Use of Human Neurons Derived via Cellular Reprogramming Methods to Study Host-Parasite Interactions of Toxoplasma gondii in Neurons

Abstract
:1. Introduction
2. Generation of iPSCs and Development of the Field of Cellular Reprogramming
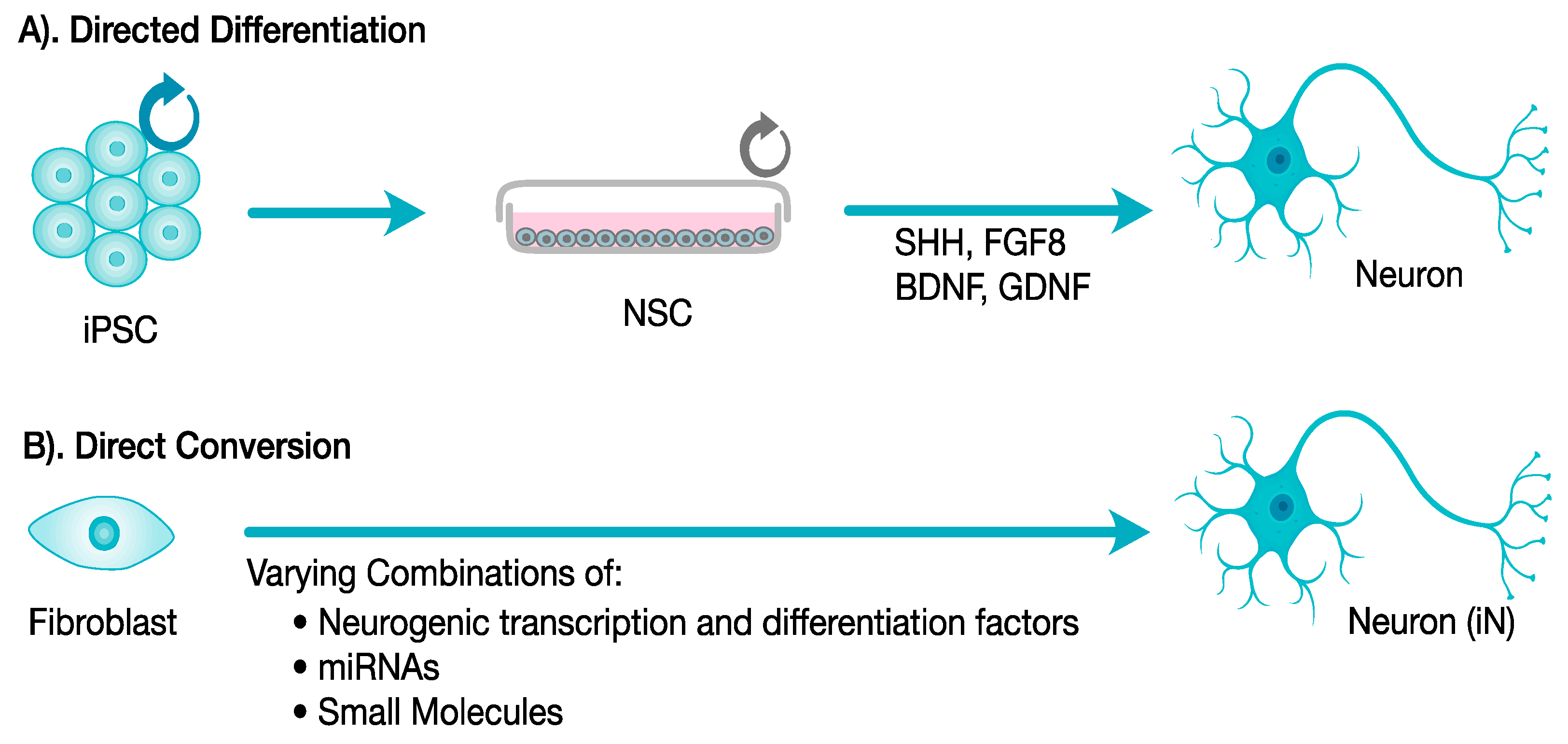
2.1. Differentiation Protocols to Derive Neurons and Neuronal Subtypes
2.2. Neural Cellular Reprogramming: Existing Limitations and Implications to Disease Modeling
3. Use of Reprogrammed Human Neurons to Study Cerebral Toxoplasmosis
3.1. Use of Reprogrammed Human Neuronal Models to Study the Host/Parasite Relationships in Neurons and to Address Outstanding Questions about Cerebral Toxoplasmosis
3.2. Advantages of Human Neuronal Models Using Reprogrammed Neurons and Applications to the Study of Cerebral Toxoplasmosis
3.2.1. Generation of Relatively Unlimited Supply of Pure, Mature Human Neurons
3.2.2. Generation of Human Neurons with Functional Characteristics
3.2.3. Disease-Specific Neurons Can Be Created
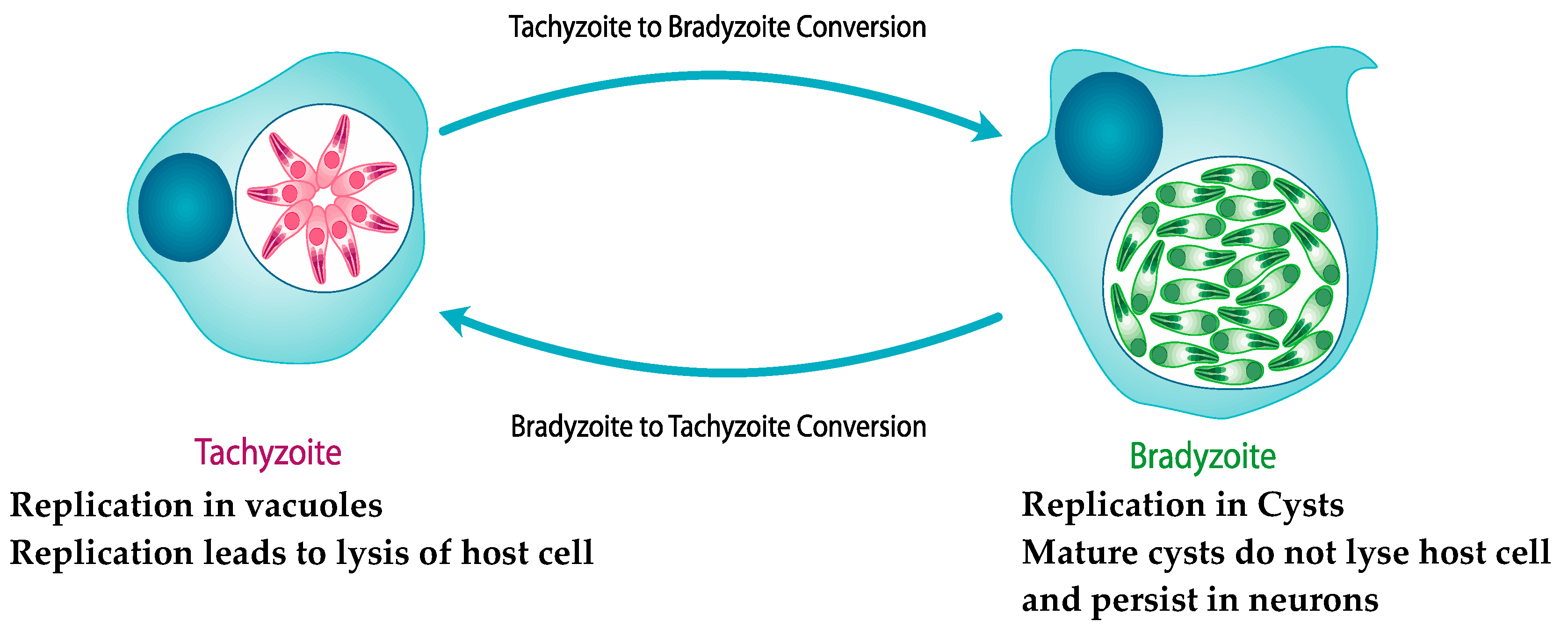
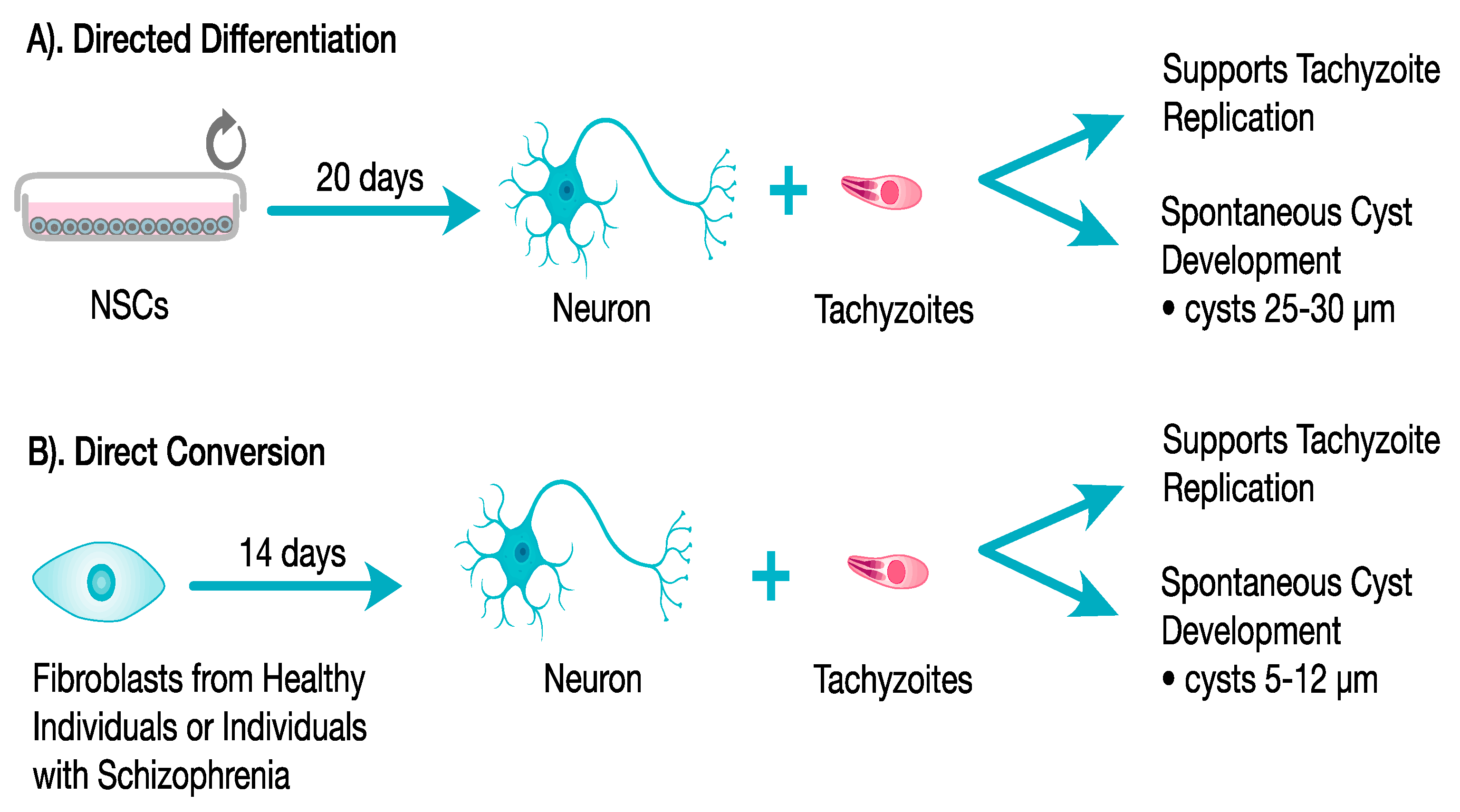
3.2.4. Provides a Model for the Study of Bradyzoite Replication and Cyst Development
3.2.5. Neuronal-Astrocyte Co-Cultures
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef]
- Ferguson, D.J.; Hutchison, W.M. An ultrastructural study of the early development and tissue cyst formation of Toxoplasma gondii in the brains of mice. Parasitol. Res. 1987, 73, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Bradyzoite-induced murine toxoplasmosis: Stage conversion, pathogenesis, and tissue cyst formation in mice fed bradyzoites of different strains of Toxoplasma gondii. J. Eukaryot. Microbiol. 1997, 44, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Okusaga, O.; Langenberg, P.; Sleemi, A.; Vaswani, D.; Giegling, I.; Hartmann, A.M.; Konte, B.; Friedl, M.; Groer, M.W.; Yolken, R.H.; et al. Toxoplasma gondii antibody titers and history of suicide attempts in patients with schizophrenia. Schizophr. Res. 2011, 133, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Groer, M.W.; Yolken, R.H.; Xiao, J.C.; Beckstead, J.W.; Fuchs, D.; Mohapatra, S.S.; Seyfang, A.; Postolache, T.T. Prenatal depression and anxiety in Toxoplasma gondii-positive women. Am. J. Obstet. Gynecol. 2011, 204, 433.e1–433.e7. [Google Scholar] [CrossRef] [PubMed]
- Sutterland, A.L.; Fond, G.; Kuin, A.; Koeter, M.W.; Lutter, R.; van Gool, T.; Yolken, R.; Szoke, A.; Leboyer, M.; de Haan, L. Beyond the association. Toxoplasma gondii in schizophrenia, bipolar disorder, and addiction: Systematic review and meta-analysis. Acta Psychiatr. Scand. 2015, 132, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Torrey, E.F.; Bartko, J.J.; Yolken, R.H. Toxoplasma gondii and other risk factors for schizophrenia: An update. Schizophr. Bull. 2012, 38, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.G.; Mortensen, P.B.; Norgaard-Pedersen, B.; Postolache, T.T. Toxoplasma gondii infection and self-directed violence in mothers. Arch. Gen. Psychiatry 2012, 69, 1123–1130. [Google Scholar] [CrossRef] [PubMed]
- Yagmur, F.; Yazar, S.; Temel, H.O.; Cavusoglu, M. May Toxoplasma gondii increase suicide attempt-preliminary results in turkish subjects? Forensic. Sci. Int. 2010, 199, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.G.; Stevens, H.; Pedersen, C.B.; Norgaard-Pedersen, B.; Mortensen, P.B. Toxoplasma infection and later development of schizophrenia in mothers. Am. J. Psychiatry 2011, 168, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Prandota, J. Possible link between Toxoplasma gondii and the anosmia associated with neurodegenerative diseases. Am. J. Alzheimers Dis. Other Dement. 2014, 29, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Miman, O.; Mutlu, E.A.; Ozcan, O.; Atambay, M.; Karlidag, R.; Unal, S. Is there any role of Toxoplasma gondii in the etiology of obsessive-compulsive disorder? Psychiatry Res. 2010, 177, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Dickerson, F.; Stallings, C.; Origoni, A.; Katsafanas, E.; Schweinfurth, L.; Savage, C.; Khushalani, S.; Yolken, R. Antibodies to Toxoplasma gondii and cognitive functioning in schizophrenia, bipolar disorder, and nonpsychiatric controls. J. Nerv. Ment. Dis. 2014, 202, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.S., Jr. Association of increased prenatal estrogen with risk factors for schizophrenia. Schizophr. Bull. 2011, 37, 946–949. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.; Patterson, P.H. Maternal infection and schizophrenia: Implications for prevention. Schizophr. Bull. 2011, 37, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Yolken, R.H.; Dickerson, F.B.; Fuller Torrey, E. Toxoplasma and schizophrenia. Parasite Immunol. 2009, 31, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Kusbeci, O.Y.; Miman, O.; Yaman, M.; Aktepe, O.C.; Yazar, S. Could Toxoplasma gondii have any role in alzheimer disease? Alzheimer Dis. Assoc. Disord. 2011, 25, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Miman, O.; Kusbeci, O.Y.; Aktepe, O.C.; Cetinkaya, Z. The probable relation between Toxoplasma gondii and parkinson’s disease. Neurosci. Lett. 2010, 475, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S. Exposure to prenatal infection and risk of schizophrenia. Front. Psychiatry 2011, 2, 63. [Google Scholar] [CrossRef] [PubMed]
- Beste, C.; Getzmann, S.; Gajewski, P.D.; Golka, K.; Falkenstein, M. Latent Toxoplasma gondii infection leads to deficits in goal-directed behavior in healthy elderly. Neurobiol. Aging 2014, 35, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Yazar, S.; Arman, F.; Yalcin, S.; Demirtas, F.; Yaman, O.; Sahin, I. Investigation of probable relationship between Toxoplasma gondii and cryptogenic epilepsy. Seizure 2003, 12, 107–109. [Google Scholar] [CrossRef]
- Palmer, B.S. Meta-analysis of three case controlled studies and an ecological study into the link between cryptogenic epilepsy and chronic toxoplasmosis infection. Seizure 2007, 16, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Prandota, J. Autism spectrum disorders may be due to cerebral toxoplasmosis associated with chronic neuroinflammation causing persistent hypercytokinemia that resulted in an increased lipid peroxidation, oxidative stress, and depressed metabolism of endogenous and exogenous substances. Res. Autism Spect Dis. 2010, 4, 119–155. [Google Scholar]
- Ngoungou, E.B.; Bhalla, D.; Nzoghe, A.; Darde, M.L.; Preux, P.M. Toxoplasmosis and epilepsy--systematic review and meta analysis. PLoS Negl. Trop. Dis. 2015, 9, e0003525. [Google Scholar] [CrossRef] [PubMed]
- Fabiani, S.; Pinto, B.; Bonuccelli, U.; Bruschi, F. Neurobiological studies on the relationship between toxoplasmosis and neuropsychiatric diseases. J. Neurol. Sci. 2015, 351, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Mahami-Oskouei, M.; Hamidi, F.; Talebi, M.; Farhoudi, M.; Taheraghdam, A.A.; Kazemi, T.; Sadeghi-Bazargani, H.; Fallah, E. Toxoplasmosis and alzheimer: Can Toxoplasma gondii really be introduced as a risk factor in etiology of alzheimer? Parasitol. Res. 2016, 115, 3169–3174. [Google Scholar] [CrossRef] [PubMed]
- Haroon, F.; Handel, U.; Angenstein, F.; Goldschmidt, J.; Kreutzmann, P.; Lison, H.; Fischer, K.D.; Scheich, H.; Wetzel, W.; Schluter, D.; et al. Toxoplasma gondii actively inhibits neuronal function in chronically infected mice. PLoS ONE 2012, 7, e35516. [Google Scholar] [CrossRef] [PubMed]
- Prandovszky, E.; Gaskell, E.; Martin, H.; Dubey, J.P.; Webster, J.P.; McConkey, G.A. The neurotropic parasite Toxoplasma gondii increases dopamine metabolism. PLoS ONE 2011, 6, e23866. [Google Scholar] [CrossRef] [PubMed]
- Parlog, A.; Harsan, L.A.; Zagrebelsky, M.; Weller, M.; von Elverfeldt, D.; Mawrin, C.; Korte, M.; Dunay, I.R. Chronic murine toxoplasmosis is defined by subtle changes in neuronal connectivity. Dis. Model. Mech. 2014, 7, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Koshy, A.A.; Dietrich, H.K.; Christian, D.A.; Melehani, J.H.; Shastri, A.J.; Hunter, C.A.; Boothroyd, J.C. Toxoplasma co-opts host cells it does not invade. PLoS Pathog. 2012, 8, e1002825. [Google Scholar] [CrossRef] [PubMed]
- Mammari, N.; Vignoles, P.; Halabi, M.A.; Darde, M.L.; Courtioux, B. In Vitro infection of human nervous cells by two strains of Toxoplasma gondii: A kinetic analysis of immune mediators and parasite multiplication. PLoS ONE 2014, 9, e98491. [Google Scholar] [CrossRef] [PubMed]
- Mammari, N.; Vignoles, P.; Halabi, M.A.; Darde, M.L.; Courtioux, B. Interferon gamma effect on immune mediator production in human nerve cells infected by two strains of Toxoplasma gondii. Parasite 2015, 22, 39. [Google Scholar] [CrossRef] [PubMed]
- Sahm, M.; Fischer, H.G.; Gross, U.; Reiter-Owona, I.; Seitz, H.M. Cyst formation by Toxoplasma gondii in vivo and in brain-cell culture: A comparative morphology and immunocytochemistry study. Parasitol. Res. 1997, 83, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Creuzet, C.; Robert, F.; Roisin, M.P.; Van Tan, H.; Benes, C.; Dupouy-Camet, J.; Fagard, R. Neurons in primary culture are less efficiently infected by Toxoplasma gondii than glial cells. Parasitol. Res. 1998, 84, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Fagard, R.; Van Tan, H.; Creuzet, C.; Pelloux, H. Differential development of Toxoplasma gondii in neural cells. Parasitol. Today 1999, 15, 504–507. [Google Scholar] [CrossRef]
- Halonen, S.K.; Lyman, W.D.; Chiu, F.C. Growth and development of Toxoplasma gondii in human neurons and astrocytes. J. Neuropathol. Exp. Neurol. 1996, 55, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, Y.; Okano, H. Modeling human neurological disorders with induced pluripotent stem cells. J. Neurochem. 2014, 129, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Revilla, A.; Gonzalez, C.; Iriondo, A.; Fernandez, B.; Prieto, C.; Marin, C.; Liste, I. Current advances in the generation of human ips cells: Implications in cell-based regenerative medicine. J. Tissue Eng. Regen. Med. 2016, 10, 893–907. [Google Scholar] [CrossRef] [PubMed]
- Ichida, J.K.; Kiskinis, E. Probing disorders of the nervous system using reprogramming approaches. EMBO J. 2015, 34, 1456–1477. [Google Scholar] [CrossRef] [PubMed]
- Russo, F.B.; Cugola, F.R.; Fernandes, I.R.; Pignatari, G.C.; Beltrao-Braga, P.C. Induced pluripotent stem cells for modeling neurological disorders. World J. Transpl. 2015, 5, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Beltrao-Braga, P.C.; Pignatari, G.C.; Russo, F.B.; Fernandes, I.R.; Muotri, A.R. In-a-dish: Induced pluripotent stem cells as a novel model for human diseases. Cytometry A 2013, 83, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.Z.; Zhang, R.X. Neurodegenerative diseases in a dish: The promise of ipsc technology in disease modeling and therapeutic discovery. Neurol. Sci. 2015, 36, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Klotz, C.; Aebischer, T.; Seeber, F. Stem cell-derived cell cultures and organoids for protozoan parasite propagation and studying host-parasite interaction. Int. J. Med. Microbiol. 2012, 302, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Okita, K.; Nakagawa, M.; Yamanaka, S. Induction of pluripotent stem cells from fibroblast cultures. Nat. Protoc. 2007, 2, 3081–3089. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nityanandam, A.; Baldwin, K.K. Advances in reprogramming-based study of neurologic disorders. Stem Cells Dev. 2015, 24, 1265–1283. [Google Scholar] [CrossRef] [PubMed]
- An, N.; Xu, H.; Gao, W.Q.; Yang, H. Direct conversion of somatic cells into induced neurons. Mol. Neurobiol. 2016, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Falk, A.; Heine, V.M.; Harwood, A.J.; Sullivan, P.F.; Peitz, M.; Brustle, O.; Shen, S.; Sun, Y.M.; Glover, J.C.; Posthuma, D.; et al. Modeling psychiatric disorders: From genomic findings to cellular phenotypes. Mol. Psychiatry 2016, 21, 1321. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Kim, Y.H.; Quinti, L.; Tanzi, R.E.; Kim, D.Y. 3D culture models of alzheimer’s disease: A road map to a “cure-in-a-dish”. Mol. Neurodegener. 2016, 11, 75. [Google Scholar] [CrossRef] [PubMed]
- Badger, J.L.; Cordero-Llana, O.; Hartfield, E.M.; Wade-Martins, R. Parkinson’s disease in a dish—Using stem cells as a molecular tool. Neuropharmacology 2014, 76, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Compagnucci, C.; Nizzardo, M.; Corti, S.; Zanni, G.; Bertini, E. In Vitro neurogenesis: Development and functional implications of ipsc technology. Cell. Mol. Life Sci. 2014, 71, 1623–1639. [Google Scholar] [CrossRef] [PubMed]
- Brennand, K.J.; Simone, A.; Jou, J.; Gelboin-Burkhart, C.; Tran, N.; Sangar, S.; Li, Y.; Mu, Y.; Chen, G.; Yu, D.; et al. Modelling schizophrenia using human induced pluripotent stem cells. Nature 2011, 473, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Shin, S.; Jha, B.S.; Liu, Q.; Sheng, J.; Li, F.; Zhan, M.; Davis, J.; Bharti, K.; Zeng, X.; et al. Efficient and rapid derivation of primitive neural stem cells and generation of brain subtype neurons from human pluripotent stem cells. Stem Cells Transl. Med. 2013, 2, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Swistowski, A.; Peng, J.; Liu, Q.; Mali, P.; Rao, M.S.; Cheng, L.; Zeng, X. Efficient generation of functional dopaminergic neurons from human induced pluripotent stem cells under defined conditions. Stem Cells 2010, 28, 1893–1904. [Google Scholar] [CrossRef] [PubMed]
- Patani, R.; Lewis, P.A.; Trabzuni, D.; Puddifoot, C.A.; Wyllie, D.J.; Walker, R.; Smith, C.; Hardingham, G.E.; Weale, M.; Hardy, J.; et al. Investigating the utility of human embryonic stem cell-derived neurons to model ageing and neurodegenerative disease using whole-genome gene expression and splicing analysis. J. Neurochem. 2012, 122, 738–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, D.E.; Melton, D. Turning straw into gold: Directing cell fate for regenerative medicine. Nat. Rev. Genet. 2011, 12, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Pre, D.; Nestor, M.W.; Sproul, A.A.; Jacob, S.; Koppensteiner, P.; Chinchalongporn, V.; Zimmer, M.; Yamamoto, A.; Noggle, S.A.; Arancio, O. A time course analysis of the electrophysiological properties of neurons differentiated from human induced pluripotent stem cells (ipscs). PLoS ONE 2014, 9, e103418. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Do-Ha, D.; Munoz, S.S.; Ooi, L. Common pitfalls of stem cell differentiation: A guide to improving protocols for neurodegenerative disease models and research. Cell. Mol. Life Sci. 2016, 73, 3693–3709. [Google Scholar] [CrossRef] [PubMed]
- Pruszak, J.; Ludwig, W.; Blak, A.; Alavian, K.; Isacson, O. Cd15, cd24, and cd29 define a surface biomarker code for neural lineage differentiation of stem cells. Stem Cells 2009, 27, 2928–2940. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.H.; Martin, J.; Elia, J.; Flippin, J.; Paramban, R.I.; Hefferan, M.P.; Vidal, J.G.; Mu, Y.; Killian, R.L.; Israel, M.A.; et al. Cell-surface marker signatures for the isolation of neural stem cells, glia and neurons derived from human pluripotent stem cells. PLoS ONE 2011, 6, e17540. [Google Scholar] [CrossRef] [PubMed]
- Passeri, E.; Wilson, A.M.; Primerano, A.; Kondo, M.A.; Sengupta, S.; Srivastava, R.; Koga, M.; Obie, C.; Zandi, P.P.; Goes, F.S.; et al. Enhanced conversion of induced neuronal cells (in cells) from human fibroblasts: Utility in uncovering cellular deficits in mental illness-associated chromosomal abnormalities. Neurosci. Res. 2015, 101, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Ladewig, J.; Mertens, J.; Kesavan, J.; Doerr, J.; Poppe, D.; Glaue, F.; Herms, S.; Wernet, P.; Kogler, G.; Muller, F.J.; et al. Small molecules enable highly efficient neuronal conversion of human fibroblasts. Nat. Methods 2012, 9, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Pfisterer, U.; Ek, F.; Lang, S.; Soneji, S.; Olsson, R.; Parmar, M. Small molecules increase direct neural conversion of human fibroblasts. Sci. Rep. 2016, 6, 38290. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Fass, D.M.; Folz-Donahue, K.; MacDonald, M.E.; Haggarty, S.J. Highly expandable human ips cell-derived neural progenitor cells (npc) and neurons for central nervous system disease modeling and high-throughput screening. Curr. Protoc. Hum. Genet. 2017, 92, 21.28.1–21.28.21. [Google Scholar] [PubMed]
- Brix, J.; Zhou, Y.; Luo, Y. The epigenetic reprogramming roadmap in generation of ipscs from somatic cells. J. Genet. Genom. 2015, 42, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Grzybek, M.; Golonko, A.; Walczak, M.; Lisowski, P. Epigenetics of cell fate reprogramming and its implications for neurological disorders modelling. Neurobiol. Dis. 2017, 99, 84–120. [Google Scholar] [CrossRef] [PubMed]
- Robinton, D.A.; Daley, G.Q. The promise of induced pluripotent stem cells in research and therapy. Nature 2012, 481, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Suh, W. A new era of disease modeling and drug discovery using induced pluripotent stem cells. Arch. Pharmacal Res. 2017, 40, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Morey, R.; O’Neil, R.C.; He, Y.; Daughtry, B.; Schultz, M.D.; Hariharan, M.; Nery, J.R.; Castanon, R.; Sabatini, K.; et al. Abnormalities in human pluripotent cells due to reprogramming mechanisms. Nature 2014, 511, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.J.; Hutchison, W.M.; Pettersen, E. Tissue cyst rupture in mice chronically infected with Toxoplasma gondii. An immunocytochemical and ultrastructural study. Parasitol. Res. 1989, 75, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Chew, W.K.; Wah, M.J.; Ambu, S.; Segarra, I. Toxoplasma gondii: Determination of the onset of chronic infection in mice and the in vitro reactivation of brain cysts. Exp. Parasitol. 2012, 130, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Takashima, Y.; Suzuki, K.; Xuan, X.; Nishikawa, Y.; Unno, A.; Kitoh, K. Detection of the initial site of Toxoplasma gondii reactivation in brain tissue. Int. J. Parasitol. 2008, 38, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Halonen, S.K.; Chiu, F.; Weiss, L.M. Effect of cytokines on growth of Toxoplasma gondii in murine astrocytes. Infect. Immun. 1998, 66, 4989–4993. [Google Scholar] [PubMed]
- Suzuki, Y.; Claflin, J.; Wang, X.; Lengi, A.; Kikuchi, T. Microglia and macrophages as innate producers of interferon-gamma in the brain following infection with Toxoplasma gondii. Int. J. Parasitol. 2005, 35, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Orellana, M.A.; Schreiber, R.D.; Remington, J.S. Interferon-gamma: The major mediator of resistance against Toxoplasma gondii. Science 1988, 240, 516–518. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Ashour, D.; Dratz, E.; Halonen, S. Use of human induced pluripotent stem cell-derived neurons as a model for cerebral toxoplasmosis. Microbes Infect. 2016, 18, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Passeri, E.; Jones-Brando, L.; Bordon, C.; Sengupta, S.; Wilson, A.M.; Primerano, A.; Rapoport, J.L.; Ishizuka, K.; Kano, S.; Yolken, R.H.; et al. Infection and characterization of Toxoplasma gondii in human induced neurons from patients with brain disorders and healthy controls. Microbes Infect. 2016, 18, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Blader, I.J.; Manger, I.D.; Boothroyd, J.C. Microarray analysis reveals previously unknown changes in Toxoplasma gondii-infected human cells. J. Biol. Chem. 2001, 276, 24223–24231. [Google Scholar] [CrossRef] [PubMed]
- Blader, I.J.; Saeij, J.P. Communication between Toxoplasma gondii and its host: Impact on parasite growth, development, immune evasion, and virulence. APMIS Acta Pathol. Microbiol. Immunol. Scand. 2009, 117, 458–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caffaro, C.E.; Boothroyd, J.C. Evidence for host cells as the major contributor of lipids in the intravacuolar network of toxoplasma-infected cells. Eukaryot. Cell 2011, 10, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Molestina, R.E.; El-Guendy, N.; Sinai, A.P. Infection with Toxoplasma gondii results in dysregulation of the host cell cycle. Cell. Microbiol. 2008, 10, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Sinai, A.P.; Payne, T.M.; Carmen, J.C.; Hardi, L.; Watson, S.J.; Molestina, R.E. Mechanisms underlying the manipulation of host apoptotic pathways by Toxoplasma gondii. Int. J. Parasitol. 2004, 34, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Fouts, A.E.; Boothroyd, J.C. Infection with Toxoplasma gondii bradyzoites has a diminished impact on host transcript levels relative to tachyzoite infection. Infect. Immun. 2007, 75, 634–642. [Google Scholar] [CrossRef] [PubMed]
- White, M.W.; Radke, J.R.; Radke, J.B. Toxoplasma development—Turn the switch on or off? Cell. Microbiol. 2014, 16, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Radke, J.R.; Guerini, M.N.; Jerome, M.; White, M.W. A change in the premitotic period of the cell cycle is associated with bradyzoite differentiation in Toxoplasma gondii. Mol. Biochem. Parasitol. 2003, 131, 119–127. [Google Scholar] [CrossRef]
- Radke, J.B.; Lucas, O.; De Silva, E.K.; Ma, Y.; Sullivan, W.J., Jr.; Weiss, L.M.; Llinas, M.; White, M.W. Apiap2 transcription factor restricts development of the toxoplasma tissue cyst. Proc. Natl. Acad. Sci. USA 2013, 110, 6871–6876. [Google Scholar] [CrossRef] [PubMed]
- Bohne, W.; Hunter, C.A.; White, M.W.; Ferguson, D.J.; Gross, U.; Roos, D.S. Targeted disruption of the bradyzoite-specific gene bag1 does not prevent tissue cyst formation in Toxoplasma gondii. Mol. Biochem. Parasitol. 1998, 92, 291–301. [Google Scholar] [CrossRef]
- Behnke, M.S.; Radke, J.B.; Smith, A.T.; Sullivan, W.J., Jr.; White, M.W. The transcription of bradyzoite genes in Toxoplasma gondii is controlled by autonomous promoter elements. Mol. Microbiol. 2008, 68, 1502–1518. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.J.; Hutchison, W.M. The host-parasite relationship of Toxoplasma gondii in the brains of chronically infected mice. Virchows Arch. A Pathol. Anat. Histopathol. 1987, 411, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Sinai, A.P.; Watts, E.A.; Dhara, A.; Murphy, R.D.; Gentry, M.S.; Patwardhan, A. Reexamining chronic Toxoplasma gondii infection: Surprising activity for a “dormant” parasite. Curr. Clin. Microbiol. Rep. 2016, 3, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Watts, E.; Zhao, Y.; Dhara, A.; Eller, B.; Patwardhan, A.; Sinai, A.P. Novel approaches reveal that Toxoplasma gondii bradyzoites within tissue cysts are dynamic and replicating entities in vivo. MBio 2015, 6, e01155–15. [Google Scholar] [CrossRef] [PubMed]
- David, C.N.; Frias, E.S.; Szu, J.I.; Vieira, P.A.; Hubbard, J.A.; Lovelace, J.; Michael, M.; Worth, D.; McGovern, K.E.; Ethell, I.M.; et al. Glt-1-dependent disruption of cns glutamate homeostasis and neuronal function by the protozoan parasite Toxoplasma gondii. PLoS Pathog. 2016, 12, e1005643. [Google Scholar] [CrossRef] [PubMed]
- Halonen, S.K.; Taylor, G.A.; Weiss, L.M. Gamma interferon-induced inhibition of Toxoplasma gondii in astrocytes is mediated by igtp. Infect. Immun. 2001, 69, 5573–5576. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.H.; Hunter, C.A. The role of astrocytes in the immunopathogenesis of toxoplasmic encephalitis. Int. J. Parasitol. 2004, 34, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Drogemuller, K.; Helmuth, U.; Brunn, A.; Sakowicz-Burkiewicz, M.; Gutmann, D.H.; Mueller, W.; Deckert, M.; Schluter, D. Astrocyte gp130 expression is critical for the control of toxoplasma encephalitis. J. Immunol. 2008, 181, 2683–2693. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Advantage | Neuron Method | Example of Possible Application |
|---|---|---|
| Unlimited supply of neurons | hiPSC-neurons or iNs |
|
| Generation of mature functional neurons | hiPSC-neurons or iNs |
|
| Creation of disease specific neurons | hiPSC-neurons or iNs 2 |
|
| Generation of in vitro model of bradyzoite growth/cyst development | hiPSC-neurons 1 or iNs |
|
| Creation of Neuron-Astrocyte Co-cultures | hiPSC-neurons 1 or iNs |
|
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halonen, S.K. Use of Human Neurons Derived via Cellular Reprogramming Methods to Study Host-Parasite Interactions of Toxoplasma gondii in Neurons. Cells 2017, 6, 32. https://doi.org/10.3390/cells6040032
Halonen SK. Use of Human Neurons Derived via Cellular Reprogramming Methods to Study Host-Parasite Interactions of Toxoplasma gondii in Neurons. Cells. 2017; 6(4):32. https://doi.org/10.3390/cells6040032
Chicago/Turabian StyleHalonen, Sandra K. 2017. "Use of Human Neurons Derived via Cellular Reprogramming Methods to Study Host-Parasite Interactions of Toxoplasma gondii in Neurons" Cells 6, no. 4: 32. https://doi.org/10.3390/cells6040032
APA StyleHalonen, S. K. (2017). Use of Human Neurons Derived via Cellular Reprogramming Methods to Study Host-Parasite Interactions of Toxoplasma gondii in Neurons. Cells, 6(4), 32. https://doi.org/10.3390/cells6040032