Methods to Monitor and Quantify Autophagy in the Social Amoeba Dictyostelium discoideum


 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
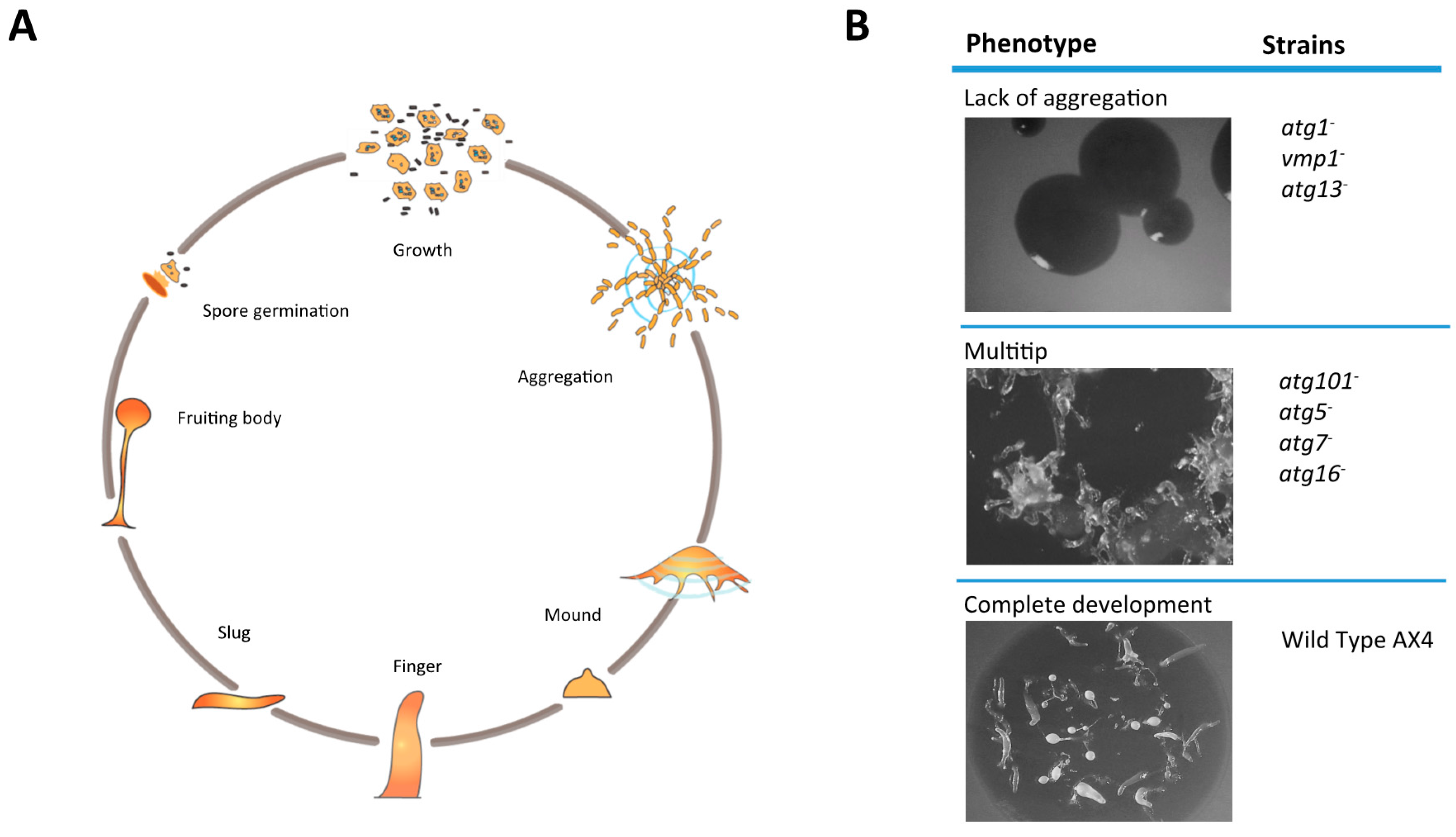
:1. Introducing Dictyostelium as a Model for Autophagy Research
2. Strategies to Induce Autophagy in Dictyostelium
3. Microscopy Techniques
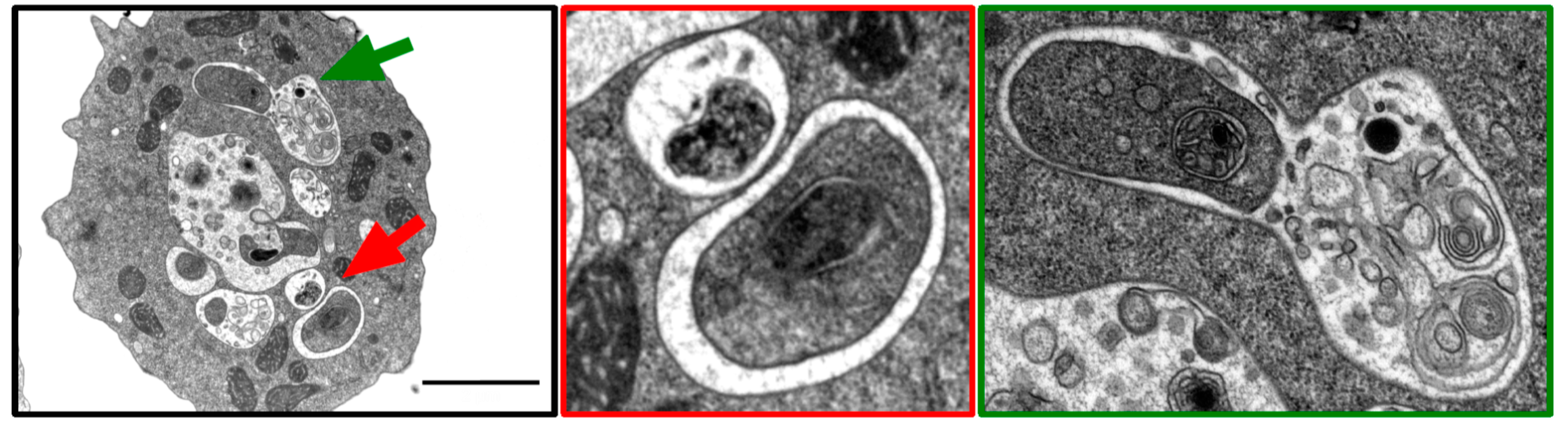
3.1. Transmission Electron Microscopy (TEM)
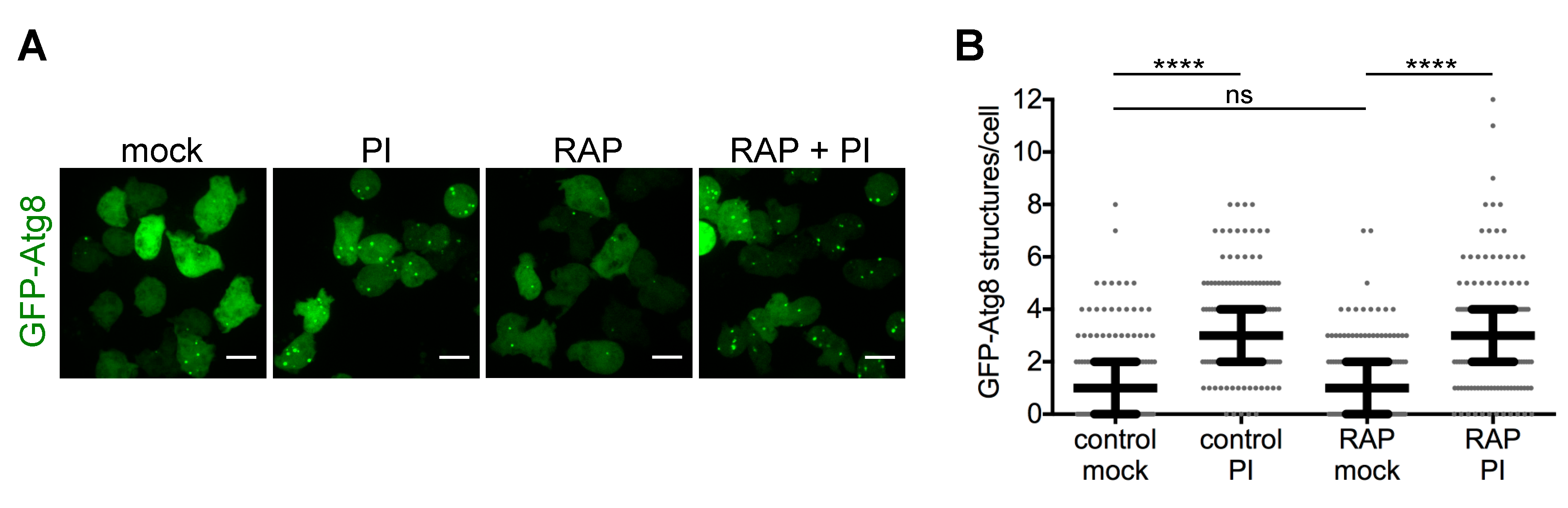
3.2. Fluorescence Microscopy
4. Measurement of Gene and Protein Expression Levels
5. Autophagic Flux Assays
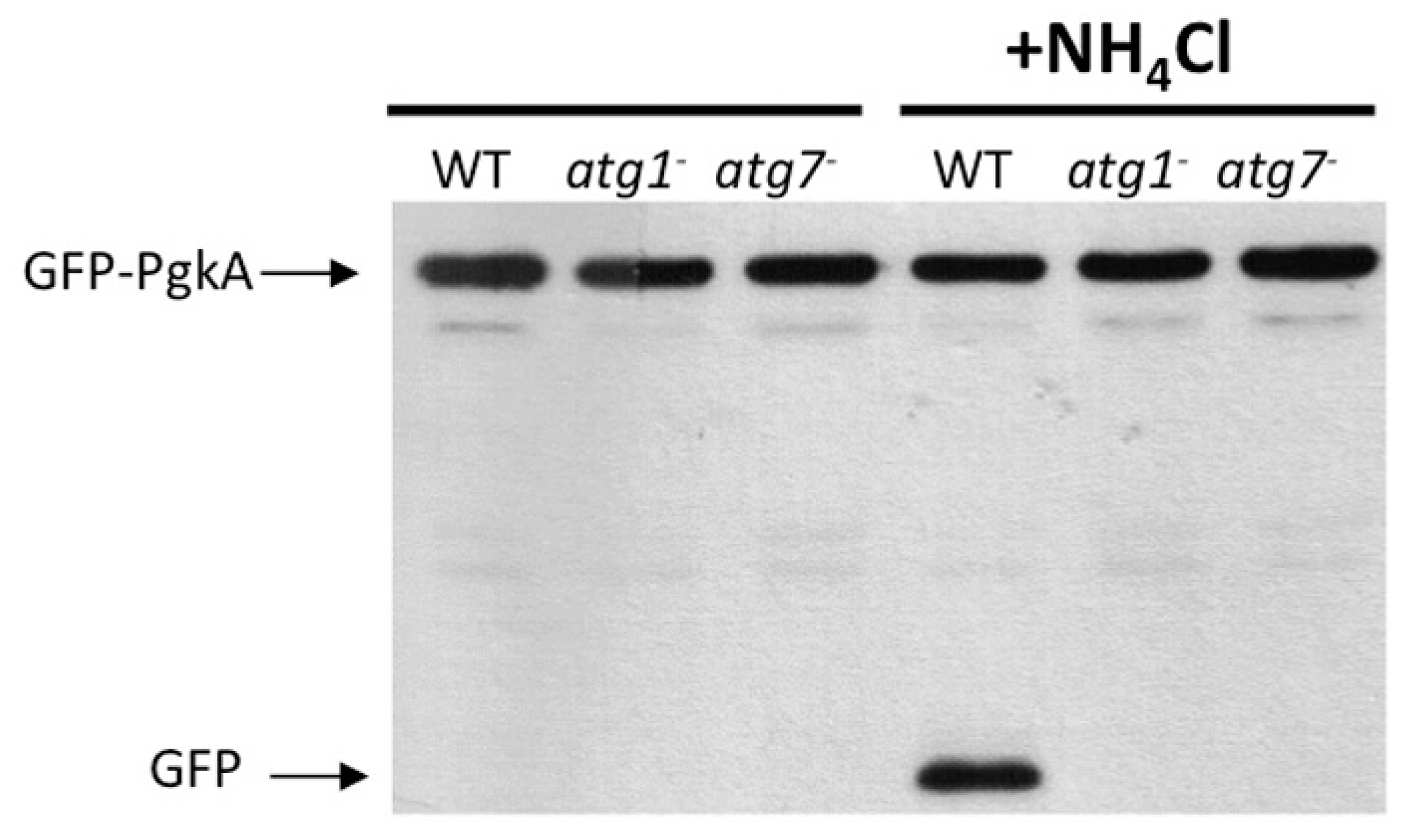
5.1. Protein Cleavage Assays
5.2. Quantification of Fluorescently-Tagged Proteins
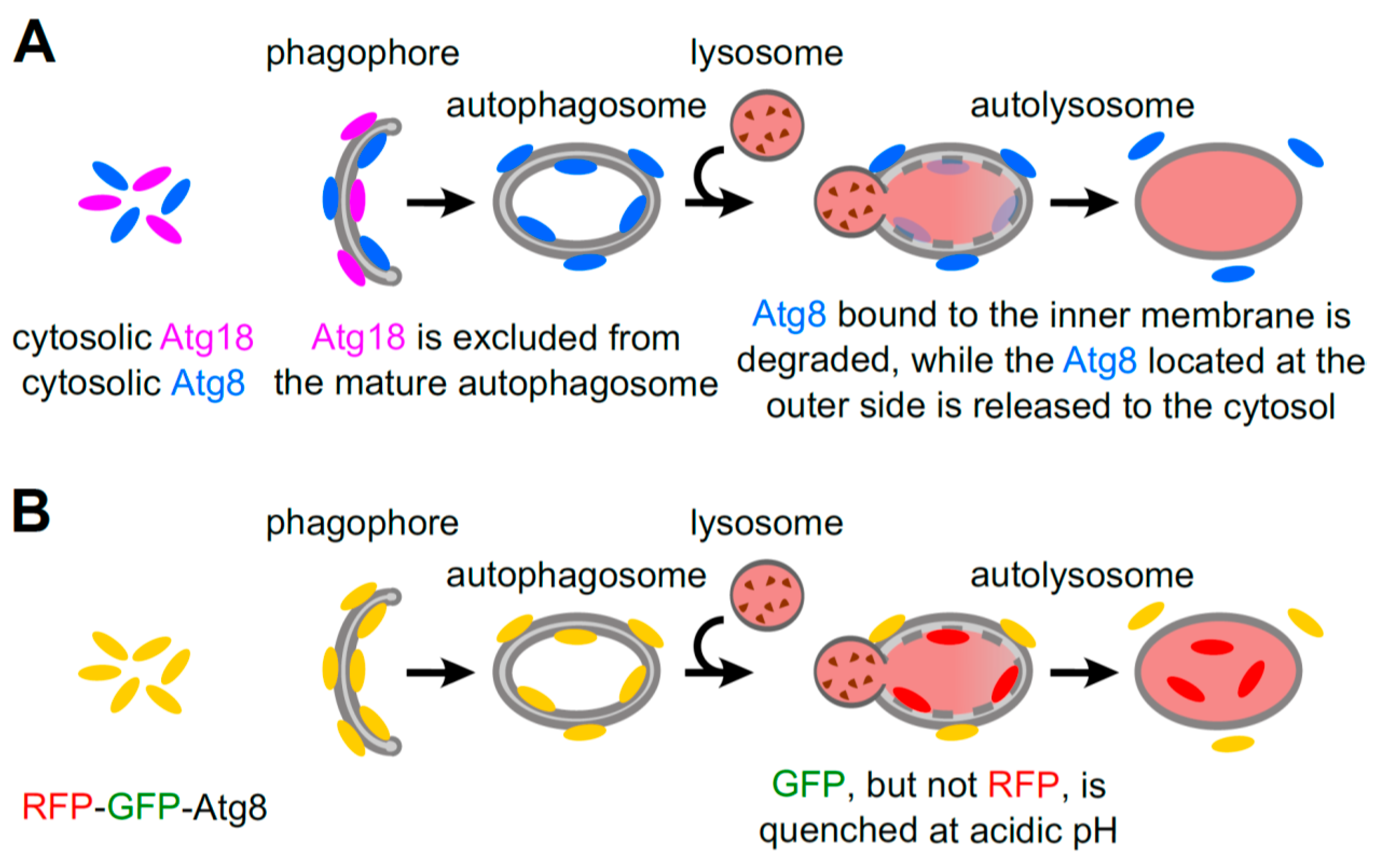
5.3. RFP-GFP-Atg8 Puncta Monitoring
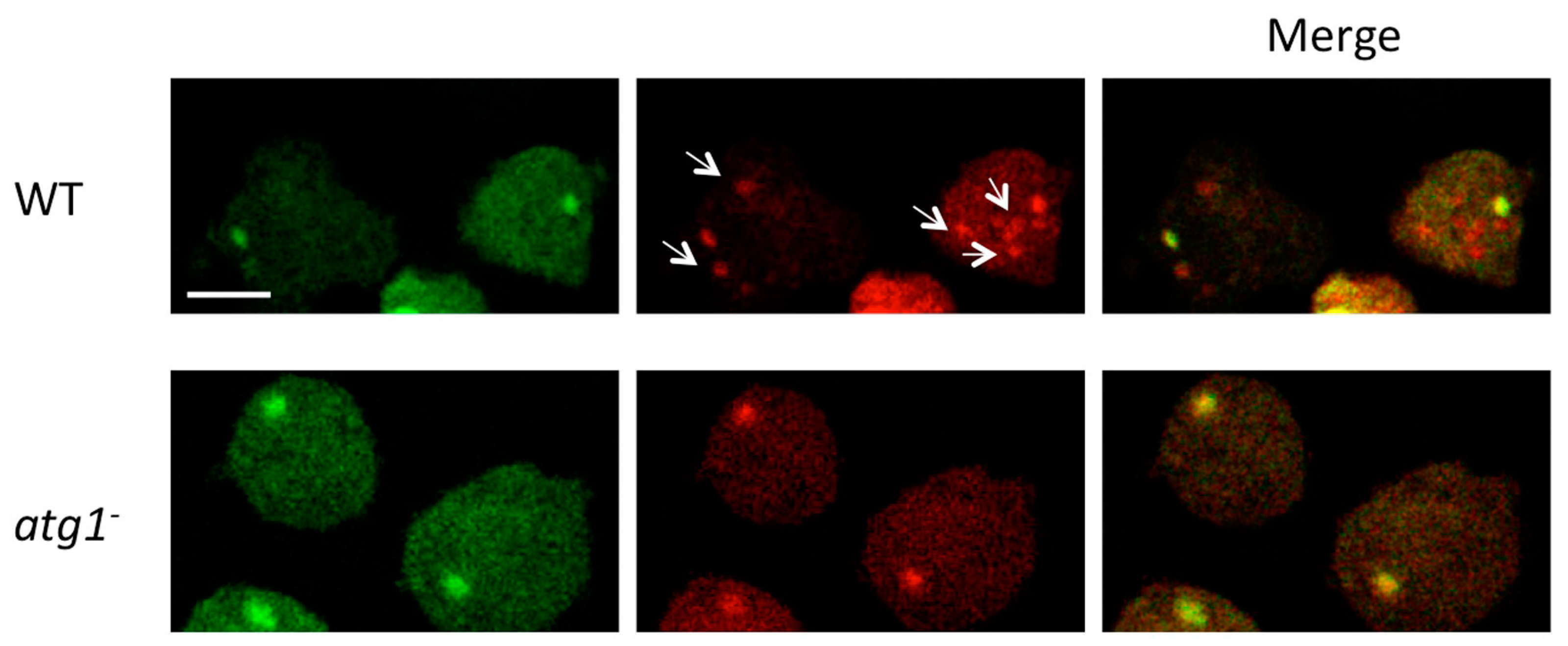
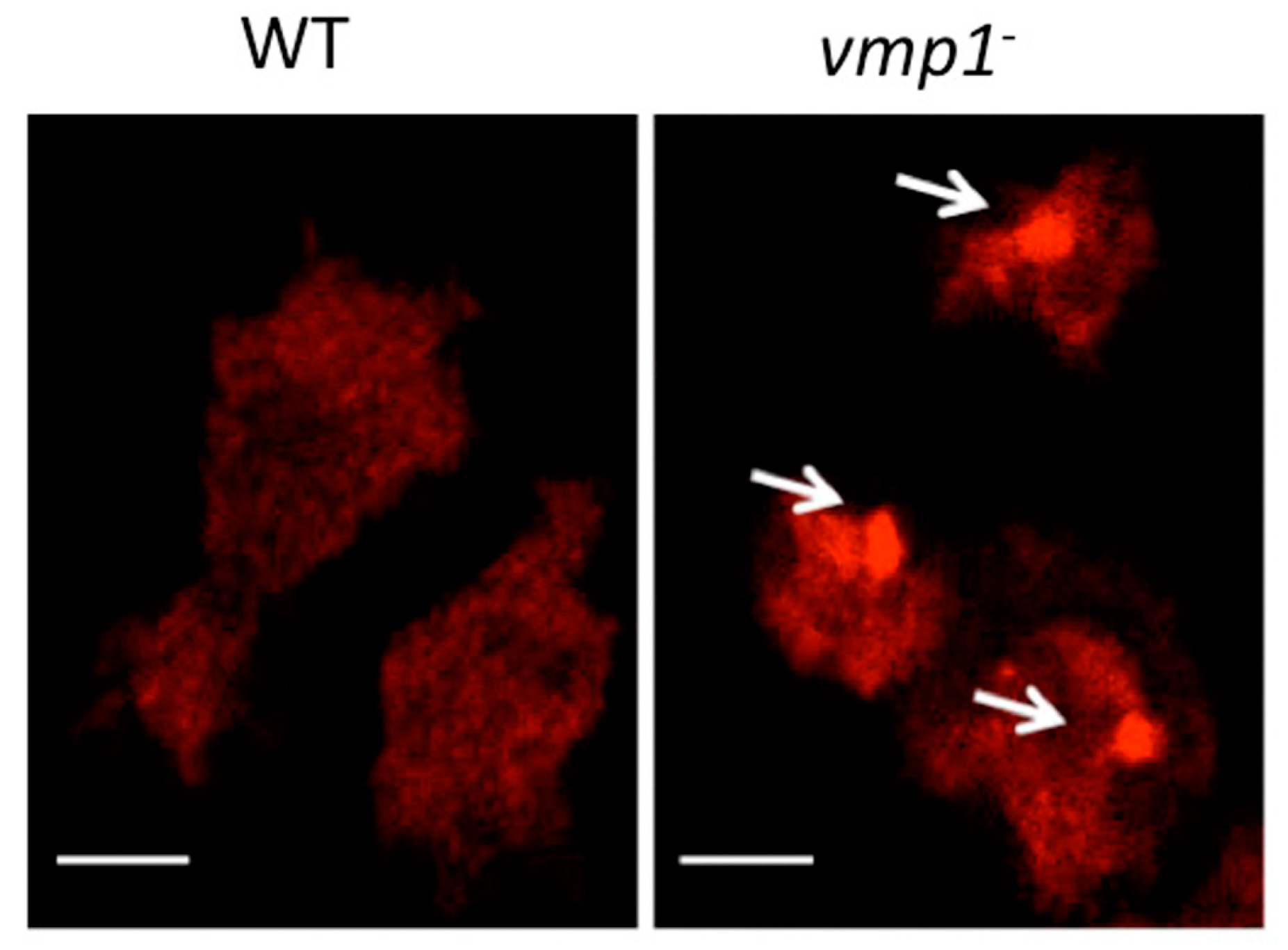
6. Determination of Ubiquitin Aggregates
7. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Onodera, J.; Ohsumi, Y. Autophagy is required for maintenance of amino acid levels and protein synthesis under nitrogen starvation. J. Biol. Chem. 2005, 280, 31582–31586. [Google Scholar] [CrossRef] [PubMed]
- King, J.S.; Veltman, D.M.; Insall, R.H. The induction of autophagy by mechanical stress. Autophagy 2011, 7, 1490–1499. [Google Scholar] [CrossRef] [PubMed]
- Yorimitsu, T.; Nair, U.; Yang, Z.; Klionsky, D.J. Endoplasmic reticulum stress triggers autophagy. J. Biol. Chem. 2006, 281, 30299–30304. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.; Celli, J. Avoidance and Subversion of Eukaryotic Homeostatic Autophagy Mechanisms by Bacterial Pathogens. J. Mol. Biol. 2016, 428, 3387–3398. [Google Scholar] [CrossRef] [PubMed]
- Lamb, C.A.; Yoshimori, T.; Tooze, S.A. The autophagosome: Origins unknown, biogenesis complex. Nat. Rev. Mol. Cell Biol. 2013, 14, 759–774. [Google Scholar] [CrossRef] [PubMed]
- Shibutani, S.T.; Yoshimori, T. A current perspective of autophagosome biogenesis. Cell Res. 2014, 24, 58–68. [Google Scholar] [CrossRef] [PubMed]
- De Duve, C. Ciba Foundation Symposium. In Lysosome; De Reuck, A., Cameron, M.P., Eds.; Little Brown and Company: Boston, MA, USA, 1963. [Google Scholar]
- Schneider, J.L.; Cuervo, A.M. Autophagy and human disease: Emerging themes. Curr. Opin. Genet. Dev. 2014, 26, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Klionsky, D.J. Autophagy wins the 2016 Nobel Prize in Physiology or Medicine: Breakthroughs in baker’s yeast fuel advances in biomedical research. Proc. Natl. Acad. Sci. USA 2017, 114, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, M.; Ohsumi, Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett. 1993, 333, 169–174. [Google Scholar] [CrossRef]
- Romeralo, M.; Escalante, R.; Baldauf, S.L. Evolution and diversity of dictyostelid social amoebae. Protist 2012, 163, 327–343. [Google Scholar] [CrossRef] [PubMed]
- Annesley, S.J.; Fisher, P.R. Dictyostelium discoideum—A model for many reasons. Mol. Cell. Biochem. 2009, 329, 73–91. [Google Scholar] [CrossRef] [PubMed]
- King, J.S. Autophagy across the eukaryotes: Is S. cerevisiae the odd one out? Autophagy 2012, 8, 1159–1162. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Braceras, S.; Mesquita, A.; Escalante, R. Dictyostelium discoideum as a model in biomedical research. In Dictyostelids. Evolution, Genomics and Cell Biology; Romeralo, M., Baldauf, S., Escalante, R., Eds.; Springer: Berlin/Heiderberg, Germany, 2013; pp. 1–34. [Google Scholar]
- Alexander, S.; Alexander, H. Lead genetic studies in Dictyostelium discoideum and translational studies in human cells demonstrate that sphingolipids are key regulators of sensitivity to cisplatin and other anticancer drugs. Semin. Cell Dev. Biol. 2011, 22, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Devreotes, P.N. Moving in the right direction: How eukaryotic cells migrate along chemical gradients. Semin. Cell Dev. Biol. 2011, 22, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Francione, L.M.; Annesley, S.J.; Carilla-Latorre, S.; Escalante, R.; Fisher, P.R. The Dictyostelium model for mitochondrial disease. Semin. Cell Dev. Biol. 2011, 22, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Ludtmann, M.H.; Boeckeler, K.; Williams, R.S. Molecular pharmacology in a simple model system: Implicating MAP kinase and phosphoinositide signalling in bipolar disorder. Semin. Cell Dev. Biol. 2011, 22, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Maniak, M. Dictyostelium as a model for human lysosomal and trafficking diseases. Semin. Cell Dev. Biol. 2011, 22, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Steinert, M. Pathogen-host interactions in Dictyostelium, Legionella, Mycobacterium and other pathogens. Semin. Cell Dev. Biol. 2011, 22, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Huber, R.J. Using the social amoeba Dictyostelium to study the functions of proteins linked to neuronal ceroid lipofuscinosis. J. Biomed. Sci. 2016, 23, 83. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Braceras, S.; Calvo, R.; Escalante, R. TipC and the chorea-acanthocytosis protein VPS13A regulate autophagy in Dictyostelium and human HeLa cells. Autophagy 2015, 11, 918–927. [Google Scholar] [CrossRef] [PubMed]
- McMains, V.C.; Myre, M.; Kreppel, L.; Kimmel, A.R. Dictyostelium possesses highly diverged presenilin/gamma-secretase that regulates growth and cell-fate specification and can accurately process human APP: A system for functional studies of the presenilin/gamma-secretase complex. Dis. Model Mech. 2010, 3, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Meyer, I.; Kuhnert, O.; Graf, R. Functional analyses of lissencephaly-related proteins in Dictyostelium. Semin. Cell Dev. Biol. 2011, 22, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Lima, W.C.; Lelong, E.; Cosson, P. What can Dictyostelium bring to the study of Pseudomonas infections? Semin. Cell Dev. Biol. 2011, 22, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Schaap, P. Evolutionary crossroads in developmental biology: Dictyostelium discoideum. Development 2011, 138, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhuchenko, O.; Kuspa, A.; Soldati, T. Social amoebae trap and kill bacteria by casting DNA nets. Nat. Commun. 2016, 7, 10938. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Soldati, T. Of Amoebae and Men: Extracellular DNA Traps as an Ancient Cell-Intrinsic Defense Mechanism. Front. Immunol. 2016, 7, 269. [Google Scholar] [CrossRef] [PubMed]
- Otto, G.P.; Wu, M.Y.; Kazgan, N.; Anderson, O.R.; Kessin, R.H. Macroautophagy is required for multicellular development of the social amoeba Dictyostelium discoideum. J. Biol. Chem. 2003, 278, 17636–17645. [Google Scholar] [CrossRef] [PubMed]
- Uchikawa, T.; Yamamoto, A.; Inouye, K. Origin and function of the stalk-cell vacuole in Dictyostelium. Dev. Biol. 2011, 352, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Duran, J.M.; Anjard, C.; Stefan, C.; Loomis, W.F.; Malhotra, V. Unconventional secretion of Acb1 is mediated by autophagosomes. J. Cell. Biol. 2010, 188, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Otto, G.P.; Wu, M.Y.; Kazgan, N.; Anderson, O.R.; Kessin, R.H. Dictyostelium macroautophagy mutants vary in the severity of their developmental defects. J. Biol. Chem. 2004, 279, 15621–15629. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, A.; Tabara, L.C.; Martinez-Costa, O.; Santos-Rodrigo, N.; Vincent, O.; Escalante, R. Dissecting the function of Atg1 complex in Dictyostelium autophagy reveals a connection with the pentose phosphate pathway enzyme transketolase. Open Biol. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, A.; Cardenal-Munoz, E.; Dominguez, E.; Munoz-Braceras, S.; Nunez-Corcuera, B.; Phillips, B.A.; Tabara, L.C.; Xiong, Q.; Coria, R.; Eichinger, L.; et al. Autophagy in Dictyostelium: Mechanisms, regulation and disease in a simple biomedical model. Autophagy 2016, 13, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Garrido, J.; Carilla-Latorre, S.; Kubohara, Y.; Santos-Rodrigo, N.; Mesquita, A.; Soldati, T.; Golstein, P.; Escalante, R. Autophagy in Dictyostelium: Genes and pathways, cell death and infection. Autophagy 2010, 6, 686–701. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Unal, C.; Matthias, J.; Steinert, M.; Eichinger, L. The phenotypes of ATG9, ATG16 and ATG9/16 knock-out mutants imply autophagy-dependent and–independent functions. Open Biol. 2015, 5, 150008. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, A.; Calvo-Garrido, J.; Carilla-Latorre, S.; Escalante, R. Monitoring autophagy in Dictyostelium. Methods Mol. Biol. 2013, 983, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Pflaum, K.; Gerdes, K.; Yovo, K.; Callahan, J.; Snyder, M.L. Lipopolysaccharide induction of autophagy is associated with enhanced bactericidal activity in Dictyostelium discoideum. Biochem. Biophys. Res. Commun. 2012, 422, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Cardenal-Munoz, E.; Arafah, S.; Lopez-Jimenez, A.T.; Kicka, S.; Falaise, A.; Bach, F.; Schaad, O.; King, J.S.; Hagedorn, M.; Soldati, T. Mycobacterium marinum antagonistically induces an autophagic response while repressing the autophagic flux in a TORC1-and ESX-1-dependent manner. PLoS Pathog. 2017, 13, e1006344. [Google Scholar] [CrossRef] [PubMed]
- Rosel, D.; Khurana, T.; Majithia, A.; Huang, X.; Bhandari, R.; Kimmel, A.R. TOR complex 2 (TORC2) in Dictyostelium suppresses phagocytic nutrient capture independently of TORC1-mediated nutrient sensing. J. Cell Sci. 2012, 125, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Swer, P.B.; Lohia, R.; Saran, S. Analysis of rapamycin induced autophagy in Dictyostelium discoideum. Indian J. Exp. Biol. 2014, 52, 295–304. [Google Scholar] [PubMed]
- Gao, M.; Yeh, P.Y.; Lu, Y.S.; Hsu, C.H.; Chen, K.F.; Lee, W.C.; Feng, W.C.; Chen, C.S.; Kuo, M.L.; Cheng, A.L. OSU-03012, a novel celecoxib derivative, induces reactive oxygen species-related autophagy in hepatocellular carcinoma. Cancer Res. 2008, 68, 9348–9357. [Google Scholar] [CrossRef] [PubMed]
- Park, M.A.; Yacoub, A.; Rahmani, M.; Zhang, G.; Hart, L.; Hagan, M.P.; Calderwood, S.K.; Sherman, M.Y.; Koumenis, C.; Spiegel, S.; et al. OSU-03012 stimulates PKR-like endoplasmic reticulum-dependent increases in 70-kDa heat shock protein expression, attenuating its lethal actions in transformed cells. Mol. Pharmacol. 2008, 73, 1168–1184. [Google Scholar] [CrossRef] [PubMed]
- de Chastellier, C.; Ryter, A. Changes on the cell surface and of the digestive apparatus of Dictyostelium discoideum during the starvation period triggering aggregation. J. Cell Biol. 1977, 75, 218–236. [Google Scholar] [CrossRef] [PubMed]
- Barisch, C.; Paschke, P.; Hagedorn, M.; Maniak, M.; Soldati, T. Lipid droplet dynamics at early stages of Mycobacterium marinum infection in Dictyostelium. Cell. Microbiol. 2015, 17, 1332–1349. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Garrido, J.; Carilla-Latorre, S.; Lazaro-Dieguez, F.; Egea, G.; Escalante, R. Vacuole membrane protein 1 is an endoplasmic reticulum protein required for organelle biogenesis, protein secretion, and development. Mol. Biol. Cell 2008, 19, 3442–3453. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Abdelmohsen, K.; Abe, A.; Abedin, M.J.; Abeliovich, H.; Acevedo Arozena, A.; Adachi, H.; Adams, C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12, 1–222. [Google Scholar] [CrossRef] [PubMed]
- Matthias, J.; Messling, S.; Eichinger, L. The two Dictyostelium autophagy eight proteins, ATG8a and ATG8b, associate with the autophagosome in succession. Eur. J. Cell Biol. 2016, 95, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Messling, S.; Matthias, J.; Xiong, Q.; Fischer, S.; Eichinger, L. The two Dictyostelium discoideum autophagy 8 proteins have distinct autophagic functions. Eur. J. Cell Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Proikas-Cezanne, T.; Takacs, Z.; Donnes, P.; Kohlbacher, O. WIPI proteins: Essential PtdIns3P effectors at the nascent autophagosome. J. Cell Sci. 2015, 128, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Garrido, J.; Escalante, R. Autophagy dysfunction and ubiquitin-positive protein aggregates in Dictyostelium cells lacking Vmp1. Autophagy 2010, 6, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Gerstenmaier, L.; Pilla, R.; Herrmann, L.; Herrmann, H.; Prado, M.; Villafano, G.J.; Kolonko, M.; Reimer, R.; Soldati, T.; King, J.S.; et al. The autophagic machinery ensures nonlytic transmission of mycobacteria. Proc. Natl. Acad. Sci. USA 2015, 112, E687–E692. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Garrido, J.; Carilla-Latorre, S.; Mesquita, A.; Escalante, R. A proteolytic cleavage assay to monitor autophagy in Dictyostelium discoideum. Autophagy 2011, 7, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, A.; Lelong, E.; Cosson, P. A measure of endosomal pH by flow cytometry in Dictyostelium. BMC Res. Notes 2009, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, P.P.; Fairn, G.D.; Grinstein, S.; Johnson, D.E. Cresyl violet: A superior fluorescent lysosomal marker. Traffic 2016, 17, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Carilla-Latorre, S.; Annesley, S.J.; Munoz-Braceras, S.; Fisher, P.R.; Escalante, R. Ndufaf5 deficiency in the Dictyostelium model: New roles in autophagy and development. Mol. Biol. Cell 2013, 24, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Lohia, R.; Jain, P.; Jain, M.; Burma, P.K.; Shrivastava, A.; Saran, S. Dictyostelium discoideum Sir2D modulates cell-type specific gene expression and is involved in autophagy. Int. J. Dev. Biol. 2017, 61, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Korolchuk, V.I.; Mansilla, A.; Menzies, F.M.; Rubinsztein, D.C. Autophagy inhibition compromises degradation of ubiquitin-proteasome pathway substrates. Mol. Cell 2009, 33, 517–527. [Google Scholar] [CrossRef] [PubMed]







© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Martín, E.; Cardenal-Muñoz, E.; King, J.S.; Soldati, T.; Coria, R.; Escalante, R. Methods to Monitor and Quantify Autophagy in the Social Amoeba Dictyostelium discoideum. Cells 2017, 6, 18. https://doi.org/10.3390/cells6030018
Domínguez-Martín E, Cardenal-Muñoz E, King JS, Soldati T, Coria R, Escalante R. Methods to Monitor and Quantify Autophagy in the Social Amoeba Dictyostelium discoideum. Cells. 2017; 6(3):18. https://doi.org/10.3390/cells6030018
Chicago/Turabian StyleDomínguez-Martín, Eunice, Elena Cardenal-Muñoz, Jason S. King, Thierry Soldati, Roberto Coria, and Ricardo Escalante. 2017. "Methods to Monitor and Quantify Autophagy in the Social Amoeba Dictyostelium discoideum" Cells 6, no. 3: 18. https://doi.org/10.3390/cells6030018
APA StyleDomínguez-Martín, E., Cardenal-Muñoz, E., King, J. S., Soldati, T., Coria, R., & Escalante, R. (2017). Methods to Monitor and Quantify Autophagy in the Social Amoeba Dictyostelium discoideum. Cells, 6(3), 18. https://doi.org/10.3390/cells6030018