1. Introduction
Allergic disorders represent a persistent challenge to global public health [
1]. Several strategies are currently available for therapy of established immunoglobulin E (IgE)-mediated allergies, such as pharmacotherapy and biologics, which neutralize IgE or inflammatory cytokines [
2,
3,
4]. Allergen-specific immunotherapy (AIT) represents the only causative and disease-modifying treatment for allergic patients [
5]. It comprises the subcutaneous [
6] or sublingual [
7] administration of natural allergen extracts and is associated with the induction of protective allergen-specific IgG and IgA antibodies, but also changes allergen-specific cellular responses of the innate and adaptive immune system [
8]. The long-term clinical efficacy of AIT was confirmed in many clinical trials [
9]. However, effective prophylactic treatment strategies for primary allergy prevention are still needed. In recent years, IgE sensitizations in early childhood have been analyzed using allergen microarrays, suggesting that allergic sensitization in children initially develops against a few molecules [
10,
11,
12]. Since the corresponding allergen sequences are known, prophylactic approaches based on defined allergen molecules become feasible. In recent years, cell therapy approaches have emerged with the potential for targeted and sustainable interventions [
13]. We aimed to develop a cell therapy strategy for achieving long-lasting prevention of IgE-mediated allergy by inducing robust allergen-specific tolerance.
In proof-of-concept studies, we have shown that irradiated BALB/c mice infused with autologous bone marrow cells (BM), that have either been transfected with a gene encoding the major grass pollen allergen, Phl p 5, or that have been retrieved from a transgenic mouse ubiquitously expressing Phl p 5, do not develop Phl p 5-specific IgE despite repetitive allergen immunizations [
14,
15,
16]. The treatment also fully prevented Phl p 5-specific T cell responses and allergic airway inflammation, whereas immune responses towards the control allergen Bet v 1 remained unaffected. We subsequently developed an irradiation-free cell therapy protocol that also successfully prevents the occurrence of allergies in an allergen-specific fashion [
15]. In these studies, we conditioned recipients only with a short course of mTOR inhibition and costimulation blockade. Moreover, we recently demonstrated that tolerance to Phl p 5 can also be induced by the transfer of Phl p 5-expressing CD19
+ B cells instead of BM cells [
17]. The robustness of this approach is underlined by the fact that Phl p 5 represents a highly immunogenic and allergenic molecule in allergic patients (11). Moreover, the allergen composition could be adapted to sensitization patterns detected early in childhood, e.g., by combining the relevant epitopes in one molecule [
10,
11,
12].
However, for eventual translation into clinical practice, major protocol enhancements would be a prerequisite. Notably, retroviral transduction of donor cells needs to be avoided since its use is currently insufficiently safe for non-life-threatening diseases. Instead, peptides or proteins stably decorating the surface of cells could be used, which might be achieved with chemical coupling.
The crosslinker 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC) has been commonly used to couple carboxyl groups to primary amines. The intravenous (iv) infusion of EDC-treated antigen (Ag)-coupled peripheral blood or splenic leukocytes has demonstrated immunomodulatory effects in rodent models of multiple sclerosis [
18], type I diabetes [
19] and transplant rejection [
20,
21]. In addition, EDC-coupling of peanut extract and ovalbumin was shown to alleviate allergic symptoms and decrease allergen-specific IgE production in a mouse model [
22]. A phase 1 clinical trial has affirmed the safety of iv infusions of encephalitogenic peptides coupled to autologous leukocytes via EDC treatment [
23]. Although the mechanism is not fully clear, EDC is known to elicit antigen-specific immunoregulatory responses, with both Treg and dendritic cells (DC) playing a central role [
24].
Alternatively, lipophilic anchors can be used for coupling antigens to the cell surface. 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-poly (ethylene glycol)-maleimide (DSPE-PEG-Mal) is one of the most commonly used reactive phospholipids for the conjugation of peptides or other ligands to plasma membranes, liposomes, or other lipid nanoparticles, facilitating otherwise challenging access to homogeneous, lipidated proteins [
25]. In contrast to EDC-coupling, this strategy represents a more physiological approach, allowing a directed anchoring of the protein in the membrane and presumably better retaining the function of other molecules on the cell surface.
Herein, we investigated chemically linking a major allergen to the cell surface of immune cells for use in prophylactic cell therapy.
2. Materials and Methods
2.1. Mice
Female BALB/c mice were purchased from Charles River Laboratories (Sulzfeld, Germany) and kept at the Centre of Biomedical Research under specific pathogen-free (SPF) conditions. Mice were used between 6 and 12 weeks of age with a body weight of 20 +/− 2 g. As a control, a previously described (12) transgenic mouse ubiquitously expressing Phl p 5 and green fluorescent protein (GFP) on a BALB/c background (referred to as Phl p 5-tg) was used. Phl p 5-tg mice were bred at the animal facility of the Medical University of Vienna. The transgene was confirmed by PCR as described (15). GFP expression acted as a surrogate for membrane-bound Phl p 5, whose presence was additionally confirmed by flow cytometry using an anti-Phl p 5 primary antibody. All experiments were approved by the local review board of the Medical University of Vienna and approved by the Austrian Federal Ministry of Science, Research and Economy, GZ: BMWFW-66.009/2021-0,276.441) and were performed in accordance with national and international guidelines for laboratory animal care.
2.2. Generation of Recombinant Phl p 5 for EDC Coupling
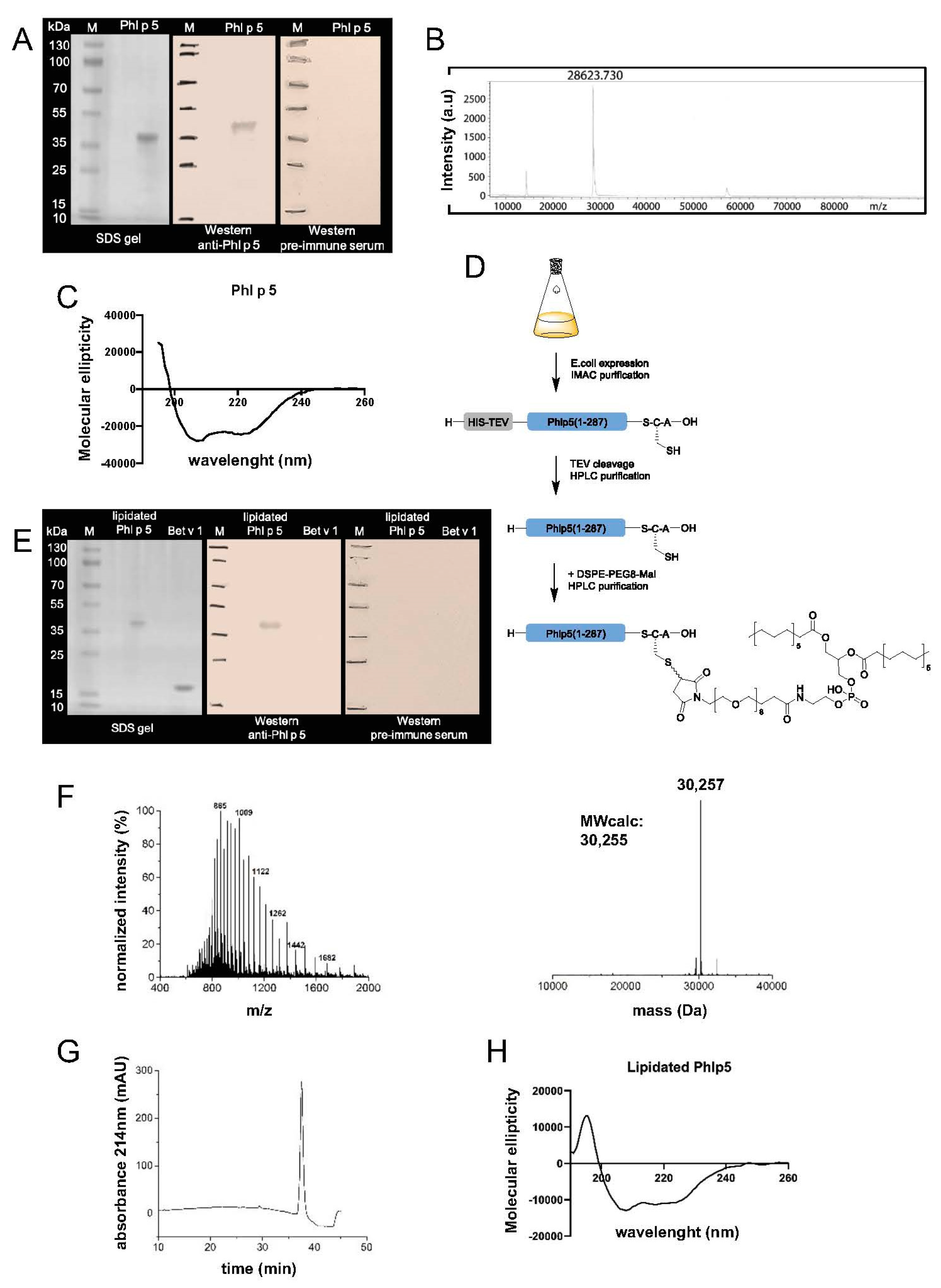
The gene encoding the major grass pollen allergen Phl p 5a was codon-optimized for
Escherichia coli (
E. coli) expression, synthesized by ATG:biosynthetics (Merzhausen, Germany) and cloned into the expression plasmid pET17b (Novagen, Darmstadt, Germany) along with an N-terminal 8x histidine tag followed by a Tobacco Etch Virus (TEV) cleavage site (
Supplementary Figure S1). The recombinant protein was expressed in
E. coli strain BL21(DE3) (Agilent, Santa Clara, CA, USA) after the addition of 1 mM isopropyl-ß-thiogalactopyranoside for 4 h at 37 °C. The protein was purified from the soluble fraction on a nickel-nitriloacetic acid (Ni-NTA) Superflow Column (Qiagen, Venlo, Netherlands) according to the manufacturer’s instructions. The soluble protein was eluted using 250 mM imidazole and dialyzed against phosphate-buffered saline (PBS) overnight in 3 mL Slide-A-lyzer G3 Cassettes with a 3.5 K molecular weight cut-off (MWco) (Thermo Fisher, Waltham, MA, USA) at 4 °C and 50 rpm. The 8x histidine tag of the purified Phl p 5 was cleaved by overnight incubation with TEV protease (1:30 TEV protease:Phl p 5 molar ratio) and 2 mM dithiothreitol (DTT) at 4 °C and 50 rpm [
26]. Endotoxins were removed using a Pierce High-Capacity Endotoxin Removal Spin Column (Thermo Fisher) and endotoxin-free water (Thermo Fisher) according to the manufacturer´s instructions. Post-removal endotoxin levels were measured on an Endosafe Portable Test System (Charles River, Lyon, France). At times, cycles of endotoxin removal and subsequent measurement had to be repeated up to three times until levels dropped below 1 endotoxin unit (EU)/mL. Phl p 5 concentration was determined by a Micro Bicinchoninic Acid (MicroBCA, Thermo Scientific) protein assay (Thermo Scientific, Waltham, MA, USA) using a Nunc-Immuno flat-bottom 96-well plate (Thermo Fisher). Phl p 5 purity was confirmed by a 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) followed by overnight Coomassie Brilliant Blue (Abcam, Cambridge, UK) staining. The molecular mass of Phl p 5 was confirmed by matrix-assisted laser desorption ionization–time of flight (MALDI-TOF) mass spectrometry (Bruker, Billerica, MA, USA).
2.3. Generation of Lipidated Phl p 5
Plasmid generation and expression conditions were the same as described before for recombinant Phl p 5 used in EDC coupling. The protein sequence differed in only 3 amino acids, including a cysteine, which were added for lipidation at the C-terminus (
Supplementary Figure S1). The protein was purified from the soluble fraction by Ni-NTA purification using a gradient from 0–500 mM imidazole in PBS buffer at 4 °C. The fractions containing the protein of interest were pooled after SDS-PAGE analysis. TEV protease was added to the sample (1:20 ratio of TEV:Phl p 5-cysteine (Cys)), followed by dialysis overnight against PBS and 1 mM DTT at 4 °C using a dialysis tubing with a 6–8 kilodalton (kDa) MWco (Spectrum Chemical, New Brunswick, NJ, USA). The cleaved His tag was removed by preparative RP-HPLC at 60 °C on a C4 column using a gradient from 5–30% acetonitrile (can) in ddH
2O/0.1% TriFluoroacetic acid (TFA) in 5 min, followed by 30–70% ACN in ddH
2O/0.1%TFA in 30 min at a flowrate of 10mL/min. The fractions containing the purified Phl p 5-Cys were analyzed by electrospray ionization (ESI) mass spectroscopy (MS), pooled, and lyophilized.
For lipidation, the purified Phl p 5-Cys was dissolved in PBS buffer pH 7.2 containing 20 mM Tris(2-carboxyethyl)phosphine (TCEP) at ~2,6 mg/mL concentration. The crosslinker DSPE-PEG8-Mal (Broadpharm, San Diego, CA, USA) was dissolved in DMSO at ~2,8 mg/mL concentration using an ultrasonic bath. For a standard reaction, 9 mg Phl p 5-Cys in 3.5 mL buffer and 2.5 mg DSPE-PEG-Mal in 900 µL dimethylsulfoxide (DMSO) were combined, and the pH was adjusted to 7.2. The reaction was vigorously mixed for 1–2 hours at room temperature and monitored by liquid chromatography (LC)-MS. Upon complete conversion to the coupling product, the reaction mixture was diluted with 10 mL of a solution containing 6 M guanidine hydrochloride at pH 4.7. The resulting Phl p 5-DSPE, further referred to as lipidated Phl p 5, was filtered and purified by reversed-phase high-performance liquid chromatography (RP-HPLC) on a C4 column with a gradient from 5–30% ACN in ddH2O/0.1%TFA in 5 min followed by 30–95% ACN in ddH2O/0.1%TFA in 40 min at 60 °C and a flowrate of 10mL/min. The fractions containing the purified, lipidated Phl p 5 were analyzed by ESI MS, pooled, and lyophilized.
2.4. Recombinant Bet v 1
Recombinant Bet v 1.0101 was produced in BL21 (DE3) Escherichia coli cells and purified by hydrophobic interaction chromatography followed by anion exchange chromatography.
2.5. Circular Dichroism Analysis
The proteins’ secondary structure was determined on a spectropolarimeter (Jasco J-810, Tokyo, Japan) using a 1mm quartz cuvette. Spectra were recorded at room temperature at a scan speed of 50 nm/min and were corrected by subtracting the corresponding baseline spectrum, which was measured for water at identical conditions. Results are the means of three measurements and are expressed as molecular ellipticity.
2.6. Western Blots
Protein gels were blotted using the Trans-Blot SD semi-dry transfer apparatus (Bio-Rad Laboratories, Hercules, CA, USA). Membranes were washed three times with PBST/bovine serum albumin (BSA) for a total of 30 min at room temperature (RT). Membranes were incubated overnight with rabbit antiserum diluted 1:5000 at 4 °C with mild agitation. The next day, the solution was discarded, and the membranes were washed three times for a total of 30 min at RT and incubated with detection antibody for rabbit antisera diluted 1:5000 for 1 h at RT with mild agitation. Next, membranes were washed three times for a total of 30 min at RT, and 2 mL AP buffer was added for pH adjustment for 2 min at RT and mild agitation. Five milliliters nitro-blue tetrazolium and 5-bromo-4-chloro-3’-indolyphosphate (NBT/BCIP) substrate was added, followed by ddH2O to stop the color reaction.
2.7. Isolation of Splenocytes
Spleens were isolated from naïve BALB/c mice, and single-cell suspensions were prepared as described [
14]. Suspensions were red blood cell (RBC)-lysed using a custom-made ammonium–chloride–potassium (ACK) buffer that consisted of 24.82 g NH
4Cl, 3 g KHCO
3, and 0.111 g ethylenediaminetetraacetic acid (EDTA) in 500 mL of sterile water (final pH 7.4). To lyse, a freshly meshed spleen was centrifuged at 300 g for 3 min in a 50 mL falcon tube. The pellet was resuspended in 2 mL of 5% fetal calf serum (FCS)/RPMI. Briefly, 0.5 mL of ACK buffer was added to 2.5 mL sterile water, and the final 3 mL solution was added to the spleen for a total of 5 mL for exactly 45 s. The falcon tube was filled up to 50 mL with 10%FCS/RPMI and centrifuged at 250 g for 10 min. After an additional wash, the suspension was filtered through a 70 μm cell strainer, and splenocytes were counted. Blood was lysed in a 15 mL falcon tube in 10 mL of sterile water for 20 s followed by the addition of 1 mL HANK medium and centrifugation at 250 g for 10 min. After careful supernatant aspiration, the pellet was resuspended in 1 mL PBS, and leukocytes were counted.
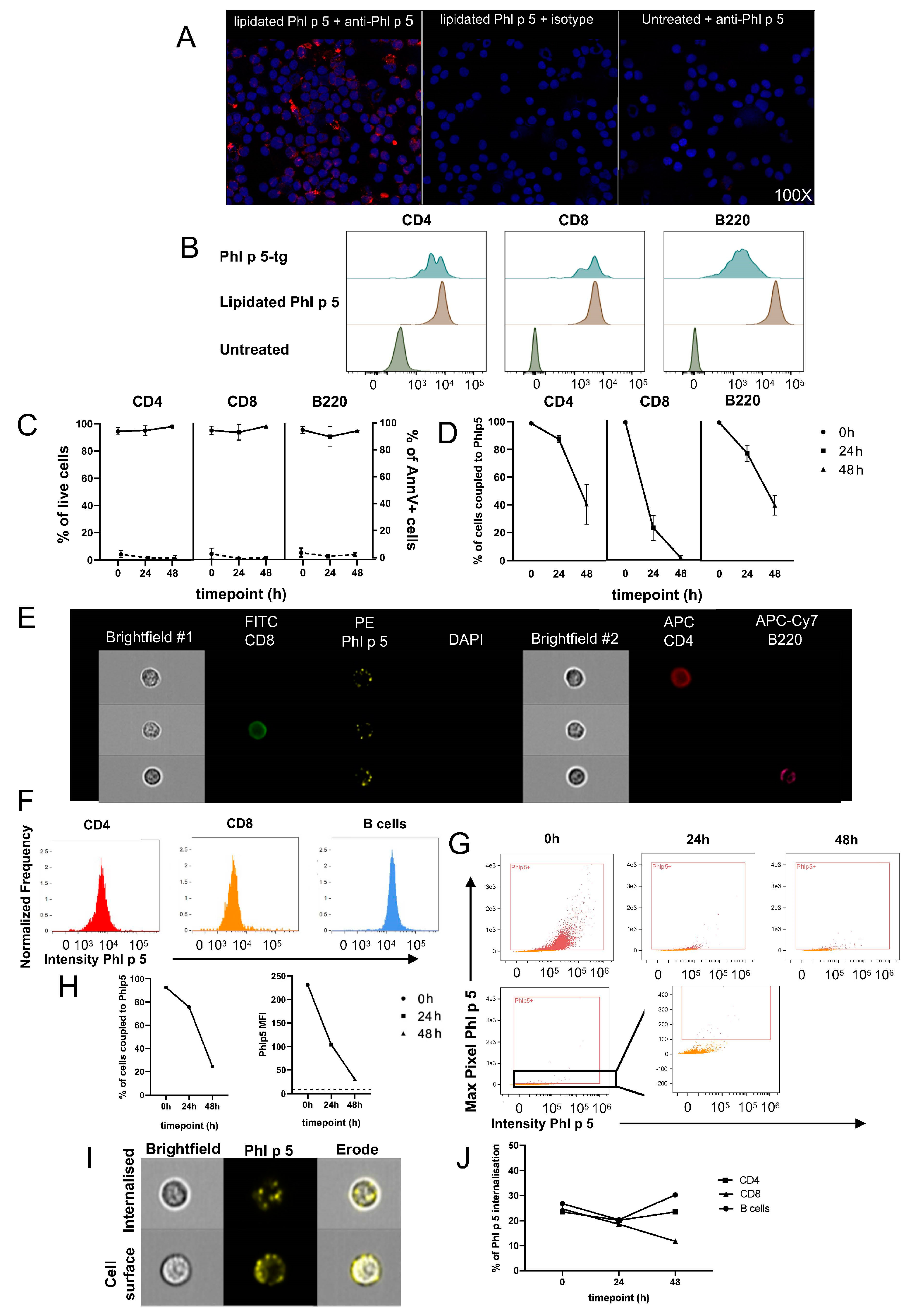
2.8. Lipidated Phl p 5 Coupling
Approximately 107 RBC-lysed BALB/c splenocytes were resuspended in 0.5 mL PBS supplemented with 100 μg purified, endotoxin-free lipidated Phl p 5. Cells were incubated at 37 °C for 1 h at 50 rpm, followed by thorough washing to remove unbound rPhl p 5.
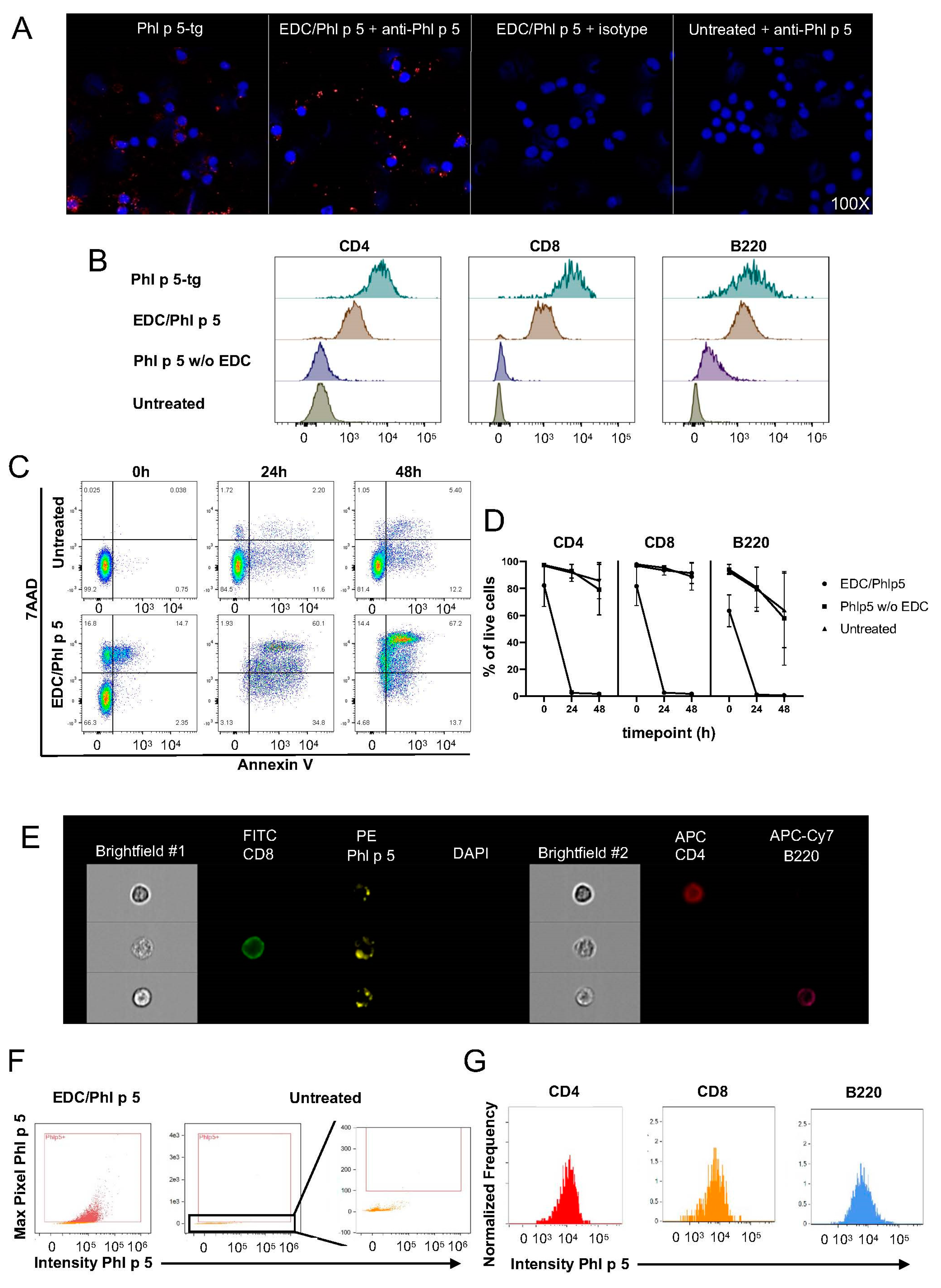
2.9. EDC Coupling
Approximately 107 RBC-lysed BALB/c splenocytes were resuspended in 0.5 mL PBS supplemented with 6.32 mg dissolved ECD and 100 μg purified, endotoxin-free rPhl p 5. Cells were incubated at 4 °C for 1 h at 50 rpm followed by thorough washing to remove unbound rPhl p 5.
2.10. Splenocyte Culture
Duplicates of 106 Phl p 5-coupled splenocytes were cultured in Greiner Cellstar 24-well plates (Sigma-Aldrich, Burlington, MA, USA) in RPMI 1640 medium (Biochrome, Cambridge, UK) supplemented with 10% FCS (Linaris, Frankfurt, Germany), PenStrep (100 U penicillin, 100 μg streptomycin/mL; Sigma), 10 mM HEPES (MP Biomedicals, Santa Ana, CA, USA), 1 mM sodium pyruvate (Sigma-Aldrich), 1x non-essential amino acids (Sigma-Aldrich), and 10 μM β-mercaptoethanol (Sigma-Aldrich). Cultures were incubated for 48 h at 37 °C and 5% CO2. The harvested cells were prepared for flow and image cytometry.
2.11. Flow Cytometry
Approximately 107 splenocytes were stained with CD4 (PE-Cy7), CD8 (APC), B220 (APC-H7), CD3 (V450), and rabbit polyclonal anti-Phl p 5 IgG (15) for 20 min at 4 °C in the dark. After washing, donkey anti-rabbit IgG (PE) was added for 15 min at 4 °C. Biotinylated Annexin V was diluted in 200 μL Annexin V buffer and incubated for 15 min at RT. The cells were washed thoroughly twice and then stained with streptavidin (FITC) in 50 μL PBS for 10 min at 4 °C. After 2 washes, the pellet was resuspended in 200 μL PBS containing 1 μL 7AAD (PercP-Cy5.5), vortexed, and analyzed immediately with a Fortessa flow cytometer (BD). Phl p 5-specific polyclonal antibodies were purified from rabbit serum raised against rPhl p 5a on a HiTrap Protein G HP antibody purification column (Merck, Rahway, NJ, USA) according to the manufacturer’s instructions.
2.12. Image Cytometry
Cells were stained with anti-Phl p 5 (rabbit polyclonal), anti-rabbit IgG (PE), anti-CD4 (APC), anti-CD8 (FITC), anti-B220 (APC-Cy7), and DAPI and run at a low rate in a concentration of 5 × 105/75 μL at 60X magnification using INSPIRE software version ISX on an ImageStream MkII image cytometer (Cytek Biosciences, Fremont, CA, USA). At least 104 focused singlets were acquired per sample. Doublets were eliminated upon gating. Single-color controls were run to set compensation. Compensation and data analysis were performed using IDEAS software version 6.2 (Cytek Biosciences).
2.13. Immunofluorescene Microscopy
Approximately 7 × 105 splenocytes were spun for 3 min at 72× g. Slides were fixed with 4% fresh neutral buffered formalin (NBF) at RT for 10 min. Slides were then rinsed in PBS and blocked with 10% goat serum in PBS at RT for 20 min. Briefly, 0.5 μg rabbit polyclonal anti-Phl p 5 was added to each slide, diluted in 100 μL 2%BSA/PBS, and incubated overnight at 4 °C in a humidified chamber. The next day, slides were washed 3 times in PBS, and 1:1000 goat anti-rabbit AF568 (Invitrogen, Waltham, MA, USA) was added in 2%BSA/PBS at RT for 1 h. Nuclei were counterstained with 1:1000 Draq5 at RT for 20 min and washed 3 times in PBS. Slides were rinsed with ddH20, mounted with ProLong Gold Antifade, and left overnight to dry. Slides were stored at 4 °C until visualization on a Nikon Ti Eclipse confocal microscope at 100× magnification.
2.14. Splenocyte Transfer
Splenocytes were isolated and prepared as described [
17]. Aliquots of 10
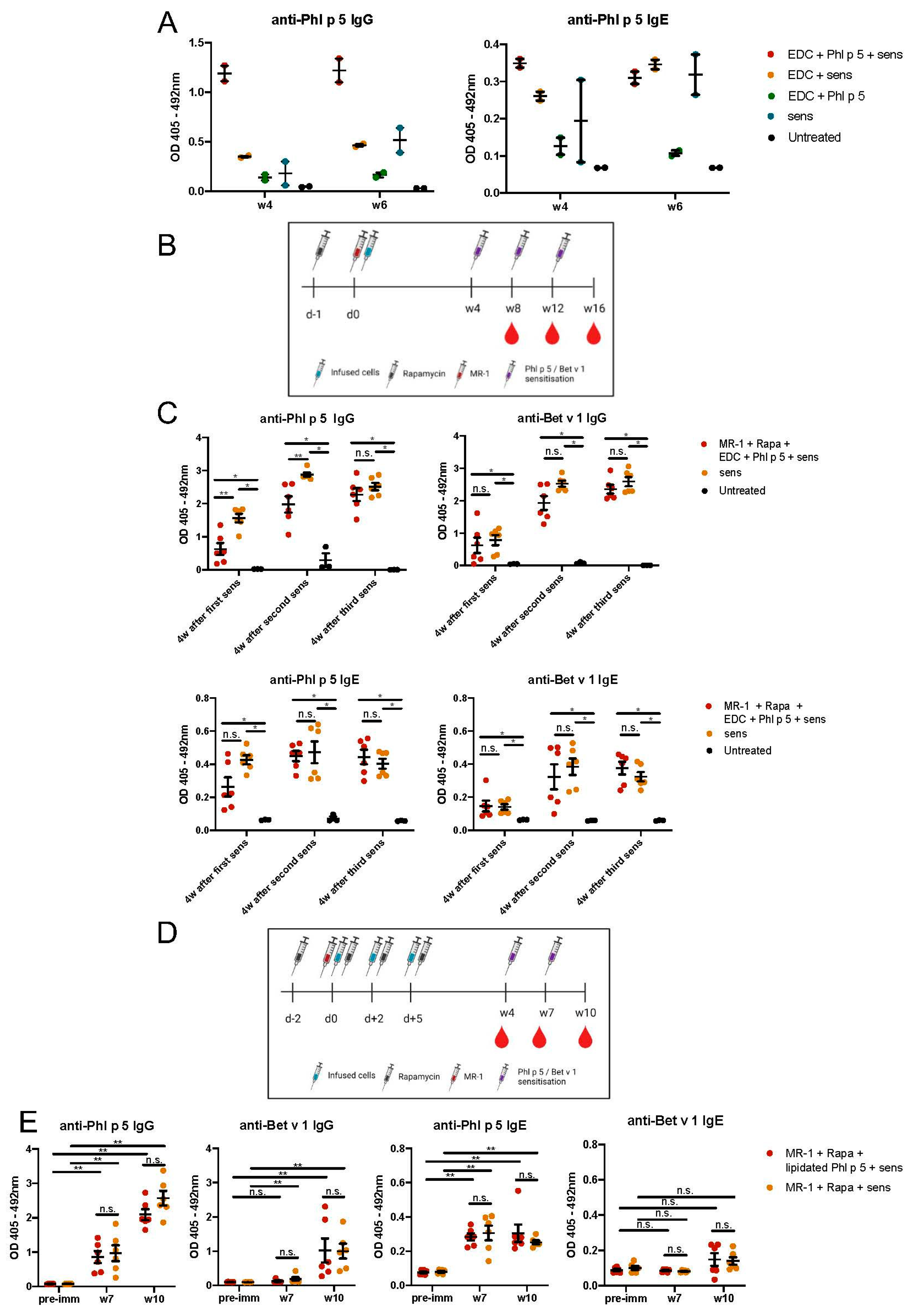
7 RBC-lysed and Phl p 5-coupled splenocytes were transplanted into the tail vein of naïve or preconditioned BALB/c recipients. Each preconditioned mouse was injected intraperitoneally (i.p.) with 50 mg/kg rapamycin (at d-1 for EDC experiments and d-2, 0, +2, +4 for lipidated experiments) and 1 mg hamster anti-mouse CD40L antibody (MR-1; Bio X Cell, Lebanon, NH, USA) at d0.
2.15. Immunisation of Mice
All groups of mice were immunized subcutaneously (sc) with 5 μg Phl p 5, as produced for EDC coupling, and 5 μg rBet v 1 adsorbed to aluminum hydroxide (Alu-Gel-S; Serva, Heidelberg, Germany) according to the manufacturer’s instructions. Mice were immunized and tail-bled at 4-week (w) intervals. Sera were stored at −20 °C until analysis.
2.16. Allergen-Specific ELISA
Allergen-specific ELISA was performed as described [
17]. Plates were coated with 5 μg of either rPhl p 5 or rBet v 1. Sera were diluted 1:20 for IgE and 1:500 for IgG isotypes. Bound antibodies were detected with monoclonal rat anti-mouse IgE or IgG1 (BD) diluted 1:1000 in 0.5%BSA/0.05%Tween20/PBS. Goat anti-rat HRP-labeled antiserum was added at a 1:2000 dilution in 0.5%BSA/0.05%Tween20/PBS at 37 °C for 30 min and then at 4 °C for 30 min. Plates were visualized with 100 μL ABTS (258 mg citric acid, 275,2 mg Na2HPO
4, 20 mg ABTS, 2 µL H
2O
2, 20 mL H
2O) per well at RT using a Spark plate reader at 490 nm (TECAN, Männedorf, Switzerland).
2.17. Statistical Analysis
The statistical analyses were performed using GraphPad Prism v8.2 (GraphPad Software, La Jolla, CA, USA). Differences between mouse groups were calculated using the two-sided Mann–Whitney U test. P-values below 0.05 were considered statistically significant. Due to the small sample size, no statistical evaluation was performed for the preliminary experiment shown in Figure 4A. Results were presented as dot plots, with lines denoting the mean and whiskers denoting the standard error of the mean (SEM). All in vivo results, with the exception of Figure 4A, depict pooled data obtained from two independent experiments.
4. Discussion
In this study, we aimed to bring a previously described cell therapy approach closer to eventual clinical translation through linking an allergen to the cell surface with safer methods than retroviral transduction. The approach is based on the prophylactic application of allergen-expressing immune cells and is associated with the establishment of molecular chimerism in recipients for several months, and the development of robust and long-lasting allergen-specific tolerance. In previous work, we could show that very low levels of chimerism (so-called microchimerism) are sufficient for tolerance induction [
15]. However, it is currently not known how long allergen expression needs to persist to achieve prevention from allergic sensitization. Here, we utilized two strategies for chemical linkage of a well-characterized and clinically important allergen, Phl p 5, to the cell surface of mouse splenocytes, including EDC-coupling and lipidated Phl p 5 modification with a lipophilic anchor. In relation to allergies, EDC has been used in two models of Th2 allergy responses: An ovalbumin-induced airway inflammation model and a whole peanut extract (WPE) food allergy model demonstrating significant antigen-specific serum IgE reduction [
22]. Our in vitro results from EDC-coupling are consistent with the published data since we observed apoptosis in antigen-coupled cells, which has been described as a prerequisite to the induction of antigen-specific tolerance [
24]. Unexpectedly and in contrast to reported protocols with other antigens, transplantation of autologous apoptotic EDC/Phl p 5 splenocytes did not prevent the Phl p 5-specific IgE and IgG response upon subsequent Phl p 5 sensitization, but indeed augmented the anti-Phl p 5 response. Consequently, we adapted the protocol by adding a short course of immunosuppression in accordance with our previously established protocols. However, except for a discrete initial reduction of Phl p 5-specific IgG and IgE responses, long-term tolerance could again not be induced. Distinct outcomes that were achieved in tolerance models using different allergens may highlight the role of the antigen itself for this approach. Phl p 5 specifically is known to be a highly immunogenic and potent allergen, eliciting strong immune responses in allergic patients. Moreover, the integrity of proteins might be differentially affected by the coupling procedure, influencing their in vivo efficacy. Furthermore, it has been suggested that Th1 responses are more successfully tolerated using EDC approaches [
28].
Whereas the EDC tolerization mechanism presumably involves anergy-inducing direct antigen presentation in the absence of costimulatory signals, chimerism-based tolerance approaches heavily rely on clonal deletion and regulatory mechanisms for tolerance induction [
29]. Adding a lipid anchor to a modified version of Phl p 5 enabled the straightforward chemical coupling of the allergen to the cell surface. The allergen was retained only for a limited time on the cell surface due to internalization, and we hypothesize that this period of time was not sufficient for tolerance induction. Internalized Phl p 5 is probably less potent in tolerization, as we observed in a previous study that cytoplasmic Phl p 5 led to less robust tolerance [
16]. We attempted to circumvent this issue by repeatedly injecting coupled cells. However, the duration of decoration did not seem to suffice even under repetitive infusions, suggesting that the decoration of donor cells with the allergen needs to be strong, stable, and (relatively) long lasting.
The present study demonstrates that chemical coupling of an allergen for cell therapy is feasible, in particular with the use of a lipid anchor. However, current strategies do not suffice for robust tolerance induction in vivo, as previously demonstrated for allergen-expressing immune cells. Further studies are needed to understand the mechanisms underlying successful tolerance induction in our cell therapy approach. It is currently not known whether central or peripheral tolerance is involved or if regulation by cytokines plays a role [
29]. Future studies will also need to focus on developing coupling strategies that lead to more stable immobilization of allergens on live immune cells.

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}



