. 2018, 662, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jo, J.; Song, J. Adiponectin Improves Long-Term Potentiation in the 5XFAD Mouse Brain. Sci. Rep. 2019, 9, 8918. [Google Scholar] [CrossRef] [PubMed]
- Ng, R.C.-L.; Jian, M.; Ma, O.K.-F.; Bunting, M.; Kwan, J.S.-C.; Zhou, G.-J.; Senthilkumar, K.; Iyaswamy, A.; Chan, P.-K.; Li, M.; et al. Chronic Oral Administration of adipoRon Reverses Cognitive Impairments and Ameliorates Neuropathology in an Alzheimer’s Disease Mouse Model. Mol. Psychiatry 2021, 26, 5669–5689. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, J.; Wang, J.; Liu, C.; Yan, H. AdipoRon Improves Cognitive Dysfunction of Alzheimer’s Disease and Rescues Impaired Neural Stem Cell Proliferation through AdipoR1/AMPK Pathway. Exp. Neurol. 2020, 327, 113249. [Google Scholar] [CrossRef] [PubMed]
- Côté, M.-P.; Gossard, J.-P. Step Training-Dependent Plasticity in Spinal Cutaneous Pathways. J. Neurosci. 2004, 24, 11317–11327. [Google Scholar] [CrossRef]
- Gómez-Pinilla, F.; Ying, Z.; Roy, R.R.; Hodgson, J.; Edgerton, V.R. Afferent Input Modulates Neurotrophins and Synaptic Plasticity in the Spinal Cord. J. Neurophysiol. 2004, 92, 3423–3432. [Google Scholar] [CrossRef]
- Ying, Z.; Roy, R.R.; Edgerton, V.R.; Gómez-Pinilla, F. Exercise Restores Levels of Neurotrophins and Synaptic Plasticity Following Spinal Cord Injury. Exp. Neurol. 2005, 193, 411–419. [Google Scholar] [CrossRef]
- Ying, Z.; Roy, R.R.; Zhong, H.; Zdunowski, S.; Edgerton, V.R.; Gomez-Pinilla, F. BDNF–Exercise Interactions in the Recovery of Symmetrical Stepping after a Cervical Hemisection in Rats. Neuroscience 2008, 155, 1070–1078. [Google Scholar] [CrossRef]
- Boulenguez, P.; Liabeuf, S.; Bos, R.; Bras, H.; Jean-Xavier, C.; Brocard, C.; Stil, A.; Darbon, P.; Cattaert, D.; Delpire, E.; et al. Down-Regulation of the Potassium-Chloride Cotransporter KCC2 Contributes to Spasticity after Spinal Cord Injury. Nat. Med. 2010, 16, 302–307. [Google Scholar] [CrossRef]
- European Medicines Agency. Available online: www.ema.europa.eu (accessed on 1 January 2019).
- US Food & Drug Administration. Available online: www.fda.gov (accessed on 1 January 2019).
- Hyun, I.; Lindvall, O.; Ährlund-Richter, L.; Cattaneo, E.; Cavazzana-Calvo, M.; Cossu, G.; De Luca, M.; Fox, I.J.; Gerstle, C.; Goldstein, R.A.; et al. New ISSCR Guidelines Underscore Major Principles for Responsible Translational Stem Cell Research. Cell Stem Cell 2008, 3, 607–609. [Google Scholar] [CrossRef]



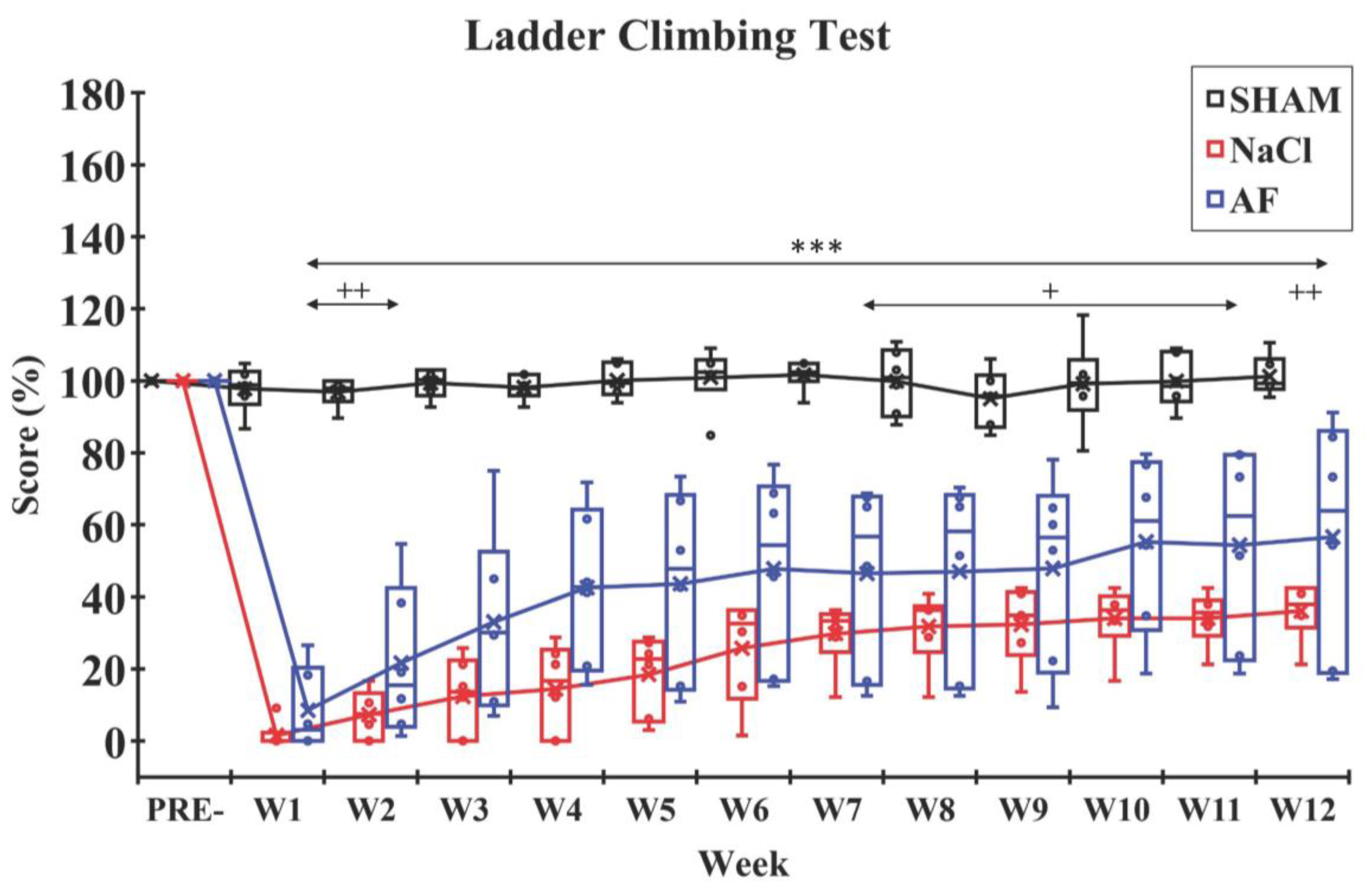
 indicate the mean and individual response, respectively. n = 5 per group.
indicate the mean and individual response, respectively. n = 5 per group.
indicate the mean and individual response, respectively. n = 5 per group.
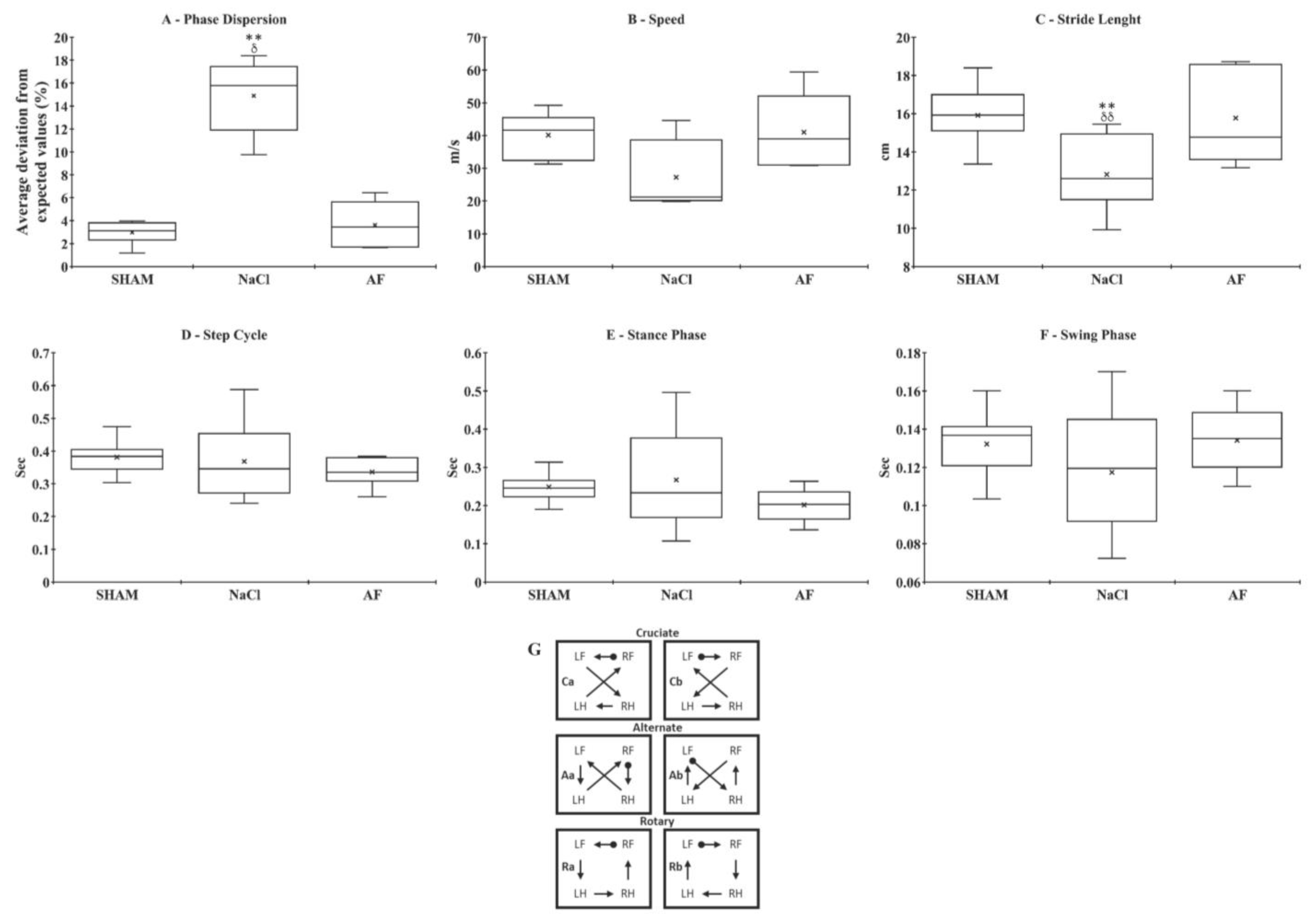
indicate the mean and individual response, respectively. n = 5 per group. indicate the mean and individual response, respectively. n = 6 per group.
indicate the mean and individual response, respectively. n = 6 per group.
indicate the mean and individual response, respectively. n = 6 per group.
indicate the mean and individual response, respectively. n = 6 per group. indicate the mean and individual response, respectively. n = 6 per group.
indicate the mean and individual response, respectively. n = 6 per group.
indicate the mean and individual response, respectively. n = 6 per group.
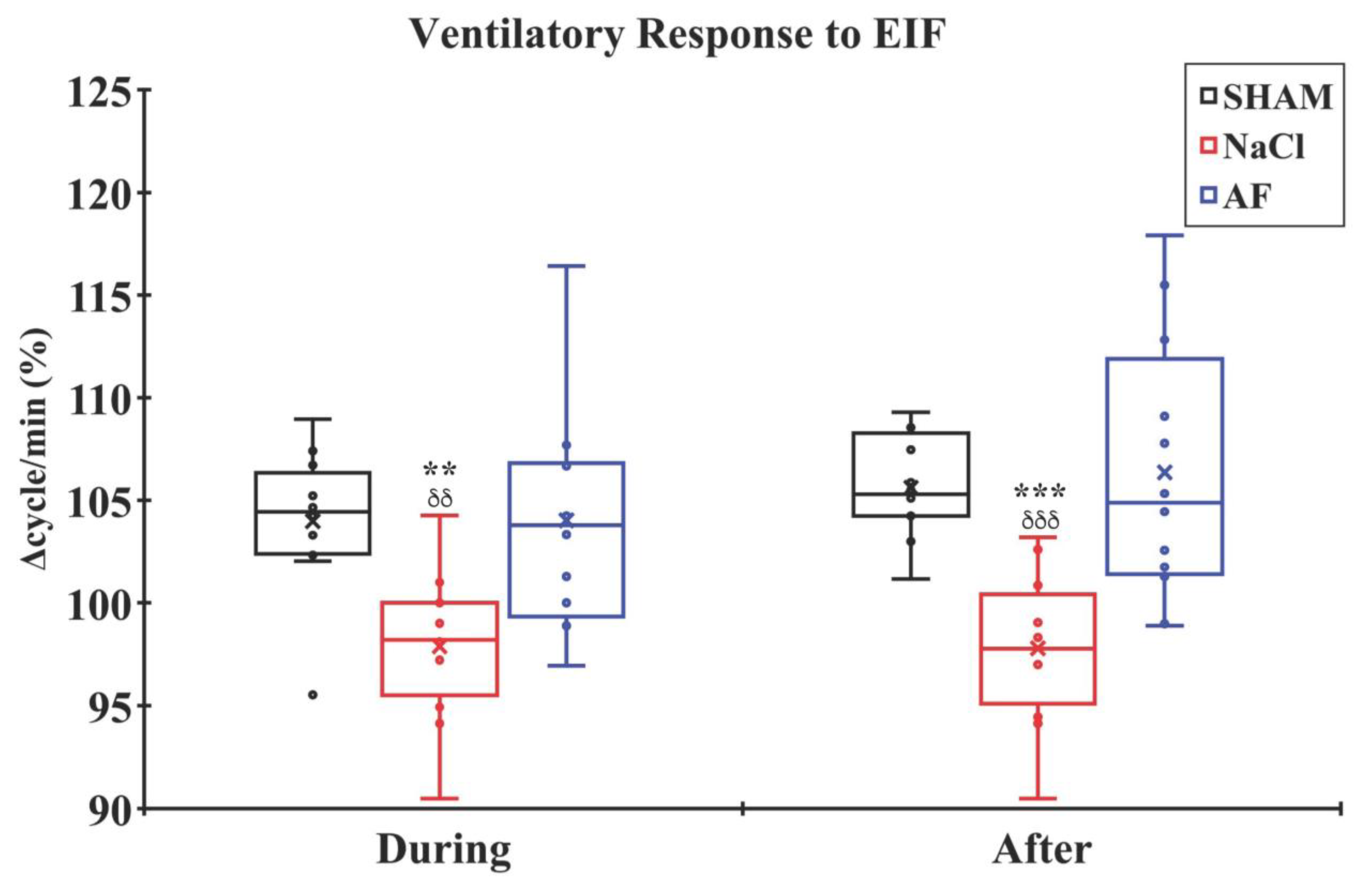
indicate the mean and individual response, respectively. n = 6 per group.
 indicate the mean and individual response, respectively. n = 6 per group.
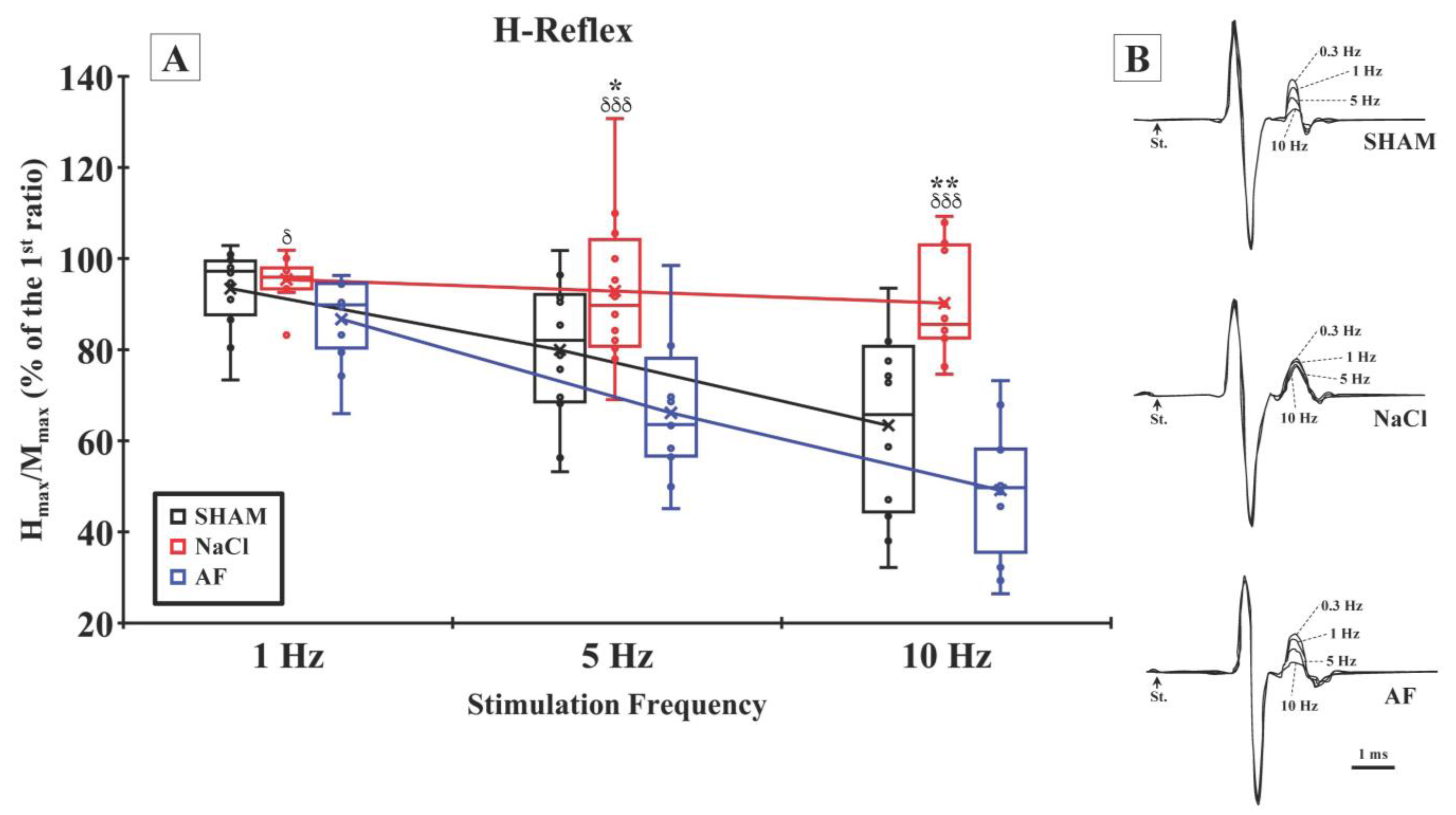
indicate the mean and individual response, respectively. n = 6 per group.
indicate the mean and individual response, respectively. n = 6 per group.
indicate the mean and individual response, respectively. n = 6 per group. indicate the mean and individual response, respectively. n = 6 per group. (B) Representative examples of H-reflex recorded at different stimulation frequencies in the SHAM (upper traces), NaCl (middle traces), and AF (bottom traces) groups.
indicate the mean and individual response, respectively. n = 6 per group. (B) Representative examples of H-reflex recorded at different stimulation frequencies in the SHAM (upper traces), NaCl (middle traces), and AF (bottom traces) groups.
indicate the mean and individual response, respectively. n = 6 per group. (B) Representative examples of H-reflex recorded at different stimulation frequencies in the SHAM (upper traces), NaCl (middle traces), and AF (bottom traces) groups.
indicate the mean and individual response, respectively. n = 6 per group. (B) Representative examples of H-reflex recorded at different stimulation frequencies in the SHAM (upper traces), NaCl (middle traces), and AF (bottom traces) groups.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonnet, M.; Ertlen, C.; Seblani, M.; Brezun, J.-M.; Coyle, T.; Cereda, C.; Zuccotti, G.; Colli, M.; Desouches, C.; Decherchi, P.; et al. Activated Human Adipose Tissue Transplantation Promotes Sensorimotor Recovery after Acute Spinal Cord Contusion in Rats. Cells 2024, 13, 182. https://doi.org/10.3390/cells13020182
Bonnet M, Ertlen C, Seblani M, Brezun J-M, Coyle T, Cereda C, Zuccotti G, Colli M, Desouches C, Decherchi P, et al. Activated Human Adipose Tissue Transplantation Promotes Sensorimotor Recovery after Acute Spinal Cord Contusion in Rats. Cells. 2024; 13(2):182. https://doi.org/10.3390/cells13020182
Chicago/Turabian StyleBonnet, Maxime, Céline Ertlen, Mostafa Seblani, Jean-Michel Brezun, Thelma Coyle, Cristina Cereda, Gianvincenzo Zuccotti, Mattia Colli, Christophe Desouches, Patrick Decherchi, and et al. 2024. "Activated Human Adipose Tissue Transplantation Promotes Sensorimotor Recovery after Acute Spinal Cord Contusion in Rats" Cells 13, no. 2: 182. https://doi.org/10.3390/cells13020182
APA StyleBonnet, M., Ertlen, C., Seblani, M., Brezun, J. -M., Coyle, T., Cereda, C., Zuccotti, G., Colli, M., Desouches, C., Decherchi, P., Carelli, S., & Marqueste, T. (2024). Activated Human Adipose Tissue Transplantation Promotes Sensorimotor Recovery after Acute Spinal Cord Contusion in Rats. Cells, 13(2), 182. https://doi.org/10.3390/cells13020182